J. Microbiol. Biotechnol. (2008), 18(4), 784–787
Synergistic Interactions Between Chitinase ChiCW and Fungicides Against
Plant Fungal Pathogens
Huang, Chien-Jui and Chao-Ying Chen*
Department of Plant Pathology and Microbiology, National Taiwan University, Taipei 106, Taiwan, China Received: June 2, 2007 / Accepted: September 1, 2007
Antifungal activity of ChiCW and synergistic interactions between ChiCW with fungicides were investigated. Conidial germinations of phytopathogenic fungi, Alternaria brassicicola, Botrytis elliptica, and Colletotrichum gloeosporioides, were inhibited by ChiCW but A. longipes was not. In addition, ChiCW showed synergistic effect with fungicides Switch (cyprodinil+fludioxonil) and tebuconazole to inhibit fungal conidial germinations. The level of synergism of ChiCW with tebuconazole was higher than that with Switch. The results indicate that ChiCW may exhibit a higher level of synergism with fungicides that have a primary effect upon membranes.
Keywords: ChiCW, synergistic interaction, fungicide, phytopathogenic fungi, tebuconazole
ChiCW of Bacillus cereus 28-9, as a modular endochitinase, consists of a signal peptide, a catalytic domain, a fibronectin type III-like domain, and a chitin-binding domain [4, 5]. In the previous study, ChiCW showed antifungal activity effectively against conidial germination of Botrytis elliptica, the causal agent of lily leaf blight. Based on this result, ChiCW was proposed to play an important role in the antagonism of B. cereus 28-9 against B. elliptica [5].
Synergistic interactions between fungal cell-wall degrading enzymes from Trichoderma harzianum and Gliocladium virens and antifungal compounds have been demonstrated [9,10]. Cell-wall degrading enzymes from Trichoderma virens and a Gram-negative bacterium, Serratia marcescens strain B2, exhibited synergistic antifungal activities with bacterial antibiotics [14, 16]. To date, many chitinases have been identified from bacteria and showed antifungal activities [3, 5]. In our previous study, the antifungal activity of a biocontrol strain, Bacillus subtilis F29-3, was enhanced by expressing the chiA gene of Bacillus circulans WL-12 in
strain F29-3 [2]. However, synergistic interactions between chitinases from Gram-positive bacteria and antifungal compounds against plant pathogenic fungi were not reported previously, to our memory. Therefore, the aim of this study was to investigate the antifungal activity of ChiCW and synergistic interactions between ChiCW with fungicides against several phytopathogenic fungi.
Four fungal species were used as test fungi in this study: Alternaria brassicicola (causing black spot disease on Brassica species), A. longipes (causing brown spot of tobacco), B. elliptica B061, and Colletotrichum gloeosporioides (causing anthracnose disease). A. brassicicola, A. longipes, and C. gloeosporioides were cultured on potato dextrose agar (PDA, Difco) at 25oC. B. elliptica was cultured on
V-8 juice agar [20% V8 juice (Campbell Soup Co.), 0.3% CaCO3, 1.8% agar] at 25oC.
For testing fungicide sensitivities of the test fungi, six formulated fungicides, benomyl (Benlate, 50% wettable powder; DuPont), carbendazim (Bavistin, 50% wettable powder; BASF), dichlorofluanid (Euparen, 50% wettable powder; Bayer), cyprodinil+fludioxonil (Switch, 62.5% water-dispersible granules; Syngenta), iprodione (Rovral, 50% wettable powder; Bayer), and tebuconazole (Folicur, 25.9% emulsifiable concentrate; Bayer), were selected. Stock solutions of the commercial formulations in sterile water were added to molten PDA, and plates were infested with mycelial disks of fungal strains obtained from actively growing cultures. Subsequently, plates were incubated at 25oC to examine fungicide sensitivities of the test fungi.
ChiCW was purified from the periplasmic protein of E. coli DH5α (pNTU55) according to the method of
Huang et al. [5]. Protein concentration was determined by Bradford’s method [1] using bovine serum albumin as the standard.
Antifungal activities of chitinase and fungicides were assayed by the method of Huang et al. [5] with a slight modification. Assay mixtures contained 10µl of a conidial
suspension (1×105conidia/ml) and an equal volume of an
enzyme or a fungicide solution. Cyprodinil+fludioxonil
*Corresponding author
Phone: 886-2-33665207; Fax: 886-2-23657735; E-mail: cychen@ntu.edu.tw
SYNERGISTIC INTERACTIONS BETWEEN ChiCW AND FUNGICIDES 785 and tebuconazole were used in bioassays for B. elliptica
and the other test fungi, respectively. In the controls, sterile distilled water was used instead of the tested solution. On the other hand, the procedure to analyze synergistic interactions between ChiCW and fungicides was conducted as described by Lorito et al. [9, 10] with some modifications. Assay mixtures contained 10µl of a conidial suspension,
9µl of an enzyme solution (made at appropriate concentration),
and 1µl of a fungicide solution (made at appropriate
concentration). In the controls, sterile distilled water was used instead of the solution containing either the enzyme or the fungicide or both. Conidial germination was examined under a light microscope and the percentage of inhibition was calculated after incubation of prepared assay mixtures at 25oC for 12 h. Each assay was triplicated.
The presence of synergism between enzyme and fungicides was determined by using Limpel’s formula [13]: Ee=(X+
Y-XY)/100, where Ee is the expected effect from additive
responses of two inhibitory compounds, and X and Y are the percentages of inhibition relative to each compound used alone. If the combination of the two agents produces any value of inhibition greater than Ee, then synergism exits.
Fungicide sensitivities of our fungal strain had to be investigated firstly in this study. Among 6 different fungicides, only tebuconazole could inhibit all test fungi (data not shown). Tebuconazole was used for further assays of all test fungi but not for B. elliptica. Switch (cyprodinil+ fludioxonil) was selected to perform further assays of B. elliptica B061 because this fungicide was recommended to control B. elliptica by the manufacturer, although B. elliptica B061 was also sensitive to tebuconazole.
Fifty % effective concentrations (EC50) of fungicides
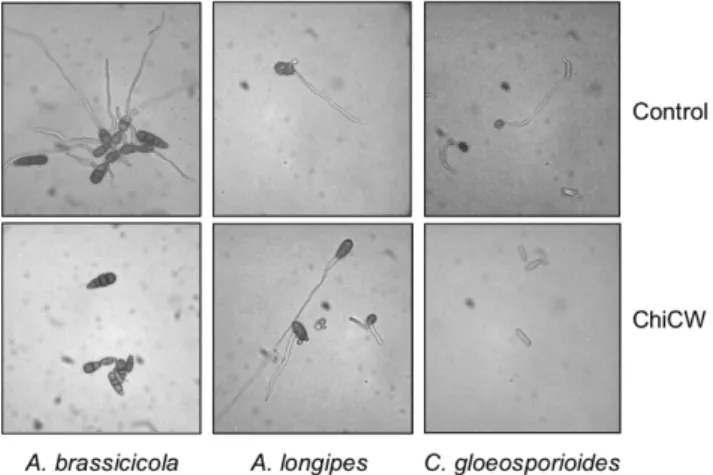
and ChiCW used alone to inhibit conidial germination were investigated (Table 1). Switch effectively inhibited conidial germination of B. elliptica, and tebuconazole can inhibited conidial germinations of A. brassicicola, A. longipes, and C. gloeoporioides. ChiCW was effective against A. brassicicola, B. elliptica, and C. gloeosporioides but not able to inhibit A. longipes (Fig. 1).
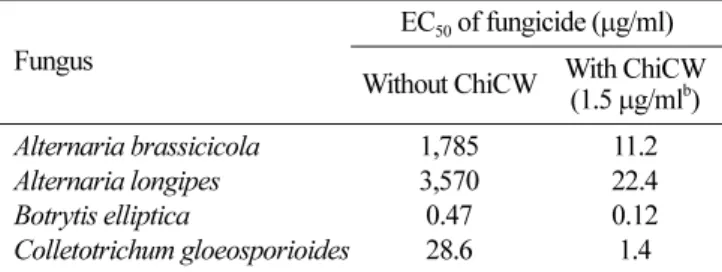
Every fungicide/ChiCW combination tested inhibited conidial germination of the test fungi and showed a substantial level of synergism (Table 2). When ChiCW (at uninhibitory concentration) was applied with Switch,
the EC50 of Switch decreased about four times (Ee=12.5).
The fungicide Switch contains two kinds of fungicides, cyprodinil and fludioxonil, with different modes of action. Cyprodinil (4-cyclopropyl-6-methyl-N-phenylpyrimidine) is an anilinopyrimidine fungicide and has been proposed to inhibit amino acid methionine biosynthesis [11]. Fludioxonil [4-(2,2-difluoro-1,3-benzodioxol-4-yl)-1H -pyrrole-3-carbonitrile], a synthetic analog of the bacterial metabolite of pyrrolnitrin, is a phenylpyrrole fungicide and demonstrates its fungicidal effect through hyperaction of the Mak protein kinase in osmotic signal transduction [6, 12]. Using a very small amount of Switch (0.47 ppm) was effective against 50% conidial germination of B. elliptica (Table 1). Furthermore, when ChiCW was applied with Switch, the EC50 of Switch decreased four times, indicating that ChiCW
can synergistically interact with Switch to inhibit conidial germination of B. elliptica.
In addition, ChiCW also exhibited synergistic interaction with tebuconazole to inhibit A. brassicicola, A. longipes, and C. gloeosporioides. Tebuconazole [1-(4-chlorophenyl)-4,4-dimethyl-3-(1,2,4-triazole-1-yl-methyl)pentane-3-ol] is a demethylation inhibitor fungicide and inhibits C14-demethylase in sterol biosynthesis [7, 8]. When ChiCW (at uninhibitory concentration) was applied with tebuconazole,
Table 1. Fifty percent effective concentrations (EC50) of two fungicidesa and ChiCW against four fungal species.
Fungus EC50 (µg/ml)
ChiCW Switch Tebuconazole
Alternaria brassicicola 3.25 -b 1785
Alternaria longipes No effect - 3570
Botrytis elliptica 3.25 0.47
-Colletotrichum gloeosporioides 3.25 - 28.6
aFungicides Switch and tebuconazole were used in bioassays for B. elliptica and other fungi, respectively. b-: not tested.
Fig. 1. Effect of ChiCW on conidial germinations of A.
brassicicola, A. longipes, and C. gloeosporioides.
Conidial suspensions were treated with purified ChiCW with sterile distilled water as the control.
786 Huang and Chen
the EC50 of tebuconazole for inhibition of C. gloeosporioides
and A. brassicicola decreased 20 and 159 times (both of Ee=0), respectively (Table 2). The result reveals that
ChiCW can significantly interact with tebuconazole to synergistically inhibit conidial germination, especially against A. brassicicola. Furthermore, the EC50 of tebuconazole for
inhibition of A. longipes dramatically decreased 159 times when using the ChiCW/tebuconazole combination (Ee=0).
However, ChiCW used alone did not inhibit conidial germination of A. longipes.
The level of synergism may be affected by the mode of action of the compounds. In the study of Lorito et al. [10], the highest levels of synergism occurred in fungal cell wall degarding enzymes with two sterol demethylation inhibitors and with gliotoxin. Sterols are required for the structure and function of membranes [15]. Combining the effect of these toxins with partial digestion of cell walls may be particularly damaging for the targeted cells and may reduce the lethal doses of the toxins [10]. In this study, the level of synergism of ChiCW with tebuconazole was higher than that with Switch (Table 2), indicating that ChiCW exhibited higher synergism with the fungicides that have primary effect upon membranes.
On the other hand, the synergistic antifungal activity of the cell-wall degrading enzyme and the antifungal compound was correlated with the cell wall composition of the target fungus [16]. In this study, three different species of fungi were used and their cell wall compositions were not exactly known. ChiCW showed inhibitory effect (when used alone) and synergistic effect (when used with fungicides) against conidial germination of A. brassicicola, B. elliptica, and C. gloeosporioides (Tables 1 and 2). The results suggest that ChiCW is able to hydrolyze the cell walls of these fungal conidia and enhance conidia to uptake fungicides.
Compared with A. brassicicola, conidial germination of A. longipes was not inhibited by ChiCW (Table 1, Fig. 1). Although both fungi belong to the genus Alternaria, ChiCW exhibited different effects on the conidial germinations of
Alternaria spp. The result, indicate that the digestion of conidial cell walls of A. longipes by ChiCW may be partial and not enough to directly damage A. longipes cells, revealing that the cell wall compositions and/or structures of A. brassicicola and A. longipes may be different. However, synergism of ChiCW with tebuconazole against A. brassicicola and A. longipes was demonstrated (Table 2), suggesting that the partial digestion of conidial cell walls of A. longipes by ChiCW is enough for tebuconazole to target membranes and to inhibit sterol biosynthesis.
Acknowledgment
This research was supported by the National Science Council of the Republic of China.
REFERENCES
1. Bradford, M. M. 1976. A rapid and sensitive method for the quantitation of microgram quantities of proteins utilizing the principle of protein-dye binding. Anal. Biochem.72: 248-254.
2. Chen, C. Y., Y. H. Wang, and C. J. Huang. 2004. Enhancement of the antifungal activity of Bacillus subtilis F29-3 by the chitinase encoded by Bacillus circulans chiA gene. Can. J. Microbiol. 50: 451-454.
3. Dahiya, N., R. Tewari, and G. S. Hoondal. 2006. Biotechnological aspects of chitinolytic enzymes: A review. Appl. Microbiol. Biotechnol. 71: 773-782.
4. Huang, C. J. and C. Y. Chen. 2005. High-level expression and characterization of two chitinases, ChiCH and ChiCW, of Bacillus cereus 28-9 in Escherichia coli. Biochem. Biophys. Res. Commun. 327: 8-17.
5. Huang, C. J., T. K. Wang, S. C. Chung, and C. Y. Chen. 2005. Identification of an antifungal chitinase from a potential biocontrol agent, Bacillus cereus 28-9. J. Biochem. Mol. Biol.
38: 82-88.
6. Kojima, K., Y. Takano, A. Yoshimi, C. Tanaka, T. Kikuchi, and T. Okuno. 2004. Fungicide activity through activation of a fungal signalling pathway. Mol. Microbiol. 53: 1785-1796.
7. Kwok, I. M. Y. and R. T. Loeffler. 1993. The biochemical mode of action of some newer azole fungicides. Pestic. Sci. 39:
1-11.
8. Lamb, D. C., D. E. Kelly, N. J. Manning, D. W. Hollomon, and S. L. Kelly. 1998. Expression, purification, reconstitution and inhibition of Ustilago maydis sterol 14α-demethylase (CYP51;
P45014DM). FEMS Microbiol. Lett. 169: 369-373.
9. Lorito, M., C. K. Hayes, A. Zoina, F. Scala, G. Del Sorbo, S. L. Woo, and G. E. Harman. 1994. Potential of genes and gene products from Trichoderma sp. and Gliocladium sp. for the development of biological pesticides. Mol. Biotechnol. 2:
209-217.
10. Lorito, M., C. Peterbauer, C. K. Hayes, and G. E. Harman. 1994. Synergistic interaction between fungal cell wall degrading enzymes and different antifungal compounds enhances inhibition of spore germination. Microbiology140: 623-629.
Table 2. Fifty percent effective concentrations (EC50) of individual
fungicidesa used either alone or with ChiCW against four fungal
species.
Fungus Without ChiCW With ChiCWEC50 of fungicide (µg/ml) (1.5µg/mlb)
Alternaria brassicicola 1,785 11.2
Alternaria longipes 3,570 22.4
Botrytis elliptica 0.47 0.12
Colletotrichum gloeosporioides 28.6 1.4
aFungicides Switch and tebuconazole were used in bioassays for B. elliptica and other fungi, respectively.
bAt this concentration, ChiCW does not inhibit spore germination of test fungi.
SYNERGISTIC INTERACTIONS BETWEEN ChiCW AND FUNGICIDES 787
11. Masner, P., P. Muster, and J. Schmid. 1994. Possible methionine biosynthesis inhibition by pyrimidinamine fungicides. Pestic. Sci. 42: 163-166.
12. Okada, A., S. Banno, A. Ichiishi, M. Kimura, I. Yamaguchi, and M. Fujimura. 2005. Pyrrolnitrin interferes with osmotic signal transduction in Neurospora crassa. J. Pestic. Sci. 30:
378-383.
13. Richer, D. L. 1987. Synergism - a patent view. Pestic. Sci. 19:
309-315.
14. Someya, N., M. Nakajima, K. Hirayae, T. Hibi, and K. Akutsu. 2001. Synergistic antifungal activity of chitinolytic enzymes and
prodigiosin produced by biocontrol bacterium, Serratia marcescens strain B2, against gray mold pathogen, Botrytis cinerea. J. Gen. Plant Pathol. 67: 312-317.
15. Vanden Bossche, H. 1990. Importance and role of sterols in fungal membranes. In: Biochemistry of Cell Walls and Membranes in Fungi. Springer-Verlag, Berlin.
16. Woo, S., V. Fogliano, F. Scala, and M. Lorito. 2002. Synergism between fungal enzymes and bacterial antibiotics may enhance biocontrol. Antonie Van Leeuwenhoek81: 353-356.