行政院國家科學委員會專題研究計畫 成果報告
金黃色葡萄球菌超級抗原在異位性皮膚炎致病機轉所扮演
的角色
計畫類別: 個別型計畫
計畫編號: NSC92-2314-B-002-224-
執行期間: 92 年 08 月 01 日至 93 年 07 月 31 日
執行單位: 國立臺灣大學醫學院小兒科
計畫主持人: 林于粲
共同主持人: 江伯倫
報告類型: 精簡報告
處理方式: 本計畫可公開查詢
中 華 民 國 93 年 11 月 4 日
Differential Susceptibility to Staphylococcal Superantigen
(SsAg)-Induced Apoptosis of CD4
ⴙ
T Cells from Atopic
Dermatitis Patients and Healthy Subjects: The Inhibitory
Effect of IL-4 on SsAg-Induced Apoptosis
1
Yu-Tsan Lin,* Chen-Ti Wang,
†Chen-Tong Hsu,* Li-Fang Wang,
‡Wen-Yi Shau,
§Yao-Hsu Yang,* and Bor-Luen Chiang
2*
§This study had two aims: 1) to determine whether there are differences between atopic dermatitis (AD) patients and healthy subjects in staphylococcal superantigen (SsAg)-induced CD4ⴙT cell activation, cytokine production, chemokine receptor expres-sion, and apoptosis; and 2) to investigate the effect of IL-4 on SsAg-induced apoptosis. By using immunofluorescence and annexin V staining, we analyzed PBMC with or without staphylococcal enterotoxin B (SEB) stimulation in the presence or absence of rIL-4 or anti-IL-4-neutralizing Abs in 15 healthy subjects and 27 AD patients. We found that SEB preferentially induced production of Th1 cytokine in SEB-reactive (TCRV3ⴙor V12ⴙor V17ⴙ) CD4ⴙT cells from healthy subjects and Th2 cytokine in those from AD patients. SEB induced up-regulation of CXCR3ⴙcells in SEB-reactive CD4ⴙT cells from healthy subjects and CCR4ⴙ cells in those from AD patients. SEB-reactive CD4ⴙT cells from AD patients were more resistant to SEB-induced apoptosis than those from healthy subjects. There was no significant difference between AD and healthy subjects in SEB-induced activation of CD4ⴙT cells. CXCR3ⴙCD4ⴙT cells were more susceptible to SEB-induced apoptosis than CCR4ⴙCD4ⴙT cells in healthy subjects. Exogenously added IL-4 inhibited SEB-induced apoptosis of SEB-reactive CD4ⴙand CXCR3ⴙCD4ⴙT cells but not of CCR4ⴙCD4ⴙT cells in healthy subjects. Inhibition of endogenous IL-4 increased SEB-induced apoptosis of SEB-reactive CD4ⴙ T cells from AD patients. These results might provide new clues to the mechanism that SsAgs contribute to the persistence and exacerbation of allergic skin inflammation in AD. The Journal of Immunology, 2003, 171: 1102–1108.
A
topic dermatitis (AD)3is a genetically determined, chronically relapsing pruritic inflammatory skin disor-der. The cellular infiltrate in lesional skin of AD patients consists of mostly CD4⫹T cells and only a small number of CD8⫹ T cells, with the CD4:CD8 ratio varying approximately from 2:1 to 5:1 (1– 4). Numerous studies indicated that CD4⫹T cells play a pivotal role in allergic skin inflammation of AD. The imbalance between Th1 and Th2 cells contributes to the pathogenesis of AD. AD is a Th2-predominant immune disorder. The elevation of se-rum IgE levels and allergic skin inflammation is provoked by Th2 cytokines such as IL-4, IL-5, and IL-13 (1).
Th1 and Th2 cells differ in their chemokine receptor expression and their responsiveness to chemokines, which may explain the phenomenon of Th1 or Th2 favoring cell infiltration in inflamma-tory sites. Among CD4⫹T cells, CXCR3 is selectively expressed on Th1 cells, whereas CCR4 is selectively expressed on Th2 cells. These findings suggest that CXCR3 and CCR4 appear to serve as
the useful surface markers for identification of Th1 and Th2 cells among CD4⫹ T cells, respectively (5– 8). In comparison with healthy subjects, recent studies showed that AD patients had a higher percentage of CCR4⫹ cells and a lower percentage of CXCR3⫹cells in the CD4⫹T cells found in peripheral blood and lesional skin (9 –11).
The skin of AD patients exhibits a striking susceptibility to col-onization and infection with Staphylococcus aureus, which can secrete exotoxins including staphylococcal enterotoxin A, B (SEB), and C and toxic shock syndrome toxin-1. These exotoxins have superantigenic activity. They may penetrate the skin barrier and contribute to the persistence and exacerbation of allergic skin inflammation in AD through the stimulation of T cells bearing the specific variable region of-chain of TCR (TCRV). This leads to the local expansion of staphylococcal superantigen (SsAg)-reac-tive T cells (12–15). In addition, these exotoxins may play the role of allergens and thus may induce the production of functionally relevant specific IgE Abs (16 –20).
SEB is the most common SsAg on the skin of AD patients, which can stimulate human T cells bearing TCRV3, 12, 14, 15, 17, or 20 (21). SEB stimulation results in sequential T cell acti-vation, proliferation, Fas expression, cytokine secretion, and is subsequently followed by apoptosis in a dose- and time-dependent manner (22). Comparing AD patients with healthy subjects, in the culture supernatants of PBMC with SEB stimulation, AD patients produced more Th2 cytokines, such as IL-4 and IL-5, and less Th1 cytokines, such as IFN-␥ (23–25).
Activation-induced apoptosis of T cells provides a mechanism for the deletion of Ag-activated T cells, thereby leading to the resolution of inflammation and playing an important role in main-taining homeostasis of the immune response (26). Recent studies
Departments of *Pediatrics,†Orthopedics, and‡Dermatology, National Taiwan
Uni-versity Hospital and§Graduate Institute of Clinical Medicine, National Taiwan
Uni-versity College of Medicine, Taipei, Taiwan
Received for publication November 25, 2002. Accepted for publication May 7, 2003. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked advertisement in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1This work was supported by Research Grants from the National Science Council and
the National Taiwan University Hospital (NTUH92-M013).
2Address correspondence and reprint requests to Dr. Bor-Luen Chiang, Department
of Pediatrics, National Taiwan University Hospital, No. 7, Chung-Shan South Road, Taipei, Taiwan. E-mail address: gicmbor@ha.mc.ntu.edu.tw
3Abbreviations used in this paper: AD, atopic dermatitis; SEB, staphylococcal
en-terotoxin B; SsAg, staphylococcal superantigen; KU, kilounit.
conducted with polarized T cell lines or long-term T cell clones indicated that Th2 cells are more resistant to activation-induced apoptosis than Th1 cells (27–31). In addition, it was shown that IL-4 can inhibit activation-induced apoptosis of murine T cells (32, 33).
The mechanism that SsAgs contribute to the persistence and exacerbation of allergic skin inflammation in AD has been clarified only partially (34). The aims of this study were to determine whether there are differences between AD patients and healthy subjects in SEB-induced activation, cytokine production, chemo-kine receptor expression, and apoptosis of peripheral blood SEB-reactive CD4⫹T cells. We also observed whether there is a dif-ference between peripheral blood CXCR3⫹CD4⫹ and CCR4⫹CD4⫹T cells from healthy subjects in the susceptibility to SEB-induced apoptosis. Furthermore, we investigated the effect of exogenously added IL-4 on SEB-induced apoptosis of peripheral blood SEB-reactive CD4⫹T cells, CXCR3⫹CD4⫹T cells, and CCR4⫹CD4⫹T cells from healthy subjects. We also investigated the effect of inhibition of endogenous IL-4 by using anti-IL-4-neutralizing Abs on SEB-induced apoptosis of peripheral blood SEB-reactive CD4⫹T cells from AD patients. These may provide some new clues to the mechanism that SsAgs contribute to the persistence and exacerbation of allergic skin inflammation in AD.
Materials and Methods
Subjects
We undertook a cross-sectional study of 27 AD patients and 15 nonatopic healthy subjects. The AD patients were made up of 15 males and 12 fe-males with a mean age of 16.2 years and they fulfilled the diagnostic criteria of Hanifin and Rajka (35). The diagnosis of AD required the pres-ence of at least three of the following basic features: 1) pruritus, 2) typical morphology and distribution of skin lesions (flexural lichenification or ex-tensor involvement), 3) chronic or chronically relapsing dermatitis, and 4) personal or family history of atopy (asthma, allergic rhinitis, or AD). All of the AD patients in this study were identified by AD specialists and re-cruited from AD special clinics. The healthy subjects were made up of 8 males and 7 females with a mean age of 20.7 years. The study was ap-proved by the ethics committee at the National Taiwan University Hospital, and informed consent was obtained from all subjects or their parents.
Abs and reagents
CyChrome-conjugated anti-CD4, conjugated anti-CXCR3, and PE-conjugated anti-CCR4 mAbs (BD PharMingen, San Diego, CA) were used for staining cells expressing the respective surface Ag. FITC-conjugated anti-TCRV3, FITC-conjugated TCRV12, and FITC-conjugated anti-TCRV17 mAbs (Immunotech, Marseille, France) were used for staining SEB-reactive cells. PE-conjugated anti-CD69 mAbs (BD PharMingen) were used for staining activated cells. PE-conjugated anti-IFN-␥ and PE-conjugated anti-IL-4 mAbs (BD PharMingen) were used for staining in-tracellular cytokines. PE-conjugated annexin V and FITC-conjugated an-nexin V (BD PharMingen) were used for staining apoptotic cells based on the binding of annexin V to phosphatidylserine exposed on the cell surface. Unreactive isotype-matched mAbs were used as controls for background staining to determine the positive and negative populations of the cells. SEB (Sigma-Aldrich, St. Louis, MO) was used to stimulate PBMC. Mo-nensin (Sigma-Aldrich) was used to inhibit cytokine secretion. Saponin (Sigma-Aldrich) was used to make the membrane permeable so that the Abs could penetrate the intracellular region. The culture medium consisted of RPMI 1640 (HyClone Laboratories, Logan, UT) supplemented with 2 mM glutamine, 100g/ml streptomycin, 100 IU/ml penicillin, 10 mM HEPES, and 10% FBS. The annexin V binding buffer (BD PharMingen) consisted of 10 mM HEPES/NaOH (pH 7.4), 140 mM NaCl, and 2.5 mM CaCl2.
Determination of serum total IgE and SEB-specific IgE levels
Serum total IgE and SEB-specific IgE levels were measured by the use of AlaSTAT kits (DPC Biermann, Bad Nauheim, Germany), which is a liq-uid-phase enzyme immunoassay.
Cell cultures and stimulation
PBMC were isolated from heparinized whole blood by Ficoll-Hypaque (Pharmacia Biotech, Uppsala, Sweden) density gradient centrifugation. PBMC (2⫻106cells/ml) from AD patients and healthy subjects were cul-tured with or without SEB (1g/ml) stimulation for 12 and 24 h (37°C/5% CO2) in 24-well plates. To demonstrate the effect of exogenously added IL-4 on SEB-induced apoptosis, PBMC from healthy subjects were cul-tured with or without SEB (1g/ml) stimulation for 12 and 24 h in the presence or absence of human rIL-4 (10 ng/ml; PeproTech, London, U.K.). To demonstrate the effect of inhibition of endogenous IL-4 on SEB-in-duced apoptosis, PBMC from AD patients were cultured with or without SEB (1g/ml) stimulation for 12 and 24 h in the presence of anti-human IL-4 neutralizing Abs (20g/ml; R&D Systems, Minneapolis, MN), which can inhibit endogenous IL-4, or in the presence of isotype-matched control Abs (20g/ml) (R&D Systems). The optimal duration of the culture and the optimal concentrations of SEB, rIL-4, and anti-IL-4-neutralizing Abs and control Abs were predetermined according to the results of preliminary kinetic and titrating experiments (data not shown).
Detection of activation marker CD69 expression on SEB-reactive CD4⫹T cells
In PBMC cultured with or without SEB stimulation for 12 h, the activation marker CD69 expression on SEB-reactive (TCRV3⫹ or V12⫹ or V17⫹) CD4⫹T cells was detected. PBMC were stained with CyChrome-conjugated anti-CD4, FITC-CyChrome-conjugated anti-TCRV3, FITC-conjugated anti-TCRV12, FITC-conjugated anti-TCRV17, and PE-conjugated anti-CD69 mAbs at 4°C for 30 min.
Detection of intracellular Th1 and Th2 cytokine production in SEB-reactive CD4⫹T cells
In PBMC cultured with or without SEB stimulation for 12 h in the presence of 10g/ml monensin, intracellular IFN-␥ (Th1 cytokine) and IL-4 (Th2 cytokine) of SEB-reactive (TCRV3⫹or V12⫹or V17⫹) CD4⫹T cells were detected. PBMC were stained with CyChrome-conjugated anti-CD4, FITC-conjugated anti-TCRV3, FITC-conjugated anti-TCRV12, and FITC-conjugated anti-TCRV17 mAbs at 4°C for 30 min. Then the cells were fixed with 4% paraformaldehyde (Sigma-Aldrich) in PBS at 24°C for 20 min, were made permeable with 0.5% saponin and 1% FBS in PBS at 24°C for 10 min, and were stained intracellularly with PE-conjugated anti-IFN-␥ or PE-conjugated anti-IL-4 mAbs at 4°C for 30 min.
Detection of CXCR3 and CCR4 expression on SEB-reactive CD4⫹T cells
In PBMC cultured with or without SEB stimulation for 12 h, CXCR3 and CCR4 expression on SEB-reactive (TCRV3⫹ or V12⫹ or V17⫹) CD4⫹T cells were detected. PBMC were stained with CyChrome-conju-gated anti-CD4, FITC-conjuCyChrome-conju-gated anti-TCRV3, FITC-conjugated anti-TCRV 12,FITC-conjugatedanti-TCRV17,andeitherPE-conjugatedanti-CXCR3 or PE-conjugated anti-CCR4 mAbs at 4°C for 30 min.
Detection of apoptosis of SEB-reactive CD4⫹T cells
In PBMC cultured with or without SEB stimulation for 12 and 24 h in the presence or absence of rIL-4 or anti-IL-4-neutralizing Abs, apoptosis of SEB-reactive (TCRV3⫹ or V12⫹or V17⫹) CD4⫹T cells was de-tected. PBMC were stained with CyChrome-conjugated anti-CD4, FITC-conjugated anti-TCRV3, conjugated anti-TCRV12, and FITC-conjugated anti-TCRV17 mAbs at 4°C for 30 min. Then the cells were resuspended in the annexin V binding buffer and stained with PE-conju-gated annexin V at 24°C for 15 min.
Detection of apoptosis of CXCR3⫹CD4⫹or CCR4⫹CD4⫹T cells
In PBMC from healthy subjects that were cultured with or without SEB stimulation for 12 and 24 h in the presence or absence of rIL-4, apoptosis of CXCR3⫹CD4⫹ or CCR4⫹CD4⫹ T cells was detected. PBMC were stained with CyChrome-conjugated CD4 and either PE-conjugated anti-CXCR3 or PE-conjugated anti-CCR4 mAbs at 4°C for 30 min. Then the cells were resuspended in the annexin V binding buffer and stained with FITC-conjugated annexin V at 24°C for 15 min.
Flow cytometric analysis
Three-color immunofluorescence analysis was performed. For each sam-ple, 100,000 cells were acquired and analyzed by a FACScan flow cytom-eter (BD PharMingen) equipped with CellQuest software. CD4⫹T cells were gated for analysis. Quadrants of dot plots were set according to the staining of isotype-matched control mAbs.
1103 The Journal of Immunology
Statistical analysis
Data are presented as means⫾ SEM of the percentages of respective sub-populations. The Mann-Whitney U test was used to compare variables between AD patients and healthy subjects. The Wilcoxon signed rank test was used to compare variables between cells from the same group of sub-jects. The STATA 7.0 statistical software (Stata, College Station, TX) was used for statistical analysis. A value of p⬍ 0.05 was considered to be statistically significant.
Results
Higher serum total IgE and SEB-specific IgE levels in AD
The total serum IgE levels in AD patients were significantly higher than those in healthy subjects (median and range, 863 and 41– 6950 kilounit (KU)/L vs 42 and 8 –171 KU/L, p⫽ 0.008). Serum SEB-specific IgE levels in AD patients were significantly higher than those in healthy subjects (median and range, 4.16 and 0.24 – 39.2 KU/L vs 0 and 0 – 0.85 KU/L, p⫽ 0.013).
The effect of SEB stimulation on the percentages of SEB-reactive cells in CD4⫹T cells is very similar in AD and healthy subjects
In both AD and healthy subjects, SEB stimulation for 12 and 24 h significantly increased the percentages of SEB-reactive (TCRV3⫹or V12⫹or V17⫹) cells in peripheral blood CD4⫹ T cells (all p⬍ 0.05). Meanwhile, there was no significant differ-ence between AD and healthy subjects in the effect of SEB stim-ulation for 12 and 24 h on the percentages of SEB-reactive cells in CD4⫹T cells ( p⫽ 0.825 for 12 h, p ⫽ 0.192 for 24 h; Fig. 1).
SEB-induced activation of SEB-reactive CD4⫹T cells is very similar in AD and healthy subjects
In both AD and healthy subjects, SEB stimulation for 12 h signif-icantly increased the percentages of activated (CD69⫹) cells in peripheral blood SEB-reactive (TCRV3⫹or V12⫹or V17⫹) CD4⫹T cells (both p⬍ 0.05). Meanwhile, there was no significant difference between AD and healthy subjects in SEB-induced acti-vation of SEB-reactive CD4⫹T cells ( p⫽ 0.713; Fig. 2).
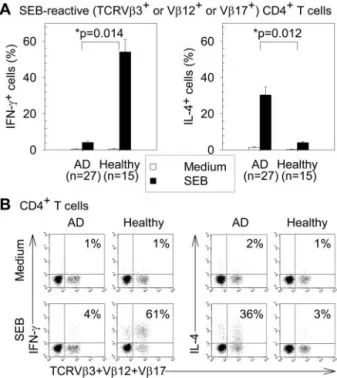
SEB preferentially induced production of Th1 cytokine in SEB-reactive CD4⫹T cells from healthy subjects and Th2 cytokine in those from AD patients
Without SEB stimulation, no intracellular IFN-␥ (Th1 cytokine) or IL-4 (Th2 cytokine) was detectable in peripheral blood
SEB-reac-tive (TCRV3⫹or V12⫹or V17⫹) CD4⫹T cells from AD or healthy subjects. In both AD and healthy subjects, SEB stimulation for 12 h significantly increased the percentages of IFN-␥⫹cells and IL-4⫹ cells in SEB-reactive CD4⫹T cells (all p ⬍ 0.05). SEB-induced increase of the percentages of IFN-␥⫹cells in
SEB-FIGURE 3. SEB-induced production of IFN-␥ and IL-4 in peripheral blood SEB-reactive CD4⫹T cells from AD patients and healthy subjects. PBMC were cultured with or without SEB (1g/ml) stimulation for 12 h. A, The means⫾ SEM of the percentages of IFN-␥⫹or IL-4⫹cells in SEB-reactive (TCRV3⫹or V12⫹or V17⫹) CD4⫹T cells. B, Repre-sentative dot plots by gating on CD4⫹T cells. The percentages of IFN-␥⫹ or IL-4⫹cells in SEB-reactive (TCRV3⫹or V12⫹or V17⫹) CD4⫹T cells are indicated.ⴱ, p value for comparison of AD patients with healthy subjects for the difference between the cell percentages with and without SEB stimulation.
FIGURE 1. Effect of SEB stimulation on the percentages of SEB-reac-tive cells in peripheral blood CD4⫹T cells from AD patients and healthy subjects. PBMC were cultured with or without SEB (1g/ml) stimulation for 12 and 24 h. Data represent the means⫾ SEM of the percentages of SEB-reactive (TCRV3⫹or V12⫹or V17⫹) cells in CD4⫹T cells.ⴱ, p value for comparison of AD patients with healthy subjects for the dif-ference between the cell percentages with and without SEB stimulation.
FIGURE 2. SEB-induced activation of peripheral blood SEB-reactive CD4⫹T cells from AD patients and healthy subjects. PBMC were cultured with or without SEB (1g/ml) stimulation for 12 h. Data represent the means⫾ SEM of the percentages of activated (CD69⫹) cells in SEB-reactive (TCRV3⫹or V12⫹or V17⫹) CD4⫹T cells.ⴱ, p value for comparison of AD patients with healthy subjects for the difference between the cell percentages with and without SEB stimulation.
reactive CD4⫹T cells in healthy subjects was significantly greater than that in AD patients ( p⫽ 0.014). In contrast, the SEB-induced increase of the percentages of IL-4⫹cells in SEB-reactive CD4⫹ T cells in AD patients was significantly greater than that in healthy subjects ( p⫽ 0.012; Fig. 3).
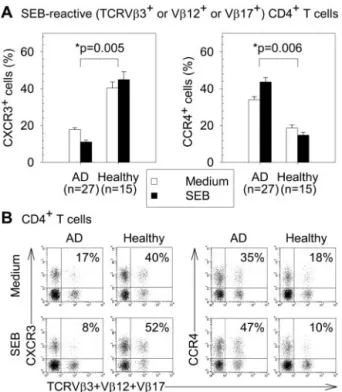
SEB induced up-regulation of CXCR3⫹cells in SEB-reactive CD4⫹T cells from healthy subjects and CCR4⫹cells in those from AD patients
In peripheral blood SEB-reactive (TCRV3⫹ or V12⫹ or V17⫹) CD4⫹T cells without SEB stimulation, AD patients had significantly higher percentages of CCR4⫹cells and lower per-centages of CXCR3⫹cells than healthy subjects (both p⬍ 0.05). SEB stimulation for 12 h significantly increased the percentages of CCR4⫹cells and decreased the percentages of CXCR3⫹cells in SEB-reactive CD4⫹T cells from AD patients (both p⬍ 0.05). In contrast, SEB stimulation for 12 h significantly increased the per-centages of CXCR3⫹ cells and decreased the percentages of CCR4⫹cells in SEB-reactive CD4⫹T cells from healthy subjects (both p⬍ 0.05). There were significant differences between AD and healthy subjects in the effects of SEB stimulation on the per-centages of CXCR3⫹cells ( p⫽ 0.005) and CCR4⫹cells ( p⫽ 0.006) in SEB-reactive CD4⫹T cells (Fig. 4).
SEB-reactive CD4⫹T cells from AD patients were more resistant to SEB-induced apoptosis than those from healthy subjects
In peripheral blood SEB-reactive (TCRV3⫹ or V12⫹ or V17⫹) CD4⫹T cells cultured for 24 h without SEB stimulation,
AD patients had significantly lower percentages of spontaneous apoptotic cells than healthy subjects (3.5%⫾ 0.4% vs 6.4% ⫾ 1%,
p⬍ 0.05). In both AD and healthy subjects, SEB stimulation for
24 h significantly increased the percentages of apoptotic cells in SEB-reactive CD4⫹T cells (both p⬍ 0.05). AD patients had a sig-nificantly lower “percentage of induced apoptotic cells in SEB-reactive CD4⫹T cells” than healthy subjects ( p⫽ 0.001), which is defined as the SEB-induced increase of the percentage of apoptotic cells and is determined by subtracting the percentage of apoptotic cells without SEB stimulation from the percentage of apoptotic cells with SEB stimulation (Fig. 5).
Exogenously added IL-4 inhibited SEB-induced apoptosis of SEB-reactive CD4⫹T cells from healthy subjects
In peripheral blood SEB-reactive (TCRV3⫹ or V12⫹ or V17⫹) CD4⫹T cells from healthy subjects that were cultured for 24 h without SEB stimulation, there was no significant difference in the percentages of spontaneous apoptotic cells between those in the presence of rIL-4 and those in the absence of rIL-4 (4.7%⫾ 0.7% vs 6.4%⫾ 1%). In the presence or absence of rIL-4, SEB stimulation for 24 h significantly increased the percentages of apo-ptotic cells in SEB-reactive CD4⫹T cells from healthy subjects (both p⬍ 0.05). The addition of rIL-4 to SEB-stimulated cells from healthy subjects for 24 h significantly reduced the percent-ages of SEB-induced apoptotic cells in SEB-reactive CD4⫹T cells
FIGURE 4. Effect of SEB stimulation on CXCR3 and CCR4 expression on peripheral blood SEB-reactive CD4⫹ T cells from AD patients and healthy subjects. PBMC were cultured with or without SEB (1g/ml) stimulation for 12 h. A, The means⫾ SEM of the percentages of CXCR3⫹ or CCR4⫹cells in SEB-reactive (TCRV3⫹or V12⫹or V17⫹) CD4⫹ T cells. B, Representative dot plots by gating on CD4⫹T cells. The per-centages of CXCR3⫹ or CCR4⫹ cells in SEB-reactive (TCRV3⫹ or V12⫹or V17⫹) CD4⫹T cells are indicated.ⴱ, p value for comparison of AD patients with healthy subjects for the difference between the cell percentages with and without SEB stimulation.
FIGURE 5. SEB-induced apoptosis of peripheral blood SEB-reactive CD4⫹ T cells from AD patients and healthy subjects and the effect of exogenously added IL-4 on SEB-induced apoptosis of these cells from healthy subjects. PBMC were cultured with or without SEB (1g/ml) stimulation for 12 and 24 h in the presence or absence of rIL-4 (10 ng/ml). A, Representative dot plots by gating on CD4⫹T cells cultured for 24 h. The percentages of apoptotic cells determined by annexin V staining in SEB-reactive (TCRV3⫹or V12⫹or V17⫹) CD4⫹T cells are indi-cated. B, Time kinetics of SEB-induced apoptosis. Data represent the means⫾ SEM of “the percentage of induced apoptotic cells in SEB-reactive (TCRV3⫹or V12⫹or V17⫹) CD4⫹T cells,” which is de-termined by subtracting the percentage of apoptotic cells without SEB stimulation from the percentage of apoptotic cells with SEB stimulation.ⴱ, p value for comparison of two groups for the percentages of SEB-induced apoptotic cells.
1105 The Journal of Immunology
( p⫽ 0.028). In SEB-reactive CD4⫹T cells with SEB stimulation for 24 h, there was no significant difference in the percentages of SEB-induced apoptotic cells between those from healthy subjects in the presence of rIL-4 and those from AD patients ( p⫽ 0.805; Fig. 5).
Inhibition of endogenous IL-4 increased SEB-induced apoptosis of SEB-reactive CD4⫹T cells from AD patients
In peripheral blood SEB-reactive (TCRV3⫹ or V12⫹ or V17⫹) CD4⫹ T cells from AD patients that were cultured for 24 h without SEB stimulation, there was no significant difference in the percentages of spontaneous apoptotic cells among those in the presence of anti-IL-4-neutralizing Abs, those in the presence of control Abs, and those in the absence of any added Abs (5.9%⫾ 1.1% vs 4.9%⫾ 1.3% vs 4.2% ⫾ 0.9%). In all three groups of cells, SEB stimulation for 24 h significantly increased the percent-ages of apoptotic cells (all p⬍ 0.05). The addition of anti-IL-4-neutralizing Abs, which can inhibit endogenous IL-4, to SEB-stim-ulated cells from AD patients for 24 h significantly increased the percentages of SEB-induced apoptotic cells in SEB-reactive CD4⫹ T cells ( p⫽ 0.0002 compared with the addition of control Abs,
p⫽ 0.001 compared with no addition of any Abs). In SEB-reactive
CD4⫹T cells from AD patients with SEB stimulation for 24 h, there was no significant difference in the percentages of SEB-in-duced apoptotic cells between those in the presence of control Abs and those in the absence of any added Abs ( p⫽ 0.455; Fig. 6).
CXCR3⫹CD4⫹T cells were more susceptible to SEB-induced apoptosis than CCR4⫹CD4⫹T cells in healthy subjects
There was no significant difference in the percentages of sponta-neous apoptotic cells between peripheral blood CXCR3⫹CD4⫹ and CCR4⫹CD4⫹T cells from healthy subjects that were cultured for 24 h without SEB stimulation (12.5%⫾ 1.4% vs 10.6% ⫾ 0.9%). In both CXCR3⫹CD4⫹ and CCR4⫹CD4⫹ T cells from healthy subjects, SEB stimulation for 24 h significantly increased the percentages of apoptotic cells (both p⬍ 0.05). The percentages of SEB-induced apoptotic cells in CXCR3⫹CD4⫹T cells were significantly higher than those in CCR4⫹CD4⫹T cells in healthy subjects ( p⫽ 0.028; Fig. 7).
Exogenously added IL-4 inhibited SEB-induced apoptosis of CXCR3⫹CD4⫹T cells but not of CCR4⫹CD4⫹T cells in healthy subjects
In peripheral blood CD4⫹T cells from healthy subjects that were cultured for 24 h without SEB stimulation, there was no significant difference in the percentages of spontaneous apoptotic cells among CXCR3⫹CD4⫹T cells in the absence of rIL-4, CXCR3⫹CD4⫹T cells in the presence of rIL-4, CCR4⫹CD4⫹T cells in the absence of rIL-4, and CCR4⫹CD4⫹T cells in the presence of rIL-4 (12.5%
⫾ 1.4% vs 11.6% ⫾ 0.6% vs 10.6% ⫾ 0.9% vs 10.2% ⫾ 1.3%).
In all four groups of cells, SEB stimulation for 24 h significantly increased the percentages of apoptotic cells (all p⬍ 0.05). The addition of rIL-4 to SEB-stimulated cells from healthy subjects for
FIGURE 6. The effect of inhibition of endogenous IL-4 on SEB-in-duced apoptosis of peripheral blood SEB-reactive CD4⫹T cells from AD patients. PBMC were cultured with or without SEB (1g/ml) stimulation for 12 and 24 h in the presence of anti-IL-4-neutralizing Abs (20g/ml), in the presence of control Abs (20g/ml), or in the absence of any added Abs. A, Representative dot plots by gating on CD4⫹T cells cultured for 24 h. The percentages of apoptotic cells determined by annexin V staining in SEB-reactive (TCRV3⫹or V12⫹or V17⫹) CD4⫹T cells are in-dicated. B, Time kinetics of SEB-induced apoptosis. Data represent the means⫾ SEM of “the percentage of induced apoptotic cells in SEB-reactive (TCRV3⫹or V12⫹or V17⫹) CD4⫹T cells,” which is de-termined by subtracting the percentage of apoptotic cells without SEB stimulation from the percentage of apoptotic cells with SEB stimulation.ⴱ, p value for comparison of two groups for the percentages of SEB-induced apoptotic cells.
FIGURE 7. SEB-induced apoptosis and the effect of exogenously added IL-4 on SEB-induced apoptosis of peripheral blood CXCR3⫹ CD4⫹ or CCR4⫹CD4⫹T cells from healthy subjects. PBMC were cultured with or without SEB (1g/ml) stimulation for 12 and 24 h in the presence or absence of rIL-4 (10 ng/ml). A, Representative dot plots by gating on CD4⫹T cells cultured for 24 h. The percentages of apoptotic cells deter-mined by annexin V staining in CXCR3⫹CD4⫹or CCR4⫹CD4⫹T cells are indicated. B, Time kinetics of SEB-induced apoptosis. Data represent the means⫾ SEM of “the percentage of SEB-induced apoptotic cells in CXCR3⫹CD4⫹or CCR4⫹CD4⫹ T cells,” which is determined by sub-tracting the percentage of apoptotic cells without SEB stimulation from the percentage of apoptotic cells with SEB stimulation.ⴱ, p value for com-parison of two groups for the percentages of SEB-induced apoptotic cells.
24 h significantly reduced the percentages of SEB-induced apo-ptotic cells in CXCR3⫹CD4⫹ T cells ( p ⫽ 0.046) but not in CCR4⫹CD4⫹T cells ( p⫽ 0.225). In CD4⫹T cells from healthy subjects with SEB stimulation for 24 h in the presence of rIL-4, there was no significant difference in the percentages of SEB-in-duced apoptotic cells between CXCR3⫹CD4⫹and CCR4⫹CD4⫹ T cells ( p⫽ 0.080; Fig. 7).
Discussion
This study showed that AD patients and healthy subjects differed markedly in SEB-induced cytokine production, chemokine recep-tor expression, and apoptosis of peripheral blood SEB-reactive CD4⫹T cells. However, there was no significant difference be-tween AD patients and healthy subjects in the effect of SEB stim-ulation on the percentages of SEB-reactive cells in CD4⫹T cells and SEB-induced activation of SEB-reactive CD4⫹T cells.
Our results indicated that SEB stimulation significantly in-creased the percentages of SEB-reactive cells in peripheral blood CD4⫹T cells and significantly activated SEB-reactive CD4⫹ T cells in both AD and healthy subjects. Meanwhile, there was no significant difference between AD and healthy subjects in the ef-fect of SEB stimulation on the percentages of SEB-reactive cells in CD4⫹T cells and SEB-induced activation of SEB-reactive CD4⫹ T cells (Figs. 1 and 2). In addition, previous studies showed that the application of SEB on intact skin induced local up-regulation of SEB-reactive T cells and dermatitis in both AD patients and healthy subjects (15, 36). These findings suggest that both the pro-liferative and activation response of CD4⫹T cells to SEB stimu-lation are similar in AD patients and healthy subjects.
In this study, intracellular cytokine staining was used to detect cytokine production at the single-cell level. We found that SEB preferentially induced production of IFN-␥ (Th1 cytokine) in pe-ripheral blood SEB-reactive CD4⫹T cells from healthy subjects and IL-4 (Th2 cytokine) in those from AD patients (Fig. 3). These results are compatible with previous reports in that cytokine pro-duction was detected by the measurement of cytokine levels in the cell culture supernatants (23–25). Moreover, this study and previ-ous reports revealed that serum SEB-specific IgE levels in AD patients were significantly higher than those in healthy subjects (16 –20). With regard to chemokine receptor expression, this study showed that AD patients had significantly higher percentages of CCR4⫹cells and lower percentages of CXCR3⫹cells than healthy subjects in SEB-reactive CD4⫹T cells without SEB stimulation. Furthermore, SEB induced up-regulation of CXCR3⫹ cells in SEB-reactive CD4⫹ T cells from healthy subjects and CCR4⫹ cells in those from AD patients (Fig. 4). These findings suggest that SEB preferentially induces a Th1 immune response in healthy subjects and a Th2 immune response in AD patients.
This study showed that SEB induced apoptosis of peripheral blood SEB-reactive CD4⫹T cells in both AD and healthy subjects. It also induced apoptosis of both CXCR3⫹CD4⫹ and CCR4⫹CD4⫹T cells from healthy subjects. However, SEB-reac-tive CD4⫹T cells from AD patients were more resistant to spon-taneous apoptosis and SEB-induced apoptosis than those from healthy subjects (Fig. 5). Because it is difficult to detect intracel-lular cytokines and apoptosis simultaneously in the same single cell, we used CXCR3 and CCR4 as the surface markers for iden-tification of Th1 and Th2 cells among CD4⫹T cells, respectively, as suggested by previous reports (5– 8). We found that CXCR3⫹CD4⫹T cells were more susceptible to SEB-induced apo-ptosis than CCR4⫹CD4⫹T cells in healthy subjects although they had a similar susceptibility to spontaneous apoptosis (Fig. 7). This might imply that Th2 cells are more resistant to SEB-induced ap-optosis than Th1 cells in the subjects with normal cytokine
bal-ances. Previous studies indicating that Th2 cells are more resistant to activation-induced apoptosis than Th1 cells were conducted ei-ther with polarized T cell lines or with long-term T cell clones (27–31). Our study is the first one to demonstrate the susceptibility to activation-induced apoptosis of T cells in human disease. In addition, the fact of using freshly isolated lymphocytes in our study rules out that in vitro manipulations, such as selection due to multiple restimulation (27–30) or the use of exogenous factors for polarizing cells toward specific cytokine profiles (31), may modify the susceptibility of cells to proapoptotic stimuli.
This study demonstrated that exogenously added IL-4 inhibited SEB-induced apoptosis of peripheral blood SEB-reactive CD4⫹T cells from healthy subjects although it had no effect on the spon-taneous apoptosis of these cells (Fig. 5). This result is compatible with previous reports indicating IL-4 can inhibit SsAg-induced apoptosis of murine T cells (32, 33). In addition, this study showed that inhibition of endogenous IL-4 by using anti-IL-4-neutralizing Abs increased SEB-induced apoptosis of SEB-reactive CD4⫹ T cells from AD patients, although it had no effect on the spontane-ous apoptosis of these cells (Fig. 6). This result suggests that en-dogenous IL-4 can inhibit SEB-induced apoptosis of SEB-reactive CD4⫹T cells from AD patients. These findings imply that CD4⫹ T cells from AD patients following SEB stimulation produced more IL-4, which can inhibit SEB-induced apoptosis, than those from healthy subjects was a partial reason why CD4⫹T cells from AD patients were more resistant to SEB-induced apoptosis than those from healthy subjects. Moreover, our study showed that ex-ogenously added IL-4 inhibited SEB-induced apoptosis of CXCR3⫹ CD4⫹ T cells but not of CCR4⫹ CD4⫹ T cells in healthy subjects although it had no effect on the spontaneous apo-ptosis of these cells. CXCR3⫹CD4⫹and CCR4⫹CD4⫹T cells from healthy subjects had a similar susceptibility to SEB-induced apoptosis in the presence of exogenously added IL-4 (Fig. 7). These are new findings that have never been reported. To clarify whether it implies that IL-4 inhibits SEB-induced apoptosis of Th1 cells but not of Th2 cells would require further study. The low susceptibility to SEB-induced apoptosis of CCR4⫹CD4⫹T cells in the absence of exogenously added IL-4 was a possible reason why the inhibitory effect of exogenously added IL-4 on SEB-in-duced apoptosis was not significant in these cells.
The results of this study suggest a potential mechanism that SsAgs contribute to the persistence and exacerbation of allergic skin inflammation in AD as in the following cases. SsAg stimu-lation of skin T cells in AD patients induces a Th2 immune re-sponse, including the up-regulation of IL-4-producing cells and CCR4-expressing cells. The intrinsic property of Th2 cells and/or intralesional cytokine microenvironment such as the presence of IL-4 may lead to the inhibition of SsAg-induced apoptosis of skin T cells in AD patients. This may impair deletion of intralesional SsAg-activated T cells and resolution of allergic skin inflammation after SsAg stimulation. The injured skin barrier results in further penetration of SsAgs. Long-term exposure of skin T cells to SsAgs in AD patients may lead to preferential loss of Th1 cells and the predominance of more long-lived Th2 cells. The “vicious circle” continues as long as SsAgs are present in the skin. On the other hand, SsAg stimulation of skin T cells through the injured skin barrier induces a Th1 immune response in healthy subjects. Rapid SsAg-induced apoptosis of skin Th1 cells in healthy subjects re-sults in an adequate resolution of the immune response after SsAg stimulation. Therefore, this may imply that SsAgs can “induce” AD in the subjects with a genetic predisposition to atopy, but they cannot induce AD in those without this predisposition.
In contrast to the major role of CD4⫹T cells in allergic skin inflammation of AD, the minor role of CD8⫹ T cells in AD is 1107 The Journal of Immunology
implied (1). There was limited evidence for involvement of CD8⫹ T cells in AD. It has been shown that SEB stimulation induced production of Th2-like cytokines such as IL-4, IL-5, and IL-13, but not of IFN-␥, in both CD4⫹and CD8⫹T cells isolated from le-sional skin of AD patients (37), which suggests that CD8⫹T cells have more than bystander functions in AD. In the future, we will conduct further study to clarify the role of CD8⫹T cells in the mechanism that SsAgs contribute to the persistence and exacerba-tion of allergic skin inflammaexacerba-tion in AD.
In conclusion, this study demonstrated that SEB preferentially induced production of Th1 cytokine in peripheral blood SEB-re-active CD4⫹T cells from healthy subjects and Th2 cytokine in those from AD patients. SEB induced up-regulation of CXCR3⫹ cells in SEB-reactive CD4⫹ T cells from healthy subjects and CCR4⫹cells in those from AD patients. SEB-reactive CD4⫹ T cells from AD patients were more resistant to SEB-induced apo-ptosis than those from healthy subjects. However, there was no significant difference between AD patients and healthy subjects in SEB-induced activation of SEB-reactive CD4⫹T cells. Further-more, CXCR3⫹CD4⫹T cells were more susceptible to SEB-in-duced apoptosis than CCR4⫹CD4⫹T cells in healthy subjects. This study also showed that exogenously added IL-4 inhibited SEB-induced apoptosis of SEB-reactive CD4⫹ T cells and CXCR3⫹CD4⫹T cells but not of CCR4⫹CD4⫹T cells in healthy subjects. Inhibition of endogenous IL-4 by using anti-IL-4-neutraliz-ing Abs increased SEB-induced apoptosis of SEB-reactive CD4⫹T cells from AD patients. These results provide further understanding of the mechanism that SsAgs contribute to the persistence and exacer-bation of allergic skin inflammation in AD, which is important for exploring the specific therapy and prevention of AD.
Acknowledgments
We thank Bo-Shiou Lin, Ya-Hui Chuang, and Ching-Yi Wu for technical assistance and Yin-Hsiu Chien for help in blood sampling.
References
1. Herz, U., R. Bunikowski, and H. Renz. 1998. Role of T cells in atopic dermatitis: new aspects on the dynamics of cytokine production and the contribution of bacterial superantigens. Int. Arch. Allergy Immunol. 115:179.
2. Leung, D. Y., A. K. Bhan, E. E. Schneeberger, and R. S. Geha. 1983. Charac-terization of the mononuclear cell infiltrate in atopic dermatitis using monoclonal antibodies. J. Allergy Clin. Immunol. 71:47.
3. Willemze, R., C. B. Graaff-Reitsma, J. Cnossen, W. A. Van Vloten, and C. J. Meijer. 1983. Characterization of T-cell subpopulations in skin and periph-eral blood of patients with cutaneous T-cell lymphomas and benign inflammatory dermatoses. J. Invest. Dermatol. 80:60.
4. Zachary, C. B., M. H. Allen, and D. M. MacDonald. 1985. In situ quantification of T-lymphocyte subsets and Langerhans cells in the inflammatory infiltrate of atopic eczema. Br. J. Dermatol. 112:149.
5. Bonecchi, R., G. Bianchi, P. P. Bordignon, D. D’Ambrosio, R. Lang, A. Borsatti, S. Sozzani, P. Allavena, P. A. Gray, A. Mantovani, and F. Sinigaglia. 1998. Differential expression of chemokine receptors and chemotactic responsiveness of type 1 T helper cells (Th1s) and Th2s. J. Exp. Med. 187:129.
6. Imai, T., M. Nagira, S. Takagi, M. Kakizaki, M. Nishimura, J. Wang, P. W. Gray, K. Matsushima, and O. Yoshie. 1999. Selective recruitment of CCR4-bearing Th2 cells toward antigen-presenting cells by the CC chemokines thymus and activation-regulated chemokine and macrophage-derived chemokine. Int. Immu-nol. 11:81.
7. Kim, C. H., L. Rott, E. J. Kunkel, M. C. Genovese, D. P. Andrew, L. Wu, and E. C. Butcher. 2001. Rules of chemokine receptor association with T cell polar-ization in vivo. J. Clin. Invest. 108:1331.
8. Sallusto, F., D. Lenig, C. R. Mackay, and A. Lanzavecchia. 1998. Flexible pro-grams of chemokine receptor expression on human polarized T helper 1 and 2 lymphocytes. J. Exp. Med. 187:875.
9. Nakatani, T., Y. Kaburagi, Y. Shimada, M. Inaoki, K. Takehara, N. Mukaida, and S. Sato. 2001. CCR4 memory CD4⫹T lymphocytes are increased in peripheral blood and lesional skin from patients with atopic dermatitis. J. Allergy Clin. Immunol. 107:353.
10. Wakugawa, M., K. Nakamura, T. Kakinuma, N. Onai, K. Matsushima, and K. Tamaki. 2001. CC chemokine receptor 4 expression on peripheral blood CD4⫹T cells reflects disease activity of atopic dermatitis. J. Invest. Dermatol. 117:188.
11. Yamamoto, J., Y. Adachi, Y. Onoue, Y. S. Adachi, Y. Okabe, T. Itazawa, M. Toyoda, T. Seki, M. Morohashi, K. Matsushima, and T. Miyawaki. 2000. Differential expression of the chemokine receptors by the Th1- and Th2-type effector populations within circulating CD4⫹T cells. J. Leukocyte Biol. 68:568. 12. Bunikowski, R., M. E. Mielke, H. Skarabis, M. Worm, I. Anagnostopoulos, G. Kolde, U. Wahn, and H. Renz. 2000. Evidence for a disease-promoting effect of Staphylococcus aureus-derived exotoxins in atopic dermatitis. J. Allergy Clin. Immunol. 105:814.
13. Ha, S. J., H. J. Lee, D. G. Byun, and J. W. Kim. 1998. Expression of T cell receptor V chain in lesional skin of atopic dermatitis. Acta Derm. Venereol. 78:424. 14. Neuber, K., C. Loliger, I. Kohler, and J. Ring. 1996. Preferential expression of
T-cell receptor V-chains in atopic eczema. Acta Derm. Venereol. 76:214. 15. Skov, L., J. V. Olsen, R. Giorno, P. M. Schlievert, O. Baadsgaard, and
D. Y. Leung. 2000. Application of staphylococcal enterotoxin B on normal and atopic skin induces up-regulation of T cells by a superantigen-mediated mecha-nism. J. Allergy Clin. Immunol. 105:820.
16. Bunikowski, R., M. Mielke, H. Skarabis, U. Herz, R. L. Bergmann, U. Wahn, and H. Renz. 1999. Prevalence and role of serum IgE antibodies to the Staphylococ-cus aureus-derived superantigens SEA and SEB in children with atopic derma-titis. J. Allergy Clin. Immunol. 103:119.
17. Leung, D. Y., R. Harbeck, P. Bina, R. F. Reiser, E. Yang, D. A. Norris, J. M. Hanifin, and H. A. Sampson. 1993. Presence of IgE antibodies to staphy-lococcal exotoxins on the skin of patients with atopic dermatitis: evidence for a new group of allergens. J. Clin. Invest. 92:1374.
18. Lin, Y. T., W. Y. Shau, L. F. Wang, Y. H. Yang, Y. W. Hwang, M. J. Tsai, P. N. Tsao, and B. L. Chiang. 2000. Comparison of serum specific IgE antibodies to staphylococcal enterotoxins between atopic children with and without atopic dermatitis. Allergy 55:641.
19. Nomura, I., K. Tanaka, H. Tomita, T. Katsunuma, Y. Ohya, N. Ikeda, T. Takeda, H. Saito, and A. Akasawa. 1999. Evaluation of the staphylococcal exotoxins and their specific IgE in childhood atopic dermatitis. J. Allergy Clin. Immunol. 104:441. 20. Tada, J., Y. Toi, H. Akiyama, J. Arata, and H. Kato. 1996. Presence of specific
IgE antibodies to staphylococcal enterotoxins in patients with atopic dermatitis. Eur. J. Dermatol. 6:552.
21. Kappler, J., B. Kotzin, L. Herron, E. W. Gelfand, R. D. Bigler, A. Boylston, S. Carrel, D. N. Posnett, Y. Choi, and P. Marrack. 1989. V-specific stimulation of human T cells by staphylococcal toxins. Science 244:811.
22. Weber, A. K., U. Wahn, and H. Renz. 2000. Superantigen-induced T cell death by apoptosis: analysis on a single cell level and effect of IFN-␥ and IL-4 treat-ment. Int. Arch. Allergy Immunol. 121:215.
23. Campbell, D. E., and A. S. Kemp. 1997. Proliferation and production of inter-feron-␥ (IFN-␥) and IL-4 in response to Staphylococcus aureus and staphylo-coccal superantigen in childhood atopic dermatitis. Clin. Exp. Immunol. 107:392. 24. Konig, B., K. Neuber, and W. Konig. 1995. Responsiveness of peripheral blood mononuclear cells from normal and atopic donors to microbial superantigens. Int. Arch. Allergy Immunol. 106:124.
25. Neuber, K., K. Steinrucke, and J. Ring. 1995. Staphylococcal enterotoxin B af-fects in vitro IgE synthesis, interferon-␥, interleukin-4 and interleukin-5 produc-tion in atopic eczema. Int. Arch. Allergy Immunol. 107:179.
26. Lynch, D. H., F. Ramsdell, and M. R. Alderson. 1995. Fas and FasL in the homeostatic regulation of immune responses. Immunol. Today 16:569. 27. Janssen, O., A. Stocker, R. Sanzenbacher, H. H. Oberg, M. A. Siddiqi, and
D. Kabelitz. 2000. Differential regulation of activation-induced cell death in in-dividual human T cell clones. Int. Arch. Allergy Immunol. 121:183.
28. Oberg, H. H., B. Lengl-Janssen, D. Kabelitz, and O. Janssen. 1997. Activation-induced T cell death: resistance or susceptibility correlate with cell surface Fas ligand expression and T helper phenotype. Cell. Immunol. 181:93.
29. Ramsdell, F., M. S. Seaman, R. E. Miller, K. S. Picha, M. K. Kennedy, and D. H. Lynch. 1994. Differential ability of Th1 and Th2 T cells to express Fas ligand and to undergo activation-induced cell death. Int. Immunol. 6:1545. 30. Varadhachary, A. S., S. N. Perdow, C. Hu, M. Ramanarayanan, and P. Salgame.
1997. Differential ability of T cell subsets to undergo activation-induced cell death. Proc. Natl. Acad. Sci. USA 94:5778.
31. Zhang, X., T. Brunner, L. Carter, R. W. Dutton, P. Rogers, L. Bradley, T. Sato, J. C. Reed, D. Green, and S. L. Swain. 1997. Unequal death in T helper cell (Th)1 and Th2 effectors: Th1, but not Th2, effectors undergo rapid Fas/FasL-mediated apoptosis. J. Exp. Med. 185:1837.
32. Aroeira, L. S., and A. Martinez. 1999. The role of IL-4 in the staphylococcal enterotoxin B-triggered immune response: increased susceptibility to shock and deletion of CD8V8⫹T cells in IL-4 knockout mice. Eur. J. Immunol. 29:1397. 33. Vella, A. T., S. Dow, T. A. Potter, J. Kappler, and P. Marrack. 1998. Cytokine-induced survival of activated T cells in vitro and in vivo. Proc. Natl. Acad. Sci. USA 95:3810.
34. Taskapan, M. O., and P. Kumar. 2000. Role of staphylococcal superantigens in atopic dermatitis: from colonization to inflammation. Ann. Allergy Asthma Im-munol. 84:3.
35. Hanifin, J., and G. Rajka. 1980. Diagnostic features of atopic dermatitis. Acta Derm. Venereol. 92:44.
36. Strange, P., L. Skov, S. Lisby, P. L. Nielsen, and O. Baadsgaard. 1996. Staph-ylococcal enterotoxin B applied on intact normal and intact atopic skin induces dermatitis. Arch. Dermatol. 132:27.
37. Akdis, M., H. U. Simon, L. Weigl, O. Kreyden, K. Blaser, and C. A. Akdis. 1999. Skin homing (cutaneous lymphocyte-associated antigen-positive) CD8⫹T cells respond to superantigen and contribute to eosinophilia and IgE production in atopic dermatitis. J. Immunol. 163:466.