Journal
of
Orthopaedic
ELSEVIER
Journal of Orthopaedic Research 30 (2002) 295-302Research
www.elsevier.com/locate/orthresNitric oxide induces osteoblast apoptosis through
the de novo synthesis of
Bax protein
*
Ruei-Ming Chen
a,Hwa-Chang Liu
b,*,Yi-Ling Lin
a,Wen-Chyi Jean
',
Jui-Shui Chen
b,Jyh-Horng Wang
''
Deptrrtinent of Anesthesiology. Taipei Medical College. Wan-Fang Hospital, N o 11 I , Sec. 3, Hsing-Lung Road, Taipei 116, T a i w ~ ~ n . ROC Department of Orthopedic Surgery, National Taiwan Unioersity Hospital. No. 7, Chung-Shun South Road, Taipei 100, Taiwan, R O CAbstract
Nitric oxide (NO) plays a crucial role in the physiological and pathophysiological regulations of osteoblast functions. This study
is designed to evaluate the toxic effects of NO released by sodium nitroprusside (SNP), an
NOdonor, on neonatal Wistar rat
calvarial osteoblasts from the analyses of cell viability, alkaline phosphatase (ALP) activity, cell morphology, apoptotic cells, ter-
minal deoxynucleotidyl transferase-mediated dUTP nick end-label (TUNEL) assay, DNA ladder, and immunocytochemistry and
Western blot for proapoptotic Bax protein. SNP increased the levels of nitrite, an oxidative product of NO, in the culture medium of
osteoblasts in concentration- and time-dependent manners, and altered cell morphologies to round and shrinkage shapes. Ad-
ministration
of osteoblasts with SNP resulted in concentration- and time-dependent decreases of cell viability and ALP activity.
Analysis of apoptotic cells revealed that SNP increased the percentages
ofosteoblasts processing apoptosis. Analyses of TUNEL
and DNA ladder showed that SNP caused DNA fragmentation. Pretreatment with cycloheximide, an inhibitor of protein synthesis,
partially blocked SNP-induced osteoblast apoptosis. Imunocytochemical and immunoblotting analyses revealed that SN P increased
Bax protein in osteoblasts. This study suggests that SNP could increase the levels of NO in osteoblasts, and cause osteoblast
apoptosis possibly through the de novo synthesis of proapoptotic Bax protein.
0
3003 Orthopaedic Research Society. Published byElsevier Science Ltd. All rights reserved.
Introduction
Osteoblasts play an important role in bone metabo-
lism. One of the primary functions
of
bone tissues is
structural support. Bone structure is maintained by bone
remodeling, a process that is characterized by the cou-
pling of osteoblast-mediated bone formation and osteo-
clast-mediated bone resorption [33]. Imbalance in the
remodeling process would lead to pathophysiological
diseases such as osteoporosis and osteoarthritis [9].
Varieties of systemic and local mediators modulate os-
teoblast or osteoclast metabolism and contribute to
regulation of bone remodeling [9,13].
Nitric oxide (NO) is a gaseous free radical synthesized
from L-arginine by calcium-dependent constitutive or
calcium-independent inducible
NO synthases [31,28,38].
'*Part of this study has been presented at the 46th Annual Meeting of the Orthopaedic Research Society, March 2000, Orlando, FL, USA.
* Corresponding author. Tel.: +886-2-23123456 ext. 5688; fax: +886- 2-23956988.
E-muil address: hcliu@~ha.mc.ntu.edu.tw (H.-C. Liu).
The diatomic free radical has been implicated as an im-
portant regulator of vaso-regulation, neuronal trans-
mission, immune response, and cell apoptosis [3,38]. NO
is detectable in control osteoblasts and is also inducible
after treatment
of
proinflammatory cytokines, mechan-
ical stress, or fluid flow [10,14,19,34-36,391. NO has the
potential role to modulate proliferation and differenti-
ation
of osteoblasts and bone resorption activity of
osteoclasts, and to mediate the effects of proinflamma-
tory cytokines and mechanical stress. The free radical
is able to regulate bone maintenance and remodeling
[5,8,15,22]. In an animal model of inflammation-induced
osteoporosis, NO is induced and associated with the
reduction of osteoblast numbers, the increase of osteo-
clast numbers, and the decrease of bone mineral density
[l]. The induction
of
NO in osteoblasts following treat-
ment with proinflammatory cytokines promotes cell
death
[11,27]. However, the death mechanism induced
by
NO is still little known.
This study is designed to evaluate the cytotoxic effects
of NO on osteoblasts. Sodium nitroprusside (SNP) is an
NO donor used to be the source of NO. The use of NO
0736-0266/02/$ - see front matter 0 3002 Orthopaedic Research Society. Published by Elsevier Science Ltd. All rights reserved P1I : S 0 7 3 6 - 0 2 6 6 ( 0 1 ) 0 0 0 8 6 - 9
296 R.-M. Chen cet ul. I Journal of Ortlzopuedic Rescurch 170 (17003) 295-3017
donors has biochemical advantages because the donors
permit the investigation of NO'S role in signaling trans-
duction pathways without interfering with NOS-
involved second messenger systems. SNP is a clinical
medicine that is decomposed to NO under the presence
of biological systems, reducing agents or visible light [2].
Administration of osteoblasts with SNP has been re-
ported to affect osteoblastic metabolism [3
11.
To eval-
uate the toxic effects of NO on osteoblasts, the bone
cells are prepared from neonatal Wistar rat calvariae
and treated with
SNP.
The cytotoxic mechanism is de-
termined from the aspects of the cells' morphologies,
ALP activity, cell viability, apoptotic assay, DNA frag-
mentation, and immunocytochemical and immunoblot-
ting analyses for proapoptotic Bax protein.
Materials and methods
Osteoblast prepurution, culture, S N P treutrnent and NO deeterminution Osteoblasts were prepared from 3-day-old Wistar rat calvariae following the sequentially enzymatic digestion method as described previously [33]. The primary osteoblasts were maintained in Dul- becco's modified Eagle medium (DMEM) (Gibco, BRL, Grand Island, NY, USA) supplemented with a IO'Yn heat-inactivated fetal bovine serum and 100 unitslml penicillin and 100 pg/ml streptomycin in 250 ml polystyrene tissue culture flasks at 37°C in a humidified atmosphere of 5% COz. SNP (Sigma, St. Louis, MO, USA) was dissolved in a PBS buffer (0.14 M NaCI, 2.6 mM KCI, 8 mM Na2HP04, 1.5 mM K H2 P0 4 ) to 200 mM as a stock solution, stored at -20°C and pro- tected from light for use in related experiments. Osteoblasts were treated with various concentrations of SNP for different time intervals. Osteoblast morphologies were observed and photographed by a re- verse-phase microscope. The amounts of nitrite in the culture medium of osteoblasts were determined by the protocol provided in the Bi- oxytech NO assay kit (OXIS International, Portland, OR, USA). A.ssuys 11j cell uiahility und A L P uc-tirity
After treatment with SNP, osteoblast viability was determined by a colorimetric 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazoliuin bro- mide (MTT) assay described previously [4]. ALP activity of osteoblasts was determined by detecting the formation ofg-nitrophenol, a product of p-nitrophenyl phosphate catalyzed by ALP, following the colori- metric procedure provided in Sigma Diagnostics Alkaline, Acid and Prostatic Acid Phosphotase kit (Sigma Diagnostics, St. Louis, MO, USA).
Analysis of upoptotic cL"s
Using a flow cytometer, apoptotic osteoblasts were determined to detect DNA fragments in nuclei stained by propidium iodide according to the method of Nicoletti et al. [29]. After treatment, osteoblasts were harvested and fixed in cold 80% ethanol. Following a process of cen- trifugation and washing, the fixed cells were stained with propidium iodide and analyzed using a FACScan flow-cytometer (FACS Calibur, Becton Dickinson, San Joes, CA, USA) on the basis of a 560 nm dichroic mirror and a 600 nm band pass filter.
T U N E L ussuy
After treatment, untreated and SNP-treated osteoblasts were har- vested. and DNA fragmentation in osteoblastic nuclei was detected following the protocol of Oncogene Research Products TdT-FragEL" DNA Fragmentation Detection Kit (Oncogene Research Products, MA, USA).
D N A ludder unulysi.,
Genomic DNA from control and SNP-treated osteoblasts were analyzed by a classical DNA electrophoresis method for the determi- nation of the DNA ladder. Following treatment, osteoblasts were harvested and lysed with a lysis buffer ( 5 % sarcosyl, 10 mM Tris-CI, 10 mM EDTA and 20 units of proteinase K ) at 50°C overnight. The rat osteoblast lysate was treated with 10 pg DNase-free RNase for 1 hour, and then was extracted with phenol-chloroform solution several times. The water layer was electrophoretically separated in a 1 .?"In agarose gel containing 0.1 pglml ethedium bromide. The DNA bands were visu- alized and photographed under UV-light exposure.
Irnn~unoi~ytochernicul anulysis
Control and SNP-treated osteoblasts were harvested for immuno- cytochemical analysis of Bax protein following the standard protocol provided by the VECTASTAIN ABC kit (Vector Laboratories, CA, USA). After treatment, osteoblasts were harvested, and spun to mi- croslides using a cytospin-3 (Shandon Scientific Limited. Cheshire, England). The slides were fixed in 80% ethanol, incubated with 1%
normal fetal bovine serum for 20 min, and reacted with the mouse monoclonal antibody against rat Bax protein, which is diluted 1: 100 in TBS buffer (10 mM Tris-C1 pH 7.5, 100 mM NaCI, 0.1% Tween 20) for another 2 h. After washing, the slides were incubated with a mouse IgG antibody conjugated with horseradish peroxidase that is diluted I :I00 at room temperature for 45 min. The slides were colorimetric after reacting with 3,3'-diaminobenzidine and hydrogen peroxide. Irnmunoblotting unulysis
Sodium dodecyl sulfate-polyacryamide gel electrophoresis (SDS- PAGE) and protein transfer were carried out following the method of Chen et al. [7]. Immunodetection of Bax protein was determined using a mouse monoclonal antibody against rat Bax protein (R&D Systems, MN, USA). The intensities of protein bands in the Western blots were quantified with the aid of an IS-1000 Digital Imaging system (Alpha Innotech, San Leandro, CA, USA) as described elsewhere [6].
Stutisticul analysis
The statistical significance of difference between control- and drug- treated groups was evaluated by Student's t-test. A P value <0.05 was considered statistically significant. The difference between drug-treated groups was considered to have a statistical significance when a P value of Duncan Multiple Rang test was less than 0.05.
Results
Nitrite, an oxidative product of
NO, was detectable
in untreated osteoblasts, and exposure to
0.5, 1.0, 1.5
and 2.0 mM SNP for 16 h resulted in 41940, 63940, fivefold
and sixfold increases of nitrite, respectively (Table
1).
Administration of osteoblasts with
1.5 and 2 mM SNP
caused 29% and 58% osteoblast death, respectively, but
SNP at <1 mM was not cytotoxic to the cells. SNP at
0.5,
1.0, 1.5 and 2 mM decreased 24% 52940, 55% and
59% ALP activity in osteoblasts, respectively.
Nitrite in the culture medium of osteoblasts pre-
treated with 2
mM
SNP for 2, 4,
8, 16 and 24 h was
augmented by about 2-, 3-,
5-,
6- and %folds, respec-
tively (Table
2).
Treatment of osteoblasts with 2 mM
SNP for
8, 16 and 24 h caused 18%. 58% and 69% cell
death, respectively. ALP activity of osteoblast exposed
to 2 mM SNP for 4, 8,
16 and 24 11 was reduced by 16%1,
R . - h t Chen rt ul. I Journal of Orthopurdic Rescarch 20 (2002 I 295302 297 Table 1
Concentration-dependent effects of SNP on nitrite production os- teoblastic viability and ALP activity
~
S N P Nitrite Cell viability ALP activity
( m M ) ( P M ) (OD at 570 nm) (OD at 410 nm) 0 3.2 i 0.6 0.86 f 0.12 0.63 f 0.08 0.1 4.0 f 0.7 0.80 f 0.09 0.53 f 0.07 0.5 4 . 5 f O . P 0.91 f O . 1 1 0.48 f 0.04* 1
.o
5.2 f 0.8* 0.95 f 0.07 0.30 f 0.07" 1.5 14.9 f 3.V 0.61 f 0.06' 0.28 f 0.06' 2.0 18.6*
5.2* 0.36 f 0.05* 0.26 f 0.02' Osteoblasts were treated with 0. 0.1, 0.5, 1.0, 1.5 and 2.0 mM SNP for16 h. The levels of nitrite in culture medium were assayed by the Griess reaction. Osteoblastic viability and ALP activity were determined us- ing colometric methods as described in Materials and Methods. Each value was represented Mean
*
S.E. for n = 6 .*Value significantly different from the respective control, P
<
0.05.Table 2
Time-dependent effects of SNP on nitrite production osteoblastic vi- ability and ALP activity
Time Cell viability ALP activity Nitrite (pM) ( h ) (OD at 570 nm) (OD at 410 nm) 0 1.03 f 0.09 0.75 f 0.06 2.6 f 0.7 I 0.96 f 0.02 0.73 f 0.06 5.3 3~ 0.6" 4 0.98 f 0.02 0.46 f 0.02' 8.1 i 1.4' 8 0.84 f 0.03' 0.41 f 0.01* 13.8 f 2.1' 16 0.43 i 0.02' 0.32
*
0.02 1 6 . 5 1 1.9* 24 0.32 f 0.02' 0.25 f 0.03' 21.4 f 3.2* 9Osteoblasts were treated with 2 mM SNP for 0, 2, 4, 8, 16 and 24 h. Viability and ALP activity of osteoblasts were determined using col- ometric methods as described in Materials and Methods. The levels of nitrite in culture medium were detected by the Griess reaction. Each value was represented Mean f S.E. for n = 6.
*Value significantly different from the respective control, P
<
0.05.25%, 50%
and
70% respectively decreases of ALP ac-
tivities, respectively.
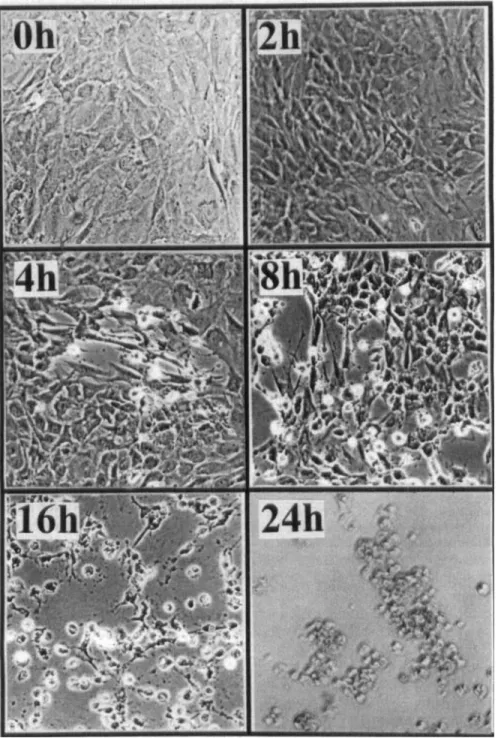
Osteoblast morphologies were observed and photo-
graphed following treatments of
2 mM SNP for 2, 4, 8,
16 and 24 h. In the 4 h-treated group, some osteoblasts'
morphologies were altered by SNP (Fig.
1). In the 8 h-
treated group, rounded and floated osteoblasts were
markedly observed. In the 16 h-treated group,
500/0
os-
teoblasts became shrunk, rounded and floated. In the 24
h-treated group, almost
80'% of the osteoblasts were
detached and floated.
Exposure of osteoblasts to 1.5 and 2 mM SNP for 16
h led to
17Y0 and 65% cells undergoing apoptosis, re-
spectively (Table
3 ) . The percentages of apoptotic cells
in osteoblasts pretreated with 2 mM SNP for 4, 8, 16
and 24h were increased by 2.5'1/0,
7.3% 61% and 71%
respectively. There was no statistical difference between
the control and
2 h-treated groups. TUNEL assay re-
vealed that SNP caused DNA fragmentation in
os-
teoblastic nuclei (Fig. 3). Genomic DNA from the
control and SNP-treated osteoblasts were isolated and
electrophoretically separated in agarose gel (Fig. 3).
Administration of osteoblasts with SNP caused 400
base-pair fragments of the DNA ladder, but the effect
was not observed in untreated osteoblasts (lanes 2 and
3).
Exposure of osteoblasts to cycloheximide, an inhibi-
tor of protein translation, did not affect cell apoptosis
(Fig. 4). Pretreatment with cycloheximide for
30 min
and then co-treated with SNP partially blocked
SNP-
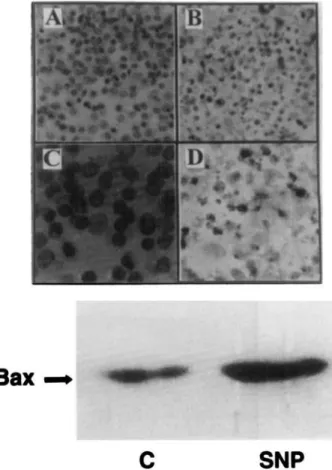
induced osteoblast apoptosis. Immunocytochemical
analysis using a mouse monoclonal antibody against rat
Bax protein showed that SNP increased the levels of Bax
protein in osteoblasts with brown signals (Fig. 5 , top
panel, B and D). In comparison, control osteoblasts
presented blue signals (Fig. 5, top panel, A and C ) .
Immunoblotting analysis revealed that SNP caused a
fivefold increase in the intensity of Bax protein (Fig. 5,
bottom panel).
Discussion
Administration of SNP increases the levels of NO in
osteoblasts and further causes oxidative stress to the
bone cells. SNP is a clinical medicine used as a vasodi-
lator for vasocardiac diseases because the NO donor can
be decomposed to
NO, a messaging molecule for re-
laxation of blood vessels [2]. The detailed mechanism of
SNP decomposition to
NO is still unknown. Several
lines of evidence have reported that the NO donor can
be decomposed
to
NO
under the presence of biological
systems, reducing agents or visible light [2,20]. N O has a
short half-life to be easily oxidized to nitrite and nitrate
[37]. The present study reveals that the levels of nitrite in
rat osteoblasts were concentration- and time-depen-
dently augmented following SNP treatment. NO is a
reactive oxygen species, whose increase of the free radi-
cal will result in the increase of oxidative stress on os-
teoblasts and the further modulation of osteoblastic
physiology or pathophysiology.
NO could regulate osteoblast survival and metabo-
lism. In parallel
to the increase of
NO in osteoblasts,
SNP causes concentration- and time-dependent de-
creases of osteoblast viability. Osteoblasts play a crucial
role in bone formation. Suppression of osteoblast vi-
ability or proliferation would interfere with the dynamic
balance of bone remodeling, and lead to pathophysio-
logical conditions
of
bone tissues
[9].
Previous studies
reveal that NO, over-induced by proinflammatory cyto-
kines, has a pathogenic role when it acts as a mediator
of these cytokines for the stimulation of osteoblast death
[22,27]. This study shows that
NO released by
an
extracellular NO donor also has the similar cytotoxic
effects on osteoblasts as intracellular NO induction.
ALP is a marker enzyme for osteoblast metabolism
[9]. Treatment of osteoblasts with SNP results in
298
Fig. 1. Effects of SNP on alternations of osteoblast morphologies. Osteoblasts prepared from neonatal Wistar rat calvariae were treated with 2 mM SNP for 0, 2, 4, 8, 16 and 24 h. Cell morphologies were observed and photographed using a reverse phase-contrast microscopy (100s).
concentration- and time-dependent decreases of ALP
activity. The reduction of ALP activity caused by NO
may be partially due to the cytotoxic effects of
NO on
osteoblasts. McPherson et al. [23] reported that the NO
donors SNP and SNAP only at high concentrations
(>O.
1 mM) were able to inhibited cell viability and ALP
activity. The present data has the similar results that low
concentrations of SNP
(<O.
1 mM) were not cytotoxic to
osteoblasts, however, SNP at high concentrations would
modulate osteoblast metabolism and lead to cell death.
There are several lines of evidence presented here to
demonstrate that NO induces osteoblast death mainly
through the apoptotic signaling pathway. Apoptosis,
also known as programmed cell death, is an energy-
dependent model
of cell death. Cells processing apop-
tosis possess certain characteristics, including typically
morphological alternation, as well as chromatin con-
densation and cleavage [25,26]. From morphological
observations, the present study has shown that admin-
istration of SNP changes osteoblast shapes into shrun-
R.-M Chen et 01. I Journul o f Orthopucdil. Rrseurclz 20 (2002) 295-302 299 Table 3
Concentration- and time-dependent effects of SNP on osteoblast apoptosis
Apoptotic cells ("YO)
SNP (mM) Time ( h ) 0 1.5 50 .3 0 1.3 hO.4 0. I 1.010.1 I l.S+0.4 0.5 0.9 f 0.2 4 2.5 50.3" 1
.o
1.8 f0.4 8 7.3 f 1.7' 1.5 26.5 f 6.9' 16 60.7 5 9.3' 2.0 65.1 f 5.8' 24 70.6 & 2.5' 7Osteoblasts were treated with 0, 0.1, 0.5, 1.0, 1.5 and 2.0 mM SNP for 16 h or with 2 mM SNP for 0, 2, 4, 8, 16 and 24 h. The percentages of apoptotic cells in osteoblasts were determined by a flow cytometric method as described in Materials and Methods. Each value was rep- resented Mean f S . E . for n = 6.
*Value significantly different from the respective control,
P
<
0.05.ken and rounded outlines. According to the TUNEL
assay and DNA electrophoresis, this study has revealed
that SNP can cause DNA fragmentation. Cells pro-
cessing apoptosis can be detected by a flow cytometer to
determine unequivocal hypodiploid DNA peaks derived
from the reduced DNA content in apoptotic nuclei
following PI staining [29]. The percentage of apoptotic
cells in osteoblasts exposed to SNP apparently increases.
This is the first demonstration that NO induces apop-
tosis of primary normal rat calvarial osteoblasts from
various aspects of cell morphology, apoptotic cells, and
DNA fragmentation. Dypbukt et al. [12] report that
high concentrations of NO will cause
LDH
release
and necrotic cell death. SNP at high concentrations has
the possibility of inducing osteoblast death partially
through necrotic mechanisms.
CONTROL
NO could directly or indirectly damage osteoblast
DNA.
The roles of NO in cells have been implicated to
have protective, regulatory, or deleterious effects [37]. As
a pathophysiological molecular effector, the chemical
biology of NO can be divided into direct and indirect
pathways. NO is a high energy radical and has the
ability to cause DNA fragmentation by direct attack or
indirect activation of serious signal transduction [3,37].
Previous studies have reported that NO induced by
proinflammatory cytokines, tumor necrosis
factor-ell,
interleukin-1p and interferon-y, or an NO donor, S-
nitroso-N-acetyl-D,L-penicillamine
would lead to DNA
fragmentation and cell death of a mouse clonal osteo-
genic cell line, MC3T3-El cell [11,27]. The present
study, using the primary culture system of normal rat
calvarial osteoblasts, has shown that administration of
the bone cells with SNP causes nuclear DNA fragmen-
tation by the analyses of apoptotic cells and the DNA
ladder. Therefore, it is possible that NO at high concen-
tration would directly or indirectly damage osteoblast
nuclei and lead to DNA fragmentation.
De novo synthesis of proapoptotic Bax protein could
be involved in the apoptotic mechanism of NO-induced
osteoblast death. Cycloheximide, an inhibitor of protein
translation, partially suppresses NO-induced osteoblast
apoptosis. This means that de novo protein synthesis is
involved in the mechanism of NO-induced osteoblast
apoptosis. Further evidence presented here from the
analyses of immunocytochemistry and Western blot re-
veal that SNP significantly elevates the levels of pro-
apototic Bax protein in osteoblasts. Bax protein is an
important component in the NO-induced p53 indepen-
dent apoptotic mechanism because the increase
of Bax
SNP
Fig. 2. SNP-induced DNA fragmentation in osteoblastic nuclei. Osteoblasts prepared from neonatal Wistar rat calvariae were treated with 2 mM SNP for 16 h. Control (C) and SNP-treated osteoblasts were harvested, and DNA fragmentation in osteoblastic nuclei were determined using the TUNEL assay as described in Materials and Methods ( 4 0 0 ~ ) .
300 R.-hf. Chrn et ul. I Journul of Orthopedic Research -70 (47002) 4795-302
M
C
8
1
1
r*
C CH SNP CH+SNP
Fig. 4. Protective effects of cycloheximide (CH ) on SNP-induced os- teoblast apoptosis. Osteoblasts prepared from neonatal rat calvariae were pretreated with 3.6 pM CH for 1 h, and then treated with 2 mM SNP for another 16 h. Apoptotic cells from control (C), CH-, SNP- and C H and SNP-treated osteoblasts were analyzed by flow cytometry as described in Materials and Methods. Each value was represented as MeanfS.E. for n = 9. Value significantly different from the respective control, P
<
0.05.1018
208
Fig. 3. SNP-induced DNA ladder in osteoblasts. Osteoblasts prepared from neonatal rat calvariae were treated with 2 mM SNP for 16 h. Genomic DNA from control ( C ) and SNP-treated osteoblasts were isolated and electrophoretically separated in 1.2% agarose gel con- taining 0.1 pglml ethedium bromide. M: DNA marker.
protein would unbalance the ratio of the proapoptotic
protein and antiapoptotic protein Bcl-3, then activate
caspases activities, and further leads to cell apoptosis [3].
Varieties of studies on different types of cells, including
T-cells, activated macrophages, primary hippocampal
neurons,
PC12, HelLa and HL-60 cells, have similar
results as the osteoblasts presented in this study
-that
the elevation of cellular Bax protein contributes
to cell
apoptosis [I 7,18,24,30]. Previous studies have shown
that accumulation
of p53, caspase activation and cyto-
chrome
L'relocation are involved in NO-induced cell
apoptosis [3]. Hikiji et al. [I61 show that peroxynitrite, a
combined oxidant
of
NO and superoxide, is able to
suppress
ALP activity and osteocalcin expression in
MC3T3-EI cells. Our other unpublished data reveals
that SNP increases intracellular reactive oxygen species
Bax
-
C
SNP
Fig. 5. Immunocytochemical and immunoblotting analyses of pro- apoptotic Bax protein in osteoblasts. Osteoblasts prepared from neo- natal rat calvariae were treated with 2 mM SNP for 16 h. The intact cells and cytosolic proteins were prepared for immunocytochemicdl (top panel) and immunoblotting (bottom panel) analyses, respectively. A mouse monoclonal antibody against rat Bax protein was used in these two assays to detect Bax protein in osteoblasts. The proteins were loaded at 100 pg. Top panel: A and C - control groups, B and D -
SNP-treated groups; A and B - Z O O * , C and D - 400 * . Bottom panel:
R.-M. Chen et al. I Journal of Orthopaedic Research 20 (2002) 295-302 30 1
and modulates mitochondria1 membrane potential.
Therefore, NO itself or its oxidative metabolite such as
peroxynitrite can directly induce or indirectly activate
biological signaling molecules or events to cause osteo-
blast apoptosis.
In conclusion, the present study has used primary
normal osteoblasts as an experimental model to reveal
that
SNP can increase the levels of NO, alternates cell
morphologies, decreases cell viability and
ALP
activity,
increases the percentage of apoptotic cells, and causes
DNA fragmentation. By checking the characteristics of
apoptosis, this study has demonstrated that SNP in-
duces osteoblast death possibly through an apoptotic
mechanism, and the mechanism may be involved by the
de novo synthesis of proapoptotic Bax protein. To find
other potential signaling effectors
or
pathways that
mediate NO-induced osteoblast apoptosis is our next
interesting subject.
Acknowledgements
This study was supported by grants NSC88-23 14-B-
002-256 and NSC89-23 14-B-002-396 from the National
Science Council, ROC.
References
[I] Armour KE, Van’T Hof RJ, Grabowski PS, Reid DM, Ralston SH. Evidence for a pathogenic role of nitric oxide in inflamma- tion-induced osteoporosis. J Bone Miner Res 1999;14:213742. [2] Bates JN, Baker MT, Guerra Jr. R, Harrison DG. Nitric oxide
generation from nitroprusside by vascular tissue. Evidence that reduction of the nitroprusside anion and cyanide loss are required. Biochem Pharmacol 1991;42:s15745.
[3] Brune B, von Knethen A, Sandau KB. Nitric oxide (N0):an effector of apoptosis. Cell Death Diff 1999;6:969-75.
[4] Carmichael J, DeGraff WG, Gazdar AF, Minna JD, Mitchell JB. Evaluation of a tetrazolium-based semiautomated colorimetric assay:assessment of chemosensitivity testing. Cancer Res 1987; 47:93642.
[5] Chae HJ, Park RK, Chung HT, Kang JS, Kim MS, Choi DY, Bang BG, Kim HR. Nitric oxide is a regulator of bone remodelling. J Pharm Pharmacol 1997;49:897-902.
[6] Chen RM, Chou MW, Ueng TH. Induction of cytochrome P450 1A in hamster liver and lung by 6-nitrochrysene. Arch Toxicol 1998;72:395401.
[7] Chen RM, Chou MW, Ueng TH. Induction of cytochrome P450 IA l in human hepatoma HepG2 cells by 6-nitrochrysene, Toxicol Lett 2000;117:69-77.
[8] Chow JW, Fox SW, Lean JM, Chambers TJ. Role of nitric oxide and prostaglandins in mechanically induced bone formation. J Bone Miner Res 1998;13:1039-44.
[9] Collin-Osdoby P, Nickols GA, Osdoby P. Bone cell function, regulation, and communication:a role for nitric oxide. J Cell Biochem 1995;57(2):399408.
[lo] Damoulis PD, Hauschka PV. Cytokines induce nitric oxide production in mouse osteoblasts. Biochem Biophys Res Commun 1994;201(2):92&31.
[I I] Damoulis PD, Hauschka PV. Nitric oxide acts in conjunction with proinflammatory cytokines to promote cell death in osteoblasts. J Bone Miner Res 1997;12:412-22.
[I21 Dypbukt JM, Ankarcrona M, Burkitt M, Sjoholm A, Strorn K, Orrenius S, Nicotera H. Different prooxidant levels stimulate growth, trigger apoptosis, or produce necrosis of insulin-secreting RINm5F cel1s:the role of intracellular polyamines. J Biol Chem [13] Evans DM, Ralston SH. Nitric oxide and bone. J Bone Miner Res
1996;11:300-5.
[I41 Heflich MH, Evans DE, Grabowski PS, Pollock JS, Ohshima H, Ralston SH. Expression of nitric oxide synthase isoforms in bone and bone cell cultures. J Bone Miner Res 1997;12:1108- 15.
[I51 Hikiji H, Shin WS, Oida S, Takato T, Koizumi T, Toyo-oka T. Direct action of nitric oxide on osteoblastic differentiation. FEBS Lett 1997;410:23842.
[I61 Hikiji H, Shin WS, Koizumi T, Takato T, Susami T, Koizumi Y, Okai-Matsuo Y, Yoyo-Oka T. Peroxynitrite production by TNF- a and IL-I b:implication for suppression of osteoblastic differen- tiation. Am J Physiol 2000;278:E1031-7.
[I71 Hortelano S, Bosca L. 6-Mercaptopurine decreases the Bcl-2IBax ratio and induces apoptosis in activated splenic B lymphocytes. Mol Pharmacol 1997;5 1 :4 14-2 1.
[I81 Ina K et al. Resistance of Crohn’s disease T cells to multiple apoptotic signals is associated with a Bcl-2IBax mucosal imbal- ance. J Immunol 1999;163:1081-90.
[19] Johnson DL, McAllister TN, Frangos JA. Fluid flow stimulates rapid and continuous release of nitric oxide in osteoblasts. Am J Physiol 1996;271:E205-8.
[20] Kowaluk EA, Seth P, Fung HL. Metabolic activation of sodium nitroprusside to nitric oxide in vascular smooth muscle. J Pharmacol Exp Ther 1992;262:91&22.
[21] Lamas S, Marsden PA, Li GK, Tempst P, Michel T. Endothetial nitric oxide synthase:molecular cloning and characterization of a distinct constitutive enzyme isoform. Proc Natl Acad Sci USA 1992;89:6348-52.
[22] Lowik CW, Nibbering PH, van de Ruit M, Papapoulos SE. Inducible production of nitric oxide in osteoblast-like cells and in fetal mouse bone explants is associated with suppression of osteoclastic bone resorption. J Clin Invest 1994;93: 1465-72.
[23] MacPherson H, Noble BS, Ralston SH. Expression and functional role of nitric oxide synthase isoforms in human osteoblast-like cells. Bone 1999;24: 179-85.
[24] Matsuzaki H, Tamatani M, Mitsuda N, Namikawa K, Kiyama H, Mijake S, Tohyama M. Activation of Akt kinase inhibits apoptosis and changes in Bcl-2 and Bax expression induced by nitric oxide in primary hippocampal neurons. J Neurochem
1999;73:203746.
[25] Messmer UK, Bruen B. Nitric oxide-induced apoptosis: p53- dependent and p53-independent signalling pathways. Biochem J 1996;319:299-305.
[26] Messmer UK, Briine B. Nitric oxide (NO) in apoptotic versus necrotic RAW 264.7 macrophage cell death: the role of NO-donor exposure, NAD’ content, and p53 accumulation. Arch Biochem Biophys 1996;327: 1-10.
[27] Mogi M, Kinpara K, Kondo A, Togari A. Involvement of nitric oxide and biopterin in proinflammatory cytokine-induced apop- totic cell death in mouse osteoblastic cell line MC3T3-El. Biochem Pharmacol 1999;58(2):649-54.
[28] Moncada S, Palmer RM, Higgs EA. Nitric oxide: physiology, pathophysiology and pharmacology. Pharmacol Rev 1991;43:109- 42.
[29] Nicoletti I, Migliorati G, Pagliacci MC, Grignani F, Riccardi C. A rapid and simple method of measuring thymocyte apoptosis by 1994;269:30553-60.
302 R - M . Chen et al. I Journal of Orthopaedic Research 20 (2002) 295-302 propidium iodide staining and flow cytometry. J Immunol Meth
[30] Okuno S, Shimizu S, Ito T, Nomura M, Hamada E, Tsujimoto Y, Matsuda H. Bcl-2 prevents caspase-independent cell death. J Biol Chem 1998;273:34272-7.
[31] Otsuka E, Hirano K, Matsushita S, Inoue A, Hirose S, Yama- guchi A, Hagiwara H. Effects of nitric oxide from exogenous nitric oxide donors on osteoblastic metabolism. Eur J Pharmacol 1998; 34935-50.
[32] ParEtt AM. Bone remodeling and bone loss: understanding the pathophysiology of osteoporosis. Clin Obstet Gynecol 1987;30789-811.
[33] Partridge NC, Alcorn D, Michelangeli
VP,
Kemp BE, Ryan GB, Martin TJ. Functional properties of hormonally responsive cultured normal and malignant rat osteoblastic cells. Endocrinol- [34] Pitsillides AA, Rawlinson SC, Suswillo RF, Bourrin S, Zaman G, Lanyon LE. Mechanical strain-induced NO production by bone 1991;139271-9.ogy 1981;1082 13-9.
ce1ls:a possible role in adaptive bone (re)modeling. FASEB J 1995;9(2): 16 14-22.
[35] Ralston SH, Todd D, Helfrich M, Benjamin N, Grabowski PS. Human osteoblast-like cells produce nitric oxide and express in- ducible nitric oxide synthase. Endocrinology 1994; 135:330-6. [36] Riancho JA, Zarrabeitia MT, Fernandez-Luna JL, Gonzalez-
Macias J. Mechanisms controlling nitric oxide synthesis in osteoblasts. Mol Cell Endocrinol 1995;107(2):87-92.
[37] Wink DA, Mitchell JB. Chemical biology of nitric oxide: insights into regulatory, cytotoxic, and cytoprotective mechanisms of nitric oxide.
Free
Radic Biol Med 1998;25:43&56.[38] Xie QW, Cho HJ, Calaycay J, Mumford RA, Swiderek KM, Lee TD, Ding A, Troso T, Nathan C. Cloning and characterization of inducible nitric oxide synthase from mouse macrophages. Science 1992; 1256225-8.
1391 Zaman G et al. Mechanical strain stimulates nitric oxide produc- tion by rapid activation of endothelial nitric oxide synthase in osteocytes. J Bone Miner Res 1999;141123-31.