Active Intracellular Domain of Notch Enhances Transcriptional Activation of
CCAAT/Enhancer Binding Protein
β on a Rat Pregnancy-Specific Glycoprotein
Gene
†Hungwen Chen,*,‡,§Yichuen Chong,§and Chia-Lin Liu§
Institute of Biological Chemistry, Academia Sinica, Nankang, Taipei 115, Taiwan, and Graduate Institute of Biochemical Sciences, National Taiwan UniVersity, Taipei 10098, Taiwan
ReceiVed August 2, 1999; ReVised Manuscript ReceiVed October 27, 1999
ABSTRACT: Pregnancy-specific glycoproteins (PSGs) are primarily expressed in the placenta and become the major glycoproteins at term. To understand the regulation of PSG expression, we characterized the promoter elements of a rodent PSG gene, rnCGM3, and delineated three nuclear protein binding sites: FPI, -II, and -III in the 5′-flanking region of the gene. The FPII-binding factor is shown to be C/EBPβ, which positively regulates rnCGM3 expression [Chen, H., et al. (1995) DNA Cell Biol. 14, 681-688]. In the current study, we used the yeast one-hybrid system to isolate transcription factors binding to the FPIII site and demonstrated that a rodent Jκ recombination signal sequence binding protein, rRBPJκ-2N, bound to the FPIII site. Electrophoretic mobility shift assay with rat placental nuclear proteins revealed a constitutive occupancy of the FPIII site by RBPJκ. By transient expression analyses, we demonstrated that rRBPJκ-2N repressed the expression from an FPIII-driven SV40 promoter. However, this repression effect was counteracted by the active intracellular domain of Notch (NotchIC). Using the native rnCGM3 promoter construct, we demonstrated that the promoter activity stimulated by C/EBPβ was also repressed by rRBPJκ-2N but enhanced by NotchIC. Additionally, we found that NotchIC can stimulate expression through another RBPJκ site within the FPI site. A functional interaction between factors binding to the FPI, FPII, and FPIII sites is proposed.
Many proteins, steroid hormones, cytokines, and growth factors are released from placenta to the maternal circulation in order to maintain pregnancy and fetal development. Pregnancy-specific glycoproteins (PSGs) are a family of glycoproteins synthesized by the syncytiotrophoblast in placenta (1). Human PSGs are consistently detectable in the maternal serum during the first 2-3 weeks of pregnancy. The level of PSGs in maternal circulation increases expo-nentially to reach 200-400µg/mL and becomes the most abundant placental proteins at term (2). Although the exact function(s) of PSGs is still not known, low levels of PSGs are associated with poor pregnancy outcomes (3-6). Fur-thermore, antisera against PSGs have been shown to induce abortions in mice and monkeys (7, 8). Putative PSG receptors
have also been identified on monocytic cell lines, suggesting a possible role of PSGs in regulating the maternal immune system during pregnancy (9).
As a model to understand human PSG expression and protein functions, several rodent or murine homologues of human PSG genes have been isolated and characterized. For example, the expression of a rodent PSG gene, rnCGM1, was detected in the giant cell/spongiotrophoblast in rat placenta (10). We have also characterized two other rodent PSG genes, rnCGM6 and rnCGM3, and demonstrated that both are primarily expressed in rat placenta (11, 12). Kromer et al. (13) studied the expression kinetics of three murine PSG genes, Cea2, Cea3, and Cea4, during placental devel-opment and revealed that expression starts at the embryonic stage E12.5 and reaches the maximum steady-state level between E15.5 and E17.5. They also detected an exclusive expression of PSG transcripts in the spongiotrophoblast of the placenta at E17.5.
In an effort to study the molecular mechanisms underlying the placenta-specific expression of PSG genes, in previous studies we have sequenced the 5′-flanking region of rnCGM3 and showed that a TATA box, CATAAA, is located at the nucleotides -226 to -221 region upstream of the translation start site (11). We also analyzed the promoter of rnCGM3 and delineated two promoter elements, PI and PII, that are located upstream and downstream of the transcription start site, respectively (11). Based on DNase I footprinting analyses, two nuclear protein binding sites at nucleotides -311 to -290 (PISI) and -257 to -239 (PISII) in the PI
†This work was supported by grants (to H.C.) from the National
Science Council (Grant 88-2311-B-001-115) and Academia Sinica of Taiwan.
* Correspondence should be addressed to this author: Tel 011-886-2-27855696, ext 6090; Fax 011-886-2-27889759; E-mail [email protected].
‡Institute of Biological Chemistry, Academia Sinica.
§Graduate Institue of Biochemical Sciences, National Taiwan
University.
1Abbreviations: PSG, pregnancy-specific glycoprotein; C/EBPβ,
CCAAT/enhancer binding proteinβ; RBPJκ, Jκ recombination signal
sequence binding protein; NotchIC, intracellular domain of Notch; 3-AT, 3-aminotriazole; EMSA, electrophoretic mobility shift assay; AD, activation domain; E(spl), Enhancer of split; MHC, major histocom-patibility complex; NLS, nuclear localization signal; EBV, Epstein-Barr virus; Su(H), Suppressor of Hairless; EBNA2, Epstein-Epstein-Barr virus nuclear antigen 2; CIR, CBF1 interacting corepressor, HDAC, histone deacetylase; HES-1, Hairy enhancer of split; TCR, T-cell receptor.
10.1021/bi991786k CCC: $19.00 © 2000 American Chemical Society Published on Web 01/28/2000
element and one binding site at nucleotides -108 to -93 (PIISIII) in the PII element (see Figure 1A) were further identified (11). For simplicity, we will call PISI, PISII, and PIISIII as FPI, FPII, and FPIII, respectively, in this report. It has been shown that the factor binding to the FPII site is C/EBPβ (14), a member of the bZIP-class of transcription factors that contain a basic DNA-binding region adjacent to a leucine zipper dimerization domain.
Because PSG is specifically expressed in placenta and the level of PSG increases exponentially during placental development, we speculate that additional transcription factors, in response to environmental cues such as hormones, cytokines, and growth factors, are involved in the upregu-lation of PSG expression. In the present study, we used the yeast one-hybrid system (15) to perform an in vivo screening of a placental cDNA library to isolate nuclear factors binding to the FPIII site. We show that the rodent Jκ recombination signal sequence binding protein (RBPJκ) rRBPJκ-2N binds to the FPIII site. We further demonstrate that rRBPJκ-2N repressed the expression of an FPIII-driven promoter con-struct. We also show that the active intracellular domain of Notch (NotchIC) counteracted the repressor activities of rRBPJκ-2N. In the native context of rnCGM3 promoter, the promoter activity stimulated by C/EBPβ was also repressed by rRBPJκ-2N but enhanced by NotchIC. Additionally, we found that NotchIC stimulated C/EBPβ-dependent expression through another RBPJκ site within the FPI site using an FPIII-deleted native promoter construct. Our data suggest
that the placenta-specific expression of rnCGM3 may be connected to C/EBPβ and the Notch signaling pathway. MATERIALS AND METHODS
Library Screening and Characterization of cDNA Clones. The yeast one-hybrid system was purchased from Clontech (Palo Alto, CA). Yeast cells were transformed by standard procedures using the lithium acetate method (16). Six tandem copies of the FPIII element (5′ -GCCTGGGAAAAAACTC-3′), generated by self-ligation using T4 ligase, were placed upstream of the minimal promoter of the yeast HIS3 gene in vector pHISi-1 to generate the target reporter plasmid, p(FPIII)6HIS3. Yeast strain YM4271 (MATa ura3-52
his3-200 ade2-101 lys2-801 leu2-3 112 trp1-903 tyr1-501) was transformed with 2µg of linearized p(FPIII)6HIS3 and then plated onto a synthetic medium without histidine (SD/-His) to select for positive reporter strains. One of the strains, YM4271FPIII, has a low level of HIS3 activity and did not replicate on SD/-His plates supplemented with 30 mM 3-aminotriazole (3-AT), a competitive inhibitor of the yeast HIS3 protein. For yeast one-hybrid screening, 300 mL of YM4271FPIII cells were grown to mid-log phase in YPD medium. Cells were transformed with 20 µg of a pACT2 human placental cDNA library (a GAL4 activation domain fusion library; Clontech) and plated onto a synthetic medium minus histidine and leucine and containing 30 mM 3-AT. Approximately 5 × 106 yeast colonies were screened. Positive colonies were purified after two more rounds of streaking on selection plates. The pACT2 plasmids were rescued from the positive clones, retransformed into Es-cherichia coli strain HB101, and purified for sequencing.
Plasmid Constructs. For the C/EBPβ expression plasmid pCMVC/EBPβ, a BamHI fragment excised from pMEX-CRP2 (17) was blunt-ended and cloned into pRcCMV (Invitrogen, Carlsbad, CA). The rodent RBPJκ, rRBPJκ-2N, was isolated from a rat placentalλgt10 cDNA library using the human RBPJκ cDNA probe obtained from the yeast one-hybrid screen. The coding region of rRBPJκ-2N was ampli-fied by polymerase chain reaction (PCR) with the Pfu DNA polymerase (Stratagene, La Jolla, CA) and cloned into pRcCMV to make the pCMVrRBPJκ-2N plasmid. The murine Notch1 expression plasmid mNotchIC (18) was kindly provided by Dr. R. Kopan (Washington University, St. Louis, MO).
The plasmid p(FPIII)6SV40CAT contains six FPIII sites fused to the SV40 promoter element and the bacterial CAT gene coding sequence. To make constructs containing multimerized mutant FPIII sites, five copies of FPIIIM34 (5′-GCTAGGGAAAAAACTC-3′) and four copies of FPI-IIM56 (5′-GCCTCCGAAAAAACTC-3′) oligonucleotides were inserted in front of the SV40 promoter element to generate the p(FPIIIM34)5SV40CAT and p(FPIIIM56)4 SV40-CAT reporter plasmids, respectively. Similarly, six copies of FPI (5′-TGTCATCCCTGGGATGAGTGTT-3′) and FPIM34A (5′-TGTCATCCAGGGGATGAGTGTT-3′) oli-gonucleotides were inserted in front of the SV40 promoter element to generate the p(FPI)6SV40CAT and p(FPIM34A)6 -SV40CAT reporter plasmids, respectively. The rnCGM3 promoter construct pCGM3A containing all three FP sites was generated by inserting the rnCGM3 promoter region of nucleotides -326 to -33 relative to the translation start site FIGURE1: (A) Schematic representation of the rodent PSG gene,
rnCGM3. The three DNase I-protected regions in the promoter of rnCGM3 are depicted as solid boxes and denoted as FPI
(nucle-otides -311/-290), FPII (nucle(nucle-otides -257/-239), and FPIII (nucleotides -108/-93), respectively. An arrow denotes the transcription initiation site located at nucleotide -197 upstream of the translation start site (denoted as ATG). The lower part of the panel shows the sense-strand sequence of each DNase I-protected region in the 5′ to 3′direction and the nuclear factor binding to each region. The core binding sequences for RBPJκ in FPI and FPIII sites are underlined. (B) In vivo interaction of candidate proteins with the FPIII element. Clones 6B3 and 6M1 encoding GAL4-RBPJκ fusion proteins were introduced into the yeast strain YM4271FPIII. Cells were plated on synthetic medium minus histidine and leucine and containing 30 mM 3-AT. As controls, yeast cells were also transformed with TE (10 mM Tris-HCl, pH 8.0, and 1 mM EDTA) solution (mock), the mouse p53 (mp53), or the parental plasmid (pACT2).
in front of the luciferase gene of the pGL3-Basic plasmid (Promega, Madison, WI). The pCGM3A-∆FPIII construct is identical to pCGM3A except that the FPIII site is deleted. The pCGM3B construct contains the rnCGM3 promoter region of nucleotides -283 to -33, which includes the FPII and III sites. The pCGM3B-FPIIIM34A construct is identi-cal to pCGM3B except it contains the M34A mutation in the FPIII site. The pCGM3A-FPI&IIIM34A construct is identical to pCGM3A except it contains the M34A mutation in FPI (5′-TGTCATCCAGGGGATGAGTGTT-3′) and the M34A mutation in FPIII (5′-GCAGGGGAAAAAACTC-3′). All constructs were verified by DNA sequencing.
Electrophoretic Mobility Shift Assay. For electrophoretic mobility shift assay (EMSA), nuclear extracts of rat placentas were prepared as described by Ohlsson and Edlund (19). In vitro transcription/translation of rRBPJκ-2N cDNA was performed with the TNT-coupled wheat germ extract system (Promega). End-labeled oligonucleotide FPIII probes [1 ng; (0.5-1)× 105cpm] were incubated in the binding reaction buffer (10 mM Tris-HCl, pH 7.5, 50 mM NaCl, 0.05% NP-40, 0.5 mM DTT, and 10% glycerol) containing 1 µg of poly(dI-dC) and 2µg of placental nuclear extracts or 1 µL of in vitro-translated proteins. After incubation at room temperature for 20 min, the reaction mixtures were analyzed by electrophoresis on 5% nondenaturing polyacrylamide gels. For competition experiments, the unlabeled oligonucleotides were used at a 100-fold molar excess and incubated with the in vitro-translated protein or nuclear extracts for 20 min on ice before addition of radiolabeled probes. For proteolytic-clipping EMSA (20), the protein-DNA complex was incubated with indicated amounts of proteases on ice for 10 min and the reaction mixtures were immediately analyzed as described above.
Oligonucleotides used in EMSA were m8, 5′- GGGCACT-GTGGGAACGGAAA; KBF, 5′ -TAGAAGATGGGGAATC-CCCA; β2m, 5′-AGATGATGGGAAAGTCCCTT; and κ light chain, 5′-CAGAGGGGACTTTCCGAGAG; FPIII and its mutant oligonucleotides were as listed in Table 1.
Cell Culture, Transfection, and Reporter Gene Assays. COS-1 and COS-7 cells were grown at 37 °C in HEPES-buffered Dulbecco’s modified minimal essential medium supplemented with 4% fetal bovine serum, 100 µg/mL streptomycin, and 100 units/mL penicillin. Cells in 30-mm culture plates were incubated with 2 mL of a calcium phosphate-DNA coprecipitate containing 3µg of reporter plasmids and various amounts of expression plasmids (as indicated in figure legends) as well as an pRcCMV empty
vector to maintain a constant amount of DNA in each transfection assay. To correct for transfection efficiency, 0.5 µg of pRSVβgal was used as an internal control. Cells were harvested in the reporter lysis buffer (Promega) 48 h after transfection. CAT activity was assayed by incubating total cellular proteins in the reporter lysis buffer plus 4 mM acetyl coenzyme A and 0.1µCi of [14C]chloramphenicol (21). The assay was routinely run for 1 h with the amount of extracts required to convert 0.5-50% of the substrate to the acety-lated forms. Assays outside this range were repeated with the appropriate amount of extracts. The acetylated com-pounds were separated from chloramphenicol by thin-layer chromatography (95% chloroform-5% methanol v/v) on silica gel 60 (Merck) and quantitated by the bio-imaging analyzer, BAS-1500 (Fujifilm, Japan). Luciferase activity was assayed with the luciferase assay system (Promega). The reaction was initiated by adding luciferin, and the light emission was monitored by a luminometer (Lumat LB 9501, EG & G Berthold, Germany). Specific CAT and luciferase activities were normalized by the internal β-galactosidase activity.β-Galactosidase enzymatic activities were assayed as described (16). Protein concentrations were measured with the BCA protein assay kit (Pierce, Rockford, IL).
RESULTS
Isolation and Characterization of the FPIII-Binding Fac-tors. To identify nuclear factors binding to the FPIII site of the rnCGM3 gene (Figure 1A), we performed an in vivo screening of a pACT2 human placental cDNA library using the yeast one-hybrid system. A human placental cDNA library was used because the expression patterns of the human and rat PSG genes are very similar and a human placental cDNA library is commercially available. Ap-proximately 5 × 106yeast colonies were screened and 10 positive clones were further characterized. Four of them encoded RBPJκ initiating at different nucleotides in the exon 2 region of the human RBPJκ gene (22). The other clones will be described elsewhere. Sequence analyses indicated that these RBPJκ clones encode N-terminally truncated polypep-tides containing the intact DNA-binding domain. To verify the specificity of the DNA-protein interaction in vivo, we reintroduced into the reporter yeast cells two of the RBPJκ clones, 6M1 and 6B3. In addition, a plasmid containing the mouse p53 gene, pGAD53m, and the parental vector pACT2 (containing the GAL4 activation domain, GAL4AD) were also introduced into yeast cells as negative controls. As shown in Figure 1B, colonies appeared only in the sectors with yeast cells containing the RBPJκ clones (6M1 and 6B3). Neither mouse p53 nor GAL4AD interacted with the FPIII element. These results demonstrated that GAL4AD-RBPJκ fusion protein specifically binds to the FPIII element and mediates HIS3 transcription in vivo.
Since the rodent counterpart of human RBPJκ gene has not been characterized, we used the human RBPJκ cDNA as a probe to screen a rat placentalλgt10 cDNA library. The isolated rat RBPJκ clone, rRBPJκ-2N, contained an open reading frame for a polypeptide of 487 amino acids. Sequence comparison indicated that the isolated clone shares 99% sequence homology with the murine RBPJκ-2N at the amino acid level (data not shown), confirming that RBPJκ gene is highly conserved among different species.
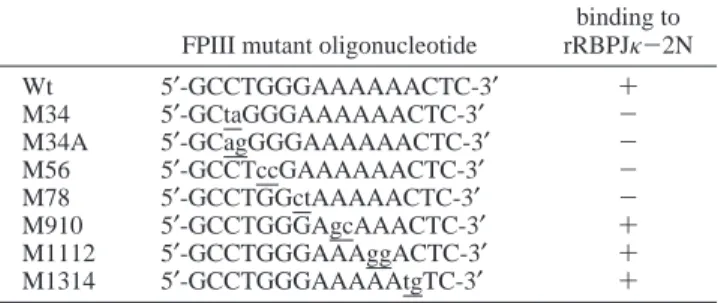
Table 1: Mutant Oligonucleotides Used for Competition Assaysa FPIII mutant oligonucleotide
binding to rRBPJκ-2N Wt 5′-GCCTGGGAAAAAACTC-3′ + M34 5′-GCtaGGGAAAAAACTC-3′ -M34A 5′-GCagGGGAAAAAACTC-3′ -M56 5′-GCCTccGAAAAAACTC-3′ -M78 5′-GCCTGGctAAAAACTC-3′ -M910 5′-GCCTGGGAgcAAACTC-3′ + M1112 5′-GCCTGGGAAAggACTC-3′ + M1314 5′-GCCTGGGAAAAAtgTC-3′ +
aEMSA was performed with labeled FPIII probe and the in vitro-translated rRBPJκ-2N. Individual mutant oligonucleotides were included at 100-fold molar excess as unlabeled competitors.
In Vitro Analysis of RBPJκ Interacting with the FPIII Element. To investigate whether RBPJκ directly binds to the FPIII element, we performed electrophoretic mobility shift assay (EMSA) with a32P-labeled FPIII probe and in vitro-translated rRBPJκ-2N protein and rat placental nuclear extracts. The estimated molecular mass of RBPJκ is 60 kDa (23) and the in vitro-translated rRBPJκ-2N we obtained approximates the predicted size (Figure 2A). As shown in Figure 2B, a DNA-protein complex was observed with both the in vitro-translated rRBPJκ-2N and the rat placental nuclear extracts (Figure 2B, lanes 2 and 9). Specific protein-DNA interaction was confirmed by the ability of an excess amount of unlabeled FPIII to block complex formation (lanes 3 and 10). To verify that the complex observed in the placental nuclear extracts was formed by RBPJκ, we tried to develop a working antiserum against rRBPJκ-2N but had no success. We therefore tried to verify the identity of the complex observed in the placental nuclear extracts using two other methods. First, we studied the competition efficiency of a series of oligonucleotides including m8, KBF, β-2m, and the κ light chain in EMSA. The m8 fragment, originally picked up as the naturally occurring candidate of the RBPJκ binding sequence from databases, was derived from the 5′ -flanking region of the Drosophila m8 gene, which is a member of the gene cluster Enhancer of split [E(spl)] (24, 25). The KBF,β-2m, and κ light-chain fragments, which all contain the NFκB binding sequence, were derived from the promoter/enhancer regions of the mouse major histocom-patibility complex (MHC) H-2Kb(26), theβ2-microglobulin (26), and the immunoglobulin κ light chain (27) genes, respectively. Depending on their sequence similarity to FPIII, these three oligonucleotides were expected to have different binding activities to RBPJκ and were purposely picked to be used as controls. As shown in Figure 2B, in both of the complexes formed with rRBPJκ-2N and the placental nuclear extracts, m8 andβ-2m had a higher potency than KBF and the κ light chain in blocking complex formation (Figure 2B, lanes 4, 6, 11, and 13). Second, we performed proteolytic-clipping band-shift assays using both protein sources. After
the DNA-protein complex formation, we treated the com-plexes with increasing amounts of trypsin. We observed two similar ladders of complexes of decreasing molecular weights from the two protein sources when the amount of trypsin was increased from 0.0005 to 0.5 unit (Figure 2C). The results suggested that the placental nuclear factor and rRBPJκ-2N have a similar conformation so that similar trypsin digestion patterns were observed. As a control, preincubation of proteins with 0.5 unit of trypsin completely abolished complex formation (Figure 2C, lanes 7 and 13). Similar results were obtained with chymotrypsin digestion (data not shown). It is not clear why we did not observe discrete bands resulted from partially digested RBPJκ-FPIII complex. However, Brou et al. (28) demonstrated that the essential DNA-binding domain of the human RBPJκ isoform, RBP3, encompasses the entire residues 7-435 region of the 486-amino acid polypeptide. Therefore, cleavage in the small regions outside the DNA-binding domain would likely result in only small differences in mobility of the complexes and might not be well resolved in our gel analysis (Figure 2C, lanes 3, 4, 9, and 10, arrows). Taken together, our data suggested that RBPJκ is a constitutive nuclear factor binding to the FPIII site of the rnCGM3 promoter in placenta.
To identify more specifically the nucleotides in the FPIII region important for rRBPJκ-2N binding, we performed competition experiments using in vitro-translated rRBPJκ-2N and a series of mutant oligonucleotides to compete with the target DNA (Table 1). Mutant oligonucleotides M34, M34A, M56, and M78 did not compete efficiently with FPIII for rRBPJκ-2N (Table 1, data not shown). These results indicated that the sequence CTGGGA in the FPIII region is the core binding sequence for rRBPJκ-2N.
Effect of RBPJκ and Notch on FPIII-Reporter Constructs. We next investigated the effect of RBPJκ on the expression of the FPIII-containing promoter construct p(FPIII)6SV40CAT, which contains six copies of FPIII site inserted in front of the SV40 promoter and the bacterial CAT gene coding sequence. Since RBPJκ is a downstream effector in the Notch signaling pathway, we examined CAT expression in the FIGURE2: Direct binding of RBPJκ to the FPIII site. (A) Product of in vitro-translated rRBPJκ-2N was monitored by incorporation of
[35S]methionine in the reaction mixtures, and the product was analyzed by SDS-PAGE and autoradiography. Numbers on the left indicate the protein molecular mass markers in kilodaltons. (B) RBPJκ EMSA with in vitro-translated rRBPJκ-2N and placental nuclear extracts. In vitro-translated rRBPJκ-2N (1µL) or 2 µg of rat placental nuclear extracts were incubated with radioactively labeled FPIII fragments in
EMSA. The DNA-protein complex formed is indicated by the arrow. The identity of the placental FPIII-binding factor was deduced to be an RBPJκ-like factor by competition assay with competitor olignucleotides m8, KBF,β-2m, and NF-κB, which are present at 100-fold
excess over the amount of the target DNA. Lanes 1 and 8 are reactions with free probe only. (C) Proteolytic-clipping assay of in vitro-translated rRBPJκ-2N- and placental nuclear protein-DNA complexes. EMSA was performed as described for panel (B) except that the protein-DNA complex was challenged with the indicated amount of trypsin (in units) for 10 min on ice before gel electrophoresis. As a control, in vitro-translated rRBPJκ-2N or placental nuclear protein was preincubated with 0.5 unit of trypsin for 10 min on ice before addition of FPIII probe (p.i. 0.5, lanes 7 and 13). Arrows indicate partially digested complexes. Lane 1 is the reaction with free probe only.
presence of rRBPJκ-2N or the active intracellular domain of Notch or both. The construct mNotchIC contains the active intracellular domain initiating at the 5′nuclear localization signal (NLS; TKKFRFEE) followed by the CDC10 domain and terminates at the 3′NLS (KGWLLDSS). As shown in Table 2, the CAT level in the presence of exogenous rRBPJκ-2N was reduced to 37% of the control. However, this repression by rRBPJκ-2N was counteracted by NotchIC. The stimulatory effect of NotchIC without rRBPJκ-2N was even more conspicuous and reached 24.21-fold over the control. The RBPJκ core sequence was required to achieve the stimulation effect observed with NotchIC since CAT levels directed by p(FPIIIM34)5SV40CAT and p(FPIIIM56)4 SV40-CAT, which contain the M34 and M56 mutations in the RBPJκ core sequence in the FPIII site (Table 1), were only 0.97- and 0.81-fold over the control, respectively (Table 2). These results suggested that Notch and RBPJκ may be involved in rnCGM3 expression through the FPIII site.
Notch Enhances the Transcriptional ActiVation of C/EBPβ
on the Natural rnCGM3 Promoter. Earlier studies on the rnCGM3 promoter indicated that C/EBPβ is a positive regulator that binds to the FPII site (14). Accordingly, interplay among C/EBPβ, RBPJκ and Notch may be involved in the modulation of rnCGM3 expression. To test this hypothesis, we placed nucleotides -326 to -33 of the 5′ -flanking region of rnCGM3 gene, which includes all three FP sites, in front of the luciferase gene in the plasmid pGL3-Basic (see Materials and Methods). The resultant rnCGM3 promoter construct was named pCGM3A. We examined the luciferase activities after cotransfecting pCGM3A with the plasmids encoding C/EBPβ, rRBPJκ-2N, or NotchIC into COS-7 cells, which are known for a low level of endogenous RBPJκ (29). As shown in Figure 3A, neither rRBPJκ-2N nor NotchIC significantly increased luciferase activity in the pCGM3A transfected COS-7 cells. Both proteins together also did not have any effect, whereas C/EBPβ increased Table 2: Effect of rRBPJκ-2N and NotchIC on FPIII- and FPI-Reporter Constructsa
construct sequence rRBPJκ-2N NotchIC fold induction p(FPIII)6SV40CAT (GCCTGGGAAAAAACTC) + - 0.37 ( 0.03
+ + 1.71 ( 0.20
- + 24.21 ( 1.48
p(FPIIIM34)5SV40CAT (GCTAGGGAAAAAACTC) - + 0.97 ( 0.01
p(FPIIIM56)4SV40CAT (GCCTCCGAAAAAACTC) - + 0.81 ( 0.08
p(FPI)6SV40CAT (TGTCATCCCTGGGATGAGTGTT) - + 9.90 ( 0.90
p(FPIM34A)6SV40CAT (TGTCATCCAGGGGATGAGTGTT) - + 1.12 ( 0.23
aCOS-1 cells were transfected with 3µg of FPIII- or FPI-reporter construct and 3 µg of pCMVrRBPJκ-2N or 1 µg of mNotchIC or both. Cells were harvested as described under Materials and Methods. Specific CAT activities directed by p(FPIII)6SV40CAT, p(FPIIIM34)5SV40CAT,
p(FPIIIM56)4SV40CAT, p(FPI)6SV40CAT, and p(FPIM34A)6SV40CAT were 0.086 ( 0.001, 0.101 ( 0.008, 0.11 ( 0.006, 0.07 ( 0.004, and
0.115 ( 0.008 nmol min-1(mg of protein)-1, respectively. Fold induction was calculated relative to the CAT expression observed in the absence of rRBPJκ-2N or mNotchIC expression plasmids for individual reporter plasmids. Mean values (SEM from three independent transfection experiments are provided.
FIGURE3: Functional interaction between mNotchIC- and C/EBPβ-stimulated luciferase expression directed by the rnCGM3 promoter
construct. (A) The rnCGM3 promoter-luciferase construct, pCGM3A, containing FPI, -II, and -III sites was generated in the promoterless and enhancerless pGL3-Basic plasmid as described under Materials and Methods. COS-7 cells were transfected with 3µg of pCGM3A
alone or plus an expression plasmid encoding either rRBPJκ-2N, C/EBPβ, or mNotchIC or various combinations of these plasmids. The
amounts (in micrograms) of expression plasmids used in each transfection are listed along the x-axis. Fold induction was calculated relative to the luciferase expression observed for pCGM3A alone. Mean values and SEM obtained from four independent transfection experiments are plotted. (B) Transient expression assays with an FPIII-deleted construct, pCGM3A-∆FPIII. The scheme for transfection was the same
as described for panel (A). Mean values and SEM obtained from four independent transfection experiments are plotted. The luciferase activities directed by pCGM3A and pCGM3A-∆FPIII were 825.75 ( 44.14 and 498.33 ( 29.06 relative light units (RLU)/µg of protein,
luciferase activity by 4.6-fold. The transactivation by C/EBPβ was decreased to 2.2-fold when it was cotransfected with 2 µg of the plasmid encoding rRBPJκ-2N. However, this repression was counteracted by mNotchIC in a dose-dependent manner. Interestingly, transactivation by C/EBPβ was further enhanced to 8.0- or 19.2-fold with 1 or 3µg of mNotchIC (Figure 3A). To examine if the stimulatory effect of NotchIC depends on the FPIII element, we cotransfected pCGM3A-∆FPIII, which is an FPIII-deleted pCGM3A construct, with plasmids encoding C/EBPβ, rRBPJκ-2N, or NotchIC. To our surprise, a similar profile of promoter activities was observed (Figure 3B), only the stimulation by NotchIC was lower than in the full-length promoter (11.96-fold versus 19.21-(11.96-fold). These data suggested that there might be another functional RBPJκ site, which is responsive to NotchIC, present in the native rnCGM3 promoter.
We inspected the rnCGM3 promoter region from nucle-otides -326 to -33 and identified another RBPJκ core sequence (CTGGGA) within the FPI site (Figure 1A, FPI, underlined). To test whether this FPI-RBPJκ core sequence contributes to the NotchIC activation on the pCGM3A-∆FPIII promoter construct mentioned above, we first per-formed transient expression assays with the p(FPI)6SV40CAT or the p(FPIM34A)6SV40CAT reporter plasmid and mNotch-IC. The p(FPI)6SV40CAT and p(FPIM34A)6SV40CAT constructs contain six copies of the FPI site and six copies of the FPIM34A mutant site (Table 2), respectively, inserted in front of the SV40 promoter and the bacterial CAT gene coding sequence. As shown in Table 2, CAT levels directed by p(FPI)6SV40CAT but not p(FPIM34A)6SV40CAT were stimulated by mNotchIC (9.9- versus 1.1-fold). These data suggested that the RBPJκ core sequence in the FPI site was responsible for NotchIC transactivation on the SV40 pro-moter. EMSA also revealed a specific binding between FPI and the in vitro-translated rRBPJκ-2N (data not shown).
We next verified the effect of the FPIII RBPJκ-binding site on the rnCGM3 promoter without the contribution from the FPI site. We made the construct pCGM3B, which consists of the rnCGM3 promoter region of nucleotides -283 to -33, containing the FPII (-263 to -233) and the FPIII (-108 to -93) sites, fused to the luciferase reporter gene. As shown in Figure 4, similar to results observed with pCGM3A, the luciferase levels directed by pCGM3B were positively regulated by C/EBPβ, repressed by rRBPJκ-2N, and further enhanced by NotchIC (Figure 4, left). Furthermore, this positive effect of NotchIC was not observed with pCGM3B-FPIIIM34A (Figure 4, middle), indicating that the FPIII RBPJκ-binding site was also required for the transcriptional activation of NotchIC. To confirm that the stimulatory effect of NotchIC was mediated only through the RBPJκ-binding sites in FPI and FPIII, we further introduced the M34A mutation into the RBPJκ core sequence (CTGGGA to AGGGGA, Tables 1 and 2) in both sites in pCGM3A and the resultant reporter construct pCGM3A-FPI&IIIM34A was used for transient expression assays. As shown in the right panel of Figure 4, the stimulatory effect of NotchIC was completely abolished. Taken together, both RBPJκ-binding sites in FPI and FPIII contributed to the NotchIC enhance-ment of the rnCGM3 promoter activities transactivated by C/EBPβ. These results also indicate a functional interaction between C/EBPβ and NotchIC.
DISCUSSION
We performed a yeast one-hybrid screening and identified RBPJκ as a nuclear factor recognizing the FPIII site in the rnCGM3 promoter. Several lines of evidence support that expression of rnCGM3 is mediated by RBPJκ in the Notch signaling pathway. First, rRBPJκ-2N repressed expression of the heterologous SV40 promoter via FPIII and this repression was counteracted by NotchIC (Table 2). Second, the RBPJκ core sequence (CTGGGA) in the FPIII and FPI sites was required for NotchIC to transactivate the SV40 promoter. This positive effect of NotchIC was abolished when residues in the RBPJκ core sequence were altered. It is unlikely that ligation of multiple copies of FPIII and FPI oligonucleotides fortuitously generated RBPJκ-binding sites in our reporter constructs because the mutant constructs also contain the same flanking sequences. Third, transcriptional activation by C/EBPβ on the rnCGM3 promoter was repressed by rRBPJκ-2N but significantly enhanced by NotchIC. Fourth, both the FPI and FPIII RBPJκ-binding sites in the rnCGM3 promoter were functional in the Notch signaling pathway because the enhancement effect of Notch-IC on C/EBPβ could be accomplished by either site, and when both sites were mutated, the enhancement effect was abolished. Taken together, we propose that expression of the rnCGM3 gene is upregulated by C/EBPβ via the FPII site and by RBPJκ/Notch via the FPI and FPIII sites. This also suggests that rnCGM3 is likely a target gene of the Notch signaling pathway, which may provide another mechanism to control PSG expression in the placenta in addition to regulations by C/EBPβ.
RBPJκ is identical to the cellular factor CBF1 that binds to the Epstein-Barr virus (EBV) C promoter (30) and the nuclear factor KBF2 that binds to the enhancer of mouse FIGURE4: Both FPIII and FPI RBPJκ-binding sites are functional for supporting the stimulatory effect of NotchIC. COS-7 cells were transfected with 3 µg of the various promoter constructs, as
indicated above each group of data, or plus the indicated amount of expression plasmid containing various combinations of rRBPJκ-2N, C/EBPβ, or mNotchIC. Mean values and SEM obtained from
three independent transfection experiments are plotted. The lu-ciferase activities directed by pCGM3B, pCGM3B-FPIIIM34A, and pCGM3A-FPI&IIIM34A were 496.5 ( 143.72, 267.83 ( 44.87, and 423.02 ( 35.9 RLU/µg of protein, respectively.
MHC class I genes (28). The Drosophila homologue of RBPJκ gene is the Suppressor of Hairless [Su(H)] gene, which is involved in the peripheral nervous system develop-ment (31,32). RBPJκ can associate with a variety of proteins including the Drosophila hairless protein (28), the EBV nuclear antigen 2 (EBNA2) (33-36), the intracellular domain of Notch protein (NotchIC) (37, 38), and a recently isolated LIM protein, KyoT2 (39). Neither EBNA2 nor NotchIC binds directly to DNA. Upon association with RBPJκ, RBPJκ/EBNA2 and RBPJκ/NotchIC complexes activate target genes through the RBPJκ site. In addition, RBPJκ interacts with two transcriptional coactivators, dTAFII110 (a subunit of TFIID) and TFIIA, to repress transcription (40). Recently, Hsieh et al. (41) reported another RBPJκ-interact-ing protein called CIR (CBF1 interactRBPJκ-interact-ing corepressor), which interacts with a histone deacetylase (HDAC) and another corepressor, SAP30. Therefore, association between RBPJκ and CIR may further recruit HDAC and other corepressor to modify chromatin structure and diminish transcriptional activities (41).
The promoter of the mouse Hairy enhancer of split (HES-1) gene contains two adjacent RBPJκ-binding sites centered at nucleotide -81 and -67, respectively, from the transcrip-tion start site (42). However, the two sites are not functranscrip-tionally equivalent because a mutation in the distal RBPJκ site abolishes activation by NotchIC, whereas a mutation in the proximal site does not affect it (43). In the present study, we characterized, in the rnCGM3 promoter, two separate RBPJκ-binding sites that are upstream and downstream, respectively, from the transcription start site. Unlike the two RBPJκ-binding sites in HES-1, the two RBPJκ-binding sites in rnCGM3 are functionally equivalent because mutations in either site do not abolish the positive effect of NotchIC. rRBPJκ-2N repressed the transcriptional activation of C/EBPβ on the rnCGM3 promoter. It is possible that rRBPJκ-2N associates with transcriptional coactivators such as TFIIA, which interferes with the interaction between C/EBPβ and the basal transcriptional machinery. rRBPJκ-2N could also interact with protein like CIR, which results in chromatin modification and reduces the transcriptional activation by C/EBPβ. These RBPJκ-associated factors may be displaced by NotchIC in mNotchIC-transfected cells. The resultant positive regulator complex RBPJκ/NotchIC further enhances the transcriptional activation of C/EBPβ on the
rnCGM3 promoter.
Neither rRBPJκ-2N, NotchIC, nor both proteins together had a significant effect on the expression of a native rnCGM3 promoter. Instead, the regulatory functions of rRBPJκ-2N and NotchIC were manifested in the presence of the FPII-binding factor C/EBPβ. This phenomenon is reminiscent of the context-dependent regulation of the T-cell receptor (TCR) R enhancer by LEF-1/TCF-1R. In this case, a specific arrangement and occupancy of the CRE- and TCF-2 protein binding sites surrounding the LEF-1/TCF-1R binding site is required for the transcriptional activation of LEF-1/TCF-1R (44). It is possible that preoccupancy of the FPII site by C/EBPβ in the rnCGM3 promoter provides a permissive condition for an effective Notch signaling pathway. We have performed yeast and mammalian two-hybrid assays (45, 46) using C/EBPβ and NotchIC and failed to detect any physical interaction between the two factors (data not shown). Additional studies are required to elucidate this functional
interaction between C/EBPβ and the Notch signaling path-way.
Interestingly, we observed that the stimulatory effect of NotchIC is stronger in the absence of exogenous rRBPJκ-2N. However, it has been shown recently that the transac-tivation activity of NotchIC is completely dependent on RBPJκ because the activity was not observed in an RBPJκ null cell line (47). It is possible that the endogenous level of RBPJκ in COS-7 is sufficient for NotchIC to enhance the transcriptional activity of C/EBPβ. This enhancement effect of NotchIC may depend on the ratio between the amounts of NotchIC and RBPJκ in the nucleus. An excess amount of exogenous RBPJκ may reduce this enhancement effect.
We have used the COS7 cell line instead of a placental cell line. A placental cell line may provide a more natural condition for transient expression assays. However, some-times the specific effects of transfected expression vectors are complicated by the existence of endogenous proteins. In comparison, in a nonplacental cell line, it may be easier to observe specific effects of the transfected expression vectors. The COS7 cells have been widely used in Notch and RBPJκ studies (39, 48, 49). In particular, this cell line contains a low level of endogenous RBPJκ, which is critical to our characterization of the specific role of RBPJκ. Moreover, the observation that C/EBPβ stimulated rnCGM3 promoter activity in COS7 cells is similar to our previous observation in a human placental cell line, HPA1 (14). Therefore, the COS7 cell line should be a valid starting material to study the interplay between C/EBPβ, NotchIC, and RBPJκ.
Using transient transfection of promoter-reporter con-structs and EMSA, we have obtained a basic framework of interaction between the three factors. However, these simpler experimental systems have their limitations. For example, the chromosomal organization of rnCGM3 gene in the living cell may play a potential role in gene activation. It has been shown that chromatin remodeling regulates the yeast PHO5 and the mouse mammary tumor virus promoter activities (50, 51). Given that the enhancement effect of NotchIC on C/EBPβ is context-dependent with the naked DNA construct, it is not clear whether these factors are accessible to their binding sites in the chromosomal context of rnCGM3. This issue deserves more attention in future studies.
One of the phenotypes in RBPJκ knockout mice is placental failure (52). It will be interesting to investigate whether silencing of the RBPJκ/Notch-regulated genes, e.g., PSG genes, contributes to the abnormal placental develop-ment in RBPJκ knockout mice. In addition, Nakayama et al. (53) reported that Notch is involved in trophoblast differentiation. Since PSGs are expressed in the giant cell/ spongiotrophoblast of the rodent placenta, additional inves-tigation is warranted to determine whether rnCGM3 expres-sion is correlated with the Notch-mediated trophoblast differentiation.
ACKNOWLEDGMENT
We thank Dr. R. Kopan for gifts of cDNA for mNotchIC, Dr. Hsou-min Li for critical reading of the manuscript, and Yung-kang Lee for technical support.
REFERENCES
1. Chou, J. Y., and Plouzek, C. A. (1992) Semin. Reprod. Endocrinol. 10, 116-126.
2. Lin, T. M., Halbert, S. P., and Spellacy, W. N. (1974) J. Clin. InVest. 54, 576-582.
3. MacDonald, D. J., Scott, J. M., Gemmell, R. S., and Mack, D. S. (1983) Am. J. Obstet. Gynecol. 147, 430-436. 4. Mantzavinos, T., Phocas, I., Chrelias, H., Sarandakou, A., and
Zourlas, P. A. (1991) Eur. J. Obstet. Gynecol. Reprod. Biol. 39, 117-122.
5. Masson, G. M., Anthony, F., and Wilson, M. S. (1983) Br. J. Obstet. Gynaecol. 90, 146-149.
6. Tamsen, L., Johansson, S. G., and Axelsson, O. (1983) J. Perinat. Med. 11, 19-25.
7. Bohn, H., and Weinmann, E. (1974) Arch. Gynakol. 217, 209-218.
8. Hau, J., Gidley-Baird, A. A., Westergaard, J. G., and Teisner, B. (1985) Biomed. Biochim. Acta 44, 1255-1259.
9. Rutherfurd, K. J., Chou, J. Y., and Mansfield, B. C. (1995) Mol. Endocrinol. 9, 1297-1305.
10. Rebstock, S., Lucas, K., Thompson, J. A., and Zimmermann, W. (1990) J. Biol. Chem. 265, 7872-7879.
11. Chen, H., Chen, C. L., and Chou, J. Y. (1994) Biochemistry 33, 9615-9626.
12. Chen, H., Plouzek, C. A., Liu, J. L., Chen, C. L., and Chou, J. Y. (1992) DNA Cell Biol. 11, 139-148.
13. Kromer, B., Finkenzeller, D., Wessels, J., Dveksler, G., Thompson, J., and Zimmermann, W. (1996) Eur. J. Biochem. 242, 280-287.
14. Chen, H., Lin, B., Chen, C. L., Johnson, P. F., and Chou, J. Y. (1995) DNA Cell Biol. 14, 681-688.
15. Wang, M. M., and Reed, R. R. (1993) Nature 364, 121-126. 16. Ausubel, F. M., Brent, R., Kingston, R. E., Moore, D. D., Seidman, J. G., Smith, J. A., and Struhl, K. (1992) Current Protocols in Molecular Biology, Greene Publishing Associates/ Wiley-Interscience, New York.
17. Williams, S. C., Cantwell, C. A., and Johnson, P. F. (1991) Genes DeV. 5, 1553-1567.
18. Kopan, R., Schroeter, E. H., Weintraub, H., and Nye, J. S. (1996) Proc. Natl. Acad. Sci. U.S.A. 93, 1683-1688. 19. Ohlsson, H., and Edlund, T. (1986) Cell 45, 35-44. 20. Laux, G., Adam, B., Strobl, L. J., and Moreau-Gachelin, F.
(1994) EMBO J. 13, 5624-5632.
21. Fordis, C. M., and Howard, B. H. (1987) Methods Enzymol. 151, 382-397.
22. Amakawa, R., Jing, W., Ozawa, K., Matsunami, N., Hamagu-chi, Y., Matsuda, F., KawaiHamagu-chi, M., and Honjo, T. (1993) Genomics 17, 306-315.
23. Matsunami, N., Hamaguchi, Y., Yamamoto, Y., Kuze, K., Kangawa, K., Matsuo, H., Kawaichi, M., and Honjo, T. (1989) Nature 342, 934-937.
24. Klambt, C., Knust, E., Tietze, K., and Campos-Ortega, J. A. (1989) EMBO J. 8, 203-210.
25. Tun, T., Hamaguchi, Y., Matsunami, N., Furukawa, T., Honjo, T., and Kawaichi, M. (1994) Nucleic Acids Res. 22, 965-971.
26. Israel, A., Kimura, A., Kieran, M., Yano, O., Kanellopoulos, J., Le Bail, O., and Kourilsky, P. (1987) Proc. Natl. Acad. Sci. U.S.A. 84, 2653-2657.
27. Sen, R., and Baltimore, D. (1986) Cell 46, 705-716. 28. Brou, C., Logeat, F., Lecourtois, M., Vandekerckhove, J.,
Kourilsky, P., Schweisguth, F., and Israel, A. (1994) Genes DeV. 8, 2491-2503.
29. Hamaguchi, Y., Yamamoto, Y., Iwanari, H., Maruyama, S., Furukawa, T., Matsunami, N., and Honjo, T. (1992) J. Biochem. (Tokyo) 112, 314-320.
30. Dou, S., Zeng, X., Cortes, P., Erdjument-Bromage, H., Tempst, P., Honjo, T., and Vales, L. D. (1994) Mol. Cell Biol. 14, 3310-3319.
31. Furukawa, T., Maruyama, S., Kawaichi, M., and Honjo, T. (1992) Cell 69, 1191-1197.
32. Schweisguth, F., and Posakony, J. W. (1992) Cell 69, 1199-1212.
33. Grossman, S. R., Johannsen, E., Tong, X., Yalamanchili, R., and Kieff, E. (1994) Proc. Natl. Acad. Sci. U.S.A. 91, 7568-7572.
34. Henkel, T., Ling, P. D., Hayward, S. D., and Peterson, M. G. (1994) Science 265, 92-95.
35. Waltzer, L., Logeat, F., Brou, C., Israel, A., Sergeant, A., and Manet, E. (1994) EMBO J. 13, 5633-5638.
36. Zimber-Strobl, U., Strobl, L. J., Meitinger, C., Hinrichs, R., Sakai, T., Furukawa, T., Honjo, T., and Bornkamm, G. W. (1994) EMBO J. 13, 4973-4982.
37. Lu, F. M., and Lux, S. E. (1996) Proc. Natl. Acad. Sci. U.S.A. 93, 5663-5667.
38. Tamura, K., Taniguchi, Y., Minoguchi, S., Sakai, T., Tun, T., Furukawa, T., and Honjo, T. (1995) Curr. Biol. 5, 1416-1423. 39. Taniguchi, Y., Furukawa, T., Tun, T., Han, H., and Honjo, T.
(1998) Mol. Cell Biol. 18, 644-654.
40. Olave, I., Reinberg, D., and Vales, L. D. (1998) Genes DeV. 12, 1621-1637.
41. Hsieh, J. J., Zhou, S., Chen, L., Young, D. B., and Hayward, S. D. (1999) Proc. Natl. Acad. Sci. U.S.A. 96, 23-28. 42. Takebayashi, K., Sasai, Y., Sakai, Y., Watanabe, T., Nakanishi,
S., and Kageyama, R. (1994) J. Biol. Chem. 269, 5150-5156. 43. Jarriault, S., Brou, C., Logeat, F., Schroeter, E. H., Kopan,
R., and Israel, A. (1995) Nature 377, 355-358.
44. Carlsson, P., Waterman, M. L., and Jones, K. A. (1993) Genes DeV. 7, 2418-2430.
45. Chien, C. T., Bartel, P. L., Sternglanz, R., and Fields, S. (1991) Proc. Natl. Acad. Sci. U.S.A. 88, 9578-9582.
46. Fearon, E. R., Finkel, T., Gillison, M. L., Kennedy, S. P., Casella, J. F., Tomaselli, G. F., Morrow, J. S., and Van-Dang, C. (1992) Proc. Natl. Acad. Sci. U.S.A. 89, 7958-7962. 47. Kato, H., Taniguchi, Y., Kurooka, H., Minoguchi, S., Sakai,
T., Nomura-Okazaki S., Tamura, K., and Honjo, T. (1997) DeVelopment 124, 4133-4141.
48. Kannabiran, C., Zeng, X., and Vales, L. D. (1997) Mol. Cell. Biol. 17, 1-9.
49. Oswald, F., Liptay, S., Adler, G., and Schmid, R. M. (1998) Mol. Cell. Biol. 18, 2077-2088.
50. Fascher, K. D., Schmitz, J., and Horz, W. (1993) J. Mol. Biol. 231, 658-667.
51. Lee, H. L., and Archer, T. K. (1994) Mol. Cell. Biol. 14, 32-41.
52. Oka, C., Nakano, T., Wakeham, A., de la Pompa, J. L., Mori, C., Sakai, T., Okazaki, S., Kawaichi, M., Shiota, K., Mak, T. W., and Honjo, T. (1995) DeVelopment 121, 3291-3301. 53. Nakayama, H., Liu, Y., Stifani, S., and Cross, J. C. (1997)
DeV. Genet. 21, 21-30. BI991786K