The methylation patterns of a disintegrin and metalloproteinase 33 gene (ADAM33) in adult asthma
Pei-Jing Yang 1, Ruei-Nian Li 2, Chi-Chih Huang 3,4, Tung-Heng Wang 3,4 ,Ying-Chin Ko 5,6, Ming-Shyan Huang 3,6#, Tsu-Nai Wang 1,6#.
1 Department of Public Health, College of Health Science, Kaohsiung Medical University, Kaohsiung, Taiwan
2. Department of Biomedical Science and Environmental Biology,
College of Life Science, Kaohsiung Medical University, Kaohsiung, Taiwan 3 Division of Pulmonary and Critical Care Medicine, Department of
Internal Medicine, Kaohsiung Medical University Hospital, Kaohsiung, Taiwan
4 Faculty of Respiratory Care, College of Medicine, Kaohsiung Medical University, Kaohsiung, Taiwan
5 Graduate Institute of Clinical Medical Science, China Medical University, Taichung, Taiwan
6 Center of Excellence for Environmental Medicine, Kaohsiung Medical University, Kaohsiung, Taiwan
# Correspondence and requests for reprints should be addressed to Dr. Tsu-Nai Wang and Ming-Shyan Huang
Department of Public Health, College of Health Science, Kaohsiung Medical University, Kaohsiung, No 100, Shih-Chuan 1st Rd, Kaohsiung, Taiwan.
Corresponding author: Dr. Tsu-Nai Wang and Ming-Shyan Huang # equal contribution for corresponding authors
Email: [email protected]
Fax: (886) 7-3110811 Phone: (886) 7-3121101-2141
Key words:adult asthma, methylation, epigenetics, ADAM33, gene
The methylation patterns of the disintegrin and metalloproteinase 33 gene (ADAM33) in adult asthma
Background: Asthma is a common chronic inflammatory respiratory disease. Previous studies have suggested that the pathogenesis of asthma may be affected by epigenetic regulation. The purpose of this study is to characterize the effect of the methylation of each CpG site in the ADAM33 (a disintegrin and metalloproteinase 33) gene on adult asthma. Methods: A human CpG island microarray was used to examine four asthmatic cases and four healthy controls, and the results suggested that there might be differences in the methylation within exon 9 of the ADAM33 gene. Therefore, we designed a case-control study with 50 asthmatic patients and 50 age- and sex-matched healthy controls to examine the relationship between the CpG methylation of the ADAM33 gene and asthma using bisulfite deoxyribonucleic acid modification and sequencing. Results: Bisulfite sequencing experiments showed that the 14 CpG sites in exon 9 of the ADAM33 gene were highly methylated (100%) in all individuals. The proportions of methylation of the 14 CpG sites in ADAM33 in the case group were not different from those of the control group. The methylation of exon 9 of this locus was not associated with age, sex, IgE levels or lung function. This study found no association between the methylation of CpG sites in exon 9 of the ADAM33 gene and adult asthma. Conclusions: The 14 CpG sites are highly methylated in the case and control groups. Further investigation of exon 9 in ADAM33 in a larger population is needed to evaluate its role in asthma.
Introduction
Asthma is one of the most common chronic diseases in children and adults. The disintegrin and metalloproteinase 33 gene (ADAM33), one of the members of the ADAM family, is located on chromosome 20p13. ADAM33 is expressed in lung fibroblasts and bronchial smooth muscle cells, and several single nucleotide polymorphisms (SNPs) in ADAM33 are associated with bronchial hyperresponsiveness (BHR) and asthma severity in China [1,2]. In addition, ADAM33 is composed of 812 amino acid residues and 22 exons [3].
Human DNA mainly consists of adenine, guanine, cytosine and thymine; however, the DNA of humans and other mammals also shows the covalent modification of cytosine with a methyl group without alterations in sequence, which is known as epigenetic regulation [4]. Previous studies have suggested that epigenetic regulation may partially regulate genes and cause asthma in response to exposure to environmental factors [5,6]. Yang et al. indicated that the hypermethylation of CpG sites in the ADAM33 promoter resulted in a loss of ADAM33 expression in bronchial epithelial cells. However, the hypomethylation of CpG sites in the ADAM33 promoter of bronchial fibroblasts leads to ADAM33 expression. In addition, no significant difference was identified in the expression of ADAM33 mRNA in bronchial biopsy specimens between healthy subjects and severe asthma patients [7]. Our study utilized a CpG island microarray to evaluate asthma-related target genes, and a ratio of >1.5 indicated that CpG sites displayed methylation. We further examined the DNA methylation status of exon 9 of ADAM33 in DNA extracted from peripheral blood leukocytes in the asthma and control groups. We performed methylation-specific PCR and bisulfite sequencing to investigate the association between the methylation status of ADAM33 and asthma.
Materials and methods
Subjects and clinical characteristics Patients
A total of 228 hospital-based asthmatic adults between the ages of 20 and 65 years old were recruited from the Division of Pulmonary and Critical Care Medicine of Kaohsiung Medical University in southern Taiwan. The participants were diagnosed with asthma by chest physicians according to the Global Initiative for Asthma (GINA) guidelines and/or spirometry demonstrating that the post-bronchodilator value increased at least 12% and 200 ml of FEV1 compared with the pre-bronchodilator value [8,9]. In total, 178 asthmatic patients were excluded: (1) 18 asthmatic patients suffered from chronic obstructive pulmonary disease (COPD), tuberculosis, emphysema or lung cancer; (2) 44 asthmatic patients were current smokers; and (3) 116 asthmatic patients were regularly taking oral cortical steroids. Only 50 patients were selected for further analysis of their methylation patterns.
The healthy controls came from communities in southern Taiwan. The control subjects were excluded if they were current smokers and had physician-diagnosed asthma, chronic obstructive pulmonary disease (COPD), tuberculosis, emphysema or lung cancer. The 50 community controls were matched 1:1 to the case groups by age (±5 years old) and sex. The asthma patients and healthy controls all agreed to complete a questionnaire; accept an assessment of lung function; provide measurements of weight, height, waist and hip circumferences and provide blood samples. Pulmonary function tests mainly included forced expiratory volume in the first second (FEV1) and forced vital capacity (FVC). The study subjects performed pulmonary function tests three times, and the highest score was recorded. The study protocol was approved by the Institutional Review Board of Kaohsiung Medical University. Written informed consent was obtained from each study subject.
Bioinformatics analysis CpG island microarray
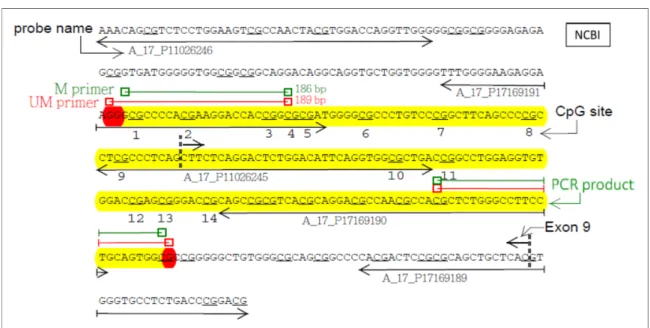
DNA was extracted using the commercial Purgene DNA isolation kit (Gentra Systems, Inc. Minneapolis, MN, USA). Only four asthmatic patients and four matched healthy controls were randomly selected for CpG island microarray analysis. The microarray used in this study was a customized 8x15K Human CpG island microarray with 243 genes that contained asthma-related genes from the Welgene Biotech Company (Agilent Methylation Microarray Service, Welgene Biotech. Co., Colorado, United States). The tiled arrays consisted of 60 oligomer probes spaced approximately 100 bp apart. Each gene contained 1 to 39 probes corresponding to different gene lengths (http://www.welgene.com.tw). Immunoprecipitated methylated DNA (IP DNA) was labeled with the Cy5 fluorophore (red), and the input genomic DNA (IN DNA) without immunoprecipitation was labeled with the Cy3 fluorophore (green). The IP DNA and IN DNA are competitively hybridized to the Agilent Human CpG Island Microarray [10,11].Feature extraction 10.5.1.1 software was used to analyze the microarray data. To ensure high reliability of our microarray data, we replicated five random experiments for each CpG site. The maximum and minimum ratios were deleted, and the ratios of the remaining 3 replicates were averaged for these CpG sites. A ratio of >1.5 indicated that the CpG sites displayed methylation [12,13]. In the Human CpG Island array, there are 22 probes in the ADAM33 gene. Only five probes (A_17_P17169189, A_17_P17169190, A_17_P11026245, A_17_P17169191 and A_17_P11026246) among the 22 probes have higher methylation levels in asthma cases than in controls. The expression ratio values of the above 5 probes were more than 1.5 in the asthma group, and the p-values were less than 0.05. These probes were then used to design the MSP primers.
Investigation of CpG islands and methylation analysis
The CpG islands in the ADAM33 gene (retrieved from the National Center for Biotechnology Information (NCBI) website) were identified using the CpG Islands Searcher website (http://www.cpgislands.com/). Our criteria for screening the CpG islands was ObsCpG/ExpCpG=0.7, %GC=60%, and the minimum length of a CpG island=500. The design of methylation-specific PCR (MSP) primers was performed using an online biological information website (http://urogene.org/methprimer /index1.html). The MSP product includes 14 CpG sites and partly maps to exon 9 of ADAM33. The size of the product amplified with the unmethylation (UM) primer was similar to the size of the product amplified with the methylation (M) primer [14,15]. The BISMA (the bisulfite sequencing DNA methylation analysis) website was used to perform the DNA methylation sequencing analysis (http://biochem.jacobs-university.de/BDPCBISMA/manual_ unique.php).
Methylation-specific PCR and bisulfite sequencing
Methylation bisulfate conversion and methylation-specific PCR were performed for our 100 study participants. Genomic DNA (200 ng) from the asthma and control subjects was modified with sodium bisulfite using the MethylCodeTM Bisulfite Conversion Kit
(Invitrogen Corporation, CA, USA) before conducting PCR. The bisulfite conversion process was performed using methylation-specific PCR (MSP). The M primer pair (methylated condition) of MSP resulted in a 186 bp product and included the forward primer 5’–GCGTTTTACGAAGGATTATCGGC-3’ and the reverse primer 5’– GCCACTACAAAAAAACCCAAAACG-3’. The UM primer pair of MSP for the unmethylated condition resulted in a 189 bp product and included the forward primer 5’-GGGTGTTTTATGAAGGATTATTGGT-3’ and the reverse primer 5’-CACCACT ACAAAAAAACCCAAAACA-3’. The methylation-specific PCR conditions were one
cycle of 95 C for 5 minutes; 40 cycles of 95 C for 30 seconds, 63 C for 1 minute, and 72 C for 30 seconds; and finally, 72 C for 30 seconds. Methylation-specific PCR was performed using the Universal Methylated Human DNA Standard kit (Zymo Research Corporation, CA, USA). Control primers were used as a positive control for bisulfite conversion processes, and methylation-specific PCR was performed without DNA as a negative control. The products of methylation-specific PCR were sequenced for all of the case and control subjects. Bisulfite sequencing was performed with the ABI Reaction Kit (ABI PRISM BigDye Terminator Cycle Sequencing Ready Reaction Kit 3.1, Applied Biosystems, CA, USA) and analyzed with an ABI Sequencer (ABI 3730 XL DNA Analyzer, Applied Biosystems, CA, USA).
Statistical analysis
Our study used an independent t-test to analyze the differences in age, IgE levels, body mass index (BMI), waist-hip ratio, lung function, and the results of the microarray between the case and control groups. The χ2 test was performed to evaluate the percentage
difference in the risk factors between the asthma and control groups. All of the statistical analyses were performed with the Statistical Package for the Social Sciences (SPSS) software (Version 14).
Results
Participant characteristics
The age and sex distributions of the case group were the same as the control group due to 1:1 matching for age and sex. The IgE concentrations and the predictive values of FEV1% and FVC% of the case group were significantly different from the control group (p<0.05). The BMI (p=0.212) and waist-hip ratios (p=0.123) of the case group were not different from the control group (Table 1).
CpG island Microarray
The 22 probes were located on exons l, 8, 9, 11, 12 and 13 of the ADAM33 gene. Five out of 22 probes in the CpG islands of ADAM33 gene (A_17_P17169189, A_17_P17169190, A_17_P11026245, A_17_P17169191 and A_17_P11026246) had a ratio value greater than 1.5 in the asthma group, which indicates the higher methylation levels (Table 2) (Figure 1A). Two probes, A_17_P11026245 and A_17_P17169191, contained CpG sites 1-5 and sites 9-10 of MSP products, respectively. The MSP amplification product includes CpG sites 1-14 and partially contains exon 9 of the ADAM33 gene. The amplification lengths of the methylation primer (M primer) (yellow) and unmethylation primer (UM primer) (red) are 186 bp and 189 bp, respectively (Figure 1A and 1B).
Methylation profiles
We found that CpG sites 1-14 were 100% methylated in both the case and control groups by bisulfite sequencing. We found that the methylation status of all 14 CpG sites of the ADAM33 gene were not associated with asthma (Figure 2).
Discussion
Many studies have found that polymorphisms within the ADAM33 gene are strongly associated with asthma and bronchial hyperresponsiveness [16-19]. To date, only one study has discussed the association between ADAM33 DNA methylation and asthma [7]. In the present study, we evaluated peripheral blood leukocyte methylation patterns in exon 9 of ADAM33 in adult asthmatics and controls. We found that the 14 CpG sites in exon 9 of the ADAM33 gene showed complete methylation in both the case and control groups. The methylation of the CpG sites in exon 9 of the ADAM33 gene is not associated with adult asthma, age, sex, IgE levels or lung function.
Based on CpG island microarray data in our study, we chose five probes on exon 9 of the ADAM33 gene to perform methylation-specific PCR and bisulfite sequencing based on a ratio of > 1.5. Previous studies have shown that the methylation status of an exon is correlated with malignancy and tumor stage [20,21]. Therefore, we hypothesized that the methylation status of the exon of the ADAM33 gene may be associated with its related diseases; the methylation level of the exon is noteworthy. However, our study showed that 14 CpG sites on exon 9 in ADAM33 were highly methylated, and there were no significant differences between the case and control groups. Similarly, Yang et al. found that the methylation status of the promoter of the ADAM33 gene was not associated with asthma [7].
Previous studies have shown that hypomethylation of the promoter of the ADAM33 gene drives the transcription and translation of ADAM33 and induces ADAM33 expression in airway fibroblasts, myofibroblasts and smooth muscles [22-25]. Van Eerdewegh et al. found that ADAM33 expression in lung fibroblasts and bronchial smooth muscle cells resulted in airway structural changes. Therefore, it could be speculated that an association exists between airway remodeling in asthma patients and ADAM33 [1,23]. A previous study also found that a demethylating agent (5-aza-2’-deoxycytidine) used to treat epithelial cells induced CpG island demethylation and ADAM33 expression [7].
Previous studies have indicated that tobacco can modify asthma airway inflammation and bronchial hyperresponsiveness in adult asthma patients [26], and smokers have reduced forced expiratory volume in one second (FEV1) compared with non-smokers [27]. Parental smoke exposure affects DNA methylation [28]. Therefore, we excluded the subjects who were smokers in the case and control groups to control for the confounding effect of smoking. Our study is a case-control study in which the temporal causality is not clear. Therefore, a prospective cohort study and confounding factors adjustment, such as diet and
occupational exposure, should be considered in further studies.
There are some limitations of this study. This study lacks consensus regarding the ideal tissue for analyzing methylation patterns in studies of respiratory diseases. Due to the difficulty of collecting other tissue types, we obtained DNA from peripheral blood leukocytes but not from airway epithelia. We could not ascertain whether the methylation patterns would reflect the patterns present in the airway; therefore, the selection of relevant tissue for the trait should be of high priority in the planning of future epigenetic studies. However, several studies have suggested that the detection of genetic alterations and methylation patterns in the peripheral blood DNA of cancer patients may serve as noninvasive markers for monitoring and diagnosing cancers in high-risk populations [29,30]. A recent study has demonstrated that the DNA methylation in blood was highly correlated with buccal cells (r>0.75) [31]. Futhermore, the sample size of the study was only 100 subjects and quite small; therefore, the findings applied to the large population group still needed further investigation. Our study found that the methylation status of exon 9 in the ADAM33 gene was not associated with adult asthma, and all 14 CpG sites showed 100% methylation in the study subjects. Exon 9 in ADAM33 was highly methylated, but further investigation of this region in a larger population is needed to evaluate its role in asthma.
Acknowledgments
This work was supported by grant NSC 99-2314-B-037-056-MY2, Taiwan.
References
1 Van Eerdewegh P, Little RD, Dupuis J, Del Mastro RG, Falls K, Simon J, Torrey D, Pandit S, McKenny J, Braunschweiger K, Walsh A, Liu Z, Hayward B, Folz C, Manning SP, Bawa A, Saracino L, Thackston M, Benchekroun Y, Capparell N, Wang M, Adair R, Feng Y, Dubois J, FitzGerald MG, Huang H, Gibson R, Allen KM, Pedan
A, Danzig MR, Umland SP, Egan RW, Cuss FM, Rorke S, Clough JB, Holloway JW, Holgate ST, Keith TP: Association of the adam33 gene with asthma and bronchial hyperresponsiveness. Nature 2002;418:426-430.
2 Jie Z, Hu Z, Bai C, Jin M: Adam33 gene polymorphisms associate with asthma susceptibility and severity in east china han population. J Asthma 48:979-985.
3 Orth P, Reichert P, Wang W, Prosise WW, Yarosh-Tomaine T, Hammond G, Ingram RN, Xiao L, Mirza UA, Zou J, Strickland C, Taremi SS, Le HV, Madison V: Crystal structure of the catalytic domain of human adam33. J Mol Biol 2004;335:129-137. 4 Nephew KP, Huang TH: Epigenetic gene silencing in cancer initiation and progression.
Cancer Lett 2003;190:125-133.
5 Miller RL, Ho SM: Environmental epigenetics and asthma: Current concepts and call for studies. Am J Respir Crit Care Med 2008;177:567-573.
6 Adcock IM, Tsaprouni L, Bhavsar P, Ito K: Epigenetic regulation of airway inflammation. Curr Opin Immunol 2007;19:694-700.
7 Yang Y, Haitchi HM, Cakebread J, Sammut D, Harvey A, Powell RM, Holloway JW, Howarth P, Holgate ST, Davies DE: Epigenetic mechanisms silence a disintegrin and metalloprotease 33 expression in bronchial epithelial cells. J Allergy Clin Immunol 2008;121:1393-1399, 1399 e1391-1314.
8 GINA: Global initiative for asthma. Global strategy for asthma management and prevention. Nhlbi/who workshop report. National institutes of health, bethesda National Heart Lung and Blood Institute 1995: publication number 95-3659.
9 Pellegrino R, Viegi G, Brusasco V, Crapo RO, Burgos F, Casaburi R, Coates A, van der Grinten CP, Gustafsson P, Hankinson J, Jensen R, Johnson DC, MacIntyre N, McKay R, Miller MR, Navajas D, Pedersen OF, Wanger J: Interpretative strategies for lung function tests. Eur Respir J 2005;26:948-968.
10 Vucic EA, Wilson IM, Campbell JM, Lam WL: Methylation analysis by DNA immunoprecipitation (medip). Methods Mol Biol 2009;556:141-153.
11 Mohn F, Weber M, Schubeler D, Roloff TC: Methylated DNA immunoprecipitation (medip). Methods Mol Biol 2009;507:55-64.
12 Grafodatskaya D, Choufani S, Ferreira JC, Butcher DT, Lou Y, Zhao C, Scherer SW, Weksberg R: Ebv transformation and cell culturing destabilizes DNA methylation in human lymphoblastoid cell lines. Genomics 2010;95:73-83.
13 Misawa A, Inoue J, Sugino Y, Hosoi H, Sugimoto T, Hosoda F, Ohki M, Imoto I, Inazawa J: Methylation-associated silencing of the nuclear receptor 1i2 gene in advanced-type neuroblastomas, identified by bacterial artificial chromosome array-based methylated cpg island amplification. Cancer Res 2005;65:10233-10242.
14 Licchesi JD, Herman JG: Methylation-specific pcr. Methods Mol Biol 2009;507:305-323.
15 Li LC, Dahiya R: Methprimer: Designing primers for methylation pcrs. Bioinformatics 2002;18:1427-1431.
16 Reijmerink NE, Kerkhof M, Koppelman GH, Gerritsen J, de Jongste JC, Smit HA, Brunekreef B, Postma DS: Smoke exposure interacts with adam33 polymorphisms in the development of lung function and hyperresponsiveness. Allergy 2009;64:898-904. 17 van Diemen CC, Postma DS, Vonk JM, Bruinenberg M, Schouten JP, Boezen HM: A
disintegrin and metalloprotease 33 polymorphisms and lung function decline in the general population. Am J Respir Crit Care Med 2005;172:329-333.
18 Raby BA, Silverman EK, Kwiatkowski DJ, Lange C, Lazarus R, Weiss ST: Adam33 polymorphisms and phenotype associations in childhood asthma. J Allergy Clin Immunol 2004;113:1071-1078.
19 Simpson A, Maniatis N, Jury F, Cakebread JA, Lowe LA, Holgate ST, Woodcock A, Ollier WE, Collins A, Custovic A, Holloway JW, John SL: Polymorphisms in a disintegrin and metalloprotease 33 (adam33) predict impaired early-life lung function. Am J Respir Crit Care Med 2005;172:55-60.
20 Kempster S, Phillips WA, Baindur-Hudson S, Thomas RJ, Dow C, Rockman SP: Methylation of exon 2 of p16 is associated with late stage oesophageal cancer. Cancer Lett 2000;150:57-62.
21 Crossen PE, Morrison MJ: Methylation status of the 3rd exon of the c-myc oncogene in b-cell malignancies. Leuk Res 1999;23:251-253.
22 Cakebread JA, Haitchi HM, Holloway JW, Powell RM, Keith T, Davies DE, Holgate ST: The role of adam33 in the pathogenesis of asthma. Springer Semin Immunopathol 2004;25:361-375.
23 Shapiro SD, Owen CA: Adam-33 surfaces as an asthma gene. N Engl J Med 2002;347:936-938.
24 Holgate ST, Yang Y, Haitchi HM, Powell RM, Holloway JW, Yoshisue H, Pang YY, Cakebread J, Davies DE: The genetics of asthma: Adam33 as an example of a susceptibility gene. Proc Am Thorac Soc 2006;3:440-443.
25 Chung KF, Barnes PJ: Cytokines in asthma. Thorax 1999;54:825-857.
26 Sandstrom T, Lundback B: Tobacco smoke: Old foe more important for asthma than commonly appreciated? Eur Respir J 2004;24:720-721.
27 Thomson NC, Chaudhuri R, Livingston E: Asthma and cigarette smoking. Eur Respir J 2004;24:822-833.
28 Breton CV, Byun HM, Wenten M, Pan F, Yang A, Gilliland FD: Prenatal tobacco smoke exposure affects global and gene-specific DNA methylation. Am J Respir Crit Care Med 2009;180:462-467.
29 Wong IH, Lo YM, Yeo W, Lau WY, Johnson PJ: Frequent p15 promoter methylation in tumor and peripheral blood from hepatocellular carcinoma patients. Clin Cancer Res
2000;6:3516-3521.
30 Widschwendter M, Apostolidou S, Raum E, Rothenbacher D, Fiegl H, Menon U, Stegmaier C, Jacobs IJ, Brenner H: Epigenotyping in peripheral blood cell DNA and breast cancer risk: A proof of principle study. PLoS One 2008;3:e2656.
31 Talens RP, Boomsma DI, Tobi EW, Kremer D, Jukema JW, Willemsen G, Putter H, Slagboom PE, Heijmans BT: Variation, patterns, and temporal stability of DNA methylation: Considerations for epigenetic epidemiology. Faseb J 24:3135-3144.
Table 1. Basic characteristics of the asthma and control groups Case group N=50 (mean ± std) Control group N=50 (mean ± std) p-value Age (years) a 46.04±12.50 46.04±12.50 Sex b Male 20 (40%) 20 (40%) Female 30 (60%) 30 (60%) IgE (IU/ml) a 299.87±701.19 74.35±154.39 <0.001* BMI (kg/m2) 24.86±3.94 23.89±3.77 0.212 Waist-hip ratio 0.84±0.09 0.82±0.07 0.123 Lung functions FEV1, % pred 87.84±17.01 93.96±12.94 <0.05 FVC, % pred 89.85±19.40 96.56±13.88 <0.05 Passive smoke No 35 (74.5%) 38 (79.2%) 0.765 Yes 12 (25.5%) 10 (20.8%)
a: the p-value was determined after the data were logarithmically transformed *p<0.05
(UCSC, hg18)
Probe name Gene name Chromoso
me Start End Case group Control group
mea na std mea na std A_17_P17169180 ADAM33 chr20 3653087 3653131 0.74 8 0.038 0.60 1 0.024 A_17_P11026242 ADAM33 chr20 3653142 3653186 1.29 5 0.109 1.28 1 0.096 A_17_P17169181 ADAM33 chr20 3653242 3653286 1.06 9 0.008 1.05 5 0.025 A_17_P17169182 ADAM33 chr20 3653359 3653403 1.01 0 0.031 0.95 7 0.078 A_17_P17169184 ADAM33 chr20 3653523 3653567 0.52 8 0.010 0.62 3 0.081 A_17_P11026243 ADAM33 chr20 3653774 3653827 1.13 5 0.094 0.80 8 0.027 A_17_P11026244 ADAM33 chr20 3653927 3653971 0.90 5 0.038 0.95 7 0.074 A_17_P17169186 ADAM33 chr20 3654094 3654138 1.15 3 0.084 1.16 9 0.086 A_17_P17169187 ADAM33 chr20 3654154 3654198 1.30 7 0.125 1.20 5 0.024 A_17_P17169189 ADAM33 chr20 3654370 3654414 1.53 8 0.136 1.45 3 0.056 A_17_P17169190 ADAM33 chr20 3654449 3654493 2.14 7 0.049 1.80 7 0.366 A_17_P11026245 ADAM33 chr20 3654525 3654569 1.88 8 0.119 1.37 7 0.319 A_17_P17169191 ADAM33 chr20 3654599 3654643 1.49 6 0.037 1.31 4 0.057 A_17_P11026246 ADAM33 chr20 3654705 3654749 1.86 8 0.150 1.53 1 0.200 A_17_P17169192 ADAM33 chr20 3654778 3654822 1.14 6 0.036 1.03 8 0.032 A_17_P11026310 ADAM33 chr20 3662203 3662247 0.82 9 0.133 0.80 0 0.077 A_17_P17169204 ADAM33 chr20 3662297 3662341 1.20 0.104 1.01 0.098
A_17_P17169205 ADAM33 chr20 3662404 3662448 1.12 9 0.021 1.14 0 0.004 A_17_P11026311 ADAM33 chr20 3662648 3662692 1.08 3 0.139 1.06 5 0.080 A_17_P11026312 ADAM33 chr20 3662983 3663027 1.47 9 0.120 1.30 2 0.143 A_17_P17169208 ADAM33 chr20 3663071 3663115 0.85 0 0.012 1.03 8 0.137 A_17_P17169209 ADAM33 chr20 3663188 3663232 0.94 0 0.028 1.01 6 0.117
All of the probe variables were evaluated with an independent t-test.
a. The means were calculated after excluding the maximum and minimum ratios of five repeated experiments.
number 1 2 3 4 5 6 7 8 9 1 0 11 12 13 14 1 5 16 1 7 18 1 9 20 2 1 22 P ro b e n am e U C S C , h g1 8 A _1 7_ P 17 16 91 80 A _1 7_ P 11 02 62 42 A _1 7_ P 17 16 91 81 A _1 7_ P 17 16 91 82 A _1 7_ P 17 16 91 84 A _1 7_ P 11 02 62 43 A _1 7_ P 11 02 62 44 A _1 7_ P 17 16 91 86 A _1 7_ P 17 16 91 87 A _1 7_ P 17 16 91 89 A _1 7_ P 17 16 91 90 A _1 7_ P 11 02 62 45 A _1 7_ P 17 16 91 91 A _1 7_ P 11 02 62 46 A _1 7_ P 17 16 91 92 A _1 7_ P 11 02 63 10 A _1 7_ P 17 16 92 04 A _1 7_ P 17 16 92 05 A _1 7_ P 11 02 63 11 A _1 7_ P 11 02 63 12 A _1 7_ P 17 16 92 08 A _1 7_ P 17 16 92 09 Exon 1 3 13 1 2 12 1 1 9 9 9 8 8 1 CpG sites 9-10 1-5 Promoter promoter
B
Figure 1: The 14 CpG sites, the five selected probes in the microarray and the two primers for methylation-specific PCR in exon 9 of ADAM33
Methylation pattern of cases Methylation pattern of controls
Case s
Control s
Figure 2: The methylation profile of 14 CpG sites in ADAM33.