ZOOTAXA
ISSN 1175-5326 (print edition)
ISSN1175-5334(online edition) Copyright © 2007 · Magnolia Press
Zootaxa 1443: 37–54 (2007) www.mapress.com/zootaxa/
Aleurocyperus humus gen. et sp. n. (Hemiptera: Aleyrodidae) from Taiwan, with
interesting feeding behaviour
CHIUN-CHENG KO1& ANIL KUMAR DUBEY
Department of Entomology, National Taiwan University, Taipei 106, Taiwan
1Corresponding author. E-mail: kocc2501@ntu.edu.tw
Abstract
Aleurocyperus humus gen. et sp. n. is described from Taiwan, feeding on Cyperus rotundus, Kyllinga brevifolia and Torulinium odoratum (Cyperaceae), also Eleusine indica (Poaceae), with illustrations of the egg, immature stages, adult male and female. The new genus is defined through a combination of tracheal furrows, marginal characters, placement of dorsal setae, and absence of first abdominal setae. Observations are also presented on feeding behaviour.
Key words: Whiteflies, new genus, new species, Cyperaceae, Poaceae, Taiwan
Introduction
The new species of whitefly described in this paper has been found breeding on cosmopolitan monocotyle-donous weeds of the families Cyperaceae and Poaceae in Taiwan. Grass-feeding whitefly species do not usu-ally feed on dicotyledonous plants, and so this species is probably restricted to feeding on these families. Detailed descriptions are provided of the egg, immatures (II, III & IV instars), and adults of both sexes. The new taxon shows similarities to Aleuromarginatus Cockerell (known to be specific to leguminous host plants), to Aleuropteridis Mound (known from ferns), and to Extensaleyrodes Bink-Moenen (known from Caesalpini-aceae). Prior to this investigation 45 genera of whitefly were on recorded from Taiwan.
The feeding behavior of this species is unusual. The immature stages are usually found feeding on the basal leaves of the host, very close to the ground. This is so, even when the upper leaves provide enough space for a colony. Puparia are sometimes hidden among sand particles that have settled over them. A few imma-tures are also found feeding on the inflorescences of these Cyperaceae, although the tissue appears to be harder than the leaf cuticle, and there is sufficient feeding space on the leaves. The immatures always orientate on the leaf surface with their anterior end upward and their posterior end downward. This behavior remains constant even when the population is high. The pattern of egg laying is also of interest, as the eggs are laid in a linear series mostly near the leaf margins. Sometimes a single egg may be laid on the ventral median area of a leaf, but eggs were not observed on the inflorescences. The orientation of the puparia on the inflorescences was the same as that on the basal parts.
Material and methods
Grass blades with nymphal whitefly were collected (Fig. 1) into A4 size polythene zip covers along with the plant inflorescence and fruits to confirm the host identity. A colony of whitefly was established in the
labora-tory to gather adults for morphological studies. Puparia were mounted using the method suggested by Martin (1987). The morphological terminology used here follows Bink-Moenen (1983), Martin (1985), and Gill (1990). Specimens for scanning electron microscopy were removed from host plants and washed in 95% eth-anol with an ultrasonic mini cleaner at 50-60 Hz for 2 minutes, then sequentially dehydrated in 95% and 100% ethanol. Specimens were critical point-dried using CO2 as the transfer fluid, then mounted on stubs and
sput-ter-coated with a gold-palladium alloy. The scanning electron microscope (JEOL S-800; Japan) was used from the Department of Entomology, National Taiwan University, Taipei, Taiwan.
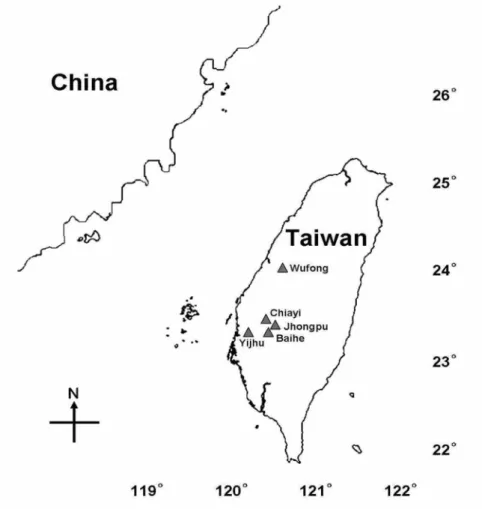
FIGURE 1. Map showing the collection localities in Taiwan.
Specimen depositories
ANIC Australian National Insect Collection, CSIRO Entomology, Canberra, ACT, Australia
BMNH Natural History Museum, London, U.K.
CDFA California Department of Food and Agriculture, Sacramento, U.S.A. IARI Indian Agricultural Research Institute, New Delhi, India
NMNH National Museum of Natural History, Tel Aviv University, Israel
NTU National Taiwan University, Taipei, Taiwan
SMTD Staatliches Museum für Tierkunde, Dresden, Germany
USNM U. S. Department of Agriculture, Beltsville, Maryland, U.S.A. (Sternorrhyncha collections of the United States National Museum of Natural History, Washington D.C.)
ZMU Zoological Museum, Universitetsparken, Department of Zoology, Copenhagen, Denmark
Aleurocyperus gen. nov.
Type species: Aleurocyperus humus sp. nov.
Diagnosis. Puparia elongate, with thin layer of colourless transparent wax on dorsum, luminous in certain
lights, deposition of wax rather prominent at thoracic and caudal tracheal pore openings and in furrows. Mar-gin irregularly crenulate. Longitudinal molting suture ill-defined but present, transverse molting suture reach-ing submedian area. Anterior and posterior marginal setae present. Paired cephalic, meso-, meta-thoracic and eighth abdominal setae present. First abdominal setae absent. Dorsum with few scattered large pores contain-ing porettes, sometimes with an associated porette. Abdominal segment sutures almost reachcontain-ing outer subdor-sum. Depressions on thoracic segments more clearly visible than on abdominal segments. Seventh abdominal segment not reduced medially. Thoracic tracheal furrows faintly indicated by margin than caudal tracheal fur-row. Thoracic tracheal folds with longitudinal ridges, caudal tracheal fold defined with stipples. Vasiform ori-fice subcordate. Operculum subcordate, filling 1/3rd of oriori-fice. Lingula tip exposed, spinose and included. Antennae short, mesad of and not extending beyond base of prolegs. Ventral abdominal setae present. All four pairs of spiracles visible. One pair of setae visible at base of rostrum.
Etymology. The genus name is derived from Aleuro+cyperus. The term ‘Aleuro–’ represents family of
the insect, ‘Aleyrodidae’ and ‘–cyperus’ is a Latin word represents sedge (Cyperaceae) from which the spe-cies was collected.
Remarks. This genus is similar to Aleuropteridis Mound (1961), Aleuromarginatus Corbett (1935) and
Extensaleyrodes Bink-Moenen (1983). It differs from Aleuropteridis in (1) dorsal disc not deflexed ventrally,
(2) absence of glands at base of marginal teeth, (3) tracheal pores unclearly modified at margin, (4) presence of mesothoracic setae, (5) defined tracheal folds, (6) position of anterior and posterior marginal setae, (7) presence of typical pores on dorsum, (8) shape of operculum, (9) dorsal pores and porettes scattered not arranged in rows, (10) exposed lingula. It differs from Aleuromarginatus in (1) absence of submarginal row of papillae, (2) absence of first abdominal setae, (3) transverse molting suture reaching submedian area, (4) defined thoracic tracheal folds, (5) dorsal disc not with 21 pairs of setae (as in Aleuromarginatus), (6) pres-ence of anterior marginal setae, (7) prespres-ence of typical pores on dorsum, (8) apically truncated operculum, (9) a pair of setae at base of rostrum, (10) ill-defined longitudinal molting suture, (11) presence of thoracic tra-cheal furrows, (12) seventh abdominal segment not reduced medially. It differs from Extensaleyrodes in (1) transverse molting suture reaching outer submedian area, (2) transverse molting suture not curved upward from middle, (3) presence of thoracic tracheal furrows, (4) shape of legs, (5) position of caudal and posterior marginal setae, (6) presence of typical dorsal pores, (7) position of cephalic setae, (8) absence of cephalic sub-dorsal, first abdominal and abdominal submedian setae, (9) antennae inside of legs, (10) anterior abdominal spiracles distinct.
From other genera of grass-feeding whiteflies, such as Agrostaleyrodes Ko (Ko et al. 2001), this new genus differs in characters of vasiform orifice, absence of raised longitudinal ridges, differentiated tracheal pores, and details of dorsal setae. It differs from Aleurocybotus Quaintance and Baker (1914) in the absence of first abdominal setae and details of dorsal setae and the antenna of male and female not extending beyond pos-terior margin of proleg. Moreover, it differs from Vasdavidius Russell (2000) in details of dorsal setae, defined tracheal pores, notched vasiform orifice and concealed lingula; and from Corbettia Dozier (1934) in having tracheal pores, absence of chain of small pores on dorsum and prominent marginal setae.
Aleurocyperus humus sp. nov.
(Figs. 5–67)
Puparium (Figs. 5, 30, 66, 67). Pale to white, with secretion of transparent colourless wax on dorsum, lumi-nous in certain lights, but white wax deposition prominent at tracheal pore openings and sometimes in
fur-rows; broadly elliptical in shape, broadest at third and fourth abdominal segments, less dimorphic, % 1.47– 1.53 mm long, & 2.00–2.02 mm long, % 0.77–0.80 mm wide, & 1.02–1.08 mm wide, found singly on the lower surface of leaves. Overlapping population of previous instars was observed. Margin (Fig. 6) smooth or irregularly crenulate, if crenulate, 8–14 crenulations in 0.1 mm. Caudal tracheal comb (Fig. 7) and thoracic tracheal combs (Figs. 8, 31) unclearly differentiated from margin. Anterior and posterior marginal setae at lat-eral margins of puparia, each 2.75 µm long.
FIGURES 2–4. Aleurocyperus humus Ko and Dubey, sp. nov., 2, egg; 3, second instar; 4, third instar.
Dorsum. Submargin not separated from dorsal disc. Submedian pockets and depressions present on all the cephalothoracic and abdominal segment sutures. Longitudinal molting suture very faint but present and reach-ing margin, transverse moltreach-ing suture reachreach-ing outer submedian area. Cephalothoracic sutures appear to be made up of broken lines (Fig. 9). Submarginal striations absent but reticulations present in the form of polyg-onal structures. Thoracic and caudal tracheal furrows defined without lateral margins but easily recognizable due to irregular striations. Dorsal pores usually in geminate pairs (Fig. 10). Median length of seventh abdomi-nal segment almost equal to sixth (Fig. 32). Eye spots absent.
Chaetotaxy. A pair of minute setae each on meso- and metathorax 1.25 µm and 2 µm long, respectively, eighth abdominal setae anterolaterad of vasiform orifice, 1.25 µm long. Cephalic setae tiny, present. First abdominal setae absent. Caudal setae 4.25 µm long, placed far from caudal opening. Interestingly, the caudal setae are at the lateral margins of the eighth abdominal segment instead of at the posterior (Fig. 5), a condition not previously reported for any whitefly species.
FIGURES 5–10. Aleurocyperus humus Ko and Dubey, sp. nov., 5, fourth instar; 6, margin; 7, caudal tracheal furrow; 8,
thoracic tracheal furrow; 9, cephalothoracic sutures; 10, dorsal pore.
Vasiform orifice (Figs. 11, 33, 34). Subcordate, % and & 7.00 µm long, % 6.75–6.87 µm wide, & 7.50–8.00 µm wide, notched at hind end, inner lateral and caudal margin with irregular ridges (Fig. 34) and tiny setae. Operculum subcordate, truncate at caudal end, % 3.75–3.87 µm long, & 4.25–4.50 µm long; % 4.12–4.50 µm wide, & 4.75–5.25 µm wide. Lingula setose, exposed and included.
Venter (Fig. 35). A pair of ventral abdominal setae 4.25 µm long, 4.75–6.50 µm apart. A pair of setae at base of meso- and metathoracic legs, 0.50 and 0.62 µm long, respectively. A pair of setae at base of rostrum, 1.25 µm long, 1.75 µm apart. Antennal base near apical end of prolegs, pads on apical end of legs faintly indi-cated. Antenna reaching midpoint of prothoracic leg, apical end keel-like (Fig. 12). No clear difference was seen in antennal length of male and female puparia. Thoracic tracheal folds with longitudinal striations (leaf surface impression) (Fig. 36) and caudal tracheal fold (Fig. 37) with fine dots, dots in caudal tracheal fold absent on submarginal area.
Adult male. Length from vertex to claspers, 1.425 mm. Upper and lower compound eyes not joined
(Figs. 13, 39). A pair of facet-like structures is seen in intervals of upper and lower compound eyes, many microtrichia in intervals. Antenna (Figs. 14, 38): Seven-segmented, elongate with many rows of concentric rings of microtrichia, segment I 2.75 µm long; segment II 5.75 µm long, three minute setae present; segment III cylindrical, longest, 16 µm long, armed with two primary sensoria and one sensorial cone, sensoria located subapically and close together; segment IV sub-cylindrical, 5 µm long, segment V sub-cylindrical, 6 µm long, a primary sensorium subapically; segment VI 5 µm long; segment VII sub-cylindrical, 4 µm long, a sensorial cone subapically, a primary sensorium and an apical seta. Wings: Forewing (Fig. 15) 127.5 µm long, 47.5 µm
wide, widest at apical 1/3rd region, not mottled, wing venation reduced to radius and cubitus, not reaching to margin, radius slightly curved downward in middle; hindwing (Fig. 16) 105 µm long, 40 µm wide, widest at apical 1/3rd region, not mottled. Legs: Mesotibia (Figs. 17A, 17B) 38.5 µm long, with rows of spines, proxi-mal tarsus 11 µm long, 8 long and 3 sproxi-mall setae present, distal tarsus 8.5 µm long; hindtibia (Figs. 18A, 18B, 40) 47.75 µm long, proximal tarsus 13.25 µm long, 11 long and 5 small setae on proximal tarsus, distal tarsus 9.25 µm long, each distal tarsus ending with 2 claws and 1 paronychium, four setae on distal tarsus of meso-and hindtibia; metatibial comb with 17 spines in a row, metatibial brush absent. Four abdominal plates (Fig. 19), each with a minute seta on outer margin, and 2 setae on inner margin, position of setae varies. Anterior and posterior plates usually reduced in size.
FIGURES 11–16. Aleurocyperus humus Ko and Dubey, sp. nov., 11, vasiform orifice; 12, proleg and antennae; 13, eye,
male; 14, antenna, male; 15, forewing, male; 16, hindwing, male.
Genitalia (Figs. 20, 41, 42). Clasper broadest at basal 1/3rd region, 4.37 µm wide at base, 13 µm long, 20– 22 pairs of setae (usually 5 pairs on outer margin, 8 pairs on centre, 6 pairs on inner margin and 1 pair subapi-cally), subapical tooth present (Fig. 43). Aedeagus 9.5 µm long, 1.5 µm wide at base, shorter than clasper, thick at base and slightly constricted in middle, swollen subapically and bluntly pointed (Fig. 44). Operculum slightly constricted at lateral and apical middle margins, apical lateral margins without setae; lingula exposed and setose (Figs. 21, 45); genital plate with three pairs of long setae, placed dorsally in oblique row.
Adult female (Figs. 62, 63). Most characters are that of male. Vertex to tip of ovipositor 1.63 mm long.
Upper and lower compound eyes not joined (Figs. 22, 51, 52), some facets pigmented, a pair of small facet-like structures are seen in intervals of upper and lower compound eyes (Fig. 52), sometimes not clear; inter-vals filled with many microthrichia. Antenna (Figs. 23, 46): Seven-segmented, many concentric rings of microtrichia, segment I 3.25 µm long; segment II 7.25 µm long with 6 setae; segment III longest, 13.75 µm
long; segment IV 3 µm long; segment V 5 µm long; segment VI 5.75 µm long; segment VII 3.87 µm long; 4 primary sensoria (Fig. 50)- 2 on segment III sub-apically (Fig. 47), these 2 sensoria are not close together as in male, and 1 each on segment V and VII sub-apically, segment VII with a apical seta (Fig. 48), 1.5 µm long. Two sensorial cones- 1 each on segment VI and VII subapically (Fig. 49), sensorial cone absent on sixth antennal segment of male but present in female, interestingly, sensorial cone absent on third antennal segment of female but present in male, perhaps, it is a useful documentation of sexual dimorphism in adults. Wings: As for males, forewing 146.25 µm long, 52.5 µm wide; hindwing 122.5 µm long, 43.75 µm wide, radius not reaching apical margin. Legs: Mesotibia (Figs. 24A, 24B) 39.5 µm long, proximal tarsus 10.75 µm long, 9 long and 3 small setae, distal tarsus 9.25 µm long, 1 seta and 5 long setae; hindtibia (Figs. 25A, 25B, 54, 55) with a row of 19–21 setae, 51.25 µm long, proximal tarsus 13.25 µm long, 12 long and 3 small setae present, distal tarsus 9.25 µm long, 1 small and 5 long setae present, each distal tarsus ending in a pair of claws and a paronychium (Fig. 53). Two abdominal plates (Fig. 26), each with 1 minute seta on outer margin and 2 setae on inner margin, position of setae varies.
FIGURES 17–20. Aleurocyperus humus Ko and Dubey, sp. nov., 17A, mesotibia dorsal view on slide, male; 17B,
mesotibia ventral view on slide, male; 18A, hindtibia dorsal view on slide, male; 18B, hindtibia ventral view on slide, male; 19, abdominal plates, male; 20, genitalia, male.
FIGURES 21–25. Aleurocyperus humus Ko and Dubey, sp. nov., 21, lingula, male; 22, compound eye, female; 23,
antenna, female; 24A, mesotibia, dorsal view on slide, female; 24B, mesotibia, ventral view on slide, female; 25A, hindtibia dorsal view on slide, female; 25B, hindtibia, ventral view on slide, female.
Genitalia (Figs. 27, 56). Two pairs of long setae on gonapophysis on outer subapical margin and 1 pair on basal region. A pair of setae on median area of unpaired gonapophysis (Fig. 57), unpaired gonapophysis diverted apically. Gonapophysis with row of transverse bands only on apical inner margin, giving serrated look to outer apical margin (Figs. 58, 59). Microtrichia abundant on gonapophysis and basal abdominal seg-ment. A pair of setae on eighth abdominal segment, lateral to basal part of female genitalia present. Opercu-lum subrectangular, concave in middle, 3.5 µm long, 5.25 µm wide at base, anterolateral margins with a seta (Figs. 28, 60, 61); lingula setose, excluded and two-segmented, basal segment longer (3 µm long) than apical segment (2.25 µm long), however, exposed basal segment is smaller than apical segment. Cement gland like a coiled ribbon and constricted at many places (Fig. 29).
Egg (Figs. 2, 64). Elliptical, surface smooth and shining in certain lights, 33.8 µm long, 12.5 µm wide.
Distal end broadly acute and basal end with a pedicle attached to the leaf surface, pedicle 7.5 µm long. Mostly on lower surface and lateral margins of leaves; laid singly, sometimes in rows.
Second instar (Fig. 3). Broadly elliptical, 0.52–0.62 mm long, 0.25–0.37 mm wide; margin smooth or
irregularly crenulate, if crenulate, 10–12 major crenulations in 0.1 mm. Anterior and posterior marginal setae 1.75 and 2.25 µm long respectively, eighth abdominal setae 0.38 µm long, caudal setae 1.75 µm long and first
abdominal setae absent. A pair of setae on submedian area of meso- and metathorax present. Thoracic and caudal tracheal furrows indicated without definite margins, distinguished by semicircular markings or striae. Longitudinal molting suture reaching margin, transverse molting suture reaching submedian region. Cephal-othoracic and abdominal segment sutures visible. Vasiform orifice subcordate, 3.25–3.5 µm long, 4.00–4.25 µm wide; operculum subrectangular, 2.00–2.75 µm long, 2.87–3.25 µm wide. Lingula exposed, setose and included. Caudal furrow visible without definite margins, 4.12–5.25 µm long. A pair of minute setae at base of prolegs, 0.5 µm long, mesolegs 0.37 µm long, metalegs 0.12 µm long and at base of rostrum 1.12 µm long. Antennae small, 1.87 µm long, above prolegs, the distance between base of proleg and antenna measured 2.75 µm. Thoracic tracheal folds without dots but caudal tracheal fold indicated with dots. Legs conical and pads not clear. All four pairs of spiracles visible.
Third instar (Figs. 4, 65). Broadly elliptical, 1.05–1.10 mm long, 0.57 mm wide. Margin smooth or
irreg-ularly crenulate, if crenulate, 19–26 crenulations in 0.1 mm. Anterior and posterior marginal setae each 2.75 µm. Eighth abdominal setae 1.25 µm, caudal setae 2.25 µm, first abdominal setae absent. A pair of setae on submedian area of meso- and metathorax, each 0.62 µm. Cephalothoracic and abdominal segment sutures and pockets visible. Longitudinal molting suture faint, transverse molting suture visible, reaching submedian area. Vasiform orifice subcordate, 5.75–6.12 µm long, 5.5–5.75 µm wide; operculum subrectangular, 3.25– 3.50 µm long, 4.00–4.5 µm wide. Lingula exposed, setose and included. A pair of setae at base of mesotho-racic legs 0.5 µm long, metathomesotho-racic legs 0.75 µm long. Pair of setae at base of rostrum 1 µm long, 0.87 µm apart. Antennae anterior to prolegs, curved and forming hook-shaped structure, distance between bases of pro-leg and antenna 1.87 µm. Paired ventral abdominal setae 2.00 µm long, 0.42 µm apart.
FIGURES 26–29. Aleurocyperus humus Ko and Dubey, sp. nov., 26, abdominal wax plates, female; 27, genitalia,
FIGURES 30–33. Aleurocyperus humus Ko and Dubey, sp. nov., 30, puparium dorsal view; 31, thoracic tracheal pore;
FIGURES 34–37. Aleurocyperus humus Ko and Dubey, sp. nov., 34, vasiform orifice; 35, puparium ventral view; 36,
thoracic tracheal fold; 37, caudal tracheal fold.
Holotype puparium. Taiwan, Tainan Hsien, Baihe, 28.ix.2003, on Eleusine indica (Poaceae), (C. C. Ko)
FIGURES 38–41. Aleurocyperus humus Ko and Dubey, sp. nov., 38, antenna, male; 39, compound eye, male; 40,
metat-ibial comb on hindtibia in dorsal view, male; 41, genitalia ventral view, male.
Paratypes. Taiwan, Tainan Hsien, Baihe, 207 puparia, 18 second instar, 23 third instar, 15 %, 22 & on 52
slides, 28.ix.2003, on Eleusine indica, C. C. Ko (one paratype slide deposited each in the collections of ANIC, BMNH, CDFA, IARI, NMNH, NTU, SMTD, ZMU, ZSI and rest in NTU collection); Chiai Hsien, Jhongpu, 10 puparia, 24.vi.2006, on Cyperus rotundus (Cyperaceae), C. C. Ko, 102 puparia on 9 slides, 15 puparia, 8 second instar, 7 third instar, 8 %, 12& on 16 slides, Taichung Hsien, Wufong, 30.viii.2005, on Cyperus
rotun-dus, C. C. Ko; one puparium on a slide, 21.ii.2004, on Cyperus rotunrotun-dus, C. C. Ko; Chiai Hsien, Jhongpu, 15
puparia, 6 second instar, 5 third instar, 8 % , 4 &, on 18 slides, 24.vi.2006, on Cyperus rotundus, C. C. Ko; Chiai Hsien, Yijhu, 12 puparia on 4 slides, 10.ix.2006, on Cypersus rotundus, C. C. Ko; Chiai Hsien, Jhongpu, 10 puparia on 2 slides, 17.ix.2006, on Cyperus rotundus, C. C. Ko; Tainan Hsien, Baihe, 10 puparia, 11 sec-ond instar, 15 third instar, 20 %, 18 & on 17 slides, 29.viii.2004, on Torulinium odoratum (Cyperaceae), Chiai Hsien, Jhongpu, 14 puparia on 5 slides, 17.ix.2006, on Kyllinga brevifolia, C. C. Ko (all NTU COLL.).
FIGURES 42–45. Aleurocyperus humus Ko and Dubey, sp. nov., 42, genitalia dorsal view, male; 43, subapical tooth on
claspers, male; 44, aedeagus apical end; 45, vasiform orifice and lingula, male.
Host plants. Cyperaceae: Cyperus rotundus, Kyllinga brevifolia, Torulinium odoratum; Poaceae:
Eleusine indica.
Distribution. Known only from Taiwan.
Etymology. The species epithet is a Latin word ‘humus’, it means ground, soil or earth, emphasizing the
unique behaviour of the species feeding at the basal region of its host.
Acknowledgements
We thank Jon Martin, BMNH for confirming the identity of the new genus and species, and for evaluation of the manuscript. The plants were identified by S. C. Wu (NTU), and H. F. Hung (NTU) assisted with the scan-ning electron microscope study. Publication is supported by part of a grant received under postdoctoral pro-gram from National Taiwan University, Taiwan (NTU 95R0044), and National Science Council (NSC95-2621-B-002-012).
FIGURES 46–49. Aleurocyperus humus Ko and Dubey, sp. nov., 46, antenna, female; 47, primary sensoria on antennal
FIGURES 50–53. Aleurocyperus humus Ko and Dubey, sp. nov., 50, primary sensoria; 51, compound eye, female; 52,
FIGURES 54–57. Aleurocyperus humus Ko and Dubey, sp. nov., 54, metatibial comb on hindtibia, dorsal view, female;
55, metatibial comb on hindtibia, lateral view, female; 56, genitalia, female; 57, genitalia, setae on unpaired gonopophy-sis, female.
FIGURES 58–61. Aleurocyperus humus Ko and Dubey, sp. nov., 58, gonapophysis apical end, dorsal view, female; 59,
gonapophysis apical end, lateral view, female; 60, vasiform orifice and lingula, female; 61, vasiform orifice and lingula, female.
FIGURES 62–67. Aleurocyperus humus Ko and Dubey, sp. nov., 62, adult female, lateral view; 63, adult female, lateral
view; 64, egg attached to leaf margin; 65, third instar; 66, puparium; 67, infestation on leaf.
References
Bink-Moenen, R.M. (1983) Revision of the African whiteflies (Aleyrodidae), mainly based on a collection from Tchad. Monografieen
Nederlandse Entomologische Vereniging, 10, 1–210.
Corbett, G.H. (1935) On new Aleurodidae (Hem.). Annals and Magazine Natural History, 10, 240–252.
Dozier, H.L. (1934) Descriptions of new genera and species of African Aleyrodidae. Annals and Magazine of Natural History, 14, 184–192.
Gill, R.J. (1990) The morphology of whiteflies. Pp. 13–46. In D Gerling [ed.] Whiteflies: Their Bionomics, Pest Status and
Manage-ment. Intercept, Andover.
Ko, C.C., Chou, L.Y. & Wu, W.J. (2001) Agrostaleyrodes arcanus, a new genus and species of whitefly (Homoptera: Aleyrodidae) from Taiwan. Zoological Studies, 40, 177–186.
Martin, J.H. (1985) The whitefly of New Guinea (Homoptera: Aleyrodidae). Bulletin of the British Museum of Natural History
(Ento-mology), 50, 303–351.
Martin, J.H. (1987) An identification guide to common whitefly pest species of the world (Homoptera: Aleyrodidae). Tropical Pest
Management, 33, 298–322.
Mound, L.A. (1961) A new genus and four new species of whitefly from ferns (Homoptera, Aleyrodidae). Revue Zoologie Botanique
Africaines, 64, 127–132.
Quaintance, A.L. & Baker, A.C. (1914) Classification of the Aleyrodidae Part II. Technical Series, U. S. Department of Agriculture
Bureau of Entomology, 27, 95–109.
Russell, L.M. (2000) Notes on the family Aleyrodidae and its subfamilies: Redescription of the genus Aleurocybotus Quaintance and Baker and description of Vasdavidius, a new genus (Homoptera: Aleyrodidae). Proceedings of the Entomological Society of