Genetic polymorphisms of the promoter region of dopamine
D2 receptor and dopamine transporter genes and
alcoholism among four aboriginal groups and Han Chinese
in Taiwan
Wei J. Chen
a, Chia-Hsiang Chen
b, Jielen Huang
c, Yun-Pung P. Hsu
d, See-Voon Seow
c,
Chiao-Chicy Chen
eand Andrew T.A. Cheng
ca
Institute of Epidemiology, College of Public Health, National Taiwan University, Taipei, Taiwan;b Depart-ment of Psychiatry, Tzu-Chi General Hospital, and Institute of Human Genetics, Tzu-Chi Medical College, Hualien City, Taiwan;cInstitute of Biomedical Sciences, Academia Sinica, Taipei, TaiwandDepartment of Life Science, National Tsing Hua University, Hsinchu, Taiwan;eDepartment of Adult Psychiatry, Taipei City Psychiatric Center, Taipei, Taiwan
Correspondence to Professor Andrew T.A. Cheng, Institute of Biomedical Sciences, Academia Sinica, Taipei, Taiwan. E-mail: bmandrew@ccvax.sinica.edu.tw
Received 15 January 2001; accepted 7 August 2001
This study aims to examine the relationship between the functional polymorphism at the promoter region of the
( ) ( )
dopamine D2 receptor DRD2 gene i.e. –141C InsrrrrrDel and variable number of tandem repeat polymorphism at the 3⬘
( ) ( )
untranslated region of the dopamine transporter DAT gene SLC6A3 with alcoholism in a case–control study. The
( ) ( )
cases ns203 were alcohol dependents with withdrawal symptoms, and the controls ns213 were sex- and ethnicity-matched individuals who were screened to exclude those with alcohol problems among four aboriginal groups (Atayal, Ami, Bunun, and Paiwan and Han Chinese in Taiwan. To control for potential confounding factors, we excluded) tobacco abusers from control subjects in part of the analysis and compared the distribution of the genetic polymorphisms in alcoholics with severe medical complications versus those with less severe medical complications. There were no differences in allele and genotype frequencies of these two distinct genetic markers between alcoholics and control subjects in these five different ethnic groups. There was no significant linkage disequilibrium between the –141C
( )
polymorphism and two other DRD2 polymorphisms TaqI A and NcoI . The results remained unchanged when cases were limited to alcoholics with more severe medical complications or when tobacco abusers were excluded from control subjects. The results suggest that both the DRD2 promoter region and the DAT gene do not play a significant role in conferring vulnerability to alcoholism. Psychiatr Genet 11:187–195䊚 2001 Lippincott Williams & Wilkins.
Keywords: alcoholism, genetics, dopamine D2 receptor, dopamine transporter, promoter
INTRODUCTION
Alcoholism is a complex mental disorder that most probably results from the interplay between genetic
Ž
predisposition and environmental factors Cloninger, .
1987; Merikangas, 1990 . In search of genetic sus-ceptibility to alcoholism, the dopaminergic neuro-transmission in the mesolimbic system has been extensively investigated because of its involvement in the reinforcing effects of rewarding behavior ŽKoob, 1992; Nestler et al., 1993; Noble, 1996 . Ani-. mal studies have shown that alcohol can enhance
Ž
the dopamine release in the rat brain Brodie et al.,
.
1990; Weiss et al., 1993 , alcohol preference and
sensitivity are markedly reduced in mice lacking the
Ž . Ž .
dopamine D2 receptor DRD2 Phillips et al., 1998 ,
and blockade of DRD2 disrupts ongoing alcohol Ž
drinking in genetic drinking rat Myers and Robin-.
son, 1999 . The association between the human DRD2 gene and alcoholism was first reported by
Ž .
Blum et al. 1990 . They found an allelic association
of severe alcoholism with the TaqI A1 allele, which was located 10 kb downstream of the end of the 3⬘ untranslated region of the DRD2 gene. However, the results of subsequent association studies have been conflicting, with some replicating and some not
Ž
replicating the finding reviewed by Ferguson and .
Goldberg, 1997; Noble, 1998 .
these findings. First, since there are large variations in DRD2 allele frequencies among population groups ŽKidd et al., 1998 , the association might be due to. confounding by population admixture if cases and control subjects were from different ethnic groups. Nevertheless, combined analyses of independent studies of non-Hispanic Caucasian alcoholics and non-alcoholics have shown the association to be
Ž .
significant Noble, 1998 . However, several family-based association studies employing either the tran-smissionrdisequilibrium test or affected family-based association tests, which are free from popula-tion stratificapopula-tion, failed to find such an associapopula-tion
Ž
between the DRD2 gene and alcoholism
Neis-wanger et al., 1995; Edenberg et al., 1998; Blomqvist
.
et al., 2000 . Second, several studies have reported
that the DRD2 A1 allele is associated with a variety of other substance use disorders, such as nicotine, opioids, cocaine, and polysubstances, and it has been stressed that control subjects should be screened to exclude individuals with these drug problems. Third, since alcoholism is a heterogeneous disorder, the association between the DRD2 A1 allele and the disease might exist only in those with more severe alcoholism. However, there is no clear consensus about the definition of severity. Whether the medi-cal complications commonly seen in more severe cases reflect the severity of alcohol dependence per se or are merely a medical consequence of alcohol
Ž
consumption has been debated Conneally and
. Sparkes, 1998 .
Given that the TaqI A and other nearby polymor-phisms do not affect the protein sequence, it is probable that the association is due to linkage dis-equilibrium with unknown functional polymor-phisms. Recently, a functional polymorphism at the 5⬘ promoter region of the DRD2 gene was identified
Ž .
by Arinami et al. 1997 . The DRD2 promoter
poly-Ž .
morphism is a cytosine insertionrdeletion InsrDel variant at the ᎐141 nucleotide position. An expres-sion assay showed that the C Ins allele has higher
functional activity than the Del allele, indicating
that this polymorphism may regulate the DRD2
Ž .
gene expression in¨i¨o. Ishiguro et al. 1998 found
that the ᎐141C polymorphism was associated with alcoholism in Japanese. Paradoxically, the trend that the frequency of the᎐141C Ins allele was increased in Japanese alcoholic subjects as compared with control subjects was opposite to that expected. Be-sides, Ishiguro et al. found that there is no
signifi-cant linkage disequilibrium between the ᎐141C and TaqI A polymorphisms. Several subsequent studies failed to find an association between the ᎐141C polymorphism and alcoholism, and could not find
linkage disequilibrium between the ᎐141C and the Ž
TaqI A polymorphisms Gelernter et al., 1998;
.
Blomqvist et al., 2000; Noble et al., 2000 . Given the
limited number of studies that have examined this polymorphism in alcoholics, the role of the ᎐141C polymorphism of the DRD2 gene in the susceptibil-ity to alcoholism warranted further investigation.
Another way to test the involvement of dopamin-ergic transmission in alcoholism is to examine ge-netic polymorphisms of the dopaminergic system other than dopamine receptor. The dopamine
Ž .
transporter DAT is located at the presynaptic neu-ron, and is responsible for the re-uptake of do-pamine from the synaptic cleft to terminate the action of dopamine neurotransmission. Abnormal distribution and density of DAT were found in the
Ž .
brain of alcoholics Tiihonen et al., 1995 . Thus,
Ž DAT might have interactions with alcohol
Gold-.
man, 1995 . The variable number of tandem repeat ŽVNTR polymorphisms of the DAT gene locus. Ž
.
symbol SLC6A3 is a 40 bp repeat located at the 3⬘ Ž
untranslated region of the DAT1 gene
Vanden-. Ž .
bergh et al., 1992 . Gelernter et al. 1994 reported
an allelic association between the frequency of the nine-repeat allele of the SLC6A3 and cocaine-induced paranoia. Two subsequent population asso-ciation studies on alcoholism found that either the frequency of the seven-repeat allele of the SLC6A3 was increased in Japanese alcohol-dependent patients with a defective aldehyde dehydrogenase-2
Ž U . Ž .
allele ALDH2 2 Muramatsu and Higuchi, 1995
or the frequency of the nine-repeat allele of the SLC6A3 was increased in German alcoholic subjects
Ž .
with withdrawal symptoms Sander et al., 1997 .
Ž .
Schmidt et al. 1998 further found that the A9 allele
is associated with more severe effects of alcohol withdrawal. However, a recent family-based associa-tion study with a new sample of German alcoholics failed to find a significant association of the
nine-re-Ž
peat allele of the SLC6A3 and alcoholism Franke et
.
al., 1999 . Similar to the case for the DRD2 gene,
the interpretation of these association studies should be cautioned because there are large variations in SLC6A3 polymorphism frequencies among
popula-Ž .
tion groups Kang et al., 1999 . The relationship
between the SLC6A3 polymorphism and alcoholism needs further exploration.
In a prior community-based association study, we examined the relationship between two polymor-phisms of DRD2 and alcoholism in four aboriginal
Ž
groups and Han Chinese in Taiwan Chen et al.,
.
1997a . Features of the study included both cases and control subjects being matched by ethnicity,
Ž
DSM-Ž . III-R American Psychiatric Association, 1987 crite-ria of alcohol dependence and having withdrawal
.
symptoms , control subjects were screened to ex-clude individuals with alcohol problems, and tobacco abusers among controls were excluded from part of the analyses. We found a borderline association
Ž between alcoholism and TaqI A1 in the Ami Ps
.
0.08 and an association between alcoholism and Ž
NcoI N1, located on exon 6 of the DRD2 Sarkar et
. Ž .
al., 1991 , in Han Chinese Ps0.01 . Results of
haplotype analysis further showed that the fre-quency of haplotype A1N1 was higher in alcoholic
Ž subjects than in control subjects for the Ami Ps
. Ž .
0.01 and Han Chinese Ps0.03 .
To explore the nature of these associations fur-ther, in this study we aim to examine the relation-ship between the functional polymorphism at the promoter region of the DRD2 gene and the VNTR polymorphism of the SLC6A3 with alcoholism using the same sample as our previous study. We also examined whether there is linkage disequilibrium between the DRD2 promoter region and the TaqI A and the NcoI polymorphisms. To further control of potential confounders, we excluded tobacco abusers from control subjects in part of the analysis and compared the distribution of the genetic polymor-phisms in alcoholics with severe medical tions versus those with less severe medical complica-tions.
METHODS
Subjects and clinical assessments
The ascertainment of subjects examined in this study Ž
has been described in detail elsewhere Chen et al.,
.
1996, 1997a,b . In brief, subjects who met the
DSM-Ž .
III-R American Psychiatric Association, 1987 cri-teria of alcohol dependence with withdrawal symp-toms were included as cases. The control subjects
Ž were subjects who had no alcohol problems either never drank or only occasionally drank in low
quan-.
tity and were matched on the basis of ethnicity and sex to cases. Older eligible subjects were preferred as matched controls to avoid misclassification in phenotype. Any subjects born from an inter-ethnic marriage were excluded. Related subjects were also eliminated from eligibility. The alcoholic status of the subjects was assessed by a psychiatrist or well-trained research assistants with a semi-structured clinical interview schedule for alcoholism, which in-cludes the alcoholism section of the Schedules for
Ž Clinical Assessment in Neuropsychiatry World
.
Health Organization, 1999 and other relevant items. The reliability of the instrument has been reported
Ž
to be satisfactory Cheng and Chen, 1995; Cheng et
.
al., 2001 .
In addition to psychiatric assessment, information on medical complications and smoking history were also collected. As explained in detail in Chen et al.
Ž1997a , we subgrouped the alcoholics as more se-. vere if they had any one of the following
complica-Ž . Ž
tions: 1 impairment of liver function those without hepatitis B and C virus seropositivity and a combina-tion of an alanine aminotransferase level )35 IUrl . and an aspartate aminotransferase level )35 IUrl ; Ž .2 peripheral neuropathy if subjects reported thatŽ alcohol consumption produced sensations of tingling
. Ž .
or numbness in the feet ; and 3 hallucinations. These criteria were selected because they could be determined either by laboratory tests or by self-reports of high accuracy in a field survey setting. To exclude a possible confounding by tobacco abuse among controls, those who have smoked at least one pack of cigarettes per day for at least 1 year were deleted from part of the analysis.
The aboriginal subjects were ascertained from ei-ther a longitudinal community survey among four
Ž .
aboriginal groups Atayal, Ami, Bunun, and Paiwan ŽCheng & Chen, 1995; Chen & Cheng, 1997 , or.
Ž
referred to us via local health workers Chen et al.,
.
1997b . Almost all cases were identified from the community and many of them have been treated for various complications of alcohol drinking, although few of them had ever been treated for alcoholism
per se due to poor accessibility of psychiatric
ser-vices. The Han alcoholic subjects were recruited Ž
from both community Chinshan and Sanchi
Town-. Ž
ship and clinical settings Taipei City Psychiatric .
Center and Chinshan Health Station in northern Taiwan, while the Han control subjects were
re-Ž
cruited from both the community Chinshan and .
Sanchi Township and the Health Screening Ward at Ž
National Taiwan University Hospital Chen et al.,
. 1996 .
Ž
In total, 203 alcoholic subjects 168 men and 35
. Ž
women and 213 control subjects 178 men and 35 .
women were recruited for this study. The mean age for each group ranged from 41.2 to 51.1 years for cases and from 54.2 to 61.0 years for control sub-jects. However, because of genotyping failure or shortage of DNA material, the actual number of subjects varied for the analysis of the DRD2 pro-moter and the DAT1 VNTR polymorphisms.
Polymerase chain reaction-based genotyping
Genomic DNA samples extracted from blood were used as templates for polymerase chain reaction
ŽPCR amplification. The genotyping of the func-. tional polymorphism at the promoter region of the
Ž .
DRD2 gene i.e. ᎐141C InsrDel was based on the
Ž .
method described by Arinami et al. 1997 with
modification. In brief, PCR was carried out in a volume of 25l reaction mixture containing sense
primer 5
⬘-ACTGGCGAGCAGACGGTGAG-GACCC-3⬘ and antisense primer
5⬘-TGCGCGCGT-GAGGCTGCCGGTTCGG-3⬘. After an initial
Ž denaturation at 95⬚C for 5 min, amplification 30
.
cycles was performed with denaturation at 95⬚C for 1 min, annealing at 68⬚C for 1 min, and extension at 72⬚C for 1 min. The PCR amplification yielded a 304 Ž bp DNA fragment. After amplification, an aliquot 5
. Ž
l of PCR product was incubated with BstN1 New .
England Biolabs, Beverly, MA, USA at 60⬚C
overnight. The digested PCR products were sub-jected to electrophoresis in 3% agarose gel, stained with ethidium bromide, and visualized under UV
Ž .
light. The Ins allele a cytosine insertion remained
Ž uncut with the size of 304 bp, while the Del allele a
.
cytosine deletion yielded two DNA fragments of 160 and 144 bp.
The PCR-based genotyping of a 40-bp repeat poly-morphism at the 3⬘ untranslated region of the hu-man DAT gene was essentially as described by Sano
Ž .
et al. 1993 . In brief, PCR amplification was carried
out in a 25 l reaction mixture, containing sense
primer 5
⬘-TGTGGTGTAGGGAACGGCCTGAGA-3⬘ and antisense primer
5⬘-TGTTGGTCTGCAG-GCTGCCTGCAT-3⬘. After initial denaturation at 95⬚C for 3 min, 30 cycles of PCR were carried out with the following conditions: denaturation at 95⬚C for 1 min, annealing at 60⬚C 1 for min, and exten-sion at 72⬚C for 1 min. The PCR products were separated in 2% agarose gel by electrophoresis, stained with ethidium bromide and visualized under UV light with a marker standard on the same gel.
The genotypings of TaqI A and NcoI polymor-phism have been described in our previous report ŽChen et al., 1997a ..
Statistical analysis
The differences in allele frequency or genotype dis-tribution between cases and controls were examined
Ž .
by Fisher’s exact test two-tailed , and the Hardy᎐Weinberg equilibrium was tested by the 2
test with Yates’ continuity correction. Frequencies of two-locus haplotype were estimated using Hill’s Ž1974 iteration algorithm. For convenience, we de-. noted the frequency of the rarer allele at the first locus p and that at the second locus q: pFqF0.5.
The maximum-likelihood estimate of linkage
dis-equilibrium statistic D was computed as Dsf ᎐11
pq, where f11 is the frequency of the haplotype with the rare allele at both loci. Let n denote number of
subjects, the statistic of the test for Ds0 was
ŽD 2 n. r p 1 ᎐ p q 1 ᎐ q , which has the dis-w Ž . Ž .x 2 tribution asymptotically with one degree of freedom Ždf . The maximum possible value of. D is p 1Ž ᎐ q ,. denoted as Dma x, and the percentage of Dmax
Ž .
achieved by D is D⬘sDrDma x Lewontin, 1988 . All statistical tests did not adjust for multiple compar-isons since the association between the DRD2 gene and alcoholism has been reported previously. P
-0.05 was considered significant, and P-0.1 as
borderline. RESULTS
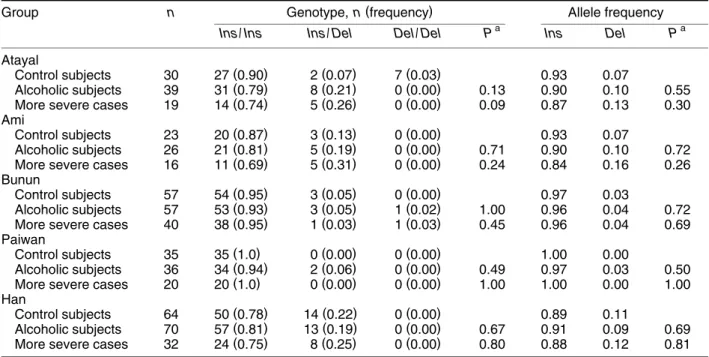
The genotype distribution of the ᎐141C InsrDel polymorphism of the DRD2 gene did not deviate from Hardy᎐Weinberg equilibrium in the four abo-riginal groups and Han Chinese. Table 1 summarizes the distribution of the genotype and allele frequency of this polymorphism of the DRD2 gene. There were no significant differences in the genotype and allele frequency of this DRD2 polymorphism between al-coholic subjects and control subjects in the five ethnic groups from Taiwan. If we limited the al-coholic cases to those with more severe complica-tions, the difference in genotype distribution between alcoholic subjects and control subjects became
Ž .
borderline for the Atayal Ps0.09 .
If control subjects who have had tobacco abuse Žone Atayal, one Ami, seven Bunun, two Paiwan,
.
and 15 Han are excluded from the analysis, the Ž
frequencies of the three genotypes InsrIns,
.
InsrDel, and DelrDel were respectively as follows:
0.90, 0.07, and 0.03 for the Atayal; 0.86, 0.14, and 0.00 for the Ami; 0.96, 0.04, and 0.00 for the Bunun; 1.00, 0.00, and 0.00 for the Paiwan; and 0.82, 0.18, and 0.00 for the Han. These were similar to those of the original control subjects. There were no differ-ences in the genotype distribution of the ᎐141C
InsrDel polymorphism of the DRD2 gene between
the tobacco-abuse-free control subjects and al-coholic subjects for all five groups. If only more severe cases were included for the comparison, the difference in genotype distribution between al-coholic subjects and the tobacco-abuse-free control
Ž subjects remained borderline for the Atayal Ps
. 0.09 .
Since the ᎐141C polymorphism is located on the same DRD2 region with TaqI A and NcoI, we further examined whether there was linkage disequi-librium between the ᎐141C InsrDel polymorphism
TABLE 1. Genotype and allele frequencies of the᎐141C Ins/Del polymorphism of the DRD2 gene among Taiwanese aborigines and Han Chinese
( )
Group n Genotype, n frequency Allele frequency
a a
Ins / Ins Ins / Del Del / Del P Ins Del P
Atayal ( ) ( ) ( ) Control subjects 30 27 0.90 2 0.07 7 0.03 0.93 0.07 ( ) ( ) ( ) Alcoholic subjects 39 31 0.79 8 0.21 0 0.00 0.13 0.90 0.10 0.55 ( ) ( ) ( )
More severe cases 19 14 0.74 5 0.26 0 0.00 0.09 0.87 0.13 0.30
Ami ( ) ( ) ( ) Control subjects 23 20 0.87 3 0.13 0 0.00 0.93 0.07 ( ) ( ) ( ) Alcoholic subjects 26 21 0.81 5 0.19 0 0.00 0.71 0.90 0.10 0.72 ( ) ( ) ( )
More severe cases 16 11 0.69 5 0.31 0 0.00 0.24 0.84 0.16 0.26
Bunun ( ) ( ) ( ) Control subjects 57 54 0.95 3 0.05 0 0.00 0.97 0.03 ( ) ( ) ( ) Alcoholic subjects 57 53 0.93 3 0.05 1 0.02 1.00 0.96 0.04 0.72 ( ) ( ) ( )
More severe cases 40 38 0.95 1 0.03 1 0.03 0.45 0.96 0.04 0.69
Paiwan ( ) ( ) ( ) Control subjects 35 35 1.0 0 0.00 0 0.00 1.00 0.00 ( ) ( ) ( ) Alcoholic subjects 36 34 0.94 2 0.06 0 0.00 0.49 0.97 0.03 0.50 ( ) ( ) ( )
More severe cases 20 20 1.0 0 0.00 0 0.00 1.00 1.00 0.00 1.00
Han ( ) ( ) ( ) Control subjects 64 50 0.78 14 0.22 0 0.00 0.89 0.11 ( ) ( ) ( ) Alcoholic subjects 70 57 0.81 13 0.19 0 0.00 0.67 0.91 0.09 0.69 ( ) ( ) ( )
More severe cases 32 24 0.75 8 0.25 0 0.00 0.80 0.88 0.12 0.81
a ( )
P-value of Fisher’s exact test two-tailed comparing control subjects with alcoholic subjects or more severe cases.
and the latter two. Because the frequency of DelrDel
Ž .
was very low in all five groups range, 0᎐0.03 , the frequencies of the haplotypes involving allele Del
were also low. Unlike the significant linkage disequi-librium between the TaqI A and NcoI in all five
Ž
ethnic groups we reported previously D⬘ s
. Ž .
0.65᎐1.00 Chen et al., 1997a , neither the D⬘ value between the ᎐141C and TaqI A nor that between the ᎐141C and NcoI reached statistical significance, except for one group. The D⬘ value between the
᎐141C and TaqI A in controls and cases for each group were 1.00 and 0.58 for the Atayal, 0.88 and 1.00 for the Ami, 1.00 and 1.00 for the Bunun, 0.00 and 1.00 for the Paiwan, and 0.42 and 0.58 for the Han, respectively. Only the D⬘ value for the Bunun
Ž 2
cases reached statistical significance s 5.75, .
dfs1, P-0.0001 . The D⬘ value between the ᎐141C and NcoI in controls and cases for each group were 1.00 and 0.51 for the Atayal, 1.00 and 1.00 for the Ami, 0.16 and 1.00 for the Bunun, 0.00 and 1.00 for the Paiwan, and 0.46 and 0.65 for the Han, respec-tively. Again, only the D⬘ value for the Bunun cases
Ž 2
reached statistical significance s7.15, dfs1, .
P-0.0001 .
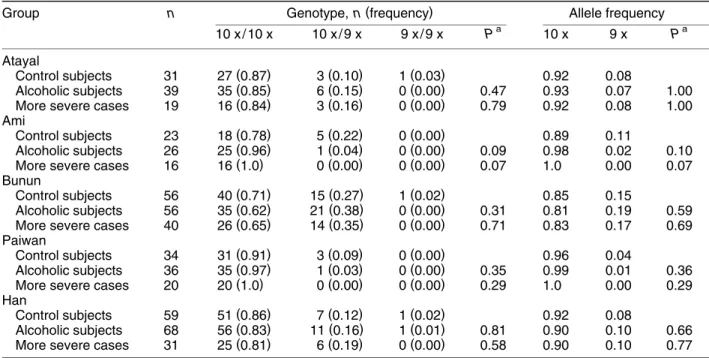
The genotype and allele frequencies of VNTR polymorphism at the 3⬘ untranslated region of SLC6A3 among four aboriginal groups and Han Chinese were summarized in Table 2. In our sam-ples, three alleles were detected, including 490 bp
Ž11 repeats , 450 bp 10 repeats , and 410 bp nine. Ž . Ž .
repeats . The 11-repeat allele was very rare in our subjects; we did not detect this allele in the Atayal, Ami, and Bunun. Only one out of 142 alleles in the Paiwan and six out of 256 alleles in the Han Chinese were found to be 11-repeat alleles. Hence, these individuals with 11 repeats were excluded from com-parison. The genotype distribution did not deviate from Hardy᎐Weinberg equilibrium in both alcoholic subjects and control subjects in these five ethnic groups. There was a borderline difference between alcoholic subjects and control subjects in both the genotype distribution and allele frequency in the
Ž .
Ami Ps0.09 . For the remaining ethnic groups,
there were no differences in genotype and allele frequencies between alcoholic groups and normal control subjects. If we limited the alcoholic cases to those with more severe complications, the difference in the genotype distribution and allele frequency between alcoholic subjects and control subjects re-mained borderline for the Ami, although the P
Ž .
value decreased slightly from 0.09 to 0.07 . Ž
The frequencies of the three genotypes 10 xr10 .
x, 10 xr9 x, and 9 xr9 x among the tobacco-abuse-free control subjects were respectively as follows: 0.90, 0.07, and 0.03 for the Atayal; 0.82, 0.18, and 0.00 for the Ami; 0.69, 0.29, and 0.02 for the Bunun; 0.90, 0.10, and 0.00 for the Paiwan; and 0.87, 0.11, and 0.02 for the Han. Again, these were similar to
TABLE 2. Genotype and allele frequencies of the VNTR polymorphism at the 3⬘ untranslated region of the DAT1 gene among Taiwanese aborigines and Han Chinese
( )
Group n Genotype, n frequency Allele frequency
a a 10 x / 10 x 10 x / 9 x 9 x / 9 x P 10 x 9 x P Atayal ( ) ( ) ( ) Control subjects 31 27 0.87 3 0.10 1 0.03 0.92 0.08 ( ) ( ) ( ) Alcoholic subjects 39 35 0.85 6 0.15 0 0.00 0.47 0.93 0.07 1.00 ( ) ( ) ( )
More severe cases 19 16 0.84 3 0.16 0 0.00 0.79 0.92 0.08 1.00
Ami ( ) ( ) ( ) Control subjects 23 18 0.78 5 0.22 0 0.00 0.89 0.11 ( ) ( ) ( ) Alcoholic subjects 26 25 0.96 1 0.04 0 0.00 0.09 0.98 0.02 0.10 ( ) ( ) ( )
More severe cases 16 16 1.0 0 0.00 0 0.00 0.07 1.0 0.00 0.07
Bunun ( ) ( ) ( ) Control subjects 56 40 0.71 15 0.27 1 0.02 0.85 0.15 ( ) ( ) ( ) Alcoholic subjects 56 35 0.62 21 0.38 0 0.00 0.31 0.81 0.19 0.59 ( ) ( ) ( )
More severe cases 40 26 0.65 14 0.35 0 0.00 0.71 0.83 0.17 0.69
Paiwan ( ) ( ) ( ) Control subjects 34 31 0.91 3 0.09 0 0.00 0.96 0.04 ( ) ( ) ( ) Alcoholic subjects 36 35 0.97 1 0.03 0 0.00 0.35 0.99 0.01 0.36 ( ) ( ) ( )
More severe cases 20 20 1.0 0 0.00 0 0.00 0.29 1.0 0.00 0.29
Han ( ) ( ) ( ) Control subjects 59 51 0.86 7 0.12 1 0.02 0.92 0.08 ( ) ( ) ( ) Alcoholic subjects 68 56 0.83 11 0.16 1 0.01 0.81 0.90 0.10 0.66 ( ) ( ) ( )
More severe cases 31 25 0.81 6 0.19 0 0.00 0.58 0.90 0.10 0.77
a ( )
P-value of Fisher’s exact test two-tailed comparing control subjects with alcoholic subjects or more severe cases.
those of the original control subjects. There were no differences in the genotype distribution of the
SLC6A3 VNTR polymorphism between the
tobacco-abuse-free control subjects and alcoholic subjects, either all cases or more severe ones only, for all five groups.
DISCUSSION
The continuing controversy over the association between the DRD2 gene and alcoholism is puzzling. On the one hand, there continue to be new results supporting this association. The quantitative trait locus mapping in animal studies consistently demon-strates that the DRD2 gene is responsible for
al-Ž
cohol drinking in rats Philips et al., 1998; Myers and
.
Robinson, 1999 . Although one earlier meta-analysis of human population association studies indicated that the associations between the TaqI A polymor-phism and alcoholism of early studies were due to
Ž
abnormally low control allele frequencies Gelernter .
et al., 1993 , several later meta-analyses of studies
among non-Hispanic Caucasians did find a
signifi-Ž .
cant association between the two Noble, 1998 . On the other hand, the magnitudes of association between alcoholism and haplotypes composing of TaqI A and several nearby polymorphisms that are closer to the DRD2 gene per se were less than TaqI
Ž .
A alone Noble et al., 2000 . An earlier study
searched for functional polymorphisms in the region Ž
of exon 2 to exon 8 in the DRD2 region Gejman et
.
al., 1994 . Although some polymorphisms were found
in the exon regions, these were rare and were not associated with alcoholism.
Under this circumstance, the functional polymor-phism of the promoter region of the DRD2 gene is of special interest because it has relatively high allele prevalence of the mutant allele. The fre-quency of the Del allele in the control subjects
ranged from 0.0 to 0.11 in the control subjects in the present study, which is close to that in Americans of
Ž .
European origin 0.11 by Gelernter et al., 1998 but
lower than those in other populations, such as Ž
Japanese 0.23 by Ishiguro et al., 1998, or 0.21 by
. Ž
Gelernter et al., 1998 and African-Americans 0.39
.
by Gelernter et al., 1998 . Intriguingly, the
frequen-cies of the Del allele in alcoholics of different
popu-lations are closer to one another: 0.03᎐0.10 in the
Ž .
present study, 0.14 Ishiguro et al., 1998 in the
Ž .
Japanese, 0.11 in Germans Blomqvist et al., 2000 ,
Ž
and 0.10 in non-Hispanic Caucasians Noble et al.,
. 2000 .
In terms of the potential association between the
InsrDel and alcoholism, however, we did not find a
positive association in our samples. This is consistent Ž
with two previous studies Blomqvist et al., 2000;
.
Noble et al., 2000 . The only study that reported an
al-coholic subjects was in a subpopulation of Japanese
Ž U . Ž
alcoholic subjects those with ALDH2 2 Ishiguro .
et al., 1998 . However, the paradoxical direction of
Ž
the association higher frequency of the Ins allele
. rather than the Del allele in alcoholic subjects
renders the finding difficult to explain. Moreover, there is no significant linkage disequilibrium between the ᎐141C InsrDel polymorphism and the TaqI A and NcoI polymorphisms of the DRD2 in our sam-ples. This is indeed consistent with all three previous
Ž
studies Gelernter et al., 1998; Ishiguro et al., 1998;
.
Blomqvist et al., 2000; Noble et al., 2000 . Taken
together, the evidence so far does not support this promoter polymorphism of the DRD2 gene playing a significant role in increasing vulnerability to al-coholism.
In addition, we examined the possible involvement of the DAT gene in alcoholism in our samples. Although positive allelic association of the VNTR polymorphism of SLC6A3 and alcoholism has been
Ž
reported Muramatsu and Higuchi, 1995; Sander et
.
al., 1997 , the physiological function of the VNTR
polymorphism of SLC6A3 remains unknown. Even in these positive studies, the allele that was found to have increased frequency in these two studies was
Ž
different the seven-repeat allele in the Japanese alcoholic subjects and the nine-repeat allele in the
.
German alcoholic subjects . However, no positive association with alcoholism was detected in this study. Our results are consistent with those of two
Ž
family-based association studies Parsian and Zhang, .
1997; Franke et al., 1999 . A recent comprehensive
search for variations in each of 15 SLC6A3 exons and several introns failed to identify any common protein coding DAT sequence variant in 150 unre-lated individuals free of neuropsychiatric disease and 228 patients of various psychiatric disorders Žincluding 64 alcohol dependents. ŽVandenbergh et
.
al., 2000 . The result indicates that there is
substan-tial evolutionary conservation of the DAT protein sequence. Given the conservation in the coding re-gion of the DAT gene and the failure of several
Ž .
association studies including this one to find an association between the VNTR polymorphism of the DAT gene and alcoholism, the possibility that the DAT gene is involved in the susceptibility to al-coholism may lie in its level-of-expression variation. Nevertheless, the SLC6A3 VNTR is a non-func-tional marker located at one end of the gene and little is known about linkage disequilibrium across this gene. Therefore, the lack of association between the VNTR polymorphism and alcoholism cannot exclude other variation in the gene if it is not in linkage disequilibrium with this VNTR marker.
The conflicting results as seen in many association studies may be attributed to several factors. One is the population admixture due to ethnic heterogen-eity in the subjects studied. In this, we use the genetically isolated ethnic groups and chose the appropriate control subjects matched with sex and gender. Moreover, the control subjects were screened to exclude those with alcohol drinking problems. When tobacco abusers were excluded from part of our analyses, the results remained unchanged. Hence, the possibility of false-negative finding in
either the DRD2 ᎐141C polymorphism or the
SLC6A3 VNTR polymorphism due to population stratification or contaminated control subjects is low in this study. Another issue pertaining to the associ-ation studies in alcoholism is whether the alcoholic subjects are of sufficient severity. In this study, we consider all of our alcoholic patients as severe be-cause they all had withdrawal symptoms in addition to satisfying the DSM-III-R criteria of alcohol de-pendence. Furthermore, if we subgrouped patients into less severe and more severe, and compared the more severe group with control subjects, the results of lack of significant associations between the two genetic polymorphisms and alcoholism remained un-changed.
Two limitations of this study should be noted. First, the sample size of each ethnic group was relatively small. Second, the level of heterozygosity
for both the DRD2 ᎐141C InsrDel and SLC6A3
polymorphisms was low in our samples. Thus, we cannot exclude the possibility of insufficient power to detect the association due to the combination of these two factors.
Acknowledgements
This work was supported by grants from the National
Ž .
Science Council, Taipei NSC 89-2320-B001-073 . The au-thors acknowledge the excellent performance of research assistants J.C. Chang, J.Y. Kao and C.L. Cheng in case identification.
REFERENCES
Ž .
American Psychiatric Association 1987 . Diagnostic and statistical manual of mental disorders, revised. 3rd edn. Washington, DC: American Psychiatric Association.
Ž .
Arinami T, Gao M, Hamaguchi H, Toru M 1997 . A functional polymorphism in the promoter region of the dopamine D2 receptor gene is associated with schizophrenia. Hum Mol Genet 6:577᎐582.
Ž .
Blomqvist O, Gelernter J, Kranzler HR 2000 . Family-based study of DRD2 alleles in alcohol and drug
depen-( )
dence. Am J Med Genet Neuropsychiatr Genet 96:659᎐664.
Blum K, Noble EP, Sheridan PJ, Montgomery A, Ritchie
Ž .
human dopamine D2 receptor gene in alcoholism.J Am Med Assoc 263:2055᎐2060.
Ž .
Brodie MS, Shefner SA, Dunwiddie TV 1990 . Ethanol increases the firing rate of dopamine neurons of the rat. Brain Res 508:65᎐69.
Ž .
Chen WJ, Cheng ATA 1997 . Incidence of first onset alcoholism among Taiwanese aborigines. Psychol Med 27:1363᎐1371.
Chen WJ, Loh EW, Hsu Y-PP, Chen C-C, Yu J-M, Cheng
Ž .
ATA 1996 . Alcohol-metabolizing genes and al-coholism among Taiwanese Han Men: independent ef-fect of ADH2, ADH3, and ALDH2. Br J Psychiatry 168:762᎐767.
Chen WJ, Lu ML, Hsu Y-PP, Chen C-C, Yu J-M, Cheng
Ž .
ATA 1997a . Dopamine D2 receptor gene and alcoholism among four aboriginal groups and Han in
( )
Taiwan.Am J Med Genet Neuropsychiatr Genet 74:129᎐136.
Ž .
Chen WJ, Loh EW, Hsu Y-PP, Cheng ATA 1997b . Alcohol dehydrogenase and aldehyde dehydrogenase genotypes and alcoholism among Taiwanese aborigines. Biol Psychiatry 41:703᎐709.
Ž .
Cheng ATA, Chen WJ 1995 . Alcoholism among four aboriginal groups in Taiwan: high prevalences and their implications. Alcohol Clin Exp Res 19:81᎐91.
Cheng ATA, Tien AY, Chang CJ, Brugha TS, Cooper JE,
Ž .
Lee CS,et al. 2001 . Cross-cultural implementation of a Chinese version of the Schedules for Clinical
Assess-Ž .
ment in Neuropsychiatry SCAN in Taiwan. Br J Psy-chiatry 178:567᎐572.
Ž .
Cloninger CR 1987 . Neurogenetic adaptive mechanisms in alcoholism.Science 236:410᎐416.
Ž .
Conneally PM, Sparkes RS 1998 . General discussion. Alcohol 16:85᎐91.
Edenberg HJ, Foroud T, Koller DL, Goate A, Rice J,
Ž .
Eerdewegh PV,et al. 1998 . A family-based analysis of
Ž .
the association of the dopamine D2 receptor DRD2 with alcoholism. Alcohol Clin Exp Res 22:505᎐512.
Ž .
Ferguson RA, Goldberg DM 1997 . Genetic markers of alcohol abuse.Clin Chim Acta 257:199᎐250.
Franke P, Schwab SG, Knapp M, Gansicke M, Delmo C,
Ž .
Zill P, et al. 1999 . DAT1 gene polymorphism in al-coholism: a family-based association study.Biol Psychia-try 45:652᎐654.
Gejman PV, Ram A, Gelernter J, Friedman E, Cao Q,
Ž .
Pickar D, et al. 1994 . No structural mutation in the dopamine D2 receptor gene in alcoholism or schizophrenia. J Am Med Assoc 271:204᎐208.
Ž .
Gelernter J, Goldman D, Risch N 1993 . The A1 allele at the D2 dopamine receptor gene and alcoholism: a re-appraisal. J Am Med Assoc 269:1673᎐1677.
Ž .
Gelernter J, Kranzler HR, Satel AL, Rao PA 1994 . Genetic association between dopamine transporter protein alleles and cocaine-induced paranoia. Neuropsychopharmacology 11:195᎐200.
Gelernter J, Kranzler H, Cubells JF, Ichinose H, Nagatsu
Ž .
T 1998 . DRD2 allele frequencies and linkage disequi-libria, including the᎐141C InsrDel promoter polymor-phism, in European᎐American, African᎐American, and Japanese subjects.Genomics 51:21᎐26.
Ž .
Goldman D 1995 . Dopamine transporter, alcoholism and other diseases. Nat Med 1:624᎐625.
Ž .
Hill WG 1974 . Estimation of linkage disequilibrium in randomly mating populations.Heredity 33:229᎐239. Ishiguro H, Arinami T, Saito T, Akazawa S, Enomoto M,
Ž .
Mitushio H,et al. 1998 . Association study between the
᎐141C InsrDel and TaqI A polymorphisms of the dopamine D2 receptor gene and alcoholism. Alcohol Clin Exp Res 22:845᎐848.
Ž .
Kang AM, Palmatier MA, Kidd KK 1999 . Global varia-tion of a 40-bp VNTR in the 3⬘-untranslated region of
Ž .
the dopamine transporter gene SLC6A3 . Biol Psychia-try 46:151᎐160.
Kidd KK, Morar B, Castiglione CM, Zhao H, Pakstis AJ,
Ž .
Speed WC, et al. 1998 . A global survey of haplotype frequencies and linkage disequilibrium at the DRD2 locus. Hum Genet 103:211᎐227.
Ž .
Koob GF 1992 . Drugs of abuse: anatomy, pharmacology and function of reward pathways.Trends Pharmacol Sci 13:177᎐184.
Ž .
Lewontin RC 1988 . On measures of gametic disequilib-rium.Genetics 120:849᎐852.
Ž .
Merikangas KR 1990 . The genetic epidemiology of al-coholism. Psychol Med 20:11᎐22.
Ž .
Muramatsu T, Higuchi S 1995 . Dopamine transporter gene polymorphism and alcoholism. Biochem Biophys Res Commun 211:28᎐32.
Ž .
Myers RD, Robinson DE 1999 . Mmu and D2 receptor antisense oligonucleotides injected in nucleus accum-bens suppress high alcohol intake in genetic drinking HEP rats. Alcohol 18:225᎐233.
Ž .
Neiswanger K, Hill SY, Kaplan BB 1995 . Association and linkage studies of the TaqI A1 allele at the do-pamine D2 receptor gene in samples of female and
( )
male alcoholics. Am J Med Genet Neuropsychiatr Genet 60:267᎐271.
Ž .
Nestler EJ, Hope BT, Widnell KL 1993 . Drug addiction: a model for the molecular basis of neural plasticity. Neuron 11:995᎐1006.
Ž .
Noble EP 1996 . Alcoholism and the dopaminergic sys-tem: a review. Addict Biol 1:333᎐348.
Ž .
Noble EP 1998 . The D2 dopamine receptor gene: a review of association studies in alcoholism and pheno-types. Alcohol 16:33᎐45.
Ž .
Noble EP, Zhang X, Ritchie TL, Sparkes RS 2000 . Haplotypes at the DRD2 locus and severe alcoholism.
( )
Am J Med Genet Neuropsychiatr Genet 96:622᎐631.
Ž .
Parsian A, Zhang ZH 1997 . Human dopamine
Ž .
transporter gene polymorphism VNTR and
al-( )
coholism. Am J Med Genet Neuropsychiatr Genet 74:480᎐482.
Phillips TJ, Brown KJ, Burkhart-Kasch S, Wenger CD,
Ž .
Kelly MA, Rubinstein M, et al. 1998 . Alcohol prefer-ence and sensitivity are markedly reduced in mice lack-ing dopamine D2 receptors. Nat Neurosci 1:610᎐615. Sander T, Harms H, Podschus J, Finckh U, Nickel B,
Ž .
Rolfs A, et al. 1997 . Allelic association of a dopamine transporter gene polymorphism in alcohol dependence with withdrawal seizures or delirium. Biol Psychiatry 41:299᎐304.
Ž .
Sano A, Kondoh K, Kakimoto Y, Kondo I 1993 . A 40-nucleotide repeat polymorphism in the human do-pamine transporter gene. Hum Genet 91:405᎐406. Sarkar G, Kapelner S, Grandy DK, Marchionni M, Civelli
Ž .
O, Sobell JL, et al. 1991 . Direct sequencing of the
Ž .
dopamine D2 receptor DRD2 in schizophrenics re-veals three polymorphisms but no structural change in the receptor.Genomics 11:8᎐14.
Schmidt LG, Harms H, Kuhn S, Rommelspacher H, Sander
Ž .
T 1998 . Modification of alcohol withdrawal by the A9 allele of the dopamine transporter gene. Am J Psychia-try 155:474᎐478.
Tiihonen J, Kuikka J, Bergstrom K, Hakola P, Karhu J,
Ž .
do-pamine re-uptake site densities in habitually violent and non-violent alcoholics. Nat Med 1:654᎐657.
Vandenbergh DJ, Persico AM, Hawkins AL, Griffin CA,
Ž .
Li X, Jabs EW, Uhl GR 1992 . Human dopamine
Ž .
transporter gene DAT1 maps to chromosome 5p15.3 and display a VNTR.Genomics 14:1104᎐1106.
Vandenbergh DJ, Thompson MD, Cook EH, Bendahhou
Ž .
E, Nguyen T, Krasowski MD, et al. 2000 . Human dopamine transporter gene: coding region conservation among normal, Tourette’s disorder, alcohol dependence
and attention-deficit hyperactivity disorder populations. Mol Psychiatry 5:283᎐292.
Ž .
Weiss F, Lorang MT, Bloom FE, Koob GF 1993 . Oral alcohol self-administration stimulates dopamine release in the rat nucleus accumbens: genetic and motivational determinants. J Pharmacol Exp Therap 267:250᎐258.
Ž .
World Health Organization 1999 . SCAN 2.1: schedules for clinical assessment in neuropsychiatry. Cambridge: Cambridge University Press.