利用簡單重複序列標記建立台灣主要稉稻品種的鑑別技術
謝廉一
1)、吳岱融
2)、胡凱康
3) 1)花蓮縣政府農業局技士 2)行政院農業委員會苗栗區農業改良場助理研究員 3)台灣大學農藝系副教授 (通訊作者) 台北市羅斯福路四段一號摘要:
為鑑別台灣地區的主要稉稻品種,本研究利用 46 個簡單重複序 列標記對25 個參試品種(系)進行分析。在經過聚合酵素連鎖反應及毛細 管電泳後,其中20 組引子在參試品種之中具有多型性,共 21 基因座, 平均對偶基因數為2.90、PIC 值為 0.38。除台稉 11 號及台稉 15 號的分 子標記多型性模式完全相同之外,其餘品種(系)均可被最少 6 個標記完全 鑑別。將品種鑑定的目標限定為具有相當栽培面積的18 個品種,則除了 台稉11 號與台稉 15 號之外,其餘品種(系)均可被最少 4 個標記完全鑑 別。配合選用長度範圍有差異的標記,將各標記PCR 增幅產物混合後進 行毛細管電泳分析,將可有效的降低分析所需的時間與分析成本。 關鍵詞:稉稻品種、品種鑑別、簡單重複序列、主成分分析前 言
水稻 (Oryza sativa L.) 是台灣地區主要的糧食作物,據中華民國九十五年農業 統計年報 (行政院農業委員會,2007) 統計民國 95 年台灣地區稉稻收穫面積達二十 二萬六千餘公頃。主要栽培品種有台稉8 號、台稉 9 號、台稉 11 號等十餘種。 為確保農戶稻穀品種及品質的一致性與兼顧其栽培管理上的便利,我國稻種繁 殖自民國48 年起即採取三級良種繁殖制度,並嚴格執行田間檢查與室內檢查作業, 以提高稻種純度。為確保稻種純度,行政院農業委員會 (以下簡稱農委會) 農糧署 種子檢查室 (前省農林廳種子檢查室) 在執行品種純度檢查過程,主要於田間檢查 時就地檢查各級採種田之植株外觀形態與生育特性,及於室內檢查時觀察所抽取樣 品之外觀性狀。然而由於多年來各育種單位沿用相近的育種材料,水稻品種間的差 異有逐漸減小的趨勢,加上種子外觀形態與生育特性容易受到外在環境的影響,因 收到日期:96 年 08 月 27 日而提高了以外觀性狀檢查異品種的困難度。
簡單重複序列 (simple sequence repeat, SSR) 是由 1 到 6 個長度鹼基對構成重複 單位 (motif) 的 DNA 序列,這種分子標記已廣泛地應用在遺傳分析與育種應用上 (Yang et al., 1994; Olufowote et al., 1997; Cho et al., 2000; Li et al., 2004),如水稻 (Wu and Tanksley, 1993; Panaud et al., 1996; Chen et al., 1997)、大麥 (Becker and Heun, 1995; Liu et al., 1996)、葡萄 (Thomas and Scott, 1993)、蕓苔 (Kresovich et al., 1995)、 玉米 (Senior and Heum, 1993)、大豆 (Akkaya et al., 1995) 及番茄 (Broun and Tanksley, 1996) 等作物。SSR 標記是屬於一種共顯性的 DNA 標記,它的對偶基因數 目變化很大,並可以簡便的利用聚合酵素連鎖反應 (Polymerase Chain Reaction, PCR) 檢驗 (Panaud et al., 1996)。SSR 標記的 PCR 增幅結果,會出現較主要條帶短少 1 至 4 個重複單位的殘跡條帶 (stutter band),由於殘跡條帶的增幅量依其短少重複單位 的數量而遞減,因此主要條帶的位置可由其殘跡條帶協助判讀 (Blair et al., 2002)。 在水稻基因組之中,已經發現大量且均勻分佈的簡單重複序列 (McCouch et al., 1997 )。Temnykh 等學者於 2001 年所建構的簡單重複序列圖譜中,包含超過 500 個 SSR 基因座,其中有 200 個 SSR 基因座是新增的。水稻為二倍體自交作物,在同一 個SSR 基因座中,通常只會顯現單套的、同質性的對偶基因,很少產生異質性的情 形,且對偶基因在稉稻的變化程度明顯的小於秈稻 (Blair et al. 2002)。 本研究的目的,是利用簡單重複序列標記,對台稉1 號至台稉 17 號,以及台農 67 號、台農 71 號 (益全香米) 等 25 個品種進行分析,期望找出一組可以鑑別出所 有參試品種(系)之引子對組合,並建立可快速鑑別不同品種的技術,供為相關單位 品種鑑別之參考。
材料與方法
一、材料 本研究使用25 個台灣稉稻品種(系),包含台稉 1-17 號,台中 189 號,台農 67、 70、71 號,高雄 139、141 號,台南 9 號及台南育 212 號等。種子來源分別為:農 委會農業試驗所國家作物種原中心 (台稉 1-13 號、台農 67 號、台南 9 號、台中 189 號、高雄 141 號)、農委會農業試驗所嘉義分所 (台農 70 號、台南育 212 號)、 農委會台中區農業改良場 (台稉 15 號)、農委會農糧署種子檢查室 (台稉 14、16、17 號、台農 71 號、高雄 139 號)。台農 67 號、台農 70 號、台中 189 號、台南 9 號 及高雄 139 號為部分台稉系列品種之親本。高雄 141 號及台南育 212 號為台稉 11 號、台稉15 號二品種之共同親本。其中台稉 2、4-6、8-10、11、13-17 號,台中 189 號,台農67、71 號,高雄 139、141 號等 18 個品種,在民國 90 年台灣地區的稻作 生產中占99%以上的面積 (臺南區農業改良場,2003)。 二、方法 各參試品種於91 年 5 月間種植,每品種取 5 至 10 粒種子進行浸種,發芽後將 種子移至塑膠杯中,生長初期置於具照明之室內,一個月後移至室外繼續生長,依 不同品種別適期分株收取3 單株葉片,以 CTAB 法 (Doyle et al., 1990) 抽取水稻葉 片DNA。經分光光度計 (U-2001 Spectrophotometer, Hitachi) 定量後,等量混合各品 種內3 單株的 DNA。
本研究所使用的SSR 標記,係選自 Temnykh 等 (2001) 的 SSR 連鎖群圖譜中, 多型性資訊含量 (polymorphism information contents, PIC) 值較高,且均勻分整個染 色體組的46 個 SSR 標記,選用 SSR 標記的名稱及其於染色體上的位置參見圖 1。 多型性測定之 PCR 反應使用熱循環反應器 (GeneAmp PCR System 9700, PE Applied Biosystems) 採 3 種黏合 (annealing) 溫度進行,反應條件為:94°C 2 分鐘; 94°C 1 分鐘、 (51°C、55°C 或 59°C) 30 秒、72°C 1 分鐘,循環 2 次;94°C 20 秒、 (51°C、55°C 或 59°C) 20 秒、72°C 30 秒,循環 30 次;72°C 30 分鐘,反應結束後產 物以4°C 保存。每一反應的總體積為 10 μL,其中包含 20 ng 水稻 DNA,引子各 0.4 μM,dNTPs 各 0.2 mM,0.6 U Taq DNA 聚合酶 (Biotools, B&M Labs)。PCR 產物以 4% Metaphor (BioWhittaker Molecular Applications) 膠片,0.5
×
TBE 緩衝液,電壓 180 V,11 小時進行電泳分析。具多型性 SSR 基因座各對偶基因的精確大小,以毛細管電泳 (PRISM 310 Genetic Analyzer, Applied Biosystems) 進行分析,為配合雷射激發螢光訊號的偵測, 另 行 訂 製 帶 有 FAM (5-carboxyfluorescein) 或 JOE (2', 7'-dimethoxy-4', 5'-dichloro-6-carboxyfluorescein) 螢光標記的 Forward 引子 (Merck, Taiwan)。PCR 反 應的配製與條件同上,但第2 階段的循環降低為 22 次。PCR 產物於稀釋 5 倍之後, 取1 μL 加上 24 μL Deionized Formamide 與 0.25 μL 片段大小標準 (GeneScan-500 Rox, Applied Biosystems),以 95˚C 處理 5 分鐘後於冰上急速冷卻。所收集到的螢光訊號 記錄,以GeneScan 3.7 軟體 (Applied Biosystems) 依據另一個螢光通道 (Rox) 中同
時讀取到的標準片段大小校正後,以鹼基對 (base pair, bp) 為單位判讀主要增幅片 段的長度。所有的DNA 樣品皆經過一次以上的 PCR/毛細管電泳分析,以確認對偶 基因大小判讀結果的可靠性。 三、資料分析 SSR 標記在不同品種(系)之間的多型性程度,依據 Ishii 與 McCouch (2000) 對 自交作物使用之PIC 計算式 2 1 1 n i ij j PIC p = = −
∑
,其中 pij是第 i 個基因座上第 j 個對 偶基因的頻度。 品種(系)之間的遺傳距離,定義為不相同基因座的數目在全部基因座數目中的 比 率 。 主 成 分 分 析 (Principal Coordination Analysis) 係 以 NTSYSpc2.2 (Exeter Software),將品種(系)間的距離矩陣,經過 Transformation 類別中的 Dcenter 雙向校 正之後,以Ordination 類別中的 Eigen 模組計算。 對於鑑別參試品種(系)所需最少 SSR 分子標記的數量,係以自行編寫的程式以 窮舉法列出限制標記數下所有可能的標記組合,並逐一驗證其鑑別參試品種(系)的 能力。結 果
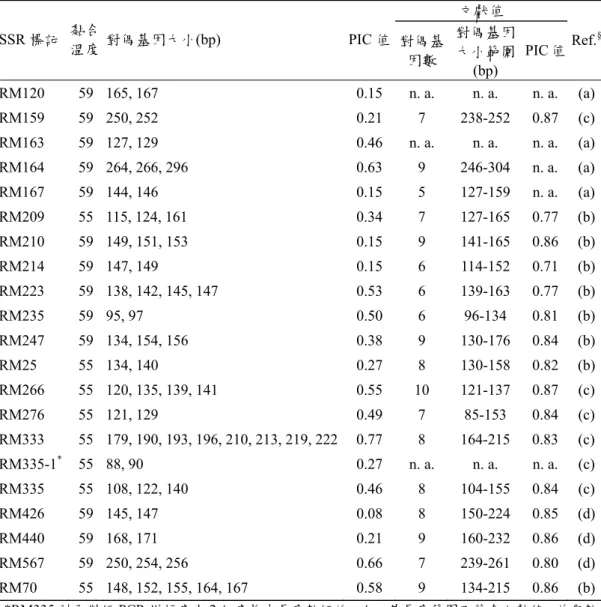
一、SSR 標記多型性 在所篩選的46 個 SSR 分子標記中,有 20 個可以在參試品種(系)之間表現出多 型性 (膠片電泳結果未顯示)。經毛細管電泳分析與判讀對偶基因大小,其最佳黏 合溫度、對偶基因大小與PIC 值見表 1。對偶基因大小在各次重複分析與判讀間的 差異皆小於0.5 bp,取最接近整數後的結果在各重複間無差異 (資料未顯示)。各基 因座對偶基因間的長度差異甚小,最小差異為2 bp 的有 14 個基因座,最小差異為 3 bp 的有 3 個基因座,最小差異在 4 bp (含)以上的有 4 個基因座。除 RM333 有 8 個對偶基因,與 RM70 有 5 個對偶基因之外,其餘標記對偶基因數的範圍在 2~4 之間;平均對偶基因數為 2.90。各標記的 PIC 值範圍由 0.08 (RM426)至 0.77 (RM333),平均值為 0.38。 各標記對偶基因的長度,除 RM335 之外,大體上皆落於文獻記載值的範圍之 內。RM335 在每一個參試的品種(系)皆可產生兩個增幅片段,較短的一組有 2 個對 偶基因,分別為88 bp 與 90 bp,其大小不在文獻值範圍之內;較長的一組有 3 個對偶基因,分別為108 bp、122 bp 與 140 bp,其範圍與文獻值相符。由於這兩組增 幅片段在各品種(系)之間並無一致性,應為兩個獨立分離的基因座。較短這組為一 未曾記載的新基因座,標示為RM335-1,其於染色體上的位置未知。 Chr. 1 Chr. 2 Chr. 3 Chr. 4 Chr. 5 Chr. 6 Chr. 7 Chr. 8 Chr. 9 Chr. 10 Chr. 11 Chr. 12 圖1. 用於篩選多型性的 46 個 SSR 分子標記,及其於 Temnykh 等 (2001) SSR 連鎖 群圖譜中的位置。以粗體顯示的為在台稉1-17 號,台中 189 號,台農 67、70、 71 號,高雄 139、141 號,台南 9 號及台南育 212 號等 25 品種(系)間具有多 型性者;CEN 為染色體中心節。
Fig 1. Forty-six SSR markers used in this research and their chromosomal locations based on linkage map of Temnykh et al. (2001). Markers that are polymorphic among Taikeng 1-17, Taichung 189, Tainung 67, 70 and 71, Kaohsiung 139 and 141, Tainan 9 and Tainan line 212 are in boldface. CEN denotes the centromere of each chromosome. RM25 52.2 CEN 10.8 RM223 17.5 RM210 9.8 38.3 CEN RM219 11.7 RM566 36.0 67.0 CEN 44.0 RM228 63.3 RM333 3.1 7.9 RM167 37.5 RM120 4.2 CEN 22.3 RM209 9.9 RM229 3.9 RM187 18.3 RM224 24.0 5.5 RM174 47.5 CEN 18.7 RM475 26.3 RM266 99.7 11.2 RM60 RM545 35.3 CEN 90.3 RM426 31.7 RM203 11.3 RM168 2.6 RM148 56.4 3.4 RM335 21.5 CEN 15.8 RM567 116.3 2.2 RM159 1.2 RM122 RM592 30.3 CEN 36.9 RM164 11.1 RM163 RM440 13.3 49.1 RM220 28.4 RM151 7.8 RM600 28.6 CEN 15.8 RM34 23.7 RM543 41.4 RM212 3.1 RM200 0.3 47.5 RM204 25.1 RM276 15.2 CEN 30.1 RM30 55.0 RM400 9.1 15.2 CEN 33.1 RM214 1.6 RM11 12.3 RM70 17.6 52.0 RM247 32.3 CEN 23.3 RM235 44.6 12.5
表一、 在參試 25 品種(系) (詳見圖 1) 之間具有多型性的 20 個 SSR 分子標記及其對 偶基因大小
Table 1. Amplified fragment length of 20 SSR markers which are polymorphic among 25 varieties and line listed in Fig 1
文獻值 SSR 標記 黏合溫度 對偶基因大小(bp) PIC 值 對偶基 因數 對偶基因 大小範圍 (bp) PIC 值 Ref. § RM120 59 165, 167 0.15 n. a. n. a. n. a. (a) RM159 59 250, 252 0.21 7 238-252 0.87 (c) RM163 59 127, 129 0.46 n. a. n. a. n. a. (a) RM164 59 264, 266, 296 0.63 9 246-304 n. a. (a) RM167 59 144, 146 0.15 5 127-159 n. a. (a) RM209 55 115, 124, 161 0.34 7 127-165 0.77 (b) RM210 59 149, 151, 153 0.15 9 141-165 0.86 (b) RM214 59 147, 149 0.15 6 114-152 0.71 (b) RM223 59 138, 142, 145, 147 0.53 6 139-163 0.77 (b) RM235 59 95, 97 0.50 6 96-134 0.81 (b) RM247 59 134, 154, 156 0.38 9 130-176 0.84 (b) RM25 55 134, 140 0.27 8 130-158 0.82 (b) RM266 55 120, 135, 139, 141 0.55 10 121-137 0.87 (c) RM276 55 121, 129 0.49 7 85-153 0.84 (c) RM333 55 179, 190, 193, 196, 210, 213, 219, 222 0.77 8 164-215 0.83 (c) RM335-1* 55 88, 90 0.27 n. a. n. a. n. a. (c) RM335 55 108, 122, 140 0.46 8 104-155 0.84 (c) RM426 59 145, 147 0.08 8 150-224 0.85 (d) RM440 59 168, 171 0.21 9 160-232 0.86 (d) RM567 59 250, 254, 256 0.66 7 239-261 0.80 (d) RM70 55 148, 152, 155, 164, 167 0.58 9 134-215 0.86 (b) *RM335 引子對經 PCR 增幅產生 2 組產物中長度較短的一組,其長度範圍不符合文獻值,並與較 長、長度範圍符合文獻值的一組獨立表現,標示為RM335-1。
*One of the two groups of fragments amplified from RM335 primer set. Due to its deviate (shorter) size range than the one listed on Temnykh et al., 2000 and apparent independence to the regular RM335 marker, this group is recognized as another locus and labeled as RM335-1.
二、參試品種(系)之遺傳關係 品種(系)間除台稉 11 號與台稉 15 號完全沒有差異之外,在 21 個 (含 RM335-1) 具多型性的SSR 標記中,品種(系)間皆有一個基因座以上的差異,而以台農 70 號與 台南育212 號間的 15 個差異為最大。將品種間差異標記數轉換為遺傳距離,進行主 成分分析的結果顯示,前3 個主成分的解釋力各為 32.6%, 24.3%及 15.2%,前 3 主 成分的累計解釋力為72.2%,品種(系)間前 2 個主成分的分佈如圖 2。在圖 2 之中, 台稉 11 號與台稉 15 號的位置重疊,顯示出以本研究的所篩選得到的 21 個 (包含 RM335-1) SSR 標記進行鑑別,將無法區別此二種品種。 主成分分析的結果亦部分顯示出參試品種(系)間的遺傳關係,如台稉 17 號選自 於台農70 號與 Milyang79 雜交後,回交台農 70 號 2 次的衍生後裔;台農 67 號為台 稉13 號的親本,高雄 139 號為高雄 141 號之親本。台稉 11 號與台稉 15 號皆選自台 南育212 號與高雄 141 號雜交的衍生 F4系統,而其重疊的落點,即位於台南育212 號與高雄141 號之間。台稉 2 號之譜系為 “台農 67 號/嘉農育 252 號//台南 9 號” , 其於圖2 的位置,也落於台農 67 號與台南 9 號之間的區間。 參試品種(系)之間具有親緣關係的還有:台南 9 號為台南育 212 號的親本,台 農67 號為台農 70 號與台中 189 號的親本,台中 189 號為台稉 3 號與台稉 7 號的親 本,台農70 號為台稉 10 號的親本,台稉 2 號為台稉 8 號與台稉 16 號的親本,台稉 4 號為台農 71 號的親本。由於這些雜交的另一親本未包含於本研究之中,因此無法 推論其親緣關係與分子標記遺傳距離間之關係。 三、最小鑑別集合 除台稉11 號與台稉 15 號之間的差異無法鑑別之外,鑑別 25 個參試品種(系)所 需最小鑑別集合的標記數為 6 個,而可能的組合共有 12 種 (資料未列出)。由於參 試品種(系)中包含已經不再使用的品種,或品種所選自的雜交親本,鑑別這些品種(系) 在實務上的用途不大。若將鑑別的目標限定為仍有相當栽培面積的18 個品種 (見材 料),則最小鑑別集合的標記數減為 4 個,可能的組合方式共有 10 種,所包含的標 記 (括弧內為其在 10 組中出現的頻度) 為 RM333 (10)、RM276 (8)、RM567 (6)、RM70 (4)、RM235 (3)、RM164 (3)、RM163 (3)、RM159 (2)與 RM266 (1)。除 RM159 之外, 這些標記的PIC 值皆相當接近 0.5 或更高。 為簡化分析的程序,減少毛細管電泳分析的次數,從這10 種標記的組合中,選 取其標記的大小有明顯區隔者,在PCR 增幅之後,將增幅產物合併分析,如 RM235
(95-97 bp),RM276 (121-129 bp),RM333 (179-222 bp)與 RM567 (250-256 bp),合併 分析的結果見圖3。
圖2. 參試 25 品種(系)遺傳距離主成分分析 (TK:台稉;TNG:台農;TN:台南; TNY:台南育;TC:台中;KH:高雄),第一主成分與第二主成分的解釋量 分別為32.6%及 24.3%。圖中台稉 11 號與台稉 15 號重疊在一起。
Fig 2. Principle coordinate analysis result of 25 cultivars/lines (TK: Taikeng; TNG: Tainung; TN: Tainan; TNY: Tainan Yu; TC: Taichung; KH: Kaohsiung). The first and second dimensions account for 32.6% and 24.3% of total variability
respectively. TK11 and TK15 were overlapped.
PCO 1 PCO 2 TK1 TK2 TK3 TK4 TK5 TK6 TK7 TK8 TK9 TK10 TK11 TK12 TK13 TK14 TK15 TK16 TK17 TNG67 TNG70 TNG71 TC189 TN9 KH139 KH141 TNY212
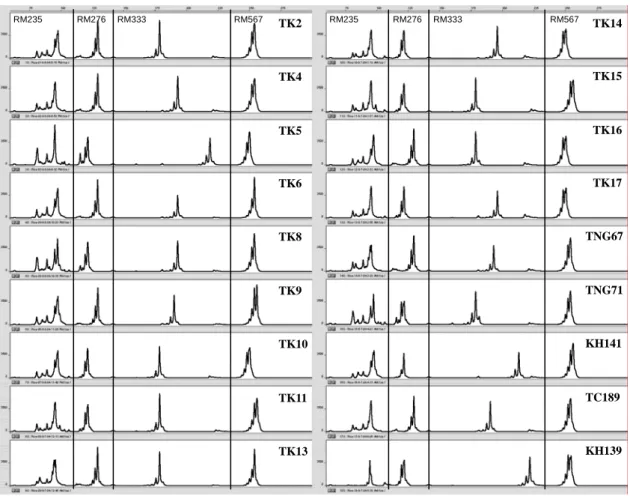
圖3. 可用於鑑別台稉 2、4-6、8-10、11、13-17 號,台中 189 號,台農 67、71 號, 高雄139、141 號等 18 個主要栽培品種的最小鑑別標記集合之一,RM235、 RM276、RM333 及 RM567 等標記 PCR 增幅產物混合進行毛細管電泳分析結 果,圖中縱軸為螢光強度,橫軸為經過校正後的片段大小,以鹼基對為單位, 品種名稱縮寫同圖2。
Fig 3. Capillary electrophoresis results of mixed amplified fragments from RM235, RM276, RM333 and RM567 as one of the 10 minimum sets of SSR markers that may identify 18 major rice varieties including Taikeng 2, 4-6, 8-10, 11, 13-17, Taichung 189, Tainung 67 and 71, Kaohsiung 139 and 141. Vertical axis is the intensity of florescence, horizontal axis is the corrected fragment size in base-pair, abbreviation for variety names are the same as Fig 2.
RM235 RM276 RM333 RM567 RM235 RM276 RM333 RM567 TK2 TK4 TK5 TK6 TK8 TK9 TK10 TK11 TK13 TK14 TK15 TK16 TK17 TNG67 TNG71 KH141 TC189 KH139
討 論
本研究依據文獻選出對偶基因數較多、或PIC 值較高的 46 個 SSR 標記參與篩 選,在台灣地區25 稉稻品種(系)之間,只有 20 個 SSR 標記表現出多型性,共得到 21 個基因座。各基因座對偶基因數之平均值為 2.90,遠低於各標記文獻值的平均 7.67 (Wu and Tanksley, 1993; Chen et al., 1997; Temnykh et al., 2000; Temnykh et al., 2001)。也由於對偶基因數較少,使得各標記的 PIC 值亦較低,平均值為 0.38,亦遠 低於各標記文獻值的平均0.83,此與 Blair et al. (2002) 認為稉稻多型性較秈稻低的 觀察吻合。不但如此,從品種的譜系資料亦顯示出參試的稉稻品種間,透過共同的 親本,有相當高程度的親緣關係,這也可能使得各品種間的遺傳組成較為接近,而 降低品種間的多樣性。 在本研究中,品種(系)之間的遺傳距離不採用一般在隨機增幅片段多型性分析 (random amplified polymorphic DNAs; RAPD) 所常用的 Jaccard similarity index (Jaccard, 1908),是因為 Jaccard similarity index 適用於以 “存在/不存在” 形式表現的 2 元資料 (binary data),而 SSR 基因座所表現的方式為多重狀態 (multi-state),且在 本研究分析的品種(系)中,並未表現 null 對偶基因、或異質結合的現象。考慮水稻 的育種過程,假設其雜交親本 (同質結合體) 之間有 n 個基因座的差異,在經過多 世代的選拔及自交之後,這些分離的基因座終將固定於兩親本之一的形式,若選出 新品種與親本 1 間有
n
1個基因座的差異,與親本 2 間有n
2個基因座的差異,則 1 2n
+
n
=
n
。由此可知以差異的基因座數目來定義遺傳距離,符合距離之基本定義。 在本研究中採用不相同基因座的數目在全部基因座數目中的比率來定義遺傳距離, 可使得遺傳距離之範圍限定於0 與 1 之間,以便與其他研究的結果比較。需注意的 是,這樣的定義與計算方法係以基因座為單位,僅能適用於全部都是同質結合體的 分析。若參試的基因型有部分表現為異質結合體,則應改用以對偶基因為單位的測 量方式。 對於兩雜交親本皆包含於分析的品種,如台稉11 號與台稉 15 (台南育 212 號/ 高雄141 號),或多數親本參試的品種,如台稉 2 號 (台農 67 號/嘉農育 252 號//台南 9 號),由 SSR 基因座之差異計算所得各品種與其親本間之遺傳距離,大致符合譜系 的紀錄。只有單一親本參試的品種中,台稉17 號 (台農 70 號/ Milyang79//台農 70 號///台農 70 號) 與其親本台農 70 號間之距離亦符合遺傳關係。其餘品種與其參試親本的關係,由於其另一親本與參試親本間的遺傳距離未知,因此無法判斷其表現 是否符合遺傳關係。但依據已驗證的關係,推測與其親本距離近的品種,可能來自 於親緣關係較近的親本組合。 SSR 標記由於各基因座具有複對偶基因的特性,使其對於親緣關係相當接近的 25 個參試稉稻品種(系)具有充分的鑑別能力,且最少只需 6 個 SSR 標記即可完全鑑 別。但SSR 標記的對偶基因間差異甚為微小,有三分之二的多型性基因座中,對偶 基因間的最小差異僅為2 bp,使得 SSR 標記分析必須藉助於高解析度的毛細管電泳 或定序膠片電泳技術,並配合與PCR 增幅產物一併分析的分子大小標準進行校正, 才能獲得準確穩定的結果,因而降低此一技術在品種鑑定上的實用性。本研究的結 果顯示,若將品種鑑定的目標依據各品種的使用程度,合理的限定於一較小範圍(如 材料中所指定的18 個品種),則最少只需 4 個 SSR 標記即可完全鑑別。配合選用長 度範圍有差異的標記,將各標記PCR 增幅產物混合後進行毛細管電泳分析,將可有 效的降低本技術所需的時間與分析成本,使得此技術可實用於台灣的稉稻品種鑑別 之用。
誌 謝
本文為第一作者碩士論文之部分資料。感謝國家作物種原中心、行政院農業委 員會農糧署種子檢查室、農業試驗所嘉義分所及台中區農業改良場提供實驗材料。 本研究承行政院農業委員會經費 (89 科技-1.1-糧 69(13)) 補助,僅此致謝。參考文獻
行政院農業委員會編。2007。中華民國九十五年農業統計年報。行政院農業委員會。 臺南區農業改良場編。2003。稻作改良年報 (民國九十年)。行政院農業委員會臺南 區農業改良場。Akkaya, M. S., R. C. Shoemaker, J. E. Specht, A. A. Bhagwat, and B. B. Cregan. 1995. Integration of simple sequence repeat DNA markers into a soybean linkage map. Crop Sci. 35:1439–1445.
Mol Biol 27:835–845.
Blair, M. W., V. Hedetale, and S. R. McCouch. 2002. Fluorescent-labeled microsatellite panels useful for detecting allelic diversity in cultivated rice (Oryza sativa L.). Theor. Appl. Genet. 105:449-457.
Broun, P. and S. D. Tanksley. 1996. Characterization and genetic mapping of simple repeat sequences in the tomato genome. Mol. Gen. Genet. 250:39–49.
Chen, X., S. Temnykh, Y. Xu, Y. G. Cho, and S. R. McCouch. 1997. Development of a microsatellite framework map providing gemone-wide coverage in rice (Oryza sativa L.). Theor. Appl. Genet. 95:553-567.
Cho, Y. G., T. Ishii, S. Temnykh, X. Chen, L. Lipovich, W. D. Park, N. Ayres, S. Cartinhour, and S. R. McCouch. 2000. Diversity of microsatellites derived from genomic libraries and GenBank sequences in rice (Oryza sativa L.). Theor. Appl. Genet. 100:713-722.
Doyle, J. J. and J. L. Doyle. 1990. Isolation of plant DNA from flash tissue. Focus 12:13-15.
Ishii, T. and S. R. McCouch. 2000. Microsatellites and microsynteny in the chloroplast genomes of Oryza and eight other Gramineae species. Theor. Appl. Genet. 100:1257-1266.
Jaccard, P. 1908. Nouvelles rescherches sur la distribution florale. Bull. Soc. Vaud. Sci. Nat., 44:223-270.
Kresovich, S., A. K. Szewc-McFadden, S. M. Bliek, and J. R. McFerson. 1995. Abundance and characterization of simple-sequence repeats (SSRs) isolated from a size-fractionated genomic library of Brassica napus L. (rapeseed). Theor. Appl. Genet. 91:206-211.
Li, C., Y. Zhang, K. Ying, X. Liang, and B. Han. 2004. Sequence variations of simple sequence repeats on chomosome-4 in two subspecies of the Asian cultivated rice. Theor. Appl. Genet. 108:392-400.
Liu, Z. W., R. M. Biyashev, M. A. Saghai Maroof. 1996. Development of simple sequence repeat DNA markers and their integration into a barley linkage map. Theor. Appl. Genet. 93:869–876.
McCouch, S. R., X. Chen, O. Panaud, S. Temnykh, Y. Xu, Y. G. Cho, N. Huang, T. Ishii, and M. Blair. 1997. Microsatellite marker development, mapping and applications in rice genetics and breeding. Plant. Mol. Biol. 35:89-99.
Olufowote, J. O., Y. Xu, X. Chen, W. D. Park, H. M. Beachell, R. H. Dilday, M. Goto, and S. R. McCouch. 1997. Comparative evaluation of within-cultivar variation of rice (Oryza sativa L.) using microsatellite and RFLP markers. Genome 38:1170-1176.
Panaud, O., X. Chen, and S. R. McCouch. 1996. Development of microsatellite markers and characterization of simple sequence length polymorphism (SSLP) in rice (Oryza sativa L.). Mol. Gen. Genet. 252:597-607.
Senior, M. L. and M. Heun. 1993. Mapping maize microsatellites and polymerase chain reaction confirmation of the targeted repeats using a CT primer. Genome 36:884–889.
Temnykh, S., W. D. Park, N. Ayres, S. Cartinhour, N. Hauck, L. Lipovich, Y. G. Cho, T. Ishii, and S. R. McCouch. 2000. Mapping and genome organization of microsatellite sequences in rice (Oryza sativa L.). Theor. Appl. Genet. 100:697-712.
Temnykh, S., G. DeClerk, A. Lukashova, L. Lipovich, S. Cartinhour, and S. McCouch. 2001. Computational and experimental analysis of microsatellites in rice (Oryza sativa L.): frequency, length variation, transposon associations, and genetic marker potential. Genome Res. 11:1441-1452.
Thomas, M. R. and N. S. Scott. 1993. Microsatellite repeats in grapevine reveal DNA polymorphisms when analyzed as sequence-tagged sites (STSs). Theor. Appl. Genet. 86: 986–990.
Wu, K. S. and S. D. Tanksley. 1993. Abundance, polymorphism and genetic mapping of microsatellites in rice. Mol. Gen. Genet. 241:225-235.
Yang, G. P., M. A. Saghai Maroof, C. G. Xu, Q. Zhang, and R. M. Biyashev. 1994. Comparative analysis of microsatellite DNA polymorphism in landraces and cultivars of rice. Mol. Gen. Genet. 245:187-194.
Variety identification among major Japonica rice cultivars of
Taiwan based on simple sequence repeat markers
Lien-Yi Hsieh
1)、Dai-Rong Wu
2)、Kae-Kang Hwu
3) 1)Associate Technical Specialist, Agriculture Bureau, Hualien County.2)Assistant Research Fellow, Miaoli District Agricultural Research and Extension station, Council of Agriculture, Executive Yuan.
3)Associate professor (corresponding author), Department of Agronomy, National Taiwan University. No 1, Sec 4, Roosevelt Rood, 106 Taipei, Taiwan, ROC
Summary:
In order to identify the major Japonica type rice cultivars grown in Taiwan, 46 simple sequence repeat (SSR) markers were screened against 25 rice cultivars. After polymerase chain reaction (PCR) and capillary electrophoresis, 20 SSR markers showed polymorphism among cultivars tested. In total, there were 21 polymorphic loci, with average 2.90 alleles per locus, averaged Polymorphic Information Content is 0.38. All cultivars can be distinguished based on polymorphic SSR markers, except Taikeng No. 11 and Taikeng No. 15. The minimum set of markers required to distinguish all 25 entries composed of 6 markers. If the varieties subject to identification are restricted to 18 major cultivars which are still in use, only 4 markers are needed to identify all but the two previously mentioned cultivars. By choosing markers with non-overlapping allele sizes, PCR products may be analyzed by multiplex capillary electrophoresis and thus greatly reduce the time and cost to conduct the analysis.Key words: Japonica rice cultivar, variety identification, simple sequence repeat, principal coordinate analysis