Combined rpoB duplex PCR and hsp65 PCR-Restriction Fragment Length Polymorphism with Capillary Electrophoresis is an Effective Algorithmand Convenient of Identification of Mycobacterial species from Clinical Isolates Chen-Cheng Huang1,9, Jiann-Hwa Chen2, Shiau-Ting Hu3,Chien-Shun Chiou4,
Wei-Chang Huang5, Jeng-Yuan Hsu5, Jang-Jih Lu6,9, and Gwan-Han Shen5,7,8
Running title: Combined rpoB and hsp65 PRA for mycobacteria identification
Corresponding author: Gwan-Han Shen
Address: No. 160, Sec. 3, Chung-Kang Rd., Taichung, Taiwan, R.O.C 40705 Tel: +886-4-23592525 ext. 3201 E-mail: [email protected]
ABSTRACT Background
Identification of mycobacteria based on PCR-restriction assay (PRA) is very rapid and simple but may also lead to misidentification due to similarities in a number of band sizes critical for discriminating among species. Capillary electrophoresis can provide computer-aided band discrimination. Herein is a novel algorithm developed for identification of mycobacteria by combined rpoB DPRA (duplex PRA) and hsp65 PCR-restriction enzyme analysis (PRA) with capillary electrophoresis.
Results
Three hundred and seventy-six AFB smear-positive BACTEC cultures, including 200 Mycobacterium tuberculosis (MTB) and 176 non-tuberculous mycobacteria (NTM), were analyzed. By combined hsp65 and rpoB DPRA, the accuracy rate was 100% (200 isolates) in MTB and 91.4% (161 isolates) in NTM. Among the discordant results (8.6%) in NTM, one isolate of M. mucogenicum and the other isolate of M. flavescens were found as new sub-types in hsp65 PRA.
Conclusions
This effective and novel identification algorithm by combined rpoB DPRA and hsp65 PRA with capillary electrophoresis can rapidly identify mycobacteria and even find new sub-types in hsp65 PRA. In addition, it also is complementary to 16S rDNA sequencing.
Background
Detection and mycobacteria identification in clinical specimens is a key issue in the therapy of pulmonary diseases because misidentification will lead to inappropriate treatment. Traditionally, mycobacterial species is identified based on growth rate, presence or absence of pigmentation, and biochemical assays of the isolates recovered from the specimens, which usually takes 1 to 2 months. The biochemical assays are time-consuming and labor-intensive, while assays for non-tuberculous mycobacteria (NTM) species can be poorly reproducible and have ambiguously interpreted results [1-2].
In contrast, molecular identification, notably PCR-restriction enzyme analysis (PRA), is rapid and simple. The hsp65 PRA method, developed by Telenti et al. in 1993, is a popular DNA-based method for mycobacteria identification [3]. Using hsp65 PRA, Wong et al. [4] reported 100% sensitivity and specificity in identifying Mycobacterium tuberculosis complex but only 74.5% sensitivity in identifying NTM species. This misidentification may occur due to similarities in a number of band sizes critical for species discrimination [3] and also due to lack of knowledge of all existing PRA profiles, especially among species that are very heterogeneous, such as M. gordonae, M. scrofulaceum, and M. terrae complex. Recently, capillary electrophoresis [5-9] provide more precise band discrimination with the aid of the computer rather than with the naked eye only.
Previously, we developed an algorithm for mycobacterial species identification from AFB smear-positive BACTEC tubes by combining the rpoB DPRA method described by Kim et al. [10] with key phenotypic characters of mycobacteria recovered from the tubes. By using the rpoB duplex PCR, we differentiate Mycobacterium tuberculosis (MTB) from NTM with a 235- base pair (bp) and a
136-bp PCR amplicons in AFB smear-positive BACTEC cultures [11]. The136-bp rpoB duplex PCR amplicon was further digested with MspI and HaeIII (rpoB DPRA) to divide NTM species into eight easily distinguishable groups (A-H) described by Kim et al. [10]. By two phenotypic characters (growth rate and photoreactivity on pigment production) and two simple biochemical assays (nitrate reduction test and Tween 80 hydrolysis test) [11], the mycobacterial species identification could be made. However, this algorithm still takes three weeks for sub-culture and biochemical tests.
In this study, we developed a rapid and effective algorithm for identification of mycobacteria by combined rpoB DPRA and hsp65 PRA with capillary electrophoresis.
METHODS
Mycobacterial isolates
Fourteen mycobacterium reference strains, including one MTB and 13 NTM strains, were studied. From January to July 2007, 376900 clinical respiratory specimens such as sputum, broncho-alveolar lavage, and aspirated secretion from endotracheal tubes were collected from Taichung Veterans General Hospital. The respiratory specimens were processed by digestion, an N-acetyl-L-cysteine-NaOH decontamination procedure, centrifugal concentration, and sputum dissolving agents [12]. The processed specimens or the concentrated specimens were inoculated into MGIT culture tubes and incubated in the BACTEC 960 instrument at 37°C until a positive signal appeared. Deposits from positive signal culture tubes (positive BACTEC cultures) were smeared on glass slides and acid-fast stained by Kinyoun method for screening of acid fast staining bacilli [12]. Mycobacteria in the positive BACTEC cultures were isolated and identified by conventional methods [13-14] .
Mycobacterial isolates identification by conventional methods
All 376 isolates from cultures in the AFB smear-positive BACTEC tubes were streaked on Löwenstein-Jensen (LJ) slants and incubated at 37°C under 5% CO2 atmosphere. Colonies on the LJ slants were used for species identification by conventional culture and biochemical methods [13-14]. This included growth rates, photoreactivity for pigment production, morphology in microcolonies on LJ slants and biochemical tests, including the following: nitrate reduction, arylsulfatase (3 days), Tween 80 hydrolysis (3 days), urease, semiquantitative catalase, tolerance to 5% NaCl and niacin production.
Genomic DNA extraction
Mycobacterial DNA was extracted from positive BACTEC cultures and the DTB specimen processing kit (Becton Dickinson) was used as previously described and according to the manufacturer’s instructions [11].
rpoB duplex PCR and rpoB DPRA
The rpoB duplex PCR were performed using genomic DNA as template and primer pairs Tbc1 (5’ CGTACGGTCGGCGAGCTGATCCAA 3’) -TbcR5 (5’
CCACCAGTCGGCGCTTGTGGGTCAA 3’) and M5 (5’
GGAGCGGATGACCACCCAGGACGTC 3’) -RM3 (5’
CAGCGGGTTGTTCTGGTCCATGAAC 3’) described by Kim et al. [10]. A 235-bp DNA PCR amplicon from MTB and a 136-bp DNA PCR amplicon from NTM were specifically amplified, respectively [10], and these two amplification products were analyzed by electrophoresis on a 2% agarose gel (Seakem LE agarose, Cambrex, East Rutherford, NJ, USA).
For rpoB DPRA, the 136-bp DNA PCR amplicon was further digested with MspI and HaeIII after completing duplex PCR and analyzed by electrophoresis on a 3% agarose gel (NuSieve 3:1 agarose, Cambrex) or capillary electrophoresis (eGene)
and the rpoB RFLP pattern were compared to eight groups described by Kim et al. [10]. The eight NTM reference strains (M. abscessus ATCC 19977, M. avium subsp. avium ATCC 25291, M. kansasii ATCC 12479, M. terrae ATCC 15755, M. szulgai ATCC 29716, M. intracellulare ATCC 13950, M. scrofulaceum ATCC 19981, M. xenopi ATCC 19250) from each rpoB group (A-H) were subjected to rpoB DPRA by capillary electrophoresis (eGene).
hsp65 PCR and hsp65PRA
The hsp65 PCR were performed using genomic DNA as template and primer Tb11(5’-ACC AAC GAT GGT GTG TCC-3’) and Tb12 (5’-CTT GTC GAA CCG CAT ACC CT-3’) described by Telenti et al. (25). A 439 bp DNA hsp65 PCR amplicon was specifically amplified from DNA extraction and the amplification product was analyzed by electrophoresis on a 2% agarose gel (Seakem LE agarose, Cambrex, East Rutherford, NJ, USA).
For hsp65 PRA, the 439 bp DNA hsp65 PCR amplicon was further digested with BstEII and HaeIII after completing hsp65 PCR and analyzed by electrophoresis on a 3% agarose gel (NuSieve 3:1 agarose, Cambrex) or capillary electrophoresis (eGene). The sizes of the restriction fragment by hsp65 PRA were compared to those reported on the PRASITE database (http://app.chuv.ch/prasite/index.html). Thirteen ATCC NTM reference strains and one MTB reference strain were subjected to hsp65 PRA by capillary electrophoresis (eGene).
eGene capillary electrophoresis
The restriction fragment sizes were estimated by the naked eye or by eGene capillary electrophoresis according to the manufacturer’s instructions using size marker ranging from 100 bp to 3 kb and alignment markers ranging from 15 bp to 3 kb.
The mycobacterial identification flow chart was shown in Figure 1. 16S rDNA sequencing
The 16S rDNA sequencing of mycobacterial DNA as the reference standard method for mycobacterial species identification was carried out using primer pair 8FPL (5’AGTTTGATCCTGGCTCAG 3’) and 1492 (5’GGTTACCTTGTTACGACT T 3’) described by Turenne et al. [15]. Species identification was performed by comparing the 16S rDNA sequence with the most related sequences obtained from the GenBank.
RESULTS
Mycobacterial identification
There were 376 AFB smear-positive BACTEC culture tubes (positive BACTEC cultures), including 200 MTB and 176 NTM-containing BACTEC cultures (another 20 contaminated bacteria with MGIT positive but AFB cultural smear negative termed as contamination had been excluded in the later evaluation).
By rpoB duplex PCR, all of the 200 MTB-containing BACTEC cultures and the 176 NTM-containing BACTEC cultures demonstrated the 235-bp and 136-bp PCR amplicons specific for MTB and NTM, respectively. Species identification were reported according to the flow chart shown in Figure 1.
Concordant results by combining rpoB DPRA and hsp65 PRA
By combining rpoB DPRA and hsp65 PRA with computer-aided capillary electrophoresis, the accuracy rate was 100% (200/200) in MTB and 91.4% (161/176) in NTM (Table 1).
hsp65 RFLP and rpoB RFLP pattern compared to the 16S rDNA sequence
rpoB RFLP pattern hsp65 RFLP pattern 16S rDNA sequence identification conventional biochemistry identification BACTEC culture number concordance rate
T M. tuberculosis type 1 M. tuberculosis M. tuberculosis 200 100%(200/ 200)
NTM NTM NTM 161 91.4%(161
/176)
A M. abscessus type1 M. abscessus M. abscessus 29
A M. abscessus type 2 M. abscessus M. abscessus 41
A M. fortuitum type 1 M. fortuitum M. fortuitum 33
A M. fortuitum type 2 M. fortuitum M. fortuitum 2
A M. peregrinum type 1 M. peregrinum M. fortuitum* 5
A M. peregrinum type 2 M. peregrinum M. fortuitum* 8
A M. peregrinum type 3 M. peregrinum M. fortuitum* 1
A M. chelonae type 1 M. chelonae M. chelonae 1
A
M. mucogenicum type 1 M. mucogenicum
M.
mucogenicum 2
A M. smegmatis type 1 M. smegmatis M. smegmatis 2
B M. avium subsp. avium
type 2 M. avium subsp. avium
M. avium
complex** 2
D M. kansasii type 1 M. kansasii M. kansasii 6
D M. kansasii type 2 M. kansasii M. kansasii 1
D M. kansasii type 6 M. kansasii M. kansasii 1
D M. triviale type 1 M. triviale M. triviale 1
F M. malmoense type 1 M. malmoense M. malmoense 2
F M. szulgai type 1 M. szulgai M. szulgai 1
F M. interjectum type 1 M interjectum M interjectum 1
G M. intracellulare type 1 M. intracellulare M. avium
complex** 14
G M. gordonae type 1 M. gordonae M. gordonae 6
G M. gordonae type 2 M. gordonae M. gordonae 1
G M. gordonae type 5 M. gordonae M. gordonae 1
Total 361
* The M.peregrinum was identified as M.fortuitum by conventional biochemical method
complex by conventional biochemical method
Discordant results by combining rpoB DPRA and hsp65 PRA
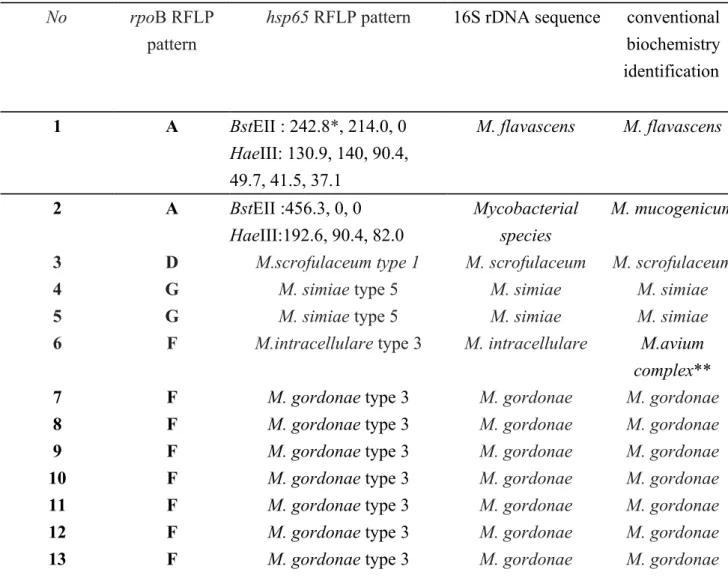
There were 15 isolates (8.6%) of NTM with discordant results in the rpoB DPRA and hsp65 PRA (Table 2). The two isolates, M. mucogenicum (A group) and M. flavescens (A group) represented new patterns not available in the databases of hsp65 PRA. The species identification of the two isolates was finally made by 16S rDNA sequencing and might be new sub-types in hsp65 PRA.
Table 2Fifteen isolates of NTM species with discordant results in rpoB RFLP and hsp65 RFLP pattern and species identification as determined by the 16S rDNA sequence
No rpoB RFLP
pattern
hsp65 RFLP pattern 16S rDNA sequence conventional biochemistry identification 1 A BstEII : 242.8*, 214.0, 0 HaeIII: 130.9, 140, 90.4, 49.7, 41.5, 37.1 M. flavascens M. flavascens 2 A BstEII :456.3, 0, 0 HaeIII:192.6, 90.4, 82.0 Mycobacterial species M. mucogenicum 3 D M.scrofulaceum type 1 M. scrofulaceum M. scrofulaceum
4 G M. simiae type 5 M. simiae M. simiae
5 G M. simiae type 5 M. simiae M. simiae
6 F M.intracellulare type 3 M. intracellulare M.avium complex**
7 F M. gordonae type 3 M. gordonae M. gordonae
8 F M. gordonae type 3 M. gordonae M. gordonae
9 F M. gordonae type 3 M. gordonae M. gordonae
10 F M. gordonae type 3 M. gordonae M. gordonae
11 F M. gordonae type 3 M. gordonae M. gordonae
12 F M. gordonae type 3 M. gordonae M. gordonae
14 F M. gordonae type 4 M. gordonae M. gordonae
15 F M. gordonae type 4 M. gordonae M. gordonae
*fragment size by the capillary electrophoresis
**The M. avium subsp. avium and M.intracellulare were identified as M.avium complex by conventional biochemical method
Development of a modified species identification algorithm
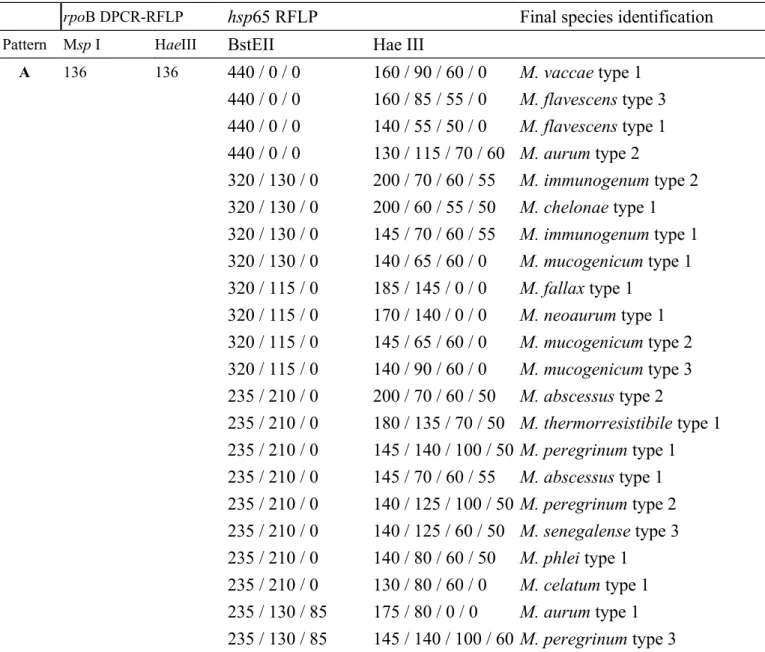
From the finding of Table 1 and 2 and using the mycobacterial identification flow chart, a modified species identification algorithm was developed by combined rpoB duplex PCR [10] and hsp65 PRA [3] including the most common 90 patterns of 40 species in Table 3. In this algorithm (Table 3), we added the M. gordonae types 3 and 4 into the F group. Thus, the accuracy rate of NTM by combined the two methods can reach 96.6% (170/176).
Table 3 A modified species identification algorithm by combined rpoB duplex PCR and hsp65 PCR-Restriction Fragment Length Polymorphism analysis
rpoB DPCR-RFLP hsp65 RFLP Final species identification
Pattern Msp I HaeIII BstEII Hae III
A 136 136 440 / 0 / 0 160 / 90 / 60 / 0 M. vaccae type 1 440 / 0 / 0 160 / 85 / 55 / 0 M. flavescens type 3 440 / 0 / 0 140 / 55 / 50 / 0 M. flavescens type 1 440 / 0 / 0 130 / 115 / 70 / 60 M. aurum type 2 320 / 130 / 0 200 / 70 / 60 / 55 M. immunogenum type 2 320 / 130 / 0 200 / 60 / 55 / 50 M. chelonae type 1 320 / 130 / 0 145 / 70 / 60 / 55 M. immunogenum type 1 320 / 130 / 0 140 / 65 / 60 / 0 M. mucogenicum type 1 320 / 115 / 0 185 / 145 / 0 / 0 M. fallax type 1 320 / 115 / 0 170 / 140 / 0 / 0 M. neoaurum type 1 320 / 115 / 0 145 / 65 / 60 / 0 M. mucogenicum type 2 320 / 115 / 0 140 / 90 / 60 / 0 M. mucogenicum type 3 235 / 210 / 0 200 / 70 / 60 / 50 M. abscessus type 2 235 / 210 / 0 180 / 135 / 70 / 50 M. thermorresistibile type 1 235 / 210 / 0 145 / 140 / 100 / 50 M. peregrinum type 1 235 / 210 / 0 145 / 70 / 60 / 55 M. abscessus type 1 235 / 210 / 0 140 / 125 / 100 / 50 M. peregrinum type 2 235 / 210 / 0 140 / 125 / 60 / 50 M. senegalense type 3 235 / 210 / 0 140 / 80 / 60 / 50 M. phlei type 1 235 / 210 / 0 130 / 80 / 60 / 0 M. celatum type 1 235 / 130 / 85 175 / 80 / 0 / 0 M. aurum type 1 235 / 130 / 85 145 / 140 / 100 / 60 M. peregrinum type 3
235 / 130 / 85 145 / 125 / 60 / 0 M. smegmatis type 1 235 / 130 / 85 140 / 125 / 60 / 50 M. senegalense type 2 235 / 120 / 85 180 / 140 / 50 / 0 M. senegalense type 4 235 / 120 / 85 145 / 120 / 60 / 55 M. fortuitum type 1 235 / 120 / 85 140 / 125 / 60 / 50 M. senegalense type 1 235 / 120 / 85 140 / 120 / 60 / 55 M. fortuitum type 2 235 / 120 / 85 135 / 90 / 85 / 0 M. fortuitum type 3 B 136 108,28 320 / 115 / 0 140 / 90 / 60 / 0 M. chitae type 1
235 / 210 / 0 145 / 130 / 0 / 0 M. avium subsp. avium type 3 235 / 210 / 0 130 / 105 / 60 / 0 M. avium subsp. avium type 2 235 / 210 / 0 130 / 105 / 0 / 0 M. avium subsp. avium type 1 235 / 210 / 0 130 / 105 / 0 / 0 M. avium subsp. paratuberculosis
type 1 C 136 76,60 235 / 120 / 85 160 / 105 / 60 / 0 M. xenopi type 1 D 75,61 or 136 440 / 0 / 0 170 / 130 / 0 / 0 M. triviale type 1 75,57,4 320 / 115 / 0 130 / 95 / 75 / 60 M. kansasii type 5 235 / 210 / 0 190 / 105 / 80 / 0 M. ulcerans type 2 (235 / 210 / 0 145 / 130 / 95 / 0 M. scrofulaceum type 1*) 235 / 210 / 0 145 / 105 / 80 / 45/20 M. marinum type 1 235 / 210 / 0 145 / 105 / 80 / 0 M. ulcerans type 1 235 / 210 / 0 130 / 105 / 80 / 0 M. kansasii type 1 235 / 130 / 85 140 / 105 / 70 / 0 M. shimodei type 1 235 / 120 / 85 130 / 115 / 75 / 60 M. kansasii type 4 235 / 130 / 85 130 / 105 / 70 / 0 M. kansasii type 6 235 / 130 / 85 130 / 105 / 0 / 0 M. kansasii type 2 235 / 130 / 85 130 / 95 / 70 / 0 M. kansasii type 3 E 75,61 or 108,28 440 / 0 / 0 145 / 130 / 0 / 0 M. simiae type 5 75,57,4 320 / 115 / 0 185 / 140 / 0 / 0 M. terrae type 2 320 / 115 / 0 180 / 130 / 0 / 0 M. terrae type 1 320 / 115 / 0 145 / 130 / 0 / 0 M. simiae type 4 320 / 115 / 0 140 / 90 / 60 / 0 M. nonchromogenicum type 2 320 / 115 / 0 140 / 60 / 50 / 0 M. terrae type 3 320 / 115 / 0 125 / 105 / 0 / 0 M. genavense type 1
235 / 210 / 0 185 / 130 / 0 / 0 M. simiae type 1 235 / 210 / 0 185 / 130 / 0 / 0 M. genavense type 2 235 / 210 / 0 155 / 140 / 0 / 0 M. simiae type 2 235 / 210 / 0 145 / 130 / 0 / 0 M. simiae type 6 235 / 210 / 0 140 / 115 / 70 / 0 M. terrae type 4 235 / 130 / 85 145 / 130 / 0 / 0 M. simiae type 3 235 / 130 / 85 130 / 105 / 70 / 0 M. gastri type 1 235 / 120 / 85 145 / 60 / 55 / 0 M. nonchromogenicum type 1 F 75,61 or 76,60 440 / 0 / 0 130 / 105 / 70 / 0 M. szulgai type 1 75,57,4 (320 / 115 / 0 130 / 115 / 60 / 0 M. gordonae type 4*) 240/210/0 130/110/0 M. interjectum (235 / 210 / 0 145 / 130 / 0 / 0 M.intracellulare type 3*) 235 / 210 / 0 115 / 105 / 0 / 0 M. asiaticum type 1 235 / 130 / 85 130 / 105 / 80 / 0 M. celatum type 2 235 / 120 / 100 145 / 105 / 80 / 0 M. malmoense type 1 235 / 210 / 0 145 / 105 / 80 / 0 M. malmoense type 2 (235 / 120 / 100 130 / 115 / 0 / 0 M. gordonae type 3*) G 75,61 or 76,32,28 (440 / 0 / 0 145 / 130 / 0 / 0 M. simiae type 5*) 75,57,4 320 / 115 / 0 130 / 110 / 70 / 60 M. gordonae type 8 320 / 115 / 0 130 / 115 / 60 / 0 M. gordonae type 4 235 / 210 / 0 145 / 130 / 0 / 0 M. intermedium type 1 235 / 210 / 0 145 / 130 / 0 / 0 M. intracellulare type 3 235 / 210 / 0 140 / 105 / 80 / 0 M. intracellulare type 2 235 / 210 / 0 130 / 115 / 0 / 0 M. gordonae type 5 235 / 210 / 0 120 / 115 / 110 / 0 M. intracellulare type 4 235 / 130 / 85 140 / 120 / 95 / 0 M. gordonae type 6 235 / 120 / 100 160 / 115 / 60 / 0 M. gordonae type 9 235 / 120 / 100 155 / 110 / 0 / 0 M. gordonae type 7 235 / 120 / 100 145 / 130 / 60 / 0 M. intracellulare type 1 235 / 120 / 100 130 / 115 / 0 / 0 M. gordonae type 3 235 / 120 / 100 130 / 110 / 95 / 0 M. gordonae type 10 235 / 120 / 85 160 / 115 / 60 / 0 M. gordonae type 1 235 / 120 / 85 215 / 110 / 0 / 0 M. gordonae type 2 H 75,61 or 66,60,10 235 / 210 / 0 145 / 130 / 95 / 0 M. scrofulaceum type 1
75,57,4 320 / 130 / 0 160 / 110 / 0 / 0 M. haemophilum type 1
T
(235 bp) 235 / 120 / 85 150 / 130 / 70 / 0 M. tuberculosis type 1
DISCUSSION
Mycobacterial species identification is generally achieved by standard culture and biochemical methods [13]. Biochemical methods are labor-intensive and time-consuming, and not all are reproducible [2, 14]. PRA or DPRA, the molecular identification of mycobacterial species using different regions of hsp65, 16S rDNA, 16S-23S rDNA spacer, dnaJ, and rpoB as an amplification target [3, 16-19], has been developed in recent years. The hsp65 PRA, the most commonly applied method, has
74 patterns of 40 species in the PRASITE database (http://app.chuv.ch/prasite/ index.html).
Previous studies [20-21] reported that hsp65 PRA provided more time-saving and accurate species identification than phenotypic or biochemical testing because more incorrect and ambiguous results critical for the clinical importance were found in phenotypic method. This was due to phenotypic homogeneity among different species and phenotypic variability within a species [20]. Some sub-species with difficult identification such as M. kansasii, or separation of rapid-growing mycobacterium (RGM) between M. abscessus and M.chelonae and M. fortuitum
group and M. smegmatis group [22]can be made by hsp65 PRA [23].
However, the limitation of hsp65 PRA has been reported in some articles [24-25]. Failure or mis-identification may occur due to similarities in a number of band sizes critical for discriminating species, including difficult to distinguish MTB complex (M. tuberculosis and M. bovis) [24], and closely related sub-species such as M. avium or M. gordonae due to sequence heterogeneity [24]. In addition, technical problems can also cause misinterpretation or incorrect identification [25]. Patterns in the PRA profiles are more complex and difficult to interpret with the naked eye, especially when more detailed sub-types are included [23].
and clinical respiratory isolates. The useful mycobacterial identification flow chart (Fig. 1) can make species identification to the sub-species level, and final species identification can be obtained instantly by concordant results of the two PRA. M. gordonae has a highly variable gene sequence with 10 sub-types in hsp65 PRA, and there are two groups in rpoB DPRA: the G and F groups. Most M. gordonae is G group, but M. gordonae types 3 and 4 by hsp65 PRA areF group (Table 1 and 2).
In addition, there were different rpoB DPRA (Table 2) in M. simaie type 5 (G group but not E group), M. scrofulaceum type 1 (D group but not H group), and M. intracellulare type 3 (F group but not G group). All of these isolates were finally confirmed by 16S rDNA sequencing. Variable numbers of restriction site for HaeIII in these species may be due to genetic sequence mutation. However, these species are included in the modified species identification algorithm although they are uncommon isolates.
Using the mycobacteria identification flow chart (Fig.1) and modified algorithm(Table 3), M. avium-intracellulare complex (MAC) can be easily divided into M. avium spp. avium and M. intracellulare by both rpoB DPRA and hsp65 PRA, which conventional method cannot be made. From the findings in Table 3, some NTM species with identical or similar hsp65 PRA can be clearly grouped by rpoB DPRA (Table 4). By combining rpoB DPRA and hsp65 PRA, ambiguous results by hsp65 PRA alone can be solved more clearly. However, M. intermedium type 1 and M. intracellulare type 3 with identical hsp65 PRA and rpoB DPRA (G group) are not further differentiated by this modified species identification algorithm and instead require 16S rDNA sequencing for confirmation.
Although 16S rDNA sequencing is the standard method for mycobacterium species identification, it does not differentiate some closely related RGM species [26]
as well as slow-growing M. kansasii and M. gastri with identical 16S rDNA sequence, which can be made by hsp65 PRA and rpoB DPRA. Some articles [6, 27] report that different methods for mycobacterial species identification often have conflicting results because one method may have limitations in failing to identify all test strains correctly. Combining more methods for mycobacterial species identification can improve the accuracy rate, avoid ambiguous results and save more time.
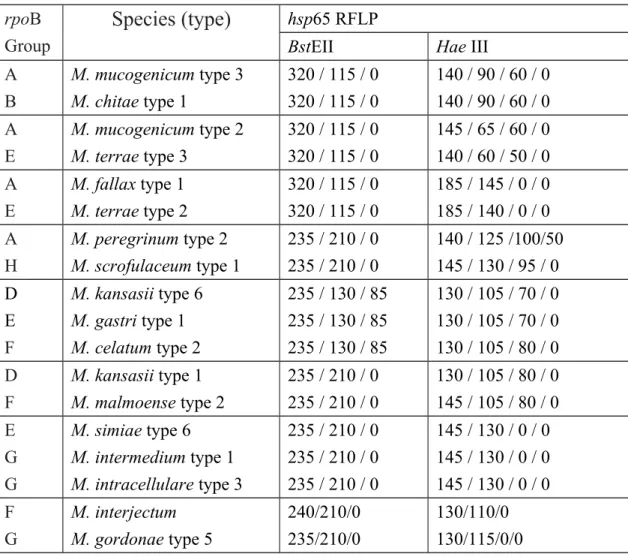
Table 4 Species with identical or similar hsp65 PRA but different groups in rpoB DPRA
rpoB Group
Species (type)
hsp65 RFLPBstEII Hae III
A B M. mucogenicum type 3 M. chitae type 1 320 / 115 / 0 320 / 115 / 0 140 / 90 / 60 / 0 140 / 90 / 60 / 0 A E M. mucogenicum type 2 M. terrae type 3 320 / 115 / 0 320 / 115 / 0 145 / 65 / 60 / 0 140 / 60 / 50 / 0 A E M. fallax type 1 M. terrae type 2 320 / 115 / 0 320 / 115 / 0 185 / 145 / 0 / 0 185 / 140 / 0 / 0 A H M. peregrinum type 2 M. scrofulaceum type 1 235 / 210 / 0 235 / 210 / 0 140 / 125 /100/50 145 / 130 / 95 / 0 D E F M. kansasii type 6 M. gastri type 1 M. celatum type 2 235 / 130 / 85 235 / 130 / 85 235 / 130 / 85 130 / 105 / 70 / 0 130 / 105 / 70 / 0 130 / 105 / 80 / 0 D F M. kansasii type 1 M. malmoense type 2 235 / 210 / 0 235 / 210 / 0 130 / 105 / 80 / 0 145 / 105 / 80 / 0 E G G M. simiae type 6 M. intermedium type 1 M. intracellulare type 3 235 / 210 / 0 235 / 210 / 0 235 / 210 / 0 145 / 130 / 0 / 0 145 / 130 / 0 / 0 145 / 130 / 0 / 0 F G M. interjectum M. gordonae type 5 240/210/0 235/210/0 130/110/0 130/115/0/0
Many capillary electrophoresis (CE)-based studies [5-9] in PCR-RFLP analysis were developed to improve band sizes discrimination. In one study by Chang et al.
[7], high-resolution CE has more precise estimates of DNA fragment sizes than the naked eye and can detected low molecular weight fragment size to 12 bp. In our study, restriction fragments <50 bp were not selected for analysis to avoid confusion with primer and primer dimer bands. We found that more hsp65 fragment differences than rpoB fragmen t(data not shown) may explain the size differences with highly variable sequence for species identification but difficult interpretation in hsp65 PRA.
Some sub-types of NTM species are relevant to clinical management, such as the M. kansasii and M. avium-intracellulare complex (MAC). M. kansasii type 1 is the most common type associated with human disease [28-30] due to its most pathogenicity. However, M. kansasii types 3-7 are most often isolated from the environment, rarely from human, and have no significant role in clinical management [28-29]. The M. avium-intracellulare complex (MAC) can be divided into M. avium subsp. avium and M. intracellulare because drug sensitivity test and clinical outcomes are different between these two sub-types of NTM species [31-32].
Taken together, it is important to identify NTM to the sub-type level not only for epidemiologic data but also for differentiating potentially pathogenic sub-types [28-29]. By combined rpoB DPRA and hsp65 PRA with capillary electrophoresis, species identification can be reported more precisely to solve the problem in difficult discrimination by hsp65 PRA band sizes. This combined method just takes 2-3 days of turnaround time in the laboratory from receipt of clinical isolates to the results reported. However, the modified algorithm has some limitation. First, it still could not discriminate M. intermedium type 1 from M. intracellulare type 3. Second, not every laboratory in the hospital could be equipped with the facilities.
Conclusion
In conclusion, the novel flow chart and modified algorithm by combined rpoB DPRA and hsp65 PRA with capillary electrophoresis can easily differentiate MTB
from NTM and further make mycobacterial species identification to sub-type level, which is helpful for clinical management. It is also complementary to 16S rDNA sequencing, and is a rapid, accurate and effective algorithm for mycobacterial species identification.
Acknowledgements
This work was supported by grants from the Center of Disease Control (DOH95 -DC - 1106) and the National Science Foundation (NSC-982A52) of Taiwan.
Author details
1Division of Respiratory and Critical Care Medicine, Department of internal
medicine, Taichung Hospital, Executive Yuan Department of health, Taiwan, ROC. 2Institute of Molecular Biology, National Chung Hsing University, Taichung 402, Taiwan. 3Institute of Microbiology and Immunology, National Yang-Ming University, Taipei, Taiwan, ROC. 4The Central Region Laboratory, Centers for Disease Control, Department of Health, Taichung 408, Taiwan. 5Division of Respiratory and Critical Care Medicine, Department of Internal Medicine, Taichung Veterans General Hospital, Taichung, Taiwan, ROC. 6Department of Laboratory Medicine, Linkou Chang-Gung Memorial Hospital, Taoyuan, Taiwan. 7Department of Respiratory Therapy, College of Health Care, China Medical University, Taiwan, ROC. 8Institute of Nursing Care in Hungeuang University, Taichung, Taiwan, ROC. 9Graduate Institute of Clinical Medical Science, China Medical University, Taichung, Taiwan, ROC
Authors’ contributions
CCH interpretated and wrote this manuscript. CSC, JHC, STH participated in the design, and analysis. GHS and WCH managed the project. JYH, JJL assisted in improving the manuscript. All authors read and approved the final manuscript. The authors declare no conflict of interest.
REFERENCES
1. Collins CH, Grange JM, Yates MD: Tuberculosis bacteriology : organization and practice. 2nd edn. Oxford ; Boston: Butterworth-Heinemann; 1997. 2. Springer B, Stockman L, Teschner K, Roberts GD, Bottger EC:
Two-laboratory collaborative study on identification of mycobacteria:
molecular versus phenotypic methods. J Clin Microbiol 1996, 34:296-303. 3. Telenti A, Marchesi F, Balz M, Bally F, Bottger EC, Bodmer T: Rapid
identification of mycobacteria to the species level by polymerase chain reaction and restriction enzyme analysis. J Clin Microbiol 1993, 31:175-178.
4. Wong DA, Yip PC, Tse DL, Tung VW, Cheung DT, Kam KM: Routine use of a simple low-cost genotypic assay for the identification of mycobacteria in a high throughput laboratory. Diagn Microbiol Infect Dis 2003, 47:421-426.
5. Chang PL, Hsieh WS, Chiang CL, Yen-Liberman B, Procop GW, Chang HT, Ho HT: Identification of individual DNA molecule of Mycobacterium tuberculosis by nested PCR-RFLP and capillary electrophoresis. Talanta 2008, 77:182-188.
6. Sajduda A, Martin A, Portaels F, Palomino JC: hsp65 PCR-restriction analysis (PRA) with capillary electrophoresis in comparison to three other methods for identification of Mycobacterium species. J Microbiol Methods 2010, 80:190-197.
7. Chang PL, Hsieh WS, Chiang CL, Tuohy MJ, Hall GS, Procop GW, Chang HT, Ho HT: The hsp65 gene patterns of less common Mycobacterium and Nocardia spp. by polymerase chain reaction-restriction fragment length polymorphism analysis with capillary electrophoresis. Diagn Microbiol Infect Dis 2007, 58:315-323.
8. Yokoyama E, Kishida K, Uchimura M, Ichinohe S: Comparison between agarose gel electrophoresis and capillary electrophoresis for variable numbers of tandem repeat typing of Mycobacterium tuberculosis. J Microbiol Methods 2006, 65:425-431.
9. Lindstedt BA, Vardund T, Aas L, Kapperud G: Multiple-locus variable-number tandem-repeats analysis of Salmonella enterica subsp. enterica serovar Typhimurium using PCR multiplexing and multicolor capillary electrophoresis. J Microbiol Methods 2004, 59:163-172.
10. Kim BJ, Hong SK, Lee KH, Yun YJ, Kim EC, Park YG, Bai GH, Kook YH: Differential identification of Mycobacterium tuberculosis complex and nontuberculous mycobacteria by duplex PCR assay using the RNA
polymerase gene (rpoB). J Clin Microbiol 2004, 42:1308-1312.
11. Shen GH, Hung CH, Hu ST, Wu BD, Lin CF, Chen CH, Wu KM, Chen JH: Combining polymerase chain reaction restriction enzyme analysis with phenotypic characters for mycobacteria identification in Taiwan. Int J Tuberc Lung Dis 2009, 13:472-479.
12. Hanna BA: Diagnosis of tuberculosis by microbiologic techniques. Philadelphia, PA, USA: Little, Brown and Company. ; 1996.
13. Witebsky FG, Kruczak-Filipov P: Identification of mycobacteria by conventional methods. Clin Lab Med 1996, 16:569-601.
14. Vossler JL: Mycobacterium tuberculosis and other non-tuberculous mycobacteria. . In Text-book of diagnostic microbiology Edited by In: Mahon C R MG, eds. . Philadephia, PA, USA: W B Saunders; 2000: 667–707
15. Turenne CY, Tschetter L, Wolfe J, Kabani A: Necessity of Quality-Controlled 16S rRNA Gene Sequence Databases: Identifying
Nontuberculous Mycobacterium Species. J Clin Microbiol 2001, 39:3637-3648.
16. Domenech P, Menendez MC, Garcia MJ: Restriction fragment length polymorphisms of 16S rRNA genes in the differentiation of fast-growing mycobacterial species. FEMS Microbiol Lett 1994, 116:19-24.
17. Lee H, Park HJ, Cho SN, Bai GH, Kim SJ: Species identification of
mycobacteria by PCR-restriction fragment length polymorphism of the rpoB gene. J Clin Microbiol 2000, 38:2966-2971.
18. Roth A, Reischl U, Streubel A, Naumann L, Kroppenstedt RM, Habicht M, Fischer M, Mauch H: Novel Diagnostic Algorithm for Identification of Mycobacteria Using Genus-Specific Amplification of the 16S-23S rRNA Gene Spacer and Restriction Endonucleases. J Clin Microbiol 2000, 38:1094-1104.
19. Takewaki S, Okuzumi K, Manabe I, Tanimura M, Miyamura K, Nakahara K, Yazaki Y, Ohkubo A, Nagai R: Nucleotide sequence comparison of the mycobacterial dnaJ gene and PCR-restriction fragment length
polymorphism analysis for identification of mycobacterial species. Int J Syst Bacteriol 1994, 44:159-166.
20. Chimara E, Ferrazoli L, Ueky SY, Martins MC, Durham AM, Arbeit RD, Leao SC: Reliable identification of mycobacterial species by PCR-restriction enzyme analysis (PRA)-hsp65 in a reference laboratory and elaboration of a sequence-based extended algorithm of PRA-hsp65 patterns. BMC Microbiol 2008, 8:48.
JS, Yim JJ: Differentiation of mycobacteria in sputa by duplex polymerase chain reaction for mycobacterial hsp65 gene. Diagn Microbiol Infect Dis 2008, 62:193-198.
22. Brown-Elliott BA, Wallace RJ, Jr.: Clinical and taxonomic status of pathogenic nonpigmented or late-pigmenting rapidly growing mycobacteria. Clin Microbiol Rev 2002, 15:716-746.
23. Kim HJ, Mun HS, Kim H, Oh EJ, Ha Y, Bai GH, Park YG, Cha CY, Kook YH, Kim BJ: Differentiation of Mycobacterial species by hsp65 duplex PCR followed by duplex-PCR-based restriction analysis and direct sequencing. J Clin Microbiol 2006, 44:3855-3862.
24. McNabb A, Eisler D, Adie K, Amos M, Rodrigues M, Stephens G, Black WA, Isaac-Renton J: Assessment of partial sequencing of the 65-kilodalton heat shock protein gene (hsp65) for routine identification of Mycobacterium species isolated from clinical sources. J Clin Microbiol 2004, 42:3000-3011. 25. Leao SC, Bernardelli A, Cataldi A, Zumarraga M, Robledo J, Realpe T, Mejia
GI, da Silva Telles MA, Chimara E, Velazco M, et al: Multicenter evaluation of mycobacteria identification by PCR restriction enzyme analysis in laboratories from Latin America and the Caribbean. J Microbiol Methods 2005, 61:193-199.
26. Ringuet H, Akoua-Koffi C, Honore S, Varnerot A, Vincent V, Berche P, Gaillard JL, Pierre-Audigier C: hsp65 sequencing for identification of rapidly growing mycobacteria. J Clin Microbiol 1999, 37:852-857. 27. Häfner B, Haag H, Geiss H-K, Nolte O: Different molecular methods for
the identification of rarely isolated non-tuberculous mycobacteria and description of new hsp65 restriction fragment length polymorphism patterns. Molecular and Cellular Probes 2004, 18:59-65.
28. da Silva Telles MA, Chimara E, Ferrazoli L, Riley LW: Mycobacterium kansasii: antibiotic susceptibility and PCR-restriction analysis of clinical isolates. J Med Microbiol 2005, 54:975-979.
29. Taillard C, Greub G, Weber R, Pfyffer GE, Bodmer T, Zimmerli S, Frei R, Bassetti S, Rohner P, Piffaretti JC, et al: Clinical implications of
Mycobacterium kansasii species heterogeneity: Swiss National Survey. J Clin Microbiol 2003, 41:1240-1244.
30. Zhang Y, Mann LB, Wilson RW, Brown-Elliott BA, Vincent V, Iinuma Y, Wallace RJ, Jr.: Molecular analysis of Mycobacterium kansasii isolates from the United States. J Clin Microbiol 2004, 42:119-125.
31. Maekura R, Okuda Y, Hirotani A, Kitada S, Hiraga T, Yoshimura K, Yano I, Kobayashi K, Ito M: Clinical and prognostic importance of serotyping
Mycobacterium avium-Mycobacterium intracellulare complex isolates in human immunodeficiency virus-negative patients. J Clin Microbiol 2005, 43:3150-3158.
32. Yamori S, Tsukamura M: Comparison of prognosis of pulmonary diseases caused by Mycobacterium avium and by Mycobacterium intracellulare. Chest 1992, 102:89-90.