Targeting L1 Cell Adhesion Molecule Using Lentivirus-Mediated Short Hairpin RNA Interference
Reverses Aggressiveness of Oral Squamous Cell Carcinoma
Shiao-Chen Hung,
†,‡I-Hui Wu,
†,‡Shui-Sang Hsue,
§Chia-Hui Liao,
|Hsien-Chi Wang,
|,⊥Pei-Hsin Chuang,
|Shian-Ying Sung,
†,|,#and
Chia-Ling Hsieh*
,†,|,#Graduate Institute of Cancer Biology, and Center for Molecular Medicine, China Medical UniVersity & Hospital, Taichung, Taiwan, Department of Dentistry, Taipei Medical
UniVersity Hospital, Taipei, Taiwan, Department of Veterinary Medicine, National Chung-Hsing UniVersity, Taichung, Taiwan, and Department of Biotechnology,
Asia UniVersity, Wufeng, Taichung, Taiwan
Received August 25, 2010; Revised Manuscript Received October 25, 2010; Accepted October 28, 2010
Abstract: The L1 cell adhesion molecule (L1CAM) has been implicated in tumor progression of many types of cancers, but its role in oral squamous cell carcinoma (OSCC) has not been investigated. In the present study, we demonstrated overexpression of L1CAM in OSCC cells, but not in normal keratinocytes, using both clinical specimens and cell lines. This overexpression demonstrated a strong correlation with less differentiation and a higher invasion potential of cancer cells, supporting the significance of L1CAM in human OSCC tumor progression. Targeting L1CAM gene expression in SCC4 cells overexpressing L1CAM using a lentivirus-mediated small hairpin RNA (shRNA) led to a significant reduction in cell proliferation in vitro via retardation of cell cycle at the G1 phase. In addition, shRNA knockdown of L1CAM strongly attenuated the migration and invasion of SCC4 cells, and this was also observed to parallel increased E-cadherin levels and decreased levels of vimentin, fibronectin, and Snail-family transcription factors, indicating that L1CAM expression was related to the epithelial-mesenchymal transition. Furthermore, while mice receiving orthotopically placed control SCC4 cells died within 40 days due to invasive tumor growth and regional lymph node metastasis, prolonged animal survival and complete suppression of tumor progression was observed in mice implanted with L1CAM-deficent SCC4 cells, further substantiating the fundamental importance of L1CAM in OSCC pathophysiology. Our findings suggested that L1CAM is a critical mediator of tumor progression in OSCC, and targeting L1CAM using lentivirus-mediated shRNA may be a useful molecular pharmaceutical approach for the treatment of advanced OSCC.
Keywords: L1 cell adhesion molecule; oral squamous cell carcinoma; epithelial-mesenchymal transition; RNA interference; targeted gene therapy
Introduction
Oral cancer consistently ranks as one of the ten most frequently diagnosed cancers in the world.
1Approximately
300,000 new cases of oral cancer occur each year, and nearly two-thirds of patients with this cancer will die of their disease.
2,3The vast majority of malignant neoplasms in the oral cavity are squamous cell carcinomas, which account for 3-5% of all malignancies and tends to be highly invasive and spread rapidly.
4Despite multidisciplinary treatment with surgery, chemotherapy, and radiation, more than 50% of patients with oral squamous cell carcinoma (OSCC) receiving
* Corresponding author. Mailing address: Center for Molecular Medicine, China Medical University Hospital, 9F, No. 6, Hsueh-Shih Road, Taichung 40402, Taiwan. E-mail: chsieh2@
mail.cmu.edu.tw. Tel: +886-4-22052121 ext 7923. Fax: +886- 4-22333496.
†
Graduate Institute of Cancer Biology, China Medical University
& Hospital.
‡
These authors contributed equally to this article.
§
Taipei Medical University Hospital.
|
Center for Molecular Medicine, China Medical University &
Hospital.
⊥
National Chung-Hsing University.
#
Asia University.
articles
10.1021/mp1002834
XXXX American Chemical Society VOL. xxx, NO. xx, 000 MOLECULAR PHARMACEUTICS A PAGE EST: 11.2
1 2 3 4 5 6 7 8 9 10 11 12 13 14
15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36
37 38 39
40
41
42
43
44
45
46
47
therapy will eventually develop recurrent and metastatic disease, which carries a particularly poor prognosis.
5Thus, pursuing novel therapeutic alternatives to standard therapy to improve the survival of patients with advanced OSCC is medically urgent. Molecular therapy that targets molecules and proteins selectively expressed by cancer cells is thought to offer a high therapeutic index for cancers.
6Characteriza- tion of the specific molecular alterations associated with OSCC cell growth, invasion, and metastasis can advance our understanding of the molecular mechanisms underlying cancer induction and progression as well provide a basis for the development of new and effective therapeutic treatments for cancer patients.
It has become clear that adhesion molecules and adhesion processes are fundamentally involved, at various levels, in all steps of tumor progression, including detachment of tumor cells from the primary site, intravasation into the blood- stream, extravasation into distant target organs, and formation of secondary lesions.
7The aberrant expression of adhesion- related molecules, such as integrins
8-11and cadherins,
12-14has been reported in several types of human cancers, and this is associated with the invasive and metastatic potential
of cancer cells as well as a poor prognosis, making these molecules potential candidates for antineoplastic targeted therapies.
The cell adhesion molecule L1 (L1CAM) has been recently identified as a key mediator of tumor progression due to its upregulation in certain human tumors.
15Structurally, L1CAM belongs to the immunoglobulin superfamily, which is characterized by an extracellular region of multiple immu- noglobulin-like domains and fibronectin type III repeats followed by a highly conserved cytoplasmic domain.
16L1CAM was first described as a neural cell adhesion molecule based on a restricted distribution in the nervous system,
17where it is involved in the control of neurite outgrowth, adhesion, fasciculation, migration, myelination, and axon guidance.
18L1CAM promotes these cellular activities by interacting via its extracellular domain with a diverse group of cell adhesion molecules, extracellular matrix molecules, and signaling receptors.
19In addition to its cell surface localization, the extracellular domain of L1CAM can be released from the cell surface by metalloproteinases, such as plasmin and ADAM10.
20,21Non-neuronal expression of L1CAM has been observed in blood cells,
22endothelial cells,
(1) Jemal, A.; Siegel, R.; Ward, E.; Hao, Y.; Xu, J.; Thun, M. J.
Cancer statistics, 2009. CasCancer J. Clin. 2009, 59 (4), 225–
49 .
(2) Parkin, D. M.; Laara, E.; Muir, C. S. Estimates of the worldwide frequency of sixteen major cancers in 1980. Int. J. Cancer 1988, 41 (2), 184–97 .
(3) Hindle, I.; Nally, F. Oral cancer: a comparative study between 1962-67 and 1980-84 in England and Wales. Br. Dent. J. 1991, 170 (1), 15–20 .
(4) Warnakulasuriya, S. Global epidemiology of oral and oropharyn- geal cancer. Oral Oncol. 2009, 45 (4-5), 309–16 .
(5) Khuri, F. R.; Shin, D. M.; Glisson, B. S.; Lippman, S. M.; Hong, W. K. Treatment of patients with recurrent or metastatic squamous cell carcinoma of the head and neck: current status and future directions. Semin. Oncol. 2000, 27 (4 Suppl. 8), 25–33 .
(6) Khalili, J. Oral cancer: risk factors, prevention and diagnostic.
Exp. Oncol. 2008, 30 (4), 259–64 .
(7) Makrilia, N.; Kollias, A.; Manolopoulos, L.; Syrigos, K. Cell adhesion molecules: role and clinical significance in cancer.
Cancer InVest. 2009, 27 (10), 1023–37 .
(8) Liapis, H.; Flath, A.; Kitazawa, S. Integrin alpha V beta 3 expression by bone-residing breast cancer metastases. Diagn. Mol.
Pathol. 1996, 5 (2), 127–35 .
(9) Brakebusch, C.; Bouvard, D.; Stanchi, F.; Sakai, T.; Fassler, R.
Integrins in invasive growth. J. Clin. InVest. 2002, 109 (8), 999–
1006 .
(10) Patriarca, C.; Alfano, R. M.; Sonnenberg, A.; Graziani, D.;
Cassani, B.; de Melker, A.; Colombo, P.; Languino, L. R.;
Fornaro, M.; Warren, W. H.; Coggi, G.; Gould, V. E. Integrin laminin receptor profile of pulmonary squamous cell and adeno- carcinomas. Hum. Pathol. 1998, 29 (11), 1208–15 .
(11) Fornaro, M.; Tallini, G.; Bofetiado, C. J.; Bosari, S.; Languino, L. R. Down-regulation of beta 1C integrin, an inhibitor of cell proliferation, in prostate carcinoma. Am. J. Pathol. 1996, 149 (3), 765–73 .
(12) Yoshida, R.; Kimura, N.; Harada, Y.; Ohuchi, N. The loss of E-cadherin, alpha- and beta-catenin expression is associated with metastasis and poor prognosis in invasive breast cancer. Int. J.
Oncol. 2001, 18 (3), 513–20 .
(13) Tomita, K.; van Bokhoven, A.; van Leenders, G. J.; Ruijter, E. T.;
Jansen, C. F.; Bussemakers, M. J.; Schalken, J. A. Cadherin switching in human prostate cancer progression. Cancer Res. 2000, 60 (13), 3650–4 .
(14) Margineanu, E.; Cotrutz, C. E.; Cotrutz, C. Correlation between E-cadherin abnormal expressions in different types of cancer and the process of metastasis. ReV. Med.-Chir. Soc. Med. Nat. Iasi 2008, 112 (2), 432–6.
(15) Gavert, N.; Ben-Shmuel, A.; Raveh, S.; Ben-Ze’ev, A. L1-CAM in cancerous tissues. Expert Opin. Biol. Ther. 2008, 8 (11), 1749–
57 .
(16) Hortsch, M. Structural and functional evolution of the L1 family:
are four adhesion molecules better than one. Mol. Cell. Neurosci.
2000, 15 (1), 1–10 .
(17) Lindner, J.; Rathjen, F. G.; Schachner, M. L1 mono- and polyclonal antibodies modify cell migration in early postnatal mouse cerebellum. Nature 1983, 305 (5933), 427–30 .
(18) Maness, P. F.; Schachner, M. Neural recognition molecules of the immunoglobulin superfamily: signaling transducers of axon guidance and neuronal migration. Nat. Neurosci. 2007, 10 (1), 19–26 .
(19) Haspel, J.; Grumet, M. The L1CAM extracellular region: a multi- domain protein with modular and cooperative binding modes.
Front. Biosci. 2003, 8, s1210-25 .
(20) Gutwein, P.; Oleszewski, M.; Mechtersheimer, S.; Agmon-Levin, N.; Krauss, K.; Altevogt, P. Role of Src kinases in the ADAM- mediated release of L1 adhesion molecule from human tumor cells. J. Biol. Chem. 2000, 275 (20), 15490–7 .
(21) Naus, S.; Richter, M.; Wildeboer, D.; Moss, M.; Schachner, M.;
Bartsch, J. W. Ectodomain shedding of the neural recognition molecule CHL1 by the metalloprotease-disintegrin ADAM8 promotes neurite outgrowth and suppresses neuronal cell death.
J. Biol. Chem. 2004, 279 (16), 16083–90 .
(22) Ebeling, O.; Duczmal, A.; Aigner, S.; Geiger, C.; Schollhammer, S.; Kemshead, J. T.; Moller, P.; Schwartz-Albiez, R.; Altevogt, P. L1 adhesion molecule on human lymphocytes and monocytes:
expression and involvement in binding to alpha v beta 3 integrin.
Eur. J. Immunol. 1996, 26 (10), 2508–16 .
B MOLECULAR PHARMACEUTICS VOL. xxx, NO. xx 48
49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
epithelial cells, and reticular fibroblasts.
23-25Recent studies have also demonstrated aberrant expression of L1CAM in several different types of human cancers, including mela- noma, glioma, renal cancer, lung cancer, colon cancer, and ovarian cancer.
26-30For some malignancies, the level L1CAM is also a significant indicator of subsequent me- tastasis and poor prognosis,
26,31strongly suggesting a tumor- promoting function of L1CAM in advanced stages of these cancers. However, expression of L1CAM in OSCC has not been examined, and its role in oral cancer progression remains elusive.
In this study, we described for the first time the significant association of L1CAM protein expression with a more aggressive phenotype of OSCC in both clinical specimens and cell lines. L1CAM-targeting shRNA was effective at inhibiting OSCC cell growth with concomitant cell-cycle retardation at the G1 phase, which led to a nearly complete suppression of the tumorigenicity in an orthotropic OSCC animal model. Moreover, modulation of L1CAM expression in OSCC cell lines also caused changes in cell motility and gene expression that favored cancer cells undergoing epithelial-mesenchymal transition. Our results demonstrated the importance of L1CAM in OSCC tumor progression and
the therapeutic potential of L1CAM-targeting shRNAs for treatment of advanced oral cancer.
Materials and Methods
Cell Cultures. Human oral squamous cell carcinoma cell lines SAS, SCC4, SCC9, and SCC25
32were a gift from Dr.
Kuo-Wei Chang at the Institute of Oral Biology, National Yang-Ming University, Taiwan. SAS cells were grown in DMEM medium with high glucose. SCC4, SCC9, and SCC25 cells were maintained in DMEM/F12 (1:1) medium (Invitrogen, Carlsbad, CA, USA) supplemented with 1%
nonessential amino acids and 200 ng/mL hydrocortisone. All media described above were supplemented with 10% fetal bovine serum (Hyclone, Logan, UT, USA) and 1% penici- llin-streptomycin (Hyclone, Logan, UT, USA). Primary human oral keratinocytes (HOKs) were purchased from ScienCell Research Laboratories (San Diego, CA, USA) and maintained in oral keratinocyte medium (ScienCell, San Diego, CA, USA) according to the manufacturer’s instructions.
Vectors and RNA Interference. RNA interference (RNAi) vectors, pLKO.1-shL1CAM (sh-L1 #1, TRCN0000063917, target sequence GCTAACCTGAAGGTTAAAGAT; sh-L1
#2, TRCN0000063914, target sequence, GCCAATGCCTA- CATCTACGTT) and a mammalian nontargeting shRNA control pLKO.1-shGFP (sh-NT, TRCN0000072178, target sequence, CAACAGCCACAACGTCTATAT) were ob- tained from the National RNAi Core Facility (Institute of Molecular Biology, Academia Sinica, Taipei, Taiwan). These RNAi vectors were constructed by inserting annealed oli- gonucleotides containing the shRNA sequence into EcoRI and AgeI sites from the downstream of U6 promoter in pLKO.1 vector. Recombinant lentivirus carrying a short- hairpin (sh) RNA were produced by cotransfecting 293FT cells with a mixture of plasmid DNA consisting of pCMV- ψR8.91(Gag/Pol/Rev), pMD.G (VSV-G envelope), and pLKO.1-shRNA vectors using TurboFect reagent (Fermentas, Glen Burnie, MA, USA) according to the manufacturer’s instructions. After 2 days of transfection, virus-containing culture supernatant was collected and used to infect SCC4 cells in combination with 8 µg/mL Polybrene (Sigma- Aldrich, St. Louis, MO, USA). Stable cell lines were selected with 2.5 µg/mL puromycin (Calbiochem, La Jolla, CA, USA) for one week. For further establishing luciferase-expressing SCC4 cells that were pretransduced with shRNA, luciferase cDNA was removed from a pGL3-basic plasmid (Promega, Madison, WI, USA) and then cloned into the S2 bicistronic retroviral vector,
33in which luciferase was driven by a retroviral long terminal repeat promoter. Retroviral produc-
(23) Debiec, H.; Christensen, E. I.; Ronco, P. M. The cell adhesion molecule L1 is developmentally regulated in the renal epithelium and is involved in kidney branching morphogenesis. J. Cell Biol.
1998, 143 (7), 2067–79 .
(24) Nolte, C.; Moos, M.; Schachner, M. Immunolocalization of the neural cell adhesion molecule L1 in epithelia of rodents. Cell Tissue Res. 1999, 298 (2), 261–73 .
(25) Pancook, J. D.; Reisfeld, R. A.; Varki, N.; Vitiello, A.; Fox, R. I.;
Montgomery, A. M. Expression and regulation of the neural cell adhesion molecule L1 on human cells of myelomonocytic and lymphoid origin. J. Immunol. 1997, 158 (9), 4413–21 .
(26) Thies, A.; Schachner, M.; Moll, I.; Berger, J.; Schulze, H. J.;
Brunner, G.; Schumacher, U. Overexpression of the cell adhesion molecule L1 is associated with metastasis in cutaneous malignant melanoma. Eur. J. Cancer 2002, 38 (13), 1708–16 .
(27) Allory, Y.; Matsuoka, Y.; Bazille, C.; Christensen, E. I.; Ronco, P.; Debiec, H. The L1 cell adhesion molecule is induced in renal cancer cells and correlates with metastasis in clear cell carcinomas.
Clin. Cancer Res. 2005, 11 (3), 1190–7 .
(28) Boo, Y. J.; Park, J. M.; Kim, J.; Chae, Y. S.; Min, B. W.; Um, J. W.; Moon, H. Y. L1 Expression as a Marker for Poor Prognosis, Tumor Progression, and Short Survival in Patients with Colorectal Cancer. Ann. Surg. Oncol. 2007, 14 (5), 1703–11 .
(29) Kettunen, E.; Nicholson, A. G.; Nagy, B.; Wikman, H.; Seppanen, J. K.; Stjernvall, T.; Ollikainen, T.; Kinnula, V.; Nordling, S.;
Hollmen, J.; Anttila, S.; Knuutila, S. L1CAM, INP10, P-cadherin, tPA and ITGB4 over-expression in malignant pleural mesothe- liomas revealed by combined use of cDNA and tissue microarray.
Carcinogenesis 2005, 26 (1), 17–25 .
(30) Daponte, A.; Kostopoulou, E.; Kollia, P.; Papamichali, R.;
Vanakara, P.; Hadjichristodoulou, C.; Nakou, M.; Samara, S.;
Koukoulis, G.; Messinis, I. E. L1 (CAM) (CD171) in ovarian serous neoplasms. Eur. J. Gynaecol. Oncol. 2008, 29 (1), 26–30 .
(31) Fogel, M.; Gutwein, P.; Mechtersheimer, S.; Riedle, S.; Stoeck, A.; Smirnov, A.; Edler, L.; Ben-Arie, A.; Huszar, M.; Altevogt, P. L1 expression as a predictor of progression and survival in patients with uterine and ovarian carcinomas. Lancet 2003, 362 (9387), 869–75 .
(32) Tan, K. D.; Zhu, Y.; Tan, H. K.; Rajasegaran, V.; Aggarwal, A.;
Wu, J.; Wu, H. Y.; Hwang, J.; Lim, D. T.; Soo, K. C.; Tan, P.
Amplification and overexpression of PPFIA1, a putative 11q13 invasion suppressor gene, in head and neck squamous cell carcinoma. Genes, Chromosomes Cancer 2008, 47 (4), 353–62 .
(33) Hsieh, C. L.; Chen, B. F.; Wang, C. C.; Liu, H. H.; Chen, D. S.;
Hwang, L. H. Improved gene expression by a modified bicistronic retroviral vector. Biochem. Biophys. Res. Commun. 1995, 214 (3), 910–7 .
VOL. xxx, NO. xx MOLECULAR PHARMACEUTICS C 92
93 94 95 96 97 98 99 100 101 102 103 104 105 106 107 108 109 110 111 112 113 114
115 116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
tion and infection were performed as described previously.
34The resultant stable cell lines were selected with 0.8 mg/
mL G418 (Calbiochem, La Jolla, CA, USA) for 10 days.
Western Blot Analysis. Protein extracts from cell lines were analyzed on SDS-polyacrylamide gels (15 µg per lane) and transferred to Hybond ECL nitrocellulose membranes (GE Healthcare Life Science, Piscataway, NJ, USA). Blots were probed with 1:1000 diluted monoclonal mouse anti- human L1CAM (NeoMarker, Fremont, CA, USA), anti-E- cadherin (Cell Signaling Technology, Danvers, MA, USA), antivimentin (NeoMarker, Fremont, CA, USA) and antifi- bronectin (BD Biosciences, Bedford, MA, USA) according to the manufacturers’ instructions. For loading control, blots were probed with an anti-EF1-R monoclonal antibody (1:
10,000; R&D Systems, Minneapolis, MN, USA). After incubation with an HRP-conjugated secondary antibody (1:
5000; GE Healthcare Life Sciences, Piscataway, NJ, USA), chemiluminescent signals were detected using an ECL Plus kit and exposed to Hyperfilm ECL (GE Healthcare Life Science, Piscataway, NJ, USA). Protein band quantification was carried out using ImageJ software.
Cell Proliferation Assay. Cells were seeded at a density of 5 × 10
3cells/well in a 96-well plate. Viable cells were measured using WST-1 reagent (Roche Applied Science, Mannhein, Germany) at each time point, followed by an additional incubation for 2 h at 37 °C. Absorbance was recorded at 450 nm using a Synergy 2 Multi-Mode micro- plate reader (BioTek, Winooski, VT, USA). All experiments were performed in triplicate.
Boyden Chamber Invasion Assays. The invasion of cancer cells was assessed using 24-well Transwell plates.
Briefly, 2 × 10
5cells in 0.5% FBS-containing media were added to the upper chamber containing 8 µm pore polycar- bonate coated with 1 mg/mL of Matrigel; the lower chamber was filled with growth medium. After a 16 h incubation, the upper surface of the membrane was scrubbed with a cotton- tipped swab. The invading cells on the lower surface of the membrane were fixed and stained with 0.5% crystal violet.
Random fields (5/membrane) were photographed at 40 × magnification and then quantified by measuring absorbance of dye extracts at 570 nm.
Wound Healing Assay. shRNA-expressing SCC4 cells resuspended in medium were seeded into 24-well plates.
When cells reached 90% confluence, a single wound was created in the center of the cell monolayer by gentle removal of the attached cells with a sterile plastic pipet tip. The debris was removed by washing with serum free medium. Cells having migrated into the wounded area or those protruding from the border of the wound were visualized and photo- graphed under a Zeiss Axioplan microscope (Carl Zeiss MicroImaging, Thornwood, NY, USA) with a 10 × objective at three preselected time points (0, 4, and 8 h). Each experiment was independently performed at least three times.
Immunohistochemistry. Archived paraffin-embedded oral cancer tissue specimens were collected with the approval of the Institutional Review Board. Written informed consent was obtained from all participants. Mouse tissues, including tongue and cervical lymph nodes, were collected at the end of the animal experiments (6 weeks after tumor implantation).
Immunohistochemical analyses were performed as described previously
35with the following modifications: Paraffin em- bedded sections were deparaffinized and rehydrated using xylene and decreasing concentrations of ethanol. Antigen retrieval was performed with a steamer by heating the slides in 10 mM sodium citrate (pH 6.0) for 10 min. Slides were incubated with mouse anti-human L1CAM monoclonal (1:
40; Lab Vision, Fremont, CA, USA) and goat anti-firefly luciferase (1:10,000; Abcam, Cambridge, U.K.) antibodies at room temperature for 1 h, followed by incubation with HRP-polymer conjugate (Upstate, Lake Placid, NY, USA) under the same conditions. The chromogenic reaction was performed with diaminobenzidine (Sigma, St. Louis, MO, USA) for 5 min, and slides were counterstained with hematoxylin.
Immunofluorescence Staining. shRNA-expressing SCC4 cells grown on Nunc chamber slides (Thermo Fisher Scientific, Rockford, IL, USA) were fixed with 4% paraform- aldehyde in PBS at room temperature for 15 min. The slides were blocked with 2% bovine serum albumin and then incubated with rabbit anti-human E-cadherin antibodies (Cell Signaling Technology, Beverly, MA, USA) and rabbit anti- human vimentin antibodies (Lab Vision, Fremont, CA, USA) in a 1:100 dilution for overnight at 4 °C. The slides were washed and then incubated with Alexa Fluor 488 (for E-cadherin detection) and Alexa Fluor 546 (for vimentin detection) goat anti-rabbit IgG (Invitrogen, Carlsbad, CA, USA) in a 1:200 dilution for 1 h at room temperature. Slides were washed and mounted for confocal microscopy.
RNA Extraction and Real-Time Quantitative PCR.
Total RNA was extracted from cell lines and first-strand cDNA was generated from 1 µg of total RNA and random primers using Moloney Murine Leukemia Virus Reverse Transcriptase (Invitrogen, Carlsbad, CA, USA) in a total volume of 20 µL. Quantitative RT-PCR was performed using a LightCycler 480 TaqMan Master kit with Universal ProbeLibrary probes (Roche Applied Science, Mannheim, Germany). The primers and probes were designed using a web-based Assay Design Center at www.universalprobeli- brary.com, and sequences are shown in the Supporting Information (Table S1). The real-time PCR reaction was conducted according to the manufacturer’s instructions, consisting of a denaturation step (10 min) and 55 cycles of amplification (95 °C for 10 s, 60 °C for 10 s, followed by single fluorescence acquisition at 72 °C for 10 s). The relative gene expression of a specific target in each group was represented as 2
-∆CT; ∆CT was determined by subtracting
(34) Hsieh, C. L.; Pang, V. F.; Chen, D. S.; Hwang, L. H. Regression of established mouse leukemia by GM-CSF-transduced tumor vaccine: implications for cytotoxic T lymphocyte responses and tumor burdens. Hum. Gene Ther. 1997, 8 (16), 1843–54 .
(35) Hsieh, C. L.; Xie, Z.; Yu, J.; Martin, W. D.; Datta, M. W.; Wu, G. J.; Chung, L. W. Non-invasive bioluminescent detection of prostate cancer growth and metastasis in a bigenic transgenic mouse model. Prostate 2007, 67 (7), 685–91 .
D MOLECULAR PHARMACEUTICS VOL. xxx, NO. xx 162
163 164 165 166 167 168 169 170 171 172 173 174 175 176 177 178 179 180 181 182 183 184 185 186 187 188 189 190 191 192 193 194 195 196 197 198 199 200 201 202 203 204 205 206 207 208 209 210 211 212 213 214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
233
234
235
236
237
238
239
240
241
242
243
244
245
246
247
248
249
250
251
252
253
254
255
256
257
258
259
260
261
262
263
264
265
266
267
the average housekeeping gene (HSPCB) CT value from the average target gene value.
Animals. All animal work was performed in accordance with a protocol approved by the Institutional Animal Care and Use Committee. Six-week-old male athymic nude (nu/
nu) mice from BioLASCO Taiwan Co. (Taipei, Taiwan) were used for the experiments. To establish the orthotropic oral cancer model, 2 × 10
5luciferase-expressing cancer cells were resuspended in 20 µL of PBS and directly injected submu- cosally into the anterior tongue of the mice, according to the published protocol.
36Tumor growth and regional me- tastasis was noninvasively monitored using an IVIS biolu- minescence imaging system (Caliper Life Sciences, Hop- kinton, MA, USA) as described previously.
37Statistical Analysis. For the in Vitro studies, all data are presented as the means ( SD. Differences between groups were analyzed using the two-tailed, unpaired Student’s t test.
A p value of less than 0.05 was considered to be significant.
In the mouse studies, Mann-Whitney rank-sum test was used for analysis.
Results
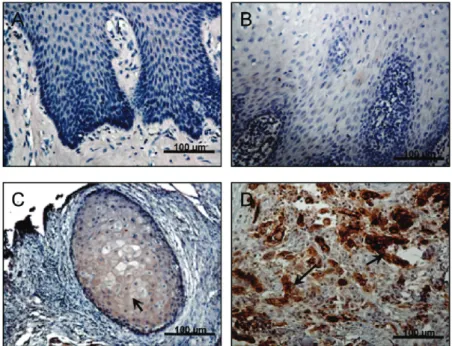
L1CAM Is Overexpressed in Cancerous Lesions of Patients with OSCC. L1CAM expression has been detected in several types of cancers but has yet to be investigated in OSCC. To investigate whether L1CAM was involved in the progression of human OSCC, we first examined the expres- sion levels of L1CAM in 25 oral tissues from patients with OSCC by immunohistochemical (IHC) staining (Figure 1).
L1CAM expression was not observed in healthy oral epithelium (Figure 1A). Oral premalignant lesions, such as hyperparakeratosis and squamous hyperplasias (Figure 1B), had no or faint L1CAM immunoreactivity in almost all epithelium cells in the hyperplasia of stratum spinosum and were considered L1CAM-negative. In contrast, positive L1CAM immunoreactivity was seen in cancerous lesions of OSCC specimens. Compared to the low-grade (well dif- ferentiated) tumors (Figure 1C) that expressed L1CAM mainly in the cytoplasm and at the cell surface of the cells, significantly increased L1CAM expression was detected in late-stage (moderated to poorly differentiated) tumors (Figure 1D). The positive correlation between L1CAM expression levels and the stage of tumor differentiation indicated the significance of L1CAM expression in human OSCC tumor biology.
L1CAM Protein Expression Is Associated with Aggres- siveness in OSCC Cell Lines. L1CAM protein levels were also examined in four independent OSCC cell lines: SAS, SCC4, SCC9, and SCC25 as well as normal oral kerati- nocytes (HOKs). Intact and cleaved L1CAM protein was
(36) Myers, J. N.; Holsinger, F. C.; Jasser, S. A.; Bekele, B. N.; Fidler, I. J. An orthotopic nude mouse model of oral tongue squamous cell carcinoma. Clin. Cancer Res. 2002, 8 (1), 293–8 .
(37) Bisanz, K.; Yu, J.; Edlund, M.; Spohn, B.; Hung, M. C.; Chung, L. W.; Hsieh, C. L. Targeting ECM-integrin interaction with liposome-encapsulated small interfering RNAs inhibits the growth of human prostate cancer in a bone xenograft imaging model.
Mol. Ther. 2005, 12 (4), 634–43 .
Figure 1. Detection of L1CAM protein in OSCC patient tissues. Representative images (200 ×; scale bar )100 µm) of immunohistochemical staining of L1CAM in OSCC tissues. (A) Normal oral epithelium exhibits negative L1CAM protein expression. (B) In squamous hyperplasic lesions, undetectable to very weak L1CAM immunoreactivity was present. (C) A well differentiated OSCC tumor stained positive with anti-L1CAM in the cytoplasm and at the cell surface of cancer cells, and (D) strong positive immunoreactivity for L1CAM was detected in poorly differentiated OSCC tumors. Arrows denote positive staining of L1CAM (C and D).
VOL. xxx, NO. xx MOLECULAR PHARMACEUTICS E 268
269 270 271 272 273 274 275 276 277 278 279 280 281 282 283 284 285 286 287
288 289 290 291 292 293 294 295 296 297 298 299 300 301 302 303 304 305 306 307 308 309 310 311 312 313 314 315 316
F1
detected in cell lysates of all of the tested cancer cell lines.
Based on Western blot analysis, a higher expression was observed in the SCC4 and SAS cell lines, and a lower expression was observed in the SCC9 and SCC25 cell lines;
L1CAM expression was undetectable in HOKs (Figure 2A).
We next examine whether the L1CAM expression pattern in the OSCC cell lines was related to their metastatic potentials using Matrigel invasion chamber assays. The strongest invasion was noted in SCC4 cells, which expressed the highest levels of L1CAM, followed by SAS, SCC9, and then SCC25 (Figure 2B). The results obtained from these cell lines are in accordance with our clinical observations (Figure 1) indicating that L1CAM protein levels are associ- ated with tumor malignancy.
Targeting L1CAM by Lentivirus-Mediated shRNA In- hibits OSCC Cell Growth and Cell Cycle Progression in Vitro. To assess the functional significance of L1CAM expression during OSCC tumor progression, RNAi was used to target L1CAM gene expression. We stably expressed lentiviral vectors expressing a short hairpin (shRNA) se- quence that targeted L1CAM (sh-L1) or a mammalian nontargeting sequence (sh-NT) in SCC4 cells, which had demonstrated high levels of L1CAM expression and invasive behavior. The two distinct shRNAs targeting L1CAM caused a dramatic reduction in L1CAM protein levels (96% and 85%, respectively) when compared with parental SCC4 cells.
The sh-NT shRNA demonstrated no effects on L1CAM expression (Figure 3A). These results indicated the effective- ness of lentivirus-mediated shRNA in silencing L1CAM gene expression in OSCC cells.
We next determined whether the altered L1CAM expres- sion in SCC4 cells could affect cell growth in Vitro. A colorimetric WST-1 assay was used to quantify cell prolif- eration. We found that silencing L1CAM in SCC4 cells (SCC4-sh-L1) led to a 50% decrease in the number of cells after 48 and 72 h of culture (p < 0.05 and p < 0.0001, respectively) when compared with the control SCC4 cells (SCC4-sh-NT) (Figure 3B). Noticeably, SCC4-sh-L1 and SCC4-sh-NT cells showed similar cell viability during the culture period, as measured by trypan blue dye exclusion assay (data not shown), suggesting that death-inducing pathways are not involved in the growth inhibition resulting from L1CAM silencing.
To understand the control mechanisms by which L1CAM regulated cell growth, we determined and compared the cell- cycle distribution of SCC4-sh-L1 and SCC4-sh-NT cells by measuring DNA content following treatment with nocoda- zole, a microtubule inhibitor, to arrest the cells during mitosis.
As shown in Figure 3C, while the majority of the SCC4- sh-NT cells accumulated in a G2/M peak (70.63%) by 8 h after nocodazole treatment and completed one cell cycle over the next 4 h, SCC4-sh-L1 cells still retained 21.11% and 17.56% of their cell population in the G1 and S phases, respectively. Together, these results demonstrated that sup- pression of L1CAM expression by shRNA caused a G1 cell- cycle arrest and cell proliferation inhibition in SCC4 cells.
Figure 2. Correlation between L1CAM expression and the aggressiveness of OSCC cell lines. (A) Representative Western blot results indicating L1CAM expression in SCC25, SCC9, SCC4, and SAS human OSCC cell lines and normal human oral keratinocytes (HOK). EF1-R protein levels are shown to have varied loading quantities.
(B) Characterization of the invasive ability of OSCC cell lines using a Matrigel chamber assay. Migrated cells through the Matrigel were stained with crystal violet 16 h after cell plating. The lower surface of the membrane was photographed (100 ×) and shown as a representative image from each cell line (upper panel). The number of invaded cells was quantified by measuring the absorbance of dye extract at 570 nm (lower panel). Results from one of three experiments are shown. Error bars indicate SD of triplicate measurements.
F MOLECULAR PHARMACEUTICS VOL. xxx, NO. xx 317
318 319 320 321 322 323 324 325 326 327 328 329 330 331 332 333 334 335 336 337 338 339 340 341 342 343 344 345 346 347 348 349 350 351 352 353 354 355 356 357 358 359 360 361 362 363 364 365 366 367 368 369 370 371 372
F2F3
L1CAM shRNA Inhibits Migration and Invasion of OSCC Cells. To elucidate the role of L1CAM in the process of OSCC metastasis, we determined the migration and invasive potential of SCC4-sh-L1 cells in Vitro. We used a wound-healing assay to assess the role of L1CAM in cancer cell migration. As shown in Figure 4A, the movement of SCC4-sh-L1 cells toward the denuded areas was much slower
than SCC-sh-NT cells. Quantitative data showed that while the distance between the wound edges of SCC4-sh-L1 cells was reduced by 12.76% and 30% at 4 and 8 h, respectively, the gap width of the SCC4-sh-NT cells was closed by 43%
at 4 h and completely filled at 8 h. In Matrigel chamber assays (Figure 4B), SCC4-sh-L1 cells exhibited a marked decrease (p < 0.001) in the number of cells invading through Figure 3. Effects of L1CAM gene silencing by shRNA on SCC4 cell growth and cell-cycle progression. (A) Western blot analysis of L1CAM expression in parental SCC4 and SCC4 cells infected with a lentiviral vector carrying a nontargeting shRNA (sh-NT) or shRNA targeting L1CAM (sh-L1). EF1-R protein levels are shown to vary in loading quantities. (B) Cell proliferation of shRNA-expressing SCC4 sublines was determined by WST-1 assay performed daily for 3 days. The relative cell number was assessed by absorbance at 450 nm and presented as the fold change relative to the day of plating (0 h). Error bars indicate SD of triplicate measurements. *p < 0.05, **p < 0.0001 versus SCC4-sh-NT. (C) Representative FACS profiles of SCC4-sh-L1 and SCC4-sh-NT cells stained with propidium iodide for DNA content at the indicated time points after adding 100 ng/mL nocodazole to accumulate cells at the G2/M transition. Data was processed using the ModFit LT program, and the percentage of cells at each phase of cell cycle is indicated.
VOL. xxx, NO. xx MOLECULAR PHARMACEUTICS G 373
374 375 376 377 378 379
380
381
382
383
384
385
386
F4the membrane after a 16 h incubation, demonstrating an approximate 65% decrease relative to that of the SCC4-sh- NT cells. These results suggested that L1CAM is important for the metastatic capacity and, thus, for the malignancy of OSCC cells.
L1CAM Gene Silencing Causes Changes in the Expression of Epithelial and Mesenchymal Markers.
Epithelial-mesenchymal transition (EMT) is a critical biological process in epithelial tumor invasion, progression, and metastasis.
38E-cadherin is a transmembrane protein that mediates the major cell-cell adhesions in epithelial cells, a function that has been linked to its role as a tumor suppressor.
39Vimentin is an intermediate filament that participates in adhesion, migration, survival, and cell signal- ing processes normally occurring in mesenchymal cells, and it is also involved in pathological or physiological processes
that require epithelial cell migration.
40A defining feature of EMT is the loss of E-cadherin expression and gain of vimentin expression.
41We next investigated if the influence L1CAM gene silencing on the migration and invasion of OSCC cells was related to EMT by determining the expres- sion level of these EMT markers. As shown in Figure 5, L1CAM silencing markedly increased the protein levels of the epithelial marker E-cadherin and reduced the levels of the mesenchymal marker vimentin in SCC4 cells, as deter- mined by immunocytochemical analyses (Figure 5A) and Western blotting (Figure 5B). Consistent with this protein profile, quantitative RT-PCR analysis (Figure 5C) showed a 3-fold increase in E-cadherin and a massive reduction ( ∼97%) in vimentin in SCC4-sh-L1 cells when compared with SCC4-sh-NT cells. Moreover, fibronectin, another mesenchymal marker, and known EMT-regulatory transcrip- tion factors, Slug and Snail, were also downregulated in SCC4-sh-L1 cells, further confirming that the altered gene
(38) Thiery, J. P. Epithelial-mesenchymal transitions in tumour progression. Nat. ReV. Cancer 2002, 2 (6), 442–54 .
(39) Beavon, I. R. The E-cadherin-catenin complex in tumour me- tastasis: structure, function and regulation. Eur. J. Cancer 2000, 36 (13 Spec. No.), 1607–20 .
(40) Steinert, P. M.; Roop, D. R. Molecular and cellular biology of intermediate filaments. Annu. ReV. Biochem. 1988, 57, 593–625 .
(41) Shook, D.; Keller, R. Mechanisms, mechanics and function of epithelial-mesenchymal transitions in early development. Mech.
DeV. 2003, 120 (11), 1351–83 .
Figure 4. Effects of L1CAM gene silencing by shRNA on migration and invasion of SCC4 cells. (A) Wound healing assays using confluent cells gently wounded through the central axis. The wounded areas were examined under
×100 magnification, and images were taken at 0, 4, and 8 h. The dashed lines indicate the leading edge of the cut that was made across the monolayer (left). Cell migration was quantified by measuring the distance between the invading front of cells and plotted as the percentage relative to the zero time point (right). Error bars indicate SD of 10 random measurements. (B) Invasion of SCC4 cells was determined by Matrigel chamber assay at 16 h after incubation. Images are representative of the migration of SCC4-sh-L1 cells from three separate experiments performed in triplicate (left). The number of invaded cells was quantified by measuring the absorbance of dye extract of crystal violet at 570 nm. Results from one of three experiments are shown (right). Error bars indicate SD of triplicate measurements. *p < 0.05, **p < 0.0001 versus SCC4-sh-NT (A and B).
H MOLECULAR PHARMACEUTICS VOL. xxx, NO. xx 387
388 389 390 391
392 393 394 395 396 397 398 399 400 401 402
403 404 405 406 407 408 409 410 411 412 413 414 415 416 417 418 419 420
F5
expression caused by L1CAM knockdown was EMT-related.
Taken together, these data suggested that targeting L1CAM expression in invasive OSCC cells reverses the EMT phenotype and, thus, attenuates the aggressiveness of these cancer cells.
Targeting L1CAM Expression by shRNA Suppresses Tumor Growth and Lymph Node Metastasis of OSCC Cells in an Orthotopic Tumor Model. To investigate of the role of L1CAM in OSCC tumorigenesis and metastasis in ViVo, we used an orthotopic xenograft model of squamous cell carcinoma of the oral tongue. The model was generated by submucosal injection of human OSCC cells into the anterior tongue of nude mice, which has been reported to develop a high incidence of cervical lymph node me- tastases.
36To monitor, in real time, the primary tumor growth
and distant metastasis noninvasively, SCC4-sh-L1 and SCC4- sh-NT cells were stably transduced with a luciferase reporter gene by retroviral infection.
37We confirmed the equal bioactivity of the luciferase reporter between SCC4-sh-L1 and SCC4-sh-NT cells using an in Vitro luciferase enzymatic assay (data not shown) prior to injection of cells into the mice. Mice that were orthotopically transplanted with lu- ciferase-transduced SCC4-sh-L1 and SCC4-sh-NT cells were visualized by bioluminescence imaging at one-week intervals.
One week after cell implantation, we observed biolumines- cence at the site of the injection in all mice, regardless of the cell line used, demonstrating the success of the orthotopic transplantation (Figure 6A, week 1). While the control SCC4- sh-NT group (n ) 7) showed a substantial increase of photon intensity in the primary site (mouth) and spreading into the Figure 5. Effects of L1CAM gene silencing by shRNA on expression of epithelial-mesenchymal transition (EMT) markers. (A) Representative bright-field images of SCC4-sh-L1 and SCC4-sh-NT cells grown on a plastic surface (200 ×; scale bar ) 100 µm) and immunofluorescence staining for E-cadherin (green) and vimentin (red) expression in cells grown on cover glass under confocal microscopy ( ×400; scale bar )20 µm). Nuclei were counterstained with DAPI (blue). (B) Western blotting analysis of EMT markers. EF1-R is shown as the protein loading control. (C) Real-time RTPCR analysis of the above cells for epithelial (black bar) and mesenchymal markers (white bar). mRNA levels in SCC4-sh-L1 cells are displayed as the fold change relative to SCC4-sh-NT cells. Data are representative of three independent experiments and shown as the mean ( SD. *p < 0.05, **p < 0.0001 versus SCC4-sh-NT.
VOL. xxx, NO. xx MOLECULAR PHARMACEUTICS I 421
422 423 424 425 426 427 428 429 430 431 432 433 434 435
436 437 438 439 440 441 442 443 444 445 446 447 448 449 450
F6