行政院國家科學委員會專題研究計畫 成果報告
二性次微米聚合體粒子之合成與鑑定以及其和蛋白質的交 互作用(3/3)
計畫類別: 個別型計畫
計畫編號: NSC94-2216-E-011-002-
執行期間: 94 年 08 月 01 日至 95 年 07 月 31 日 執行單位: 國立臺灣科技大學化學工程系
計畫主持人: 陳崇賢
報告類型: 完整報告
處理方式: 本計畫可公開查詢
中 華 民 國 95 年 8 月 11 日
行政院國家科學委員會專題計畫期末進度報告
二性次微米聚合體粒子之合成與鑑定以及和蛋白質的交互作用(3/3)
計畫編號:NSC93-2216-E-011-005 執行期限:94/08/01 ~ 95/07/31
計畫主持人:陳崇賢 國立台灣科技大學 中文摘要
本研究探討使用已知等電點(IEP)之二性 PMMA 次微米粒子(PMMA1 之 IEP = 5.2、PMMA2 之 IEP = 7.1)與蛋白質在不同 pH 環境下作用以決定未知蛋 白質pI 值之可行性。本研究以 PMMA1-lysozyme(pI = 11)、PMMA2-pepsin
(pI = 1)以及 PMMA2-BSA(pI = 5)配對作為研究對象。對於 PMMA1-
lysozyme 及 PMMA2-pepsin 配對而言,皆因為 PMMA1 與 lysozyme 及 PMMA2 與pepsin 的等電點差距頗大,因此在 UV/visible 吸收值(560 nm)-pH 數據皆 呈現兩個明顯的波峰,其中一個波峰出現在蛋白質的等電點(pI)附近,而另一 個波峰則出現在二性乳漿粒子的等電點(IEP)附近。反之,在 PMMA2-BSA 配
對的實驗中,因為兩者的等電點相當接近,因此在UV/visible 吸收值(560 nm)
-pH 的圖形中可以發現在 BSA 等電點附近僅出現單一波峰。二性乳漿粒子與蛋 白質分子之間的靜電交互作用機構可以解釋所觀察膠體現象。
Abstract
The feasibility of using single amphoteric latex of known isoelectric point (IEP) (PMMA1 with IEP = 5.2 or PMMA2 with IEP = 7.1) to interact with a protein sample at various pH values to determine the pI value of the protein was investigated. The pairs of PMMA1-lysozyme (pI = 11), PMMA2-pepsin (pI = 1) and PMMA2-BSA (pI
= 5) were chosen for study. Both the pairs of PMMA1-lysozyme and PMMA2-pepsin with a large difference between the IEP of the amphoteric latex particle in the absence of protein and the pI of the model protein showed two distinct characteristic peaks in the absorbance of colloidal particles in the presence of protein at 560 nm versus pH data; one corresponded to the pI of the protein sample and the other presumably originated from the amphoteric nature of the latex particles. On the other hand, only one characteristic peak was observed in the absorbance of colloidal particles in the presence of protein at 560 nm versus pH data for the PMMA2-BSA pair. This was attributed to the rather small difference between the IEP of the amphoteric latex particle in the absence of protein and the pI of the model protein. This characteristic peak was assigned to the pI of the protein sample. The mechanisms involved in the electrostatic interactions between the amphoteric latex particles and protein molecules were proposed to explain the observed colloidal phenomena.
Key words Amphoteric latexes - Polymethyl methacrylate – Adsorption of
proteins – Electrostatic interactions - Colloidal stability.Introduction
Submicron polymer particles exhibiting an exceedingly large particle surface area per unit mass of polymer have been evaluated as a potential polymeric support for purification of protein from a crude biological mixture.
Adsorption of protein onto the particle surface via the electrostatic interaction is the most widely used recovery method.
A variety of positively charged or negatively charged polymer particles were synthesized and used to bind proteins.
1-12
Homola and James13
prepared and characterized amphoteric polystyrene (PS) latexes. Recently, Shirahama et al.14
and Fang and Kawaguchi15
adopted different synthesis routes to prepare amphoteric polymer particles and then studied their interactions with biomolecules. The literature related to the interactions between the amphoteric latex particles and proteins is rather limited.In our previous work,
16
a series of amphoteric latexes were synthesized by the semibatch surfactant-free emulsion copolymerization of methyl methacrylate (MMA) and methacrylic acid (MAA) initiated by 2,2’-azobis(2-amidinopropane)dihydrochloride (V-50). The relatively hydrophilic polymethyl methacrylate (PMMA) was expected to give a smaller extent of protein adsorption via the nonspecific hydrophobic interaction compared to the PS counterpart. At low pH, these particles had a net positive
charge due to the ionized amino group originating from V-50. At high pH, they became negatively charged due to the ionized carboxyl group originating from MAA. Thus, there existed a pH at which point these particles exhibited a net charge of zero (isoelectric point, IEP).
The electrostatic interactions between these amphoteric particles and various proteins were then investigated
17
The nature and degree of interactions between the particles and proteins were primarily controlled by the electrostatic characteristics of the polymeric supports and proteins under the experimental conditions. It was also proposed that the isoelectric point (pI) of an unknown protein sample could be determined based on the unique colloidal instability phenomenon caused by the electrostatic attraction forces between the particles and protein molecules. The objective of this work was therefore to study the feasibility of using single amphoteric latex of known IEP to determine the pI value of protein. The results obtained from this work may also contribute to the fundamental understanding of the complicated interactions between the particles and proteins.Experimental Materials
Two monodisperse amphoteric PMMA latexes with similar particle sizes and particle size distributions but different IEP values (abbreviated as PMMA1 and PMMA2) were prepared, purified and
characterized according to Ref. 16.
Some physical properties of the purified latex products are listed in Table 1. The chemicals used in the synthesis work included MMA (Kaohsiung Monomer), MAA (Mitsubishi Rayon), V-50 (95%, Wako Pure Chemical Industries), glycine (99%, Wako Pure Chemical Industries), HCl (Nacalai Tesque), N
2
(Ching-Feng-Harng), deionized water (Barnsted, Nanopure Ultrapure Water System, specific conductance < 0.057 S/cm). Other chemicals used in this study were lysozyme (Sigma), bovine serum albumin (BSA, Sigma), pepsin (Sigma), NaOH (Riedel-de Haen), and NaCl (Riedel-de Haen). MMA was distilled under reduced pressure before use. All other chemicals were used as received.
Electrostatic interactions between latex particles and proteins.
First, the pH of the aqueous
solution of protein (500 mg/dm
3
) and the amphoteric latex was adjusted by 0.1 N HCl and 0.1 N NaOH. The total solids content of the latex particles was kept constant at 0.01% and the ionic strength of the mixture of the latex particles and protein was kept constant at 2 mM NaCl throughout this work. The latex sample with a volume of 3 cm3
was then mixed with an equal volume of the protein solution. The mixture of the latex particles and protein was mixed using a magnetic stirrer at 25 ℃ for 4 h. The dynamic protein adsorption data confirmed that four hours weresufficient to assure the equilibrium condition.
18
The zeta potential (ζ), average hydrodynamic particle diameter (dp
) and absorbance at 560 nm of the latex particles with or without adsorbed protein molecules were determined by a zeta potential meter (Malvern,Zetamaster), DLS (Otsuka, Photal LPA-3000/3100) and a
spectrophotometer (Shimadzu, UV-1601), respectively. The dilution water used for the measurements of ζ and d
p
had the same ionic strength and pH as the latex sample. The ζ and dp
data reported in this work were the average of at least ten and three measurements, respectively.
Result and Discussion
Interactions between PMMA1 and Lysozyme
For reference, the ζ of the
amphoteric latex particles PMMA1 and PMMA2 in the absence of protein as a function of pH are shown in Fig. 1. Both the data of d
p
and the absorbance of latex particles at 560 nm show distinct characteristic peaks corresponding to the IEP values of PMMA1 and PMMA2 (Fig. 2). These results suggest that significant particle flocculation occurs around the IEP of the PMMA1 or PMMA2 particles due to the diminished electrostatic repulsion force between two approaching particles. Thus, only the measurements of the absorbance of the colloidal systems at 560 nm will be presented hereinafter to qualitativelydetermine whether the network structure of aggregates forms in the protein adsorption process or not.
The data of the absorbance at 560 nm and ζ of the PMMA1 particles in the presence of lysozyme are shown in Fig.
3. The difference between the IEP of the PMMA1 particle (5.2) and the pI of the model protein lysozyme (11) is 5.8.
Although not very distinct, two characteristic peaks can be identified in the absorbance of the PMMA1 particles in the presence of lysozyme at 560 nm versus pH data; one corresponds to the pI of lysozyme and the other presumably originates from the amphoteric nature of the PMMA1 particles (Fig. 3a). At pH <
5.2, both the PMMA1 particles and lysozyme molecules are positively charged and, therefore, the exceedingly retarded adsorption of lysozyme molecules on the PMMA1 particle surfaces is experienced due to the strong electrostatic repulsion force between the PMMA1 particle and lysozyme. As pH approaches 5.2, the greatly reduced ζ induces the flocculation of colloidal particles, thereby leading to the rapid increase of the absorbance at 560 nm. It should be noted that the point at which the first maximal absorbance at 560 nm occurs is shifted to a pH value (ca. 5.5) that is slightly higher than the IEP of the PMMA1 particles in the absence of protein. This is most likely due to the adsorbed lysozyme molecules that impart additional positive charges to the PMMA1 particle surfaces. This is
further supported by comparing the ζ versus pH data in the absence and presence of lysozyme in Figs. 1 and 3b, respectively. After the first maximal absoebance at 560 nm is reached, the colloid systems in the pH range ca. 5.5-9 are still unstable as a consequence of the following two competitive processes.
First, in the absence of protein, charge reversal of the PMMA1 particles (+ →
- ) occurs at pH 5.2 and then the∣ζ∣of the PMMA1 particles increases with increasing pH (Fig. 1).
However, adsorption of the positively charged lysozyme molecules onto the negatively charged PMMA1 particles (i.e., charge neutralization) becomes more important when pH increases from 5.5 to 9. These two comparable effects counterbalance with each other and make the charge reversal of colloidal particles in the presence of lysozyme to occur at a higher pH value (Fig. 3b). In addition, the ζ data shown in Fig. 3b also indicate that both the electrostatic repulsion (5.5 < pH < 7) and attraction (7 < pH < 9) forces between the PMMA1 particle and lysozyme are rather weak in this region. These unique characteristics of the colloidal particles in the presence of lysozyme are then reflected in the data of the absorbance at 560 nm (Fig. 3a).
In the pH range 9-10, the absorbance of colloidal particles in the presence of lysozyme at 560 nm increases again to the second maximum due to the significant charge
neutralization of the negatively charged PMMA1 particles by adsorption of the positively charged lysozyme molecules (Fig. 3). Beyond pH 10, PMMA1 particles become even more negatively charged, whereas the net positive charge of lysozyme decreases continuously as the pH approaches their pI value. Under the circumstance, the electrostatic repulsion forces between two approaching PMMA1 particles and between the interactive PMMA1 particle and lysozyme are strong enough to stabilize the colloidal systems (Fig. 3b) and, hence, the absorbance of colloidal particles in the presence of lysozyme at 560 nm decreases to a level comparable to that at pH 3 (Fig. 3a).
Interactions between PMMA2 and Pepsin.
The data of the absorbance at 560 nm and ζ of the PMMA2 particles in the presence of pepsin are shown in Fig. 4.
The difference between the IEP of the PMMA2 particle in the absence of protein (7.1) and the pI of the model protein pepsin (1) is 6.1. Figure 4a clearly shows two distinct characteristic peaks in the absorbance of colloidal particles in the presence of pepsin at 560 nm versus pH data; one corresponds to the pI of pepsin and the other to the contribution of the PMMA2 particles. It should be noted that the point at which the second maximal absorbance occurs is quite far away from the IEP of the PMMA2 particles in the absence of pepsin (cf. Figs. 2b and 4a). This is
closely related to the electrostatic interactions between the PMMA2 particle and pepsin at different pH values, as illustrated by the ζ data obtained from the colloidal systems in the absence and presence of pepsin in Figs. 1 and 4b, respectively. It is also interesting to note that these two maximal absorbance values at 560 nm correspond exactly to the pH values with ζ close to 0 (Fig. 4).
In the pH range 1-7.1, the PMMA2 particles in the absence of protein carry net positive charge and the particle surface charge density first remains relatively constant and then decreases with increasing pH (Fig. 1). On the other hand, pepsin is negatively charged and the surface charge density increases with increasing pH. Thus, charge neutralization of the positively charged PMMA2 particles by adsorption of the negatively charged pepsin molecules plays an important role in the adsorption process. Furthermore, there exists a pH at which point the maximal degree of charge neutralization occurs. This is the major reason why the strongest electrostatic attraction force between the PMMA2 particle and pepsin (i.e., the second maximal absorbance at 560 nm) at a pH value between 1 and 7.1 is observed (Fig. 4). After the second maximum is achieved, the continuously decreased absorbance at 560 nm can be attributed to the decreased charge neutralization with pH, as evidenced by the increased∣ζ∣with pH in Fig. 4b.
Beyond pH 7.1, both the PMMA2 particles in the absence of pepsin and pepsin molecules carry negative charges (Fig. 1). Thus, the electrostatic repulsion force between the PMMA2 particle and pepsin predominates in the adsorption process. Retarded adsorption of pepsin molecules on the PMMA2 particles and, hence, stable colloidal systems in the presence of pepsin at pH > 7.1 are then expected. This is further supported by the low absorbance at 560 nm and high︱ζ︱of colloidal particles in the presence of pepsin at pH > 7.1 (Fig. 4).
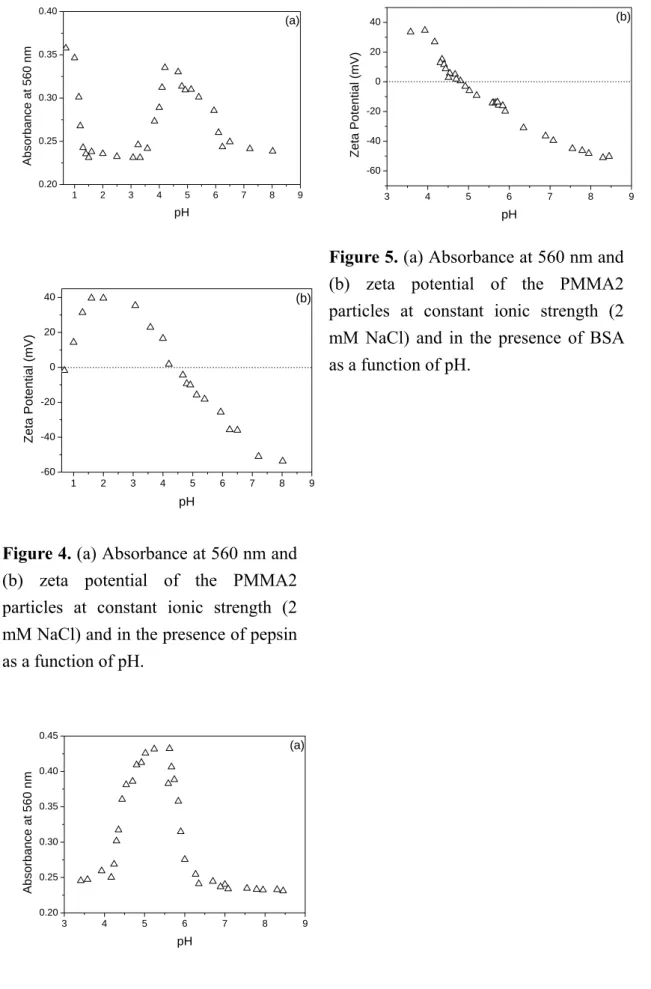
Interactions between PMMA2 and BSA.
The data of the absorbance at 560 nm and ζ of the PMMA2 particles in the presence of BSA are shown in Fig. 5. It is noteworthy that the difference between the IEP of the PMMA2 particle in the absence of protein and the pI of the model protein BSA (5) is the smallest (2.1) among the amphoteric latex-protein pairs investigated in this work. In this case, only one distinct peak is observed in the absorbance of the PMMA2 particles in the presence of BSA at 560 nm versus pH data and the maximum seems to correspond to the pI of BSA (Fig. 5a). It is apparent that the difference between the IEP of the amphoteric particle and the pI of the protein has a significant effect on the number of peaks that can be identified in the absorbance of colloidal particles in the presence of protein at 560 nm versus pH profile. Nevertheless, the point at
which the only maximal absorbance at 560 nm occurs seems to correspond to the pI of the protein sample.
At pH < 5, both the PMMA2 particles in the absence of protein and BSA molecules are positively charged and, therefore, the electrostatic repulsion force between the PMMA2 particle and BSA (i.e., the retarded BSA adsorption) is operative in the adsorption process.
However, the ζ of the PMMA2 particles in the absence of protein first remains relatively constant and then decreases with increasing pH. The surface charge density of BSA also decreases when pH increases from 3 to 5. As a result, the electrostatic repulsion force between the PMMA2 particle and BSA is greatly reduced and the amount of adsorbed BSA increases accordingly as the pH approaches 5. In this case, it is the hydrophobic interaction mechanism governs the BSA adsorption process.
Furthermore, the electrostatic repulsion force between two approaching particles is not strong to maintain adequate colloidal stability when the pH approaches 5, as shown by the ζ data in Fig. 5b. As expected, the absorbance of colloidal particles in the presence of BSA at 560 nm increases rapidly to a maximum (occurring at ca. pH 5.5) when the pH approaches 5 (Fig. 5a).
In the pH range 5-7.1, the PMMA2 particles in the absence of protein remain positively charged (Fig. 1), but BSA molecules become negatively charged. Thus, the charge neutralization
mechanism controls the BSA adsorption process and the rapidly increased amount of adsorbed BSA with pH enhances the particle surface charge density, as shown by the ζ data in Fig.
5b. The extent of charge neutralization diminishes when pH is close to 7.1. This is simply because the PMMA2 particles in the absence of protein carry a net charge of zero as the pH approaches their IEP value (7.1). This leads to the rapid decrease of the absorbance of colloidal particles in the presence of BSA at 560 nm when pH increases from 5 to 7.1 (Fig. 5a). Beyond pH 7.1, both the PMMA2 particles and BSA molecules carry net negative charges, thereby leading to the electrostatic repulsion force between the PMMA2 particle and BSA (i.e., the retarded BSA adsorption). It is then postulated that those two peaks in the absorbance of colloidal particles at 560 nm versus pH data, corresponding to the IEP of the PMMA2 particles in the absence of BSA and the pI of BSA, respectively, are overlapped with each other. This is most likely due to the very small difference between the IEP of the PMMA2 particles in the absence of protein and the pI of BSA. More research work is required to verify this speculation.
Conclusions
The feasibility of using single amphoteric latex of known IEP to interact with a protein sample at various pH values to determine the pI value of
the protein was investigated. The pairs of PMMA1 (5.2)-lysozyme (11), PMMA2 (7.1)-pepsin (1) and PMMA2-BSA (5) were chosen for study.
The numeric values in the parentheses represent the IEP values of the amphoteric latexes or the pI values of proteins. Both the pairs of PMMA1-lysozyme and PMMA2-pepsin with a large difference between the IEP of the amphoteric latex particles in the absence of protein and the pI of the model protein showed two distinct characteristic peaks in the absorbance of colloidal particles in the presence of protein at 560 nm versus pH data; one corresponded to the pI of the protein sample and the other presumably originated from the amphoteric nature of the latex particles. However, the position of the peak corresponding to the amphoteric latex may be quite different from the IEP of the latex in the absence of protein. On the other hand, only one peak was observed in the absorbance of colloidal particles in the presence of protein at 560 nm versus pH data for the PMMA2-BSA pair. This was attributed to the rather small difference between the IEP of the amphoteric latex particle in the absence of protein and the pI of the model protein. This characteristic peak was then assigned to the pI of the protein sample. The proposed method gave some promising results, but more experiments are required to verify the general validity of such an electrostatic interaction technique. As expected,
electrostatic interactions between the amphoteric particles and protein molecules and the charge reversal of colloidal particles in the presence of protein species played an important role in the protein adsorption process. The mechanisms involved in the electrostatic interactions between the amphoteric latex particles and protein molecules were proposed to explain the observed colloidal phenomena.
Reference
1. Norde, W.; Lyklema, J. J. Colloid
Interface Sci. 1978, 66, 277
2. Shirahama, H.; Takeda, K.;
Sukawa, T. J. Colloid Interface
Sci. 1986, 109, 552
3. Tamai, H.; Fujii, A.; Suzawa, T. J.
Colloid Interface Sci. 1987, 118,
1764. Kim, C. W.; Kim, S. K.; Rha, C.
In: Attia, Y. A. ed. Flocculation in
Biotechnology and Separation Systems; Elsevier: Amsterdam,
1987, p 4295. Kim, C. W.; Rha, C. Enzyme
Microb. Technol. 1987, 9, 57
6. Kim, C. W.; Rha, C. Biotechnol.Bioeng. 1989, 33, 1205
7. Sumi, Y.; Shiroya, T.; Fujimoto, K.; Wada, T.; Handa, H.;
Kawaguchi, H. Colloids Surfaces
B: Biointerfaces 1994, 2, 419
8. Ortega-Vinuesa, J. L.;Hidalgo-Alvarez, R. J. Biomater.
Sci., Polym. Ed. 1994, 6, 269
9. Ortega-Vinuesa, J. L.;Hidalgo-Alvarez, R. Biotechnol.
Bioeng. 1995, 47, 633
10. Chern, C. S.; Lee, C. K.; Chen, C.
Y.; Yeh, M. J. Colloids Surfaces B:
Biointerfaces 1996, 6, 37
11. Chern, C. S.; Lee, C. K.; Ho, C. C.
J. Polym. Sci., Part A: Polym.
Chem. 1999, 37, 1489
12. Chern, C. S.; Lee, C. K.; Ho, C. C.
Colloid Polym. Sci. 1999, 277,
97913. Homola, A.; James, R. O. J.
Colloid Interface Sci. 1977, 59,
12314. Shirahama, H.; Ohno, H.; Suzawa, T. Colloids Surfaces 1991, 60, 1 15. Fang, S. J.; Kawaguchi, H.
Colloids Surfaces A 2002, 211, 79
16. Chern, C. S.; Lee, C. K.; Chang,C. J. Colloid Polym. Sci. 2003,
281, 1092
17. Chern, C. S.; Lee, C. K.; Chang, C. J. Colloid Polym. Sci. 2004,
283, 257
18. Chern, C. S.; Lee, C. K.; Kuan, C.; Liu, K. C. Colloid Polym. Sci.
2005, 283, 917
Table 1. Some Physical Properties of Amphoteric Polymethyl Methacrylate Latexes
Latex ID PMMA1 PMMA2
[MAA] (%)
a
3 1[V-50] (%)
b
0.42 0.41d
p
(nm)c
285 310d
v
(nm)d
234 248d
w
/dn e
1.01 1.01IEP
f
5.2 7.1a
weight percentage of MAA based on total MMA in the polymerization recipeb
weight percentage of V-50 based on total water in the polymerization recipec
hydrodynamic particle diameter determined by dynamic light scatteringd
volume-average particle diameter of the latex product determined by TEMe
polydispersity index of particle size distribution of the latex product determined byTEM
f
isoelectric point of the latex product determined by a zeta potential meter2 3 4 5 6 7 8 9 10 11 -80
-60 -40 -20 0 20 40 60 80
[NaCl] = 2 mM PMMA1 PMMA2
Ze ta Pot ent ia l (mV)
pH
Figure 1. Effect of pH on the zeta
potential of amphoteric PMMA particles at constant ionic strength (2 mM NaCl) and in the absence of protein as a function of pH: (○) PMMA1, (△) PMMA2.2 4 6 8 10 12
200 400 600 800 1000
(a)
d
P(nm )
pH
[NaCl] = 2 mM PMMA1 PMMA2
3 4 5 6 7 8 9 10
0.25 0.26 0.27 0.28 0.29 0.30 0.31 0.32
(b)
Abso rb ance at 5 60 nm
pH
[NaCl] = 2 mM PMMA1 PMMA2
Figure 2. (a) Average hydrodynamic
particle diameter and (b) absorbance at 560 nm of the amphoteric PMMA particles at constant ionic strength (2 mM NaCl) and in the absence of protein as a function of pH: (○) PMMA1, (△) PMMA2.
2 3 4 5 6 7 8 9 10 11 12 13
0.20 0.25 0.30 0.35 0.40
0.45
(a)
A b sorb an ce at 56 0 nm
pH
3 4 5 6 7 8 9 10 11 12 13
-80 -60 -40 -20 0 20 40
60 (b)
Zeta Potential (mV)
pH
Figure 3. (a) Absorbance at 560 nm and
(b) zeta potential of the PMMA1 particles at constant ionic strength (2 mM NaCl) and in the presence of lysozyme as a function of pH.1 2 3 4 5 6 7 8 9 0.20
0.25 0.30 0.35 0.40
(a)
Absor b ance at 560 nm
pH
1 2 3 4 5 6 7 8 9
-60 -40 -20 0 20
40 (b)
Zeta Potential (mV)
pH
Figure 4. (a) Absorbance at 560 nm and
(b) zeta potential of the PMMA2 particles at constant ionic strength (2 mM NaCl) and in the presence of pepsin as a function of pH.3 4 5 6 7 8 9
0.20 0.25 0.30 0.35 0.40 0.45
(a)
Ab sor bance at 56 0 n m
pH
3 4 5 6 7 8 9
-60 -40 -20 0 20
40