1
行政院國家科學委員會專題研究計畫成果報告
蛻膜內巨噬細胞在懷孕初期所扮演的角色(1/3)
The r ole of decidual macr ophages in ear ly pr egnancy
計畫編號:NSC 90-2314-B-002-453
執行期限:90 年 8 月 1 日至 91 年 7 月 31 日
主持人:何弘能 臺大醫院婦產部
共同主持人:無
計畫參與人員:陳筱菲 台大醫學院免疫所博士生
一、中文摘要 哺乳動物的胎兒向來被視為一種成功 的半異體移植物,而胎兒能否在母體子宮 內正常生長發育,則取決於母體免疫系統 之趨向,即是當子宮內微環境以第二型輔 助型 T 細胞反應為主時,胚胎才能正常發 生;反之,若是以第一型細胞免疫反應為 主時,則會干擾胚胎發育。由先前已發表 文獻中得知,出現在人類蛻膜中的白血球 以巨噬細胞、子宮自然殺手細胞及 T 細胞 為主;此外,無論是在人類或是小鼠中, 巨噬細胞皆佔了 20~30% 的高比例,大多 數學者推測巨噬細胞在蛻膜中扮演了極重 要且獨佔的調控角色,例如可能藉由分泌 細胞激素或細胞驅動激素等可溶性因子來 執行其調控機制,但是仍僅止於推測階 段,這群細胞確實的重要性則有待進一步 的實驗來證實。本研究中觀得蛻膜巨噬細 胞表面表現高量的 CD30L 與 HLA-DR 分子, 表示蛻膜巨噬細胞呈現被活化狀態,但是 卻僅只表現極微量的 CD80 與 CD86 分子。 根據這些發現,我們推測蛻膜巨噬細胞可 能是借由 CD30L 與蛻膜基質表現的 CD30 交 互作用,進而調控細胞表面 CD80 與 CD86 等活化輔助因子的表現,來抑制母體 T 細 胞的正常免疫反應,使得胚胎得以不被母 體排斥而正長發育。 關鍵詞:蛻膜中的巨噬細胞、免疫調控、 細胞間交互作、CD30、CD30L Abstr actThe mammalian fetus has been considered
as a successful semiallograft after implan-tation and maternal immunomodulation can influence fetal survival. Previous studies showed that macrophages are one of the most abundant leukocytes (around 30% of the decidual leukocytes) in decidua and persist in large numbers throughout gestation. They are found primarily cluster around the implan-tation sites. Thus, they may play some regulatory roles in early pregnancy. It has been hypothesized that fetal and maternal macrophages are pivotal regulatory cells in the membrane environment. Many functions have ascribed to these cells; for example, decidual macrophages may utilize their mediator products to provide the TH2-like immune state in feto-matenal interface in early pregnancy. However, the accurate role of decidual macrophages remains unclear and needs to be further defined. In the current study, we found there are high levels of CD30L and HLA-DR expression on the surface of decidual macrophages, but low expression of CD80 and CD86 co-stimulatory molecules. Based on the observation, we propose that decidual macro-phages are modulated in CD80 and CD86 upon CD30- CD30L interaction.
Keywords: decidual macrophages,
immuno-regulation, cellular interaction, CD30, CD30L,
二、緣由與目的
2
as a successful semiallograft after implan-tation and maternal immunomodulation can influence fetal survival. Previous studies showed that not only in human, but also in murine model, macrophages are one of the most abundant cell populations in decidua and persist in large numbers throughout gestation. They are found primarily cluster around the implantation sites. Thus, they may play some regulatory roles in early pregnancy. Although macrophages are one of the major cellular populations in the early pregnant uteri (around 30% of the decidual leukocytes), there are relative few studies discussing about them because of the extreme difficulties in their isolation and purification from the uteri.
It has been hypothesized that fetal and maternal macrophages are pivotal regulatory cells in the membrane environment. Many functions have ascribed to these cells, ranging from the classical immunological roles of antigen presentation, bacteriocidal activities, immunosupperssion and the production of cytokines to phagocytosis. As we know TH2-biased maternal microenvironment is required for successful pregnancy, decidual macrophages may utilize their soluble medi- ators to achieve the local TH2-skewed immune state in feto-maternal interface in early pregnancy. However, the accurate role of decidual macrophages remains unclear and needs to be further defined.
CD30 expression is highly associated with tumor cells, but is has been also found in placenta with normal pregnancy. It implicated that there should be corresponding CD30 ligand (CD30L) adjacent to CD30. Thus it’s sensible to suggest that some signal is delivered upon CD30 ligation. There is evidence revealed that reverse signaling appears in CD30-CD30L interaction. Whether there is any CD30L expression and the consequence of the interaction happened in decidua require further investigation.
In addition, decidual macrophages derived soluble factors may act on other cells in uteri. The interaction between decidual
macro-phages and neighboring cells is thus also important.
The purposes of this study are to propose the satisfactory hypothesis for the regulatory roles of decidual macrophages in early pregnancy and make efforts to find out a reasonable immunotherapy for the patients with infertility and recurrent spontaneous abortion.
三、結果與討論
After enzyme digestion and 50-µm steel mesh filtration, single cells derived from early pregnancy were determined for surface marker expression by flow cytometer. Live cells were gated and analyzed the level of CD11b and HLA-DR at the same time. As shown in Fig. 1, all CD11b+ cells were HLA-DR+.
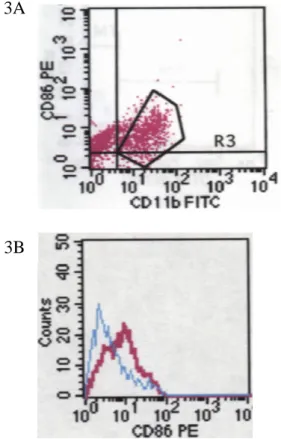
It has been implicated that decidual macrophages were activated in situ. Since CD80 and CD86 are also upregulated while macrophages become activated, we further trace the expression of these two co-stimulatory molecules. CD11b+ cells were gated (Fig. 2A, 3A) and analyzed the CD86 or CD86 expression. CD11b+HLA-DR+ were CD80- CD86low. (Fig. 2B, 3B) This is in coincidence with the previous study that indicated placental macrophages have a decreased ability to present antigen in mouse model. (PNAS. 90:462, 1993)
Fig.1 All CD11b+ cell were HLA-DR positive. One million single cells were used for flow cytometry. We gated live cells and analyzed surface CD11b (FL1)
3
It has been demonstrated that there was high level of the CD30 in human decidual cells. (Placenta. 22:309,2001; Am. J. Pathol. 145: 276, 1994) This implicated that there should be corresponding CD30 ligand (CD30L, CD153) adjacent to CD30. In addition, unpublished data supplied by co-laboratory indicated that CD86 expression was down regulated while CD30-CD30L
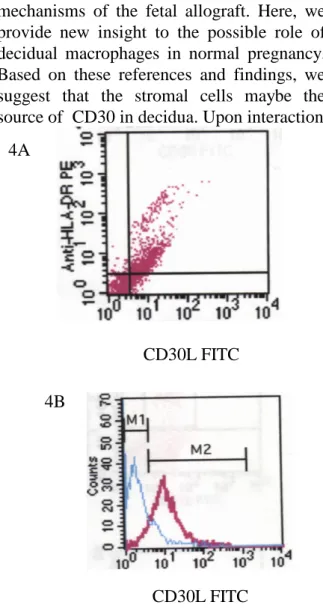
interaction. Thus, we determined the appearance of CD30L on HLA-DR+ cells and found HLA-DR+ cells were CD30L+ and the CD30L was medium expression. (Fig. 4A) Approximate 90% decidual macrophages express CD30L. (Fig.4B)
To summarize, we found that almost HLA-DR+ cells have CD11b and CD30L expression on the cell surface. It means that there are HLA-DR and CD 30L co-expression at the same time on decidual macrophages. It’s a new finding in this field. Besides, these cells were CD80- CD86low.
It has been suggested “suppressor macrophages” maybe one of the tolerance Fig. 2 CD11b+ cells were slightly CD86
positive. A. Single cells were stained with CD11b-FITC and CD86-PE. B. CD11b+ cells were gated and analyzed the CD86 expression. The blue curve represents negative control and the red curve stands for CD86 expression among CD11b+ cells.
Fig. 3 CD11b+ cells were entirely CD80 negative. A. Single cells were stained with CD11b-FITC and CD80-PE. B. CD11b+ cells were gated and analyzed the CD80 expression. The blue curve represents negative control and the red curve stands for CD80 expression among CD11b+ cells. All CD11b+ cells didn’t express CD80 on the surface. 2A 2B ..,. 3A 3B
mechanisms of the fetal allograft. Here, we provide new insight to the possible role of decidual macrophages in normal pregnancy. Based on these references and findings, we suggest that the stromal cells maybe the source of CD30 in decidua. Upon interaction
between CD30 on stromal cells and CD30L on decidual macrophages, the antigen presentation ability of decidual macrophages was diminished by down regulation of co-stimulator (CD80/CD86) expression. Thus, T cells infiltrated in deciduas become anergy and tolerant to the fetus. But what’s the exact mechanism that decidual macrophages utilized remains unclear and need further investigation.
四、計畫成果自評
In the current study, we demonstrated at
the first time that decidual macrophages have CD30L on the surface. The unique molecule may provide some signaling while ligating to the corresponding receptor, CD30. Although we have proposed one possibility to explain the phenomenon observed here, it needs more experiments and solid findings to support our suggestion. At the same time, we have been trying to isolate these decidual macrophages for further study with limited success. Currently we have finished the study in 24 normal pregnancies and 12 anembryonic pregnancies. To clarify the sophisticated network in deciduas in normal and abnormal pregnancies might answer certain important issues of the immunlogical functions of those macro- phages. And it will be benefitial to patients with reproductive disorders.
五、參考文獻
[1] Loke YW & King A. (2000) Immunological aspects of human implantation. J. Reprod. Fertil. –Suppl. 55: 83-90
[2] Haddad EK. Duclos AJ. & Baines MG. et al. (1997) Role of interferon-γ in the priming of decidual macrophages for nitric oxide pro-duction and early pregnancy loss. Cellular Immunology. 181: 68-75
[3] Krishnan L. Guilbert LJ. & Mosmann TR. et al. (1996) T helper 1 response against Leishmania major in pregnant C57BL/6 mice increases implantation failure and fetal resorptions. J. Immunol. 156: 653-662
[4] Lin H., Mosmann TR. & Wegmann TG. Et al. (1993) Synthesis of T helper 2-type cytokines at the maternal-fetal interface. J. Immunol. 151: 4562-4573
[5] Hunt JS. Miller L. & DeBrot BL. et al. (1997) Female steroid hormones regulate production of pro-inflammatory molecules in uterine leuko-cytes. J. Reprod. Immunol. 35: 87-99
[6] Kurpisz M. & Fernandez N. (1995) Immunology of Human Reproduction. BIOS Scientific Pub-lisfer Ltd., UK, pp. 313-334
[7] Vince GS. Starkey PM. & Redman CWG. Et al. (1990) Flow cytometric characterization of cell population in human pregnancy decidua and isolation of decidual macrophages. J. Immunol. Methods. 132: 181-189
Fig. 4 HLA-DR+ cells were CD30L+ and the CD30L was medium expression. A. Live cells were gated and analyzed surface CD30L-FITC (FL1) and HLA-DR PE (FL2) expression. B. HLA-DR+ cells were gated for further detection of CD30L expression.
4A
4B
CD30L FITC CD30L FITC
5
[8] Adeleye TA. Elder MG. & Sullivan MHF. (1996) Preparation of a population from human third trimester decidua. Human Reproduction. 11: 451-456
[9] Stewart IJ & Mitchell BS. (1991) The distribution of uterine macrophages in virgin and early pregnant mice. J. Anat. 179: 183-196
[10] Hunt JS & Robertson SA. (1996) Uterine macrophages and environmental programming for pregnancy success. J. Reprod. Immunol. 32: 1-25
[11] Hunt JS. (1989) Cytokine networks in the uteroplacental unit: macrophages as pivotal regulatory cells. J. Reprod. Immunol.16: 1-17
[12] Mizuno M., Aoki K. & Kimbara T. (1994) Functions of macrophages in human decidual tissue in early pregnancy. Am. J. Reprod. Immunol. 31: 180-188
[13] Pollard JW. Lin EY & Zhu L. (1998) Com-plexity in uterine macrophage responses to cytokines in mice. Biol. Reprod. 58: 1469-1475
[14] Haddad EK. Duclos AJ. & Baines MG. (1995) Early embryo loss is associated with local production of nitric oxide by decidual mono-nuclear cells. J. Exp. Med. 182: 1143-1152
[15] Greenbaum LA. Horowitz JB. & Bottomly K. et al. (1988) Autocrine growth of CD4+ T cells. Differential effects of IL-1 of helper and inflammatory T cells. J. Immunol. 140: 1555
[16] Lichman AH. Chin J. & Abbas AK. (1988) Role of IL-1 in the activation of T lymphocytes. PNAS. 85: 9699
[17] Weaver CT.; Hawrylowicz CM. & Unanue ER. (1988) T helper cell subsets require the expression of distinct costimulatory signals by antigen-presenting cells. PNAS. 85: 8181
[18] Dinarello CA. (1996) Biologic basis for IL-1 in disease. Blood. 87: 2095
[19] De M. & Wood GW. (1990) Influence of estrogen and progesterone on macrophage distribution in the mouse uterus. J. Endo-crinology. 126: 417-424
[20] Wood GW. De M., Sanford T. & Choudhuri R. (1992) Macrophage colony stimulating factor controls macrophage recruitment to the cycling mouse uterus. Developmental Biology. 152: 336-343
[21] Wood GW. Hausmann E., & Choudhuri R. (1997) Relative role of CSF-1, MCP-1/JE, and RANTES in macrophages recruitment during successful pregnancy. Molecular Reproduction and development. 46: 62-70
[22] Wood GW. Hausmann E., & Kanakaraj K. (1999) Expression and regulation of chemokine genes in the mouse uterus during pregnancy. Cytokine. 11: 1038-1045
[23] Saito S. (2000) Cytokine network at the feto-maternal interface. J. Reprod. Immunol. 47: 87-103
[24] Chwalisz K., Winterhager E., Thienel T. & Garfield RE. (1999) Synergistic role of nitric oxide and progesterone during the establishment
of pregnancy in the rat. Human Reproduction. 14: 542-552
[25] Martinez SP., Viggiano M. & Villalón M. et al. (2000) Effect of nitric oxide synthase inhibitors on ovum transport and oviductal smooth muscle activity in the rat oviduct. J. Reprod. Fertil. 118: 111-117
[26] Ariel I., Hochberg A. & Shochina M. (1998) Endothelial nitric oxide synthase immuno-reactivity in early gestation and in trophoblastic disease. J. Clin. Pathol. 51: 427-431
[27] Norman J. (1996) Nitric oxide and the myo-metrium. Pharmacol. Ther. 70: 91-100
[28] Hunt JS. Miller L., Vassmer D. & Croy BA. (1997) Expression of the inducible nitric oxide synthase gene in mouse uterine leukocytes and potential relationships with uterine function during pregnancy. Biol. of Reprod. 57: 827-836
[29] Genoveva Durán-Reyes, María del Rocío Gómez-Meléndez & Juan José Hicks-Gómez et al. (1999) Nitric oxide synthesis inhibition suppresses implantation and decreases cGMP concentration and protein peroxidation. Life sciences. 65: 2259-2268
[30] Liew FY. (1995) Regulation of lymphocyte functions by nitric oxide. Curr. Opin. Immunol. 7: 396-399
[31] Stuehr DJ. & Marletta MA. (1985) Mammalian nitrate biosysthesis: mouse macrophages pro-duce nitrite and nitrate in response to Escherichia coli lipopolysaccharide. Proc. Natl. Acad. Sci. USA 82: 7738-7742
[32] Stuehr DJ. & Marletta MA. (1987) Induction of nitrite/nitrate synthesis in murine macrophages by BCG infection, lymphokines, or interferon-γ. J. Immunol. 139: 518-525
[33] Cox FEG. & Liew FY. (1992) T-cell subsets and cytokines in parasitic infections. Immunology Today. 13: 445-448
[34] Iñigo Angulo, Joaquin Rullas & José Luis Subiza et al. (2000) Early myeloid cells are high producers of nitric oxide upon CD40 plus IFN-γ stimulation through a mechanism dependent on endogenous TNF-α and IL-1α. Eur. J. Immunol. 30: 1263-1271
[35] Niedbala W., Wei X. –Q. & Liew FY. Et al. (1999) Effects of nitric oxide on the induction and differentiation of Th1 cells. Eur. J. Immunol. 29: 2498-2505
[36] Taylor-Robinson AW. Liew FY. & Phillips RS.
Et al. (1994) Regulation of the immune response by nitric oxide differentially produced by T helper type 1 and T helper type 2 cells. Eur. J. Immunol. 24: 980-984
[37] Barnes PJ. & Liew FY. (1995) Nitric oxide and asthmatic inflammation. Immunol. Today. 16: 128-130
[38] Chang R. -H., Lin Feng M. -H., Liu W. -H. & Lai M. -Z. (1997) Nitric oxide increased interleukin-4 expression in T lymphocytes.
[39] Wei X. -Q., Charles IG. & Liew FY. (1995) Altered immune responses in mice lacking inducible nitric oxide synthase. Nature. 375: 408-411
[40] Bauer H. Jung T. & Neumann C. et al. (1997) Nitric oxide inhibits the secretion of T-helper 1-and T-helper 2-associated cytokines in activated human T cells. Immunology. 90: 205-211
[41] Mills CD. (1991) Molecular basis of “supper-ssor” macrophages: arginine metabolism via the nitric oxide synthethase pathway. J. Immunol. 146: 2719-2723
[42] Kawabe T., Isobe K. -I. & Shimokata K. et al. (1992) Immunosuppressive activity induced by nitric oxide in culture supernatant of activated rat alveolar macrophages. Immunology. 76: 72-78
[43] Bilyk N. & Holt PG. (1995) Cytokine modulation of the immunosuppressive pheno- type of pulmonary alveolar macrophage popula- tions.
Immunology. 86: 231-237
[44] Shigeki Nabeshima, Mari Nomoto & Kikuo Nomoto et al. (1999) T-cell hyporesponsiveness induced by activated macrophages through nitric oxide production in mice infected with Myco-bacterium tuberculosis. Infection and Immunity. 67: 3221-3226
[45] Roel C. van der Veen, Dietlin TA. Gray JD. & Gilmore W. (2000) Macrophage-derived nitric oxide inhibits the proliferation of activated T helper cells and is induced during antigenic stimulation of resting T cells. Cellular Imm-unology. 199: 43-49
[46] Trincheri G. (1993) Interleukin-12 and its role in the generation of TH1 cells. Immunology Today. 14: 335-337
[47] Stout RD. (1993) Macrophage activation by T cells: cognate and non-cognate signals. Curr. Opin. Immunol. 5:398-403
[48] Modolell M., Corraliza IM. & Eichmann K. et al. (1995) Reciprocal regulation of the nitric oxide synthase / arginase balance in mouse bone marrow-derived macrophages by TH1 and TH2 cytokines. Eur. J. Immunol. 25: 1101-1104
[49] Liew FY. Li Y. & Moncada S. et al. (1991) A possible novel pathway of regulation by murine T helper type-2 (Th2) cells of a Th1 cell activity via the modulation of the induction of nitric oxide synthase on macrophages. Eur. J. Immunol. 21: 2489-24