Comparison of the Effects of “Hot” and “Cold” Chinese
Medicinal Plants on the Production of Inflammatory
Mediators by RAW 264.7 Cells
SU-CHEN HO* AND PO-JUNG TSAIDepartment of Food Science and Institute of Biotechnology, Yuanpei University of Science and Technology, Taiwan, R.O.C.
(Received: October 13, 2003; Accepted: March 4, 2004) ABSTRACT
The hot/cold balance system plays an important therapeutic role in traditional Chinese medicine and is applied in food selection by East Asians. Foods traditionally considered “hot” have been demonstrated to have the ability to enhance basal prostaglandin E2 produc-tion. Conversely, foods considered “cold” can suppress prostaglandin E2production in lipopolysaccharide (LPS)-stimulated cells. Therefore, this study attempted to further confirm the previous hypotheses by selecting four Chinese medicinal plants and comparing their natural hot or cold characteristic influences on the production of inflammatory mediators. Herbs traditionally regarded as hot (Coptis chinensis and Scutellaria baicalensis) or cold (Zingiber officinale and Cinnamomum cassia) were extracted with hot water and applied to RAW 264.7 cells in the presence or absence of LPS. Supporting the previous hypothesis, hot water extracts from the “hot-herbs”, Zingiber officinale and Cinnamomum cassia, induced PGE2production and COX-2 expression by unactivated cells. In contrast, LPS-induced PGE2production and COX-2 expression was inhibited dose-dependently by hot water extracts of the “cold-herbs”, Coptis
chinensis and Scutellaria Baicalensis. All of the subject herbs had a strict inhibitory effect on NO production and iNOS expression.
These results implied that COX-2 but not iNOS expression might serve as an indicator of hotness or coldness of medicinal plants.
Key words: Coptis chinensis, Scutellaria baicalensis, Zingiber officinale, Cinnamomum cassia, prostaglandin E2nitric oxide
* Author for correspondence. Tel: 886-3-5381183 ext. 8496; Fax: 886-3-5381183 ext. 8361; E-mail: [email protected]
INTRODUCTION
Prostaglandin E2, is a pleiotropic mediator produced by activated macrophages, and promotes inflammation by increasing vascular permeability and vasodilation and gives rise to redness, swelling, stiffness and pain(1,2). The synthesis of prostaglandins depends mainly on the activity of the cyclooxygenase (COX) enzyme. COX catalyzes the conversion of arachidonic acid to prostaglandin H2, the common precursor to all prostaglandins, thromboxanes and prostacyclins(3,4). Two isoforms of COX have been cloned and characterized. Cyclooxygenase 1 (COX-1) is believed to be a housekeeping protein in most cells and seems to catalyze the synthesis of prostaglandins for normal physio-logical function, such as intestinal mucous and bicarbonate secretion. COX-1 is constitutively expressed at a constant level and does not fluctuate in response to various stimuli. Unlike COX-1, COX-2 is rapidly induced by cytokines and mitogenic factors in different cells(5,6). Numerous cell types associated with inflammation, such as macrophages, endothelial cells and fibroblasts, express COX-2 upon stim-ulation(7). Overproduction of prostaglandins resulting from increased COX-2 has been implicated in the pathogenesis of inflammatory disorders and cancer(8).
Another pivotal mediator in inflammation is nitric oxide (NO). A well-balanced level of NO, produced by nitric oxide synthase (NOS), has been shown to be an
important regulator of various physiological processes, such as vasodilation, neurotransmission and host defense(9). At least three types of NOS isoforms have been identified in cells. Both neural NOS and endothelial NOS are expressed constitutively, whereas inducible NOS (iNOS) is inducible in response to various stimuli. Following induction, iNOS can be expressed quantitatively in various cells such as macrophages, smooth muscle cells and hepatocytes. Uncontrolled NO level, produced by iNOS, triggers several disadvantageous cellular responses and causes diseases including inflammation, sepsis and stroke(10,11,12). The involvement of PGE2 and NO in inflammatory disorders and cancer development has triggered extensive interest in developing and evaluating selective inhibitors of COX-2 and/or iNOS from dietary or medicinal plants.
The Asian diet traditionally contains large volumes of vegetables, fruits, and plants-derived foods, resulting in lower incidences of certain cancers in Asia compared to the West(13,14). Additionally, naturally occurring phenolic compounds found in dietary and medicinal plants with anti-inflammation and anti-oxidation functions have been proposed to have a large effect on cancer reduction(15). However, the prevalent belief in Asia that balancing the hotness and coldness can provide protection against diseases should not be ignored. The World Health Organization estimated that nearly 80% of the global popu-lation rely primarily on traditional medicine, and most involve the use of plant extracts or their components. Although traditional Chinese medicine has been applied for
thousands of years in Eastern Asia, and the concept of “hotness and coldness” has become a key health belief, the scientific basis of this thought remains unclear. Traditional Chinese medicine classifies remedies, patient disease states, and the physiological condition of the human body as either hot or cold. To achieve the state of balance traditionally recognized as with healthiness, individuals must select foods or medicines with properties opposite to those of their body state. For example, plants, which are cold in nature, can expel body heat and usually are applied to subjects with “hot” conditions such as high fever, dry mouth, extreme thirst, noise-bleeds and flushing. Conversely, “cold” conditions require taking “heating” herbs to relieve symptoms such as slow pulse, smooth white tongue, weakness, low body temperature and shivering. Recently, Huang and Wu(16) reported that “heating” foods enhance PGE2 secretion and COX-2 expression by un-stimulated macrophages. In contrast, “cooling foods” suppress PGE2production in LPS-activated cells. Huang and Wu(16)further hypothesized that the PGE2 stimulating or inhibiting effects of food could serve as an indicator of hotness and coldness, respectively. However, because of limited classes of foods tested, the hypothesis that hot/cold natural characteristic of remedies associated with their abilities to enhance or suppress inflammatory mediators is controversial. Therefore, this study selected four widely used medicinal plants whose cold/hot nature is well known and accepted to be the subject herbs. The rhizome of Coptis chinensis and the root of Scutellaria
baicalensis, representing “cold drug”, and the rhizome of Zingiber officinale and the bark of Cinnamomum cassia,
representing “hot drug”, were investigated for a comparison of abilities to regulate the production of PGE2and NO in RAW264.7 macrophages.
MATERIALS AND METHODS I. Materials
RAW 264.7, a mouse macrophage cell line, was obtained from the American Type Culture Collection. The cells were cultured in DMEM medium supplemented with 10% heat-inactivated fetal bovine serum (Gibco BRL Life Technologies Inc., Grand Island, NY, U.S.A.) and main-tained at 37˚C in a humidified incubator containing 5% CO2. Moreover, lipopolysaccharide (LPS) and 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide (MTT) were purchased from Sigma Chemical Co. (St Louis, MO, U.S.A.).
II. Plant Materials and Extract Preparation
This study selected four Chinese medicinal plants for comparison of the effect of their natural “hot/cold” charac-teristics on the production of PGE2and NO. The plants were categorized into two classes: (1) cold: the rhizome of
Coptis chinensis and the root of Scutellaria baicalensis and
(2) hot: the rhizome of Zingiber officinale and the bark of
Cinnamomum cassia. These dried medicinal plants were
purchased from a Chinese medicine store in Taipei City (Taipei, Taiwan). The plants were cut into small species and boiled with 20 volumes of double-distilled water (1:20) for 20 min and then were centrifuged at 4000×g for 30 min to remove the marc. Supernatant was collected and freeze-dried. The residue of hot-water extracts were collected and weighed to calculate the yield of extraction. For cell culture experiment, weighed extract was dissolved in DMEM and filtered through 0.2 µm pore-size membrane and then diluted with DMEM to target concentration. III. Determination of PGE2and NO Production
To determine PGE2and NO production, the cells were scraped and suspended in DMEM medium containing 10% fetal bovine serum, then seeded at a density of 6 × 104cells per well in 96-well plastic culture dishes in 0.1 mL of medium. After 24-hr incubation at 37˚C, the adherent cells were washed three times with PBS and used for subsequent experiment. The adherent cells were incubated in 0.1-mL medium with extract from various medicinal plants, with or without 1µg/mL LPS. Eighteen hours after incubation, the medium was centrifuged at 1500×g and 4˚C for 5 min, and stored at -70˚C until assay. Moreover, cell viability was evaluated using the MTT method(17). The PGE2 concentra-tion was determined using commercial ELISA kits (Cayman Co., Ann Arbor, Mich., USA). Nitrite concentra-tion was measured as an indicator of NO producconcentra-tion based on the Griess reaction(18).
IV. Evaluation of Protein Levels of COX-2 and iNOS The RAW 264.7 cells were seeded at a density of 2 × 107cells per 6-cm culture dish in 5 mL of medium and incu-bated at 37˚C for 24 hr. After three washes, the adherent cells were incubated at 37˚C for 12 hr in 5 mL of medium in the presence or absence of 1 µg/mL LPS and the indicated concentration of hot-water extract from various herbs. One milliliter of condition medium was gathered for determining PGE2and nitrite. The adherent cells were washed three times with PBS, scraped into cold PBS and centrifuged at 500×g for 10 min at 4˚C. The cell pellets were resuspended in 0.1 mL of lysis buffer (50 mM Tris, pH 7.6, 0.01% EDTA, 1% Triton X-100, 1mM PMSF and 1µg/mL leupeptin) and centrifuged at 12,000×g for 20 min at 4˚C. Aliquots of the cell lysate used protein concentration assay by a BCA kit (Pierce Co., Rockford, IL, U.S.A.). Supernatant COX-2 and iNOS protein content were determined by immunoblot analysis. Briefly, supernatants containing 50 µg of protein were denatured, separated on 10% SDS-polyacrylamide gel and transferred to PVDF filters using a Holfer transblot cell, as instructed by the supplier. Meanwhile, filters were incu-bated in blocking solution at 4˚C overnight and then probed with primary antibodies. The antibodies used were a mouse
antibody to murine COX-2 (dilution 1/2000, Transduction Labotories, U.S.A.) and a mouse antibody to murine iNOS (dilution 1/500, Transduction Labotories, U.S.A.). The fil-ters were then incubated with secondary antibody, alkaline phosphatase conjugated anti-mouse IgG, and detected with a CDP-star luminace solution. Finally, the band intensities were quantified using a software supported photoimager (ImageMaster VDS, Amersham Pharmacia Biotech Co.). V. Statistical Analysis
All data are presented as mean ± SD for three indepen-dent experiments. The significance of differences at each sample concentration were analyzed by ANOVA and Duncan’s multiple range test using SPSS software. The paired t-test was used to evaluate the differences between the treatments and the control, with significant difference set at p < 0.05 (SPSS for Windows 10.0, SPSS Inc.).
RESULTS
I. PGE2Production in Un-activated Macrophages
Figure 1 shows the effects of hot-water extracts from
four medicinal plants on basal PGE2production in RAW 264.7 cells. Un-activated cells, after 18 h of incubation in DMEM culture medium, produced background levels of PGE2in the 0.2-0.6 ng/mL range. Hot-water extracts of
Coptis chinensis and Scutellaria baicalensis did not affect
basal PGE2production. However, Zingiber officinale and
Cinnamomum cassia displayed a biphasic effect on the
PGE2secretion by un-stimulated macrophages. At low con-centrations, varying from 50 to 200 µg/mL, Cinnamomum
cassia had an augmentative effect, with the largest induction
being at a concentration of 200 µg/mL. Moreover, the stim-ulating effect seemed to attenuate gradually with increasing concentration of the hot-water extract of Cinnamomum
cassia from 200 to 2000 µg/mL (Figure 1).
II. PGE2Production in LPS-activated Macrophages
Medium PGE2concentration increased markedly after treatment with LPS (Figure 2). Significant concentration-dependent inhibition of PGE2production was detected when cells were co-treated with LPS and various concentrations of the four plants extracts (Figure 2). The inhibitory potency of herbs could be ranked based on their IC50as Scutellaria
baicalensis (5.3 µg/mL) > Coptis chinensis (7.3 µg/mL) > Zingiber officinale (87.7 µg/mL) and Cinnamomum cassia
A. Coptis chinensis PGE 2 production (ng/mL) 12 9 6 3 0 Concentration (µg/mL) Control 1 5 10 50 100 200 Concentration (µg/mL) C. Zingiber officinale 12 9 6 3 0 PGE 2 production (ng/mL) 50 Control 100 200 500 1000 2000 a b b b a a a Concentration (µg/mL) 50 Control 100 200 500 1000 2000 PGE 2 production (ng/mL) 12 9 6 3 0 c b a d bc ab a D. Cinnamomun cassia B. Scutellaria baicalensis 9 6 3 0 12 Concentration (µg/mL) Control 1 5 10 50 100 200 PGE 2 production (ng/mL)
Figure 1. Effect of four medicinal plants on the basal PGE2production in RAW 264.7 cells. The values are expressed as means ± SD of tripli-cate tests. Means not sharing a common letter (a-d) were significantly different (p < 0.05) when analyzed by ANOVA and Duncan’s multiple range test.
(177.6 µg/mL). Importantly, the cold herbs, Coptis
chinen-sis and Scutellaria baicalenchinen-sis, showed a smaller IC50on the PGE2production than the hot-herbs. To avoid possible cytotoxic effect of herbs on PGE2production, cell viability was determined by MTT assay. Only Coptis chinensis at a concentration exceeding 200 µg/mL caused cytotoxicity, whereas other extracts at concentration of 2000 µg/mL did not influence cell viability (data not shown).
III. NO Production in Un-activated and LPS-activated
Macrophages
Analysis of NO production by measuring the nitrite with Griess reaction revealed that placing un-stimulated RAW 264.7 cells in culture medium for 18 hr produced a basal amount of nitrite (Figure 3). When the cells were incubated with hot-water extracts from these four plants in the absence of LPS, medium nitrite concentration was maintained at a background level similar to that in the un-stimulated control (data not shown). After treatment with LPS for 18 hr, the medium concentration of nitrite increased markedly compared with that of the control group (Figure 3). Like PGE2, the increase of NO was inhibited by co-treatment of cells with the four plant extracts. The
inhibitory potency of herbs could be ranked based on their IC50 as Coptis chinensis (101 µg/mL) > Scutellaria
baicalensis (104 µg/mL) > Cinnamomum cassia (1118
µg/mL) and Zingiber officinale (1388 µg/mL). IV. COX-2 and iNOS Protein Levels
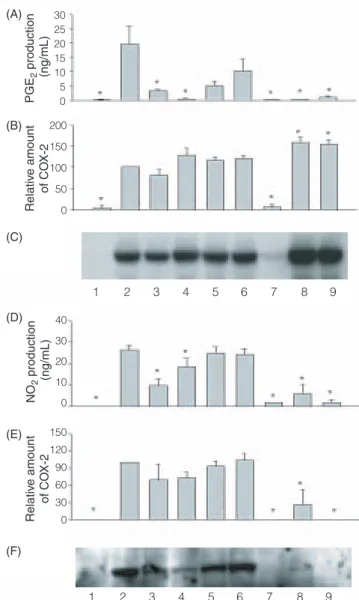
This work examined whether the inhibitory effects on inducible PGE2 and nitrite production resulted from direct effect of herbs on the expression of COX-2 and iNOS. As shown in Figure 4, RAW 264.7 cells did not express detectable COX-2 and iNOS protein when incubated in the medium without LPS for 12 hr, whereas LPS induced a marked increase in COX-2 and iNOS protein. In this study we selected two doses, namely 200 µg/mL and 2,000 µg/mL, at which the hot herbs and cold herbs had the largest inhibitory action on PGE2 production, respectively, to examine the effect of the herbs on COX-2 and iNOS expression. At 200 µg/mL of treated concentration, protein levels of COX-2 and iNOS did not change for any herbs except Coptis chinensis, which displayed a stronger but non-significant inhibitory effect than the others (Figure 4). Surprisingly, COX-2 induction by LPS was markedly atten-uated by Scutellaria baicalensis but significantly enhanced
PGE 2 production (ng/mL) 40 30 20 10 0 Concentration (µg/mL) Control 200 100 50 10 5 1 0 a a a a c c b b A. Coptis chinensis IC50 = 7.28 µg/mL PGE 2 pr oduction (ng/mL) 40 30 20 10 0 B. Scutellaria baicalensis IC50 = 5.27 µg/mL Concentration (µg/mL) Control 200 100 50 10 5 1 0 a a a a b b c c 40 30 20 10 0 PGE 2 production (ng/mL) Concentration (µg/mL) Control 2000 1000 500 200 100 50 0 a a a a b c c d C. Zingiber officinale IC50 = 87.7 µg/mL 40 30 20 10 0 PGE 2 pr oduction (ng/mL) Concentration (µg/mL) Control 2000 1000 500 200 100 50 0 a a a b c d e f D. Cinnemomun cassia IC50 = 177.6 µg/mL
Figure 2. Effect of four medicinal plants on the PGE2production in LPS-activated RAW264.7 cells. The values are expressed as means ± SD of triplicate tests. Means not sharing a common letter (a-f) were significantly different (p < 0.05) when analyzed by ANOVA and Duncan’s multiple range test.
by Zingiber officinale and Cinnamomum cassia at a con-centration of 2000 µg/mL. Moreover, the quantities of PGE2in the supernatants from each treatment were deter-mined again and were consistent with the former data presented in Figure 2. This study also was the first to report the reciprocal changes between PGE2production and COX-2 protein caused by Zingiber officinale and
Cinnamomum cassia at a concentration of 2000 µg/mL.
Relative iNOS expression in each treatment was responsi-ble for NO production level (Figure 4). Scutellaria
baicalensis, Zingiber officinale and Cinnamomum cassia at
concentrations of 2000 µg/mL inhibited iNOS protein levels by 100%, 74% and 100%, respectively, compared with the LPS-activated control group.
V. Dose-response Study
A dose-response study was undertaken to further confirm the effect of these herbs on the COX-2 expression. Western blotting analysis indicated that the protein level of COX-2 in LPS-activated macrophages gradually decreased with increasing concentrations of Scutellaria baicalensis, ranging from 200 to 2000 µg/mL (Figure 5A). Conversely,
Cinnamomum cassia markedly increased LPS-induced
COX-2 protein dose-dependently at concentrations ranging from 2,000 to 200 µg/mL (Figure 5C). Interestingly,
Cinnamomum cassia, like mitogen, could induce COX-2
expression by macrophages in the absence of LPS (Figure 5B).
DISCUSSION
Among the four medicinal plants tested in this study, hot-water extracts from the “cold” herbs, Coptis chinensis and Scutellaria baicalensis significantly inhibit PGE2 pro-duction and COX-2 expression by LPS-activated macrophages. In contrast, hot-water extracts from the “hot” plants, Zingiber officinale and Cinnamomum cassia, stimu-lated PGE2 production and COX-2 protein expression by non-activated macrophages. Consistent with Huang and Wu(16), these results demonstrated again that cold herbs inhibited and hot herbs stimulated PGE2formation, respec-tively. Furthermore, this study found that hot water extracts from the “hot plants”, Zingiber officinale and Cinnamomum
cassia stimulated COX-2 expression by LPS-activated cells
without a concomitant rise in PGE2formation. In the case of
Cinnamomum cassia, we suggest that one compound exerts
NO 2 secr ection ( µ M) 70 60 50 40 30 20 10 0 Concentration (µg/mL) Control 200 100 50 10 5 1 0 a b c cd de de e f A. Coptis chinensis IC50 = 100.8 µg/mL NO 2 secr ection ( µ M) 70 60 50 40 30 20 10 0 B. Scutellaria baicalensis IC50 = 103.7 µg/mL Concentration (µg/mL) Control 200 100 50 10 5 1 0 a b bc cd cd de de e a b c d e f g h NO 2 secr ection ( µ M) 70 60 50 40 30 20 10 0 D. Cinnamomum cassia IC50 = 1118 µg/mL Control 2000 1000 500 200 100 50 0 Concentration (µg/mL) Control 2000 1000 500 200 100 50 0 Concentration (µg/mL) NO 2 secr ection ( µ M) 70 60 50 40 30 20 10 0 C. Zingiber officinale IC50 = 1388 µg/mL a b c cd de ef g fg
Figure 3. Effect of four medicinal plants on nitric oxide (NO) production in LPS-activated RAW 264.7 cells. The values are expressed as means
± SD of triplicate tests. Means not sharing a common letter (a-f) were significantly different (p < 0.05) when analyzed by ANOVA and Duncan’s multiple range test.
transcription-inducing action begins at a low concentration and the other displays inhibitory effect on the catalytic activ-ity of COX-2 at a concentration higher than 200 µg/mL. In LPS-activated cells, the transcription-inducing effect of
Cinnamomum cassia may be over-shielded by the LPS, but
the catalytic inhibitory action of Cinnamomum cassia dis-plays continuously. Therefore, we could not observe the stimulatory effect of Cinnamomum cassia on the PGE2 pro-duction in LPS-activated cells in this study. Both the cold and hot medicinal plants examined in this study exhibited dose-dependent inhibitory effects on NO production and iNOS expression. The distinct effects of cold- and hot-
med-icinal plants on COX-2 expression but not on iNOS expres-sion implied that COX-2 might serve as a feasible indicator of distinguishing cold and hot substances in traditional Chinese medicine. Additionally, “hot” herbs represented a PGE2 and COX-2 inducing effect, which also appeared in Huang and Wu’s report(16). This fact provided another possi-bility that COX-2 inducing apossi-bility might be a determinant of cold and hot characteristics. Due to the experiment carried out lacking in the processes of absorption and metabolism in a whole body, the correlation between COX-2 and cold/hot system in vivo remains unclear and further studies are needed to clarify this problem.
It has been demonstrated that Scutellaria baicalensis displayed cytotoxicity on several cell types through inhibi-tion of PGE2and NO production(19,20). This study found that
Scutellaria baicalensis completely inhibited LPS-induced
PGE2 production and COX-2 expression by RAW 264.7 cells at concentrations of 200 and 2000 µg/mL, respectively, implied that Scutellaria baicalensis exerts its modulating action at post- and pre-translation levels. Indeed, Umeda et
al.(21)used the microsomal fraction of sheep vesicular glands as a COX enzyme source and reported that Scutellaria
baicalensis inhibited the biotransformation of arachidonic
acid, showing a post-translation regulation. Wagonin and oroxylin A, the active constituents in Scutellaria baicalensis, strongly block the expression of COX-2 through suppressing
(A) (B) (C) PGE 2 production (ng/mL) Relativ e amount of CO X-2 30 25 20 15 10 5 0 200 150 100 50 0 1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9 NO 2 production (ng/mL) Relativ e amount of CO X-2 (D) (E) (F) 40 30 20 10 0 150 120 90 60 30 0
Figure 4. Effect of four medicinal plants on COX-2 and iNOS protein
levels and PGE2and NO production in LPS-activated RAW 264.7 cells. The lanes represent cells treated without (2) or with 200 µg/mL of the hot-water extract, extracted from Coptis chinensis (3),
Scutellaria baicalensis (4), Zingiber officinale (5), Cinnamomum cas-sia (6) and 2000 µg/mL of the hot-water extract, extracted from Scutellaria baicalensis (7), Zingiber officinale (8) and Cinnamomum cassia (9), respectively, in the presence of LPS (1 µg /mL) for 12 hr.
Lane 1 presented the results content of the cells without any treatment. The values are expressed as means ± SD of triplicate tests. * p < 0.05 as determined by t-test as compared to the LPS-treated group.
(A) 1 2 3 4 5 (B) 1 2 3 4 5 (C) 1 2 3 4 5
Figure 5. Dose-response analysis of Scutellaria baicalensis and Cinnamomum cassia on COX-2 protein levels in RAW264.7 cells.
Lanes in panel A represent the COX-2 protein expression of the cells that were treated with 2000 (2), 1000 (3), 200 (4) µg/mL, respectively, of the hot-water extract of Scutellaria baicalensis in the presence of LPS (1 µg /mL) for 12 hr. Lanes in panel B and C represent the COX-2 protein expression of the cells that were treated with 2000 (2), 1000 (3), 200 (4) µg/mL, respectively, of the hot-water extract of Cinnamomum
cassia in the absence (panel B) and presence of LPS (panel C) for 12 hr.
Lane 1 and 5 in all panels represent the COX-2 protein content of the cells treated without or with LPS (1 µg /mL), respectively.
nuclear factor-kB activation(22,23). Berberine, the major component of the genera Coptis and Berberis, effectively inhibits COX-2 transcriptional activity in colon cancer cells(24). These previous results demonstrate that Coptis
chi-nensis and Scutellaria baicalensis exerted partial modulation
action on PGE2production at the pre-translation level. Additionally, induction of COX-2 and iNOS was prevented concomitantly by Scutellaria baicalensis, indicating
Scutellaria baicalensis is a plausible candidate for
develop-ing as a chemopreventive agent.
Although the reciprocal regulation on COX-2 and iNOS in activated cells caused by the hot herbs, Zingiber
officinale and Cinnamomum cassia, was contrary to the
most current finding. It seemed likely to occur, because the promoter region and the time course of expression of iNOS and COX-2 in RAW 264.7 cells was not the same. Similar opposite regulation has been observed in emodin, con-stituent of Rheum palmatum, which further supported our finding(23).
Ginger administration can relieve pain and other symp-toms in patients suffering from rheumatic disorders(25). The analgesic effect of ginger is considered to be associated with the inhibition of prostaglandin and leucotriene biosynthe-sis(26). Gingerol and structurally related pungent principles of ginger, including [8]-paradol and [8]-shogaol, have been identified as the most potent suppressors of prostaglandin synthesis(27). Although the inhibitory effect of ginger on the biotransformation of PGE2 has been demonstrated in a microsome fraction(21), intact cells(27) and humans(28), the effect of ginger on the expression of COX-2 remains unclear. In this study, we found that ginger displayed reciprocal effects on the COX-2 and PGE2in LPS-activated cells. And it needed further studies including purification of active con-stituents, investigation of regulating mechanism of single compound and interaction between active constituents, to clarify this reciprocal effect.
Few investigations have paid attention to the relation-ship between Cinnamomum cassia and inflammation. Recently, Lee et al.(29) reported that methanol extract of
Cinnamomum cassia could inhibit nitric production by
sup-pressing expression of iNOS in LPS-stimulated cells. Furthermore, cinnamaldehyde was identified as a major bioactive compound in Cinnamomum cassia(30). In addition to iNOS inhibition, this study was the first to report PGE2 inhibition and COX-2 stimulation of
Cinnamomum cassia by LPS-stimulated cells.
In conclusion, the hot herbs, Zingiber officinale and
Cinnamomum cassia, induced COX-2 protein and basal
PGE2production by RAW 264.7 macrophages. In contrast, the cold herbs, Coptis chinensis and Scutellaria baicalensis attenuated LPS-induced PGE2 production and COX-2 expression. These ex vivo analytical results might provide some cues with regard to the classification of hot/cold herbs.
ACKNOWLEDGMENTS
This study was supported by the grants from the National Science Council, Taiwan (NSC90-2320-B-264-001) and the Yuanpei University of Science and Technology, Hsinchu, Taiwan.
REFERENCES
1. Vane, J. R., Mitchell, J. A., Appleton, I., Tomlinson, A., Biohop-Bailey, D., Croxtall, J. and Willoughby, D. A. 1994. Inducible isoforms of cycloxygenase and nitric oxide synthase in inflammation. Proc. Natl. Acad. Sci. 91: 2046-2050.
2. Smith, W. L., Garavito, R. M. and Dewitt, D. L. 1996. Prostaglandin endoperoxide H synthases (cycloxyge-nase)-1 and 2. J. Biol. Chem. 271: 33157-33160. 3. DeWitt, D. L. 1991. Prostagladin endoperoxide
synthase: regulation of enzyme expression. Biochim. Biophys. Acta. 1083: 121-134.
4. Smith, W. L. and Marnett, L. J. 1991. Prostagladin endoperoxide synthase: structure and catalysis. Biochim. Biophys. Acta. 1083: 1-17.
5. Crofford, L. J., Wilder, R. L., Ristimaki, A. P., Sano, H., Remmers, E. F., Epps, H. R. and Hla, T. 1994. Cyclooxygenase-1 and -2 expression in rheumatoid synovial tissue. Effects of interleukin-1β, phorbol ester, and corticosteroids. J. Clin. Invest. 93: 1095-1101. 6. Inoue, H., Yokoyama, C., Hara, S., Tone, Y. and
Tanabe, T. 1995. Transcriptional regulation of human prostaglandin-endoperoxide synthase-2 gene by lipopolysaccharide and phorbol ester in vascular endothelial cells. J. Biol. Chem. 270: 24965-24971. 7. Hla, T., Ristimaki, A., Appleby, S. and Barriocanal, J. G.
1993. Cycloxygenase gene expression in inflammation and angiogenesis. Ann. N.Y. Acad. Sci. 696: 197-204. 8. Dubois, R. N., Abramaon, S. B., Crofford, L., Gupta, R.
A., Simon, L. S. and Van De Putte, L. B. A., Lipsky, P. E. 1998. Cyclooxygenase in biology and disease. FASEB J. 12: 1063-1073.
9. Mayer, B. and Hemmens, B. 1997. Biosynthesis and action of nitric oxide in mammalian cells. Trends Biochem. Sci. 22: 477-481.
10. MacMicking, J., Xie, Q. W. and Nathan, C. 1997. Nitric oxide and macrophage function. Ann. Rev. Immunol. 15: 323-350.
11. Szabo, C. 1995. Alterations in nitric oxide production in various forms of circulatory shock. New Horiz 3: 2-32. 12. Wheeler, A. P. and Bernard, G. R. 1999. Treating
patients with sever sepsis. New England J. Med. 340: 207-214.
13. Kolonel, L. V. 1988. Variability in diet and its relation to risk in ethnic and migration groups. Basic Life Sci. 43: 129-135.
14. Adlercreutz, H. 1990. Western diet and Western disease: some hormonal and biochemical mechanism and associ-ations. Scand. J. Clin. Lab. Invest. 50: 3-23.
23. Chen, Y. C., Yang, L. L., Lee and T. J. F. 2000. Oroxylin A inhibition of lipopolysaccharide-induced iNOS and COX-2 gene expression via suppression of nuclear factor-kB activation. Biochem. Pharmacol. 59: 1445-1457.
24. Fukuda, K., Hibiya, Y., Mutoh, M., Koshiji, M., Akao, S. and Fujiwara. H. 1999. Inhibition by berberine of cyclooxygenase-2 transcriptional activity in human colon cells. J. Ethnopharmacol. 66: 227-233.
25. Srivastava, K. C. and Mustafa, T. 1989. Ginger (Zingiber officinale) and rheumatic disorders. Med. Hypotheses 29: 25-28.
26. Srivastava, K. C. and Mustafa, T. 1992. Ginger (Zingiber officinale) in rheumatism and musculoskeletal disorders. Med. Hypotheses 39: 342-348.
27. Tjendraputra, E., Tran, V. H., Liu-Brennan, D., Roufogalis, B. D. and Duke, C. C. 2001. Effect of ginger constituents and synthetic analogues on cyclooxygenase-2 enzyme in intact cells. Bioorganic Chem. 29: 156-163.
28. Backon, J. 1986. Ginger: inhibition of thromboxane synthetase and stimulation of prostacyclin: relevance for medicine and psychiatry. Med. Hypotheses 20: 271-278.
29. Lee, H. S., Lee, S. H., Kim, Y. S., Song, C. G. and Kim, M. K. 2000. Effects of oriental medicinal plants on nitric oxide synthase activity. Food Sci. Biotech. 9: 77-82.
30. Lee, H. S., Kim, B. S. and Kim, M. K. 2002. Suppression effect of Cinnamomum cassia Bark-derived component on nitric oxide synthase. J. Agric. Food Chem. 50: 7700-7703.
Y. S., Park, K. K. and Lee, S. S. 2001. Molecular mech-anisms underlying chemopreventive activities of anti-inflammatory phytochemicals: down-regulation of COX-2 and iNOS through suppression of NF-kB activa-tion. Mutat. Res. 480-481: 243-268.
16. Huang, C. J. and Wu, M. C. 2002. Differential effects of foods traditionally regarded as heating and cooling on prostaglandin E2production by a macrophage cell line. J. Biomed. Sci. 9: 596-606.
17. Mosmann, T. 1983. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assay. J. Immunol. Methods 65: 55-63. 18. Kim, H., Lee, H. S., Chang, K. T., Ko, T. H., Baek, K.
J. and Kwon, N. S. 1995. Chloromethyl ketones block induction of nitric oxide synthase in murine macrophages by preventing activation of nuclear factor-B. J. Immunol. 154: 4741-4748.
19. Ye, F., Xui, L., Yi, J., Zhang, W. and Zhang, D. Y. 2002. Anticancer activity of Scutellaria baicalensis and its potential mechanism. J. Alternative & Complementary Med. 8: 567-572.
20. Zhang, D. Y., Wu, J., Ye, F., Xui, L., Jiang, S., Yi, J., Zhang, W., Wei, H., Sung, M., Wang, W. and Li, X. 2003. Inhibition of cancer cell proliferation and prostaglandin E2 synthesis by Scutellaria baicalensis. Cancer Res. 63: 4037-4043.
21. Umeda, M., Amagaya, S. and Ogihara, Y. 1988. Effects of certain herbal medicines on the biotransformation of arachidonic acid: a new pharmacological testing method using serum. J. Ethnopharmacol. 23: 91-98
22. Shen, S. C., Lee, W. R., Lin, H. Y., Huang, H. C., Ko, C. H., Yang, L. L. and Chen, Y. C. 2002. In vitro and in
vivo inhibitory activities of rutin, wogonin and
quercetin on lipopolysaccharide-induced nitric oxide and prostaglandin E2 production. Eur. J. Pharmacol. 446: 187-194.