國立臺灣大學生命科學院動物學研究所 碩士論文
Institute of Zoology College of Life Science
National Taiwan University Master Thesis
黃果蠅嗅覺受器 33c 的分子演化
Molecular evolution of odorant receptor 33c in Drosophila melanogaster
沈莞儒 Wan-Ju Shen
中華民國 98 年 7 月 July, 2009
指導教授: 丁照棣
方淑
Chau-Ti Ting Advisors:
Shu Fang
Acknowledgements
從2005 年夏天進入果蠅實驗室開始,四年過去了,謝謝這一路上身邊所有給
予我實質與精神上幫助的許多人,讓我從對果蠅一無所知、對演化一知半解,變 成了現在的我。
非常謝謝丁照棣老師與方淑老師這一路上的教導,無論是實驗的技巧或是思 考問題的方法,讓我明白做研究應該要有什麼樣的態度。謝謝您們包容我的許多 不足,願意耐心的提醒我、督促我,給我意見,幫助我由不同方面思考問題,甚 至不論多晚都還幫著我修改論文,真的非常謝謝您們。另外,謝謝張慧羽老師在 meeting 時總給予我實驗上的建議;謝謝王弘毅老師,從您的課堂中讓我學習到許
多論文相關的知識和方法;謝謝蔡玉真老師您對我poster 的建議,您的鼓勵讓我
不會害怕面對問題。謝謝您們擔任我的口試委員,在論文及實驗上給予許多的幫 助,讓我能更看清楚問題的方向。
謝謝曹順成老師從我還是菜鳥時,就常運用許多例子給予我實驗上的幫助;
謝謝小春,喜歡你溫暖的笑容和聲音,謝謝你在數據分析上给我的許多建議和協 助,並且告訴我該如何去樂在其中的解決問題;謝謝雅仁,有你在真好,無論什 麼大小事總有個人可以問可以談,謝謝你總像個大姊一樣照顧我這個少根筋的學 妹;謝謝其駿,願意花時間寫程式幫助我處理數據並和我討論,使我整理思緒;
謝謝小菊,總是說笑話讓大家開心,並且告訴我要想清楚自己要的是什麼;謝謝 文喬,無論多忙多累,總是耐心的解釋問題給我聽並幫我打氣;謝謝光曦,謝謝 你不時借給我許多精神食糧並給我鼓勵;謝謝承叡,因為你對研究的熱情,讓我
再度拾起找尋自己興趣的信心;謝謝Stéphane,總是雞同鴨講的和我談實驗聊生
活;謝謝姵君總是帶給我許多笑料,和我ㄧ起度過了這兩年;謝謝韋進和景賀,
常陪我討論實驗,讓我在實驗室的夜晚不會孤單;謝謝筱涵,謝謝你常告訴我不 用害怕要加油;謝謝啟發常给我許多鼓勵並幫助我很多事情;謝謝大馬常吐槽我 讓我腦筋不至於太停滯;謝謝士又、凱強、麒鳴、Kevin 和心彥,因為你們讓我的
生活更加豐富;謝謝王弘毅老師實驗室的雲露在處理RNA 技巧上的傳授;謝謝羅
竹芳老師實驗室的俊諺學長在real-time PCR 機器上的協助;謝謝楊恩誠老師實驗
室的學長姐們和錢伯在尚未完成的電生理實驗上的協助。
最後,謝謝一直陪在我身旁,我最愛的家人們,謝謝爸媽你們無論再累,還 是常常接我回家,放假還會陪我四處溜達;謝謝哥哥們常聽我吐口水,給我關懷,
並且在我晚回家的時候,和爸媽一起發動連環call。因為有你們,我才能一直走到
現在。
種種的感謝絕對無法用文字言語來傳達,但是一定要讓你們知道,因為有你 們,我才能在歡笑淚水中,完成這一本論文。謝謝你們。
摘要
果蠅的表皮碳氫化合物具有性費洛蒙的功能,在物種間的識別及生殖隔離方 面扮演很重要的角色。果蠅的嗅覺器官為觸角及小顎鬚,先前對多種常見氣味的
測試,都未發現小顎鬚上的嗅覺受器33c (Or33c)有任何反應,推測費洛蒙有可能
是此受器有反應的未知氣味。另外,在不同物種間,Or33c 已被證實受到正向選擇
的作用。為了更進一步了解果蠅Or33c 的功能以及正向選擇對於 Or33c 演化的影
響,我首先比較了Or33c 在不同族群及性別中的表現量,發現 Or33c 在黃果蠅中
的表現具有雌雄雙型性。另外,經由比較Or33c 和其縱向複製基因 Or33a 及 Or33b
(表現於觸角)的序列多型性,發現無論是在非洲族群或非非洲族群中,Or33c 皆受
到了正向選汰的作用,且Or33b 和 Or33c 間的基因區段亦受到此正向選汰的作用
而發生hitchhiking 現象。Or33c 具有一個使其能在小顎鬚表現的正向調控模段,此
模段的獲得可能與其轉換到不同感受器官上表現及所受之正向選汰有關。了解何
種因素驅動了Or33c 的正向選汰,將能更進一步釐清 Or33c 在偵測表皮碳氫化合
物上的可能功能。
關鍵字:表皮碳氫化合物,黃果蠅,嗅覺受器,正向選汰,雌雄雙型性
Abstract
Pheromonal cuticular hydrocarbons (CHs) have been demonstrated to play
important roles in species recognition and sexual isolation in Drosophila. Previous
study showed that odorant receptor 33c (Or33c) locating on one of the Drosophila
olfactory sense organs, maxillary palpi, may respond to a specific unidentified odorant
such as pheromone. In addition, positive selection on Or33c was detected between
species. To further investigate the potential function of Or33c in Drosophila
melanogaster, the expression level of Or33c between different populations and different
sexes were compared, and sexual dimorphism of this gene had been observed. Sequence
polymorphisms of Or33c and its tandemly duplicated genes, Or33a and Or33b
(expressed on the other olfactory sense organs, antennae), of D. melanogaster
populations were also compared to characterize how positive selection shapes the
evolution of Or33c in Drosophila. Analyses of the DNA sequences showed that in both
African and non-African populations there are significant signatures of positive
selection in Or33c. Or33c had acquired cis-regulatory motifs required for the gene
expression in the maxillary palp, the acquisition might involve the expression shift to
different sensory organs and/or positive selection. Understanding the driving force of
positive selection on Or33c will help us investigate the potential role of this gene in the
detection of pheromonal CHs.
Keywords: cuticular hydrocarbon, Drosophila melanogaster, olfactory receptor,
positive selection, sexual dimorphism
Contents
Acknowledgements………...i
Abstract……….………..…………..………ii
Introduction……….………...………...………1
Dispersal history of Drosophila melanogaster…………...………...………1
Reproductive isolation……….………...………...………1
Premating isolation……….………...………...……….………2
Cuticular hydrocarbons and peripheral olfactory system in Drosophila…...………4
Pheromonal communication and Or genes………...………...……..………6
Materials and Methods……...…………...………...…….………9
Fly stocks……….………9
Total RNA extraction………..………10
Quantitative RT-PCR………...……10
DNA extraction, PCR amplification, and sequencing………...…..…………11
Linkage disequilibrium………13
Sequence analyses……….…….………13
Results……….…16
Expression level of Or33c………...…16
Nucleotide polymorphism in African and non-African populations……...…18
Linkage disequilibrium………28
McDonald and Kreitman test………...…28
Predicted transmembrane helices………....31
Frequency spectrum……….…32
Tests of neutrality………36
Discussion………39
Expression pattern of Or33c………39
Demographic effect……….………39
Signature of selection………...…40
Mode of selection………..……….……….…41
Hitchhiking effect in Or33 gene family………...………..…42
References………...…43
Appendix……….………52
List of Figures
Fig. 1. Expression levels of Or33c in D. melanogaster populations………...…17 Fig. 2. Polymorphic sites of Or33 genes in the populations of D. melanogaster………19 Fig. 3. Sliding window plots of π and θw along Or33 genes sequence in African (A) and non-African (B) populations of D. melanogaster………27 Fig. 4. Linkage disequilibria of Or33 genes in the African (A) and the non-African (B) populations of D. melanogaster……….………..29 Fig. 5. Predicted topology of Or33c protein.………31 Fig. 6. Frequency spectra of Or33a gene in African (A), non-African (B) populations, and the combination of African and non-African populations (C) of D.
melanogaster………33
Fig. 7. Frequency spectra of Or33b gene in African (A), non-African (B) populations, and the combination of African and non-African populations (C) of D.
melanogaster………...……34
Fig. 8. Frequency spectra of Or33c gene in African (A), non-African (B) populations, and the combination of African and non-African populations (C) of D.
melanogaster………...……35 Fig. S1. Location of PCR primers………...…………52
List of Tables
Table 1 Primers and probes designed for quantitative RT-PCR……….…11
Table 2 Primers designed for PCR amplification………...………….…12
Table 3 Summary statistics of molecular variation for Or33 genes……….…...…26
Table 4 McDonald and Kreitman test for Or33 genes………30
Table 5 Site frequency spectrum statistics for Or33 genes...………37
Introduction
Dispersal history of Drosophila melanogaster
D. melanogaster has served as an important model system in molecular and
evolutionary genetics. Biogeographical and systematic studies suggest a colonization
history of Drosophila melanogaster. The cosmopolitan D. melanogaster originated
from tropical Africa and colonized Europe and Asia approximately 10,000 to 15,000
years ago, and followed by a relatively recent migration to the Americans and Australia
150 years ago (David and Capy 1988; Lachaise et al. 1988). Through this process of
population differentiation, many endemic variations have been generated, which could
be detected by different molecular markers, including DNA sequence data, mtDNA, and
microsatellites (Kreitman 1983; Kreitman and Aguadé 1986; Singh and Rhomberg 1987;
Hale and Singh 1991; Agis and Schlötterer 2001; Baudry, Viginier, and Veuille 2004;
Pool and Aquadro 2006; Nolte and Schlötterer 2008). Among these variations, some
may contribute to nascent reproductive isolation, thus could shed light on our
understanding towards speciation.
Reproductive isolation
Reproductive isolation could involve many complex mechanisms, including
geography, ecology, behavior, and interactions between multilocus genotypes. As a
consequence, it could arise various forms and is conventionally categorized based on
the stage in life cycle during which it has been observed: pre-mating, post-mating,
pre-zygotic, and post-zygotic (Coyne and Orr 2004). It is worth noting that in many
cases, especially allopatric ones, the reproductive isolation is just by-products of genetic
divergence because there are no forces acting to enforce reproductive compatibility,
instead of direct causes of speciation. Nonetheless, since the essential inquiry in
evolution is “why and how organisms change”, not “whether two groups can be
regarded as two species”, we could still extract hints from this kind of survey to answer
some interesting questions like: Which factor plays a more important role in divergent
process, stochastic drift or directional selection? And what kind of driving force may
involve in selection, environmental adaptation or sexual interaction? The major aims of
this study are trying to explore these questions in Drosophila melanogaster populations,
in which some premating reproductive isolation could be observed.
Premating isolation
Premating isolation could be achieved through difference in habitats, temporal
activity patterns (Marshall and Cooley 2000), modes of reproduction, and systems of
mate recognition. In the cases of mate recognition, populations come to differ in
signaling traits (of “chosen” individuals, which is usually male) and preferences (of
“choosy” individuals, which is usually female) for those traits. There are mainly two
non-exclusive theories proposed to explain how this occurs: 1) In Fisher’s runaway
model, genes for female preference become genetically correlated with genes for male
traits that are more attractive to females. Therefore, when genetic drift changes either
the trait or the preference, the runaway sexual selection could cause behavioral isolation
between different populations (Lande 1981). It is suggested this sort of mechanism may
explain the spectacular divergence in plumage and display among groups of closely
related species like the birds of paradise (Frith and Beehler 1998). 2) In sensory drive
hypothesis, both male mating traits and perceptual systems underlying female
preferences adapt to local environments (Endler 1992, 1993). Divergence occurs
because local environments impose selection of various intensity and direction on
mating signals aimed at potential mates and on sensory systems that acquire information
on predators, prey and mates (Ryan 1998). Previous works on lizards and cichlid fishes
have shown evidence supporting the hypothesis that adaptation to local light
environments drives specializations of the visual and signaling systems, bring on habitat
partitioning and speciation (Leal and Fleishman 2002; Seehausen et al. 2008).
In fruit fly Drosophila, the traits involved in mechanisms of isolation also
comprise a variety of different types, such as pheromones, courtship song, shortened
copulation time, genital morphology, and male/female sexual isolation (Coyne 1996a, b,
c; Coyne and Charlesworth 1997; Hoikkala et al. 2000; Zeng et al. 2000). The
populations surveyed in this study contain African populations of D. melanogaster in
and near Zimbabwe and non-African populations from other continents. The matings
between Zimbabwe females and cosmopolitan males are typically very rare (Wu et al.
1995; Hollocher et al. 1997). Taking its asymmetry and the existence of hybrid zones in
Africa into consideration, this incomplete premating isolation may indicate an incipient
form of speciation (Wu et al. 1995; Ting, Takahashi, and Wu 2001). Analyzing the
incipient speciation may benefit from being interfered by fewer secondary effects
occurring after speciation, providing clues to clarify what kind of gene may involve in
the primary phase of reproductive isolation.
Cuticular hydrocarbons and peripheral olfactory system in Drosophila
In D. melanogaster, males can distinguish the gender of other flies even when
most auditory and visual signals are removed. However, this ability is lost when
encountered with desat1 mutant line, whose males and females have very similar
cuticular hydrocarbon (CH) profiles (Marcillac, Grosjean, and Ferveur 2005). It is
revealed that sexual dimorphic CHs serve as pheromones for mate discrimination, and
may involve in sexual isolation just as in D. simulans/D. sechellia case (Coyne,
Crittenden, and Mah 1994).Previous studies showed that D. melanogaster populations
from Africa and other continents are different in cuticular hydrocarbons; and a mutation
in the promoter of desat2, a duplicate of desat1 coding for another desaturase, is
correlated with the assortative mating pattern described above (Takahashi et al. 2001).
In a pheromone communication system, tuning of receiver could be correlated with
changes in sender’s signal, as suggested in Ostrinia moth (Baker 2002). Thus, it would
be intriguing to survey whether there are some corresponding modifications in receiver
terminal, the olfactory receptors.
Many insects have two bilaterally symmetrical pairs of olfactory organs, the
antennae and the maxillary palps. The surfaces of these olfactory appendages are
covered with sensory hairs, called sensilla, containing olfactory receptor neurons
(ORNs) dendrites that are specialized to detect odors. In D. melanogaster adults, each
antenna contains about 1200 ORNs, and each maxillary palp contains about 120 ORNs
(Stocker 1994; Shanbhag, Müller, and Steinbrecht 1999; Shanbhag, Müller, and
Steinbrecht 2000). Each sensillum contains dendrites of one to four ORNs. The
olfactory sensilla of the antenna can be subdivided into three major morphological and
functional classes: club-shaped basiconic sensilla, long and pointed trichoid sensilla,
and short, peg-shaped coeloconic sensilla, whereas the olfactory sensilla of the
maxillary palp consist entirely of basiconic sensilla.
Search of D. melanogaster genome using bioinformatics methods predicted that
there are 62 OR proteins encoded by families of 60 Or genes through alternative
splicing (Robertson, Warr, and Carlson 2003). Drosophila OR proteins are highly
diverse and have no similarity to mammalian odorant receptors. However, closely
linked genes often share a higher degree of sequence similarity, suggesting that some
Or genes are likely to have undergone recent duplication events to generate clusters of Or genes.
There is a segregation of gene expression pattern between the two major olfactory
organs: ORs expressed in the antenna are not expressed in the maxillary palp and vice
versa. In situ hybridization showed that there are 32 Or genes expressed on the antenna
and seven Or genes expressed on the maxillary palp (Vosshall, Wong, and Axel 2000).
Pheromonal communication and Or genes
In the past several years, the pheromone detection and processing in Drosophila
male mating behaviors have been the focus. The best understood male pheromone is
cis-vaccenyl acetate (cVA), a volatile compound modulating both male and female
behaviors (Bartelt, Schaner, and Jackson 1985; Xu et al. 2005; Ejima et al. 2007;
Kurtovic, Widmer, and Dickson 2007). One olfactory receptor expressed on antenna,
Or67d, is required for cVA detection (Kurtovic, Widmer, and Dickson 2007). LUSH, an odorant binding protein, will bind cVA to activate Or67d (Xu et al. 2005). Then the
activated Or67d ORNs will target to a glomerulus called DA1 in the antennal lobe and
pass the cVA signal to the corresponding DA1 projection neurons (PNs) (Couto,
Alenius, and Dickson 2005; Schlief and Wilson 2007; Datta et al. 2008). Although both
Or67d ORNs and DA1 PNs respond to cVA equally in males and females, the DA1 PNs
form sex-specific arborization in the protocerebrum, suggesting that the cVA signal
may process differently in males and females (Datta et al. 2008). In addition to Or67d,
Gustatory receptor 32a (Gr32a) and Gr68a on the legs, Or47b, Or65a and Or88a on the antenna have also been proved d to have function of pheromone detection.
Of 60 Or genes, Odorant receptor 33c (Or33c), Or42a, and Or85e were suggested
under positive selection across eight ecologically diverse species of Drosophila
including D. melanogaster, D. mauritiana, D. simulans, D. sechellia, D. yakuba, D.
erecta, D. ananassae, and D. pseudoobscura by using maximum likelihood estimates of
dN/dS ratios (Tunstall et al. 2007). Among these three genes, Or33c and Or85e are
coexpressed in the same ORN class on the maxillary palp. The electrophysiological
experiments showed that responses of Or33c receptor to 150 odorants are much weaker
than Or85e receptor, suggesting that Or33c may respond to a specific unidentified
odorant such as pheromone (Goldman et al. 2005). To further characterize how positive
selection shapes the evolution of Or33c in Drosophila and whether the positive
selection of Or33c is related to the potential role on the detection of CH pheromones,
sequence polymorphisms of Or33c and its tandemly duplicated loci, Or33a and Or33b,
of D. melanogaster populations were compared.
Materials and Methods
Fly Stocks
Twenty-six African strains and 14 non-African strains of Drosophila melanogaster
were used in the sequence analyses. African strains were collected from Zimbabwe
(ZS2, ZS6, ZS8, ZS11, ZS29, ZS30, ZS49, ZS53, and ZS56 in Shegwa Wildlife
Reserve; ZH12, ZH18, ZH21, ZH23, ZH32, and ZH34 in Harare), Northern Zambia
(LA2, LA20, and LA66), Botswana (OK17 and OK91), Ivory Coast (Tai), and Malawi
(MW6, MW11, MW28, MW56, and MW63). The cosmopolitan lines were from several
different continents, including Europe (FR in France), North America (Canton-S in
Ohio; HG in California; Id in Indiana; Oahu in Hawaii; OR in Oregon; mm1 in North
Carolina; M97 in Miami), South America (Ica in Peru), Asia (NFS6 in Israel; QD12 and
QD18 in Japan; TWN in Taiwan), and Australia (Yv). ZS6, ZS8, ZS11, ZS30, ZH18,
ZH23, ZH32, LA20, LA66, FR, CS, HG, Id, and Ica lines were isogenic for the second
and third chromosomes. For the rest of lines which are isofemale lines, one male from
each lines was crossed with females carrying the deficiency Df(2L)Prl, Prl1 nubPrl/CyO,
which covers cytological positions 32F1-3 to 33F1-2; hemizygotes of Or33 genes could
be selected from the progeny. All flies were kept in vials containing the standard
yeast-cornmeal-agar medium and maintained at 25°C with humidity control under a
12h-12h light-dark cycle.
Total RNA extraction
Adult flies (three to five days old) were put into an empty vial for at least two
hours to void yeast before head dissection. Adults were dissected in phosphate buffered
saline to separate heads from bodies. Total RNA was extracted from heads with Trizol®
reagent according to the manufacturer’s instructions (Invitrogen). RNA pellets were
resuspended in diethyl pyrocarbonate (DEPC)-treated water. To prevent genomic DNA
contamination, the samples were treated with Deoxyribonuclease I (Invitrogen) and
stored at -80°C.
Quantitative RT-PCR
A 1-μg aliquot was used as template for cDNA synthesis, employing the
SuperScriptTM III First-Strand Synthesis system (Invitrogen) and oligo(dT) primers. The
cDNA was stored at -80°C. Quantitative RT-PCR assays were performed by the
Universal Probe Library (UPL) system (Mouritzen et al. 2005). Primers and probes for
determination of Or33c and endogenous control rp49 expression levels were designed
using the Roche Applied Science Universal Probe Library with online tools
(http://www.roche-applied-science.com/sis/rtpcr/upl/index.jsp) (table 1). Each 25 μl
reaction contained 12.5 μl of 2X FastStart Universal Probe Master (ROX), 100 nM
Universal Probe, 200 nM of each primer, and 50 ng cDNA. Reactions were subjected to
the following cycles on the Applied Biosystems 7300 Real-Time PCR System (Applied
Biosystems): step1- 95°C for 10 minutes; step 2- 95°C for 10 sec, 60°C for 1 minutes,
repeat step 2 for 39 additional cycles. Data analysis was based on the relative expression
level of Or33c to the reference gene, rp49, with a normalized calibrator method
described in manufacture’s instructions (Roche Applied Science). All reactions were
performed in triplicate, and repeated four times.
Table 1
Primers and probes designed for quantitative RT-PCR
Gene Forward (5’-3’) Reverse (5’-3’) Probe
Or33c CATCAGCGTTTGCTGGATTA TCACCAGGTTGTGGAATCG # 132 rp49 CTTCATCCGCCACCAGTC CGACGCACTCTGTTGTCG # 117
DNA extraction, PCR amplification, and sequencing
DNA was extracted from single fly following modified Puregene DNA
Purification Kit protocol (Gentra). Gene-specific primers (table 2 and appendix) were
used to perform PCR amplification. DNA sequences of PCR-amplified fragments were
sequenced on ABI PRISM® 96-capillary 3700 xl DNA Analyzer.
12
Table 2
Primers designed for PCR amplification
Forward Primer Sequence (5’-3’) Reverse primer Sequence (5’-3’) Fragment size (bp)
Or33b_ -1193F CAAGCAACCGCCGCAATCTA Or33b_-94R CCACATAGTTACGCCCATCA 1119
Or33b -563F AAAGCAGGTGGTATTGTTGGCATCG Or33b_-32R ACAATTGAAAAGGTCCTGCGGCATT 556
Or33b -877F TAATACGCCTACTTCGCAGCACTT Or33b_589R ACCAGCAACCACGCAAAATGTCAT 1489
Or33b_437F TTTTCGGCGGTGGACATAAGC Or33b_1284R CCAAAGTGAAGAAGGAGTAGG 868
Or33c_-1021F TAAATGAAGGTGTTGTAGGA Or33c_15R TGGACGATAAAAACTAAGACTG 1057
Or33c_-76F ATTTCTCGCAGTGTTTCCTCTC Or33c_1365R AACTGTTTATTTGCGTATC 1459
Linkage disequilibrium
To determine whether the association between polymorphic sites are significant or
not, the linkage disequilibrium (LD) graphs measured by r2, which is equal to the square
of the correlation coefficient between the alleles at two loci (Hill and Robertson 1968),
were constructed using data calculated form DnaSP version 5 (Librado and Rozas
2009).
Sequence analyses
Multiple sequence alignments were performed by SeqMan, EditSeq, and MegAlign
software with ClustalW method (Lasergene, DNAStar, Madison, WI). Several neutrality
tests were carried out by DnaSP version 5 (Librado and Rozas 2009). Sites containing
alignment gaps were not used in all the tests.
In the analysis of divergence between species, only coding regions were used for
the McDonald and Kreitman test (McDonald and Kreitman 1991). This test is based on
a comparison of synonymous and non-synonymous variation within and between
species. D. simulans was used as outgroup (sequence is available from Flybase). Under
the neutral theory, the ratio of replacement to synonymous fixed substitutions (R/S)
should be the same as the ratio of R/S within species.
Transmembrane (TM) helix prediction was carried out with the program
HMMTOP 2.0 (Tusnády and Simon 2001) and visualized by TMRPres2D
(Transmembrane Protein Re-presentation in 2 Dimensions tool), which automates the
creation of graphical images and models of TM proteins (Spyropoulos et al. 2004).
The following three tests are derived from θ estimated from different parts of
frequency spectrum which response to the selection differently. Tajima’s D test (Tajima
1989) is based on the normalized difference between the number of segregating sites (θ)
and the average number of nucleotide differences (π). At equilibrium between genetic
drift and selection, the expected value of D is close to zero. D is negative when there is
a relative excess of rare alleles in a folded frequency spectrum.
Fu and Li’s D* and F* test (Fu and Li 1993) are according to the expected total
length of the external and internal branches. Fu and Li’s D* test is based on the
differences between the number of singletons and the total number of mutations. Fu and
Li’s F* test is based on the differences between the number of singletons and k, the
average number of nucleotide differences between pairs of sequences. D* and F* are
negative when there is a relative excess of mid-frequency alleles in a folded frequency
spectrum.
Fay and Wu’s H test (Fay and Wu 2000) is used to measure an excess of high
compared intermediate frequency variants. This test is based on the differences between
k and θH, an estimator based on the frequency of the derived variants. Negative H value
means an excess of high-frequency-derived alleles relative to the standard neutral
model.
Results
Expression level of Or33c
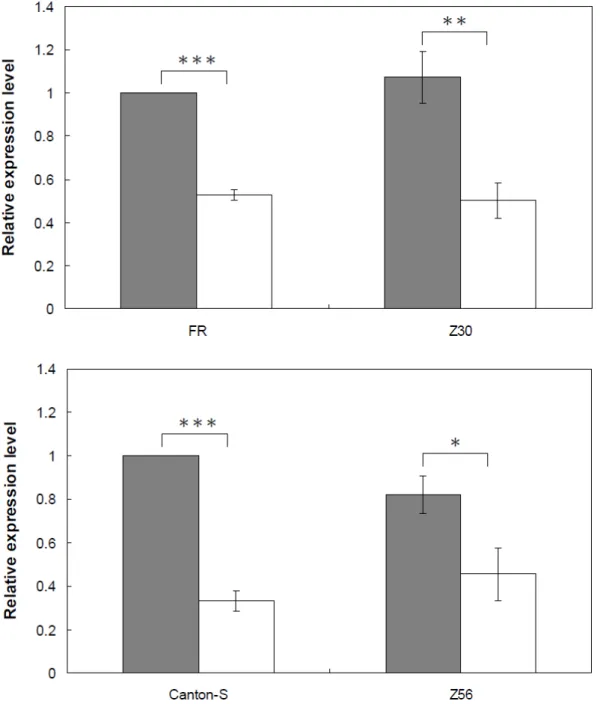
To examine whether the expression levels of Or33c are different between the
African and non-African populations, four lines from the non-African (FR and
Canton-S) and the African (Z30 and Z56) populations were divided into two groups (FR
vs. Z30; Canton-S vs. Z56) to compared by quantitative RT-PCR. The result showed
that the expression levels between these two populations have no significant difference
(two-tailed Student’s t-test, P > 0.1). Interestingly, the expression levels in males were
significantly higher than in females (fig. 1).
Fig. 1.—Expression levels of Or33c in D. melanogaster populations, as quantified by
real-time RT-PCR. All values are normalized against an internal control (rp49). Solid
bars, males; open bars, females. *P < 0.05, **P < 0.01, and *** P <0.001, two-tailed
Student’s t-test.
Nucleotide polymorphism in African and non-African populations
The sequenced region spanning Or33a, Or33b, and Or33c genes consists of 4185
bp in length. Fig. 2 summarizes the distribution of nucleotide sequence polymorphism
compared with D. simulans as outgroup, including a total of 399 polymorphic sites in
14 non-African strains and 26 African strains. Summary statistics of molecular variation
are shown in table 3. At different regions, k ranges from 5.874 to 17.145, θw and π range
from 0.01 to 0.024 in the African populations; k ranges from 4.462 to 15.725, θw and π
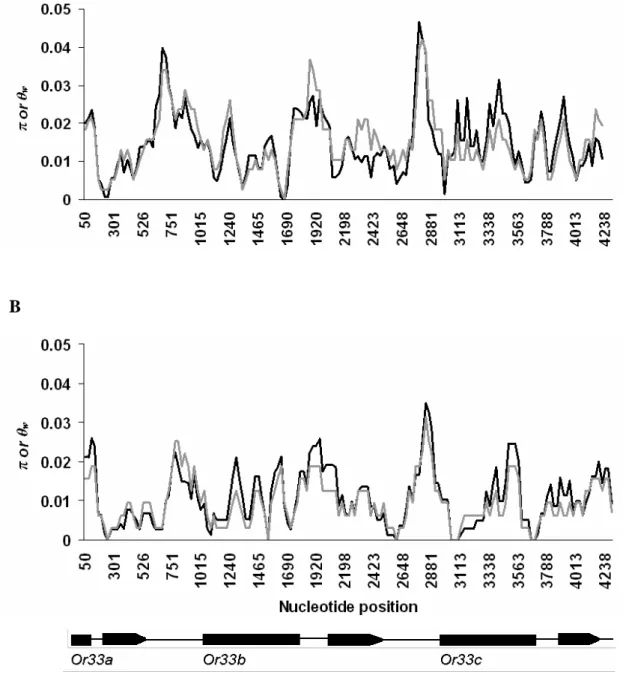
range from 0.007 to 0.014 in the non-African populations. Nucleotide diversity of Or33
genes was shown in fig. 3. The result showed that the nucleotide polymorphism in the
African population is higher than in the non-African population.
19
Fig. 2.—Polymorphic sites of Or33 genes in the populations of D. melanogaster. Polymorphisms are numbered from the initiation codon of each gene. Dots represent the same nucleotide as in the D. simulans sequence. Dashes represent alignment gaps in nucleotide sequence. R and S stand for replacements and synonymous, respectively.
20
Fig. 2.—(Continued)
21
Fig. 2.—(Continued)
22
Fig. 2.—(Continued)
23
Fig. 2.—(Continued)
24
Fig. 2.—(Continued)
25
Fig. 2.—(Continued)
Table 3
Summary statistics of molecular variation for Or33 genes
Gene Length S k θw π
African (N = 26)
Or33a 661 23 5.874 0.010 0.010
Intergenic region 494 41 10.677 0.024 0.024
Or33b 1371 73 16.551 0.013 0.015
Intergenic region 482 36 8.877 0.019 0.020
Or33c 1177 54 17.145 0.015 0.012
Non-African (N = 14)
Or33a 661 14 4.462 0.007 0.007
Intergenic region 494 20 5.396 0.012 0.014
Or33b 1371 40 15.725 0.012 0.010
Intergenic region 482 19 6.473 0.014 0.013
Or33c 1177 29 10.967 0.009 0.008
African and non-African (N = 40)
Or33a 661 28 5.576 0.009 0.011
Intergenic region 494 46 9.031 0.021 0.025
Or33b 1371 78 16.642 0.013 0.014
Intergenic region 482 39 8.259 0.018 0.020
Or33c 1177 56 15.844 0.014 0.011
Note.—S, number of segregating sites; k, the average number of nucleotide differences;
θw, Watterson’s (1975) estimates of the mutation parameter; π, nucleotide diversity.
A
B
Fig. 3.—Sliding window plots of π (black lines), and θw (gray lines) along Or33 genes sequence in African (A) and non-African (B) populations of D. melanogaster. A window length of 100 bp moved in the steps of 25 bp along the sequence was used.
Gene structure of Or33 genes was indicated by black boxes (exons) and thin lines (introns) at the bottom of the graph.
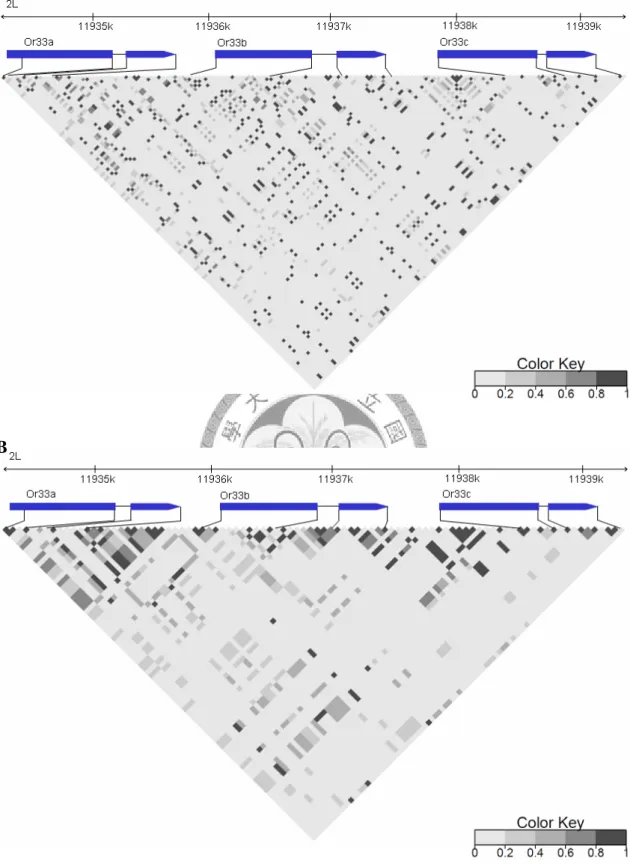
Linkage disequilibrium
The derived non-African population showed a higher level of linkage disequilibrium than the ancestral African population (fig. 4).
McDonald and Kreitman test
The DNA sequence variations of Or33a, Or33b, and Or33c coding regions were analyzed separately by the McDonald and Kreitman test (table 4). The evolutionary patterns of Or33a, Or33b, and Or33c revealed by the replacement to silent sites ratio (R/S) are different. In Or33a and Or33b, the R/S ratios between species
(D.melanogaster and D. simulans) and within species (D. melanogaster) are not significantly different. Whereas in Or33c, the R/S between species is significantly higher than that within species. These patterns are similar in both the African population and the non-African population, suggesting that positive selection operating on Or33c when D. melanogaster diverged from D. simulans.
A
B
Fig. 4.—Linkage disequilibria of Or33 genes in the African (A) and the non-African (B) populations of D. melanogaster. The degree of LD estimated by r2 is shown by different shades.
Table 4
McDonald and Kreitman test for Or33 genes
Fixed differences Polymorphic differences (between species) (within species)
R S R S G Value (P value)
African (N = 26)
Or33a 7 9 11 14 0.000 (0.987)
Or33b 15 26 19 37 0.073 (0.787)
Or33c 20 22 11 39 6.755 (0.009)
Non-African (N = 14)
Or33a 8 12 7 6 0.609 (0.435)
Or33b 15 29 12 19 0.168 (0.682)
Or33c 20 26 5 23 5.398 (0.020)
African and non-African (N = 40)
Or33a 7 9 14 15 0.085 (0.771)
Or33b 15 26 23 38 0.013 (0.909)
Or33c 20 22 12 41 6.565 (0.010)
Note.—R and S stand for the number of amino acid replacements and silent changes, respectively. Significant P values (less than 5%) are in bold.
Predicted transmembrane helices
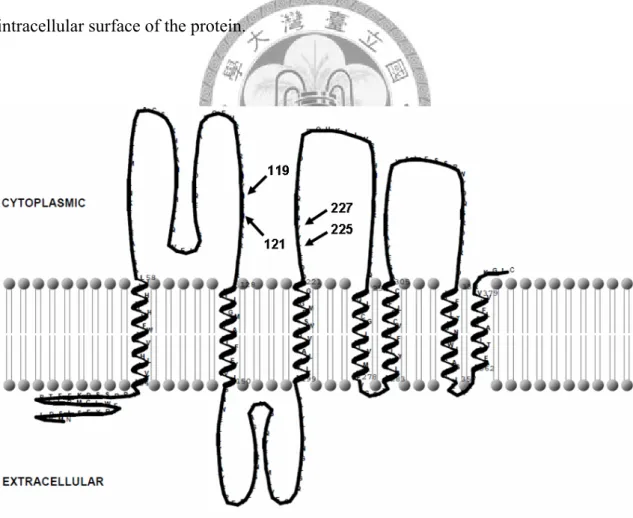
Among the fixed nucleotide changes between D. melanogaster and D. simulans, there are four sites involve in polarity changes (fig. 5). All of these four sites are located in the loop regions of the Or33c protein. The sites on the extracellular surface may involve in ligand binding, whereas intracellular sites may involve in signal transduction.
The HMMTOP 2.0 prediction result showed that Or33c protein has the regular G protein-coupled receptor (GPCR) topology with the extracellular N-terminal. This topology is different to the novel Drosophila chemoreceptor topology proposed by Benton et al. (2006), which the N-terminal is in the intracellular surface. According to the regular GPCR topology, the four sites changing amino acid polarity are on the intracellular surface of the protein.
Fig. 5.—Predicted topology of Or33c protein. The arrows indicate the sites involving polarity changes.
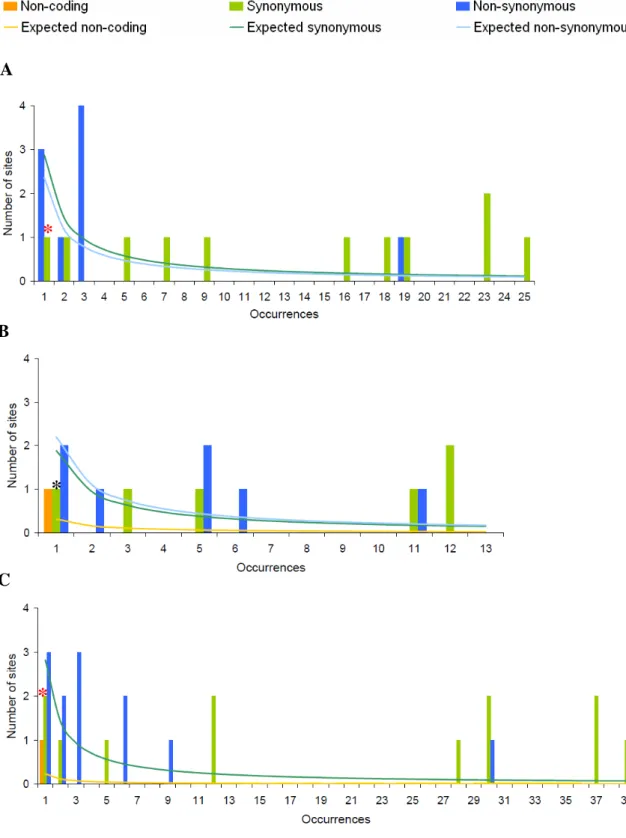
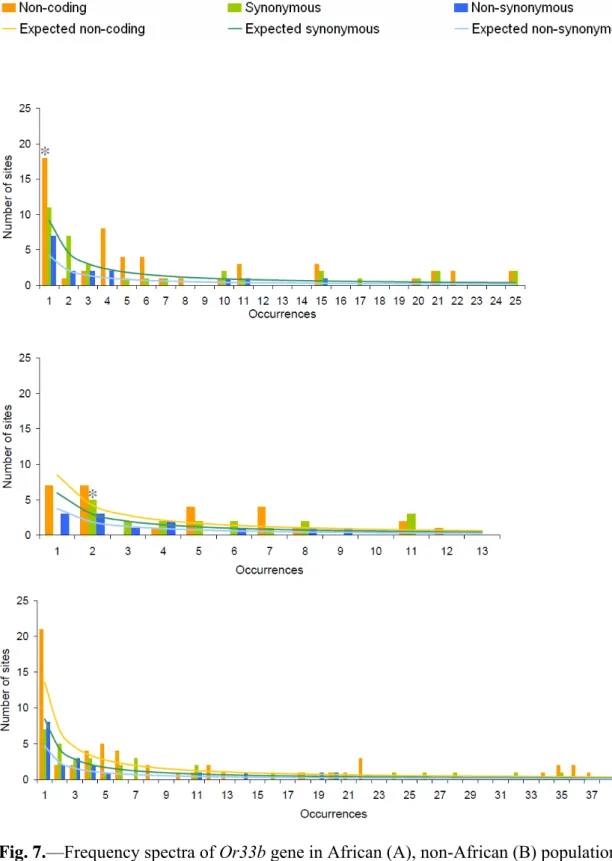
Frequency spectrum
The goodness-of-fit test revealed that the frequency spectra on the non-coding region of Or33b, and Or33c in the African population significantly departures from neutrality (P = 0.013 and 0.033, respectively). In the non-African population and the combination of African and non-African populations, Or33c significantly deviate from the neutrality (P = 0 and 0.01, respectively). The synonymous sites frequency spectra of Or33a and Or33c are significantly higher than the neutral expectation in the African population (P = 0 and 0.009, respectively) and the combination of African and
non-African populations (both P = 0). In the non-African population, Or33a and Or33b significantly departures from neutrality (P = 0.003 and 0.017, respectively) (figs. 6-8). To further analyze the frequency spectra, neutrality tests based on frequency spectrum were performed.
A
Fig. 6.—Frequency spectra of Or33a gene in African (A), non-African (B) populations, and the combination of African and non-African populations (C) of D. melanogaster.
Black star, 0.001 < P < 0.01; red star, P < 0.001.
*
*
* B
C
Fig. 7.—Frequency spectra of Or33b gene in African (A), non-African (B) populations, and the combination of African and non-African populations (C) of D. melanogaster.
Gray star, 0.01 < P < 0.05.
*
*
Fig. 8. —Frequency spectra of Or33c gene in African (A), non-African (B) populations, and the combination of African and non-African populations (C) of D. melanogaster.
Gray star, 0.01 < P < 0.05; black star, 0.001 < P < 0.01; red star, P < 0.001.
*
*
*
*
*
Tests of neutrality
Tests of neutrality based on polymorphism data were applied to the Or33a, the
intergenic region of Or33a and Or33b, Or33b, intergenic region of Or33b and Or33c,
and Or33c. Tajima’s D test, Fu and Li’s D* and F* tests of Or33a, Or33b and
intergenic region between Or33b and Or33c showed higher values in the non-African
population than in the African population. Neither the Tajima’s D test nor the Fu and
Li’ D* and F* tests detected any departure from neutral expectations in any populations.
Fay and Wu’s H test showed a significant degree of deviation from the neutral
expectation in Or33a (African: H = -1.466; non-African: H = -1.022) and in the
intergenic region between Or33b and Or33c (African: H = -1.164; H = -1.386) in D.
melanogaster (table 5).
37
Table 5
Site frequency spectrum statistics for Or33 genes
Gene Length Tajima’s D Fu and Li’s D* Fu and Li’s F* Fay and Wu’s H African (N = 26)
Or33a 661 -0.378 (0.286) -0.046 (0.462) -0.176 (0.390) -1.466 (0.021) Intergenic region 494 -0.113 (0.450) -0.085 (0.417) -0.111 (0.425) -0.269 (0.249) Or33b 1371 -0.613 (0.132) -0.948 (0.069) -0.989 (0.071) -0.298 (0.205) Intergenic region 482 -0.318 (0.315) -0.504 (0.227) -0.523 (0.245) -1.164 (0.030) Or33c 1177 0.646 (0.956) 0.031 (0.510) 0.268 (0.698) -0.075 (0.385) Non-African (N = 14)
Or33a 661 0.054 (0.541) 0.233 (0.601) 0.212 (0.589) -1.022 (0.095) Intergenic region 494 -0.594 (0.270) -0.052 (0.454) -0.228 (0.396) 0.096 (0.420)
Or33b 1371 1.088 (0.988) 1.274 (0.999) 1.405 (0.998) 0.121 (0.532) Intergenic region 482 0.347 (0.693) 0.104 (0.529) 0.195 (0.564) -1.386 (0.047)
Or33c 1177 0.697 (0.946) 0.500 (0.848) 0.636 (0.890) -0.058 (0.412)
38
Table 5 (Continued)
Gene Length Tajima’s D Fu and Li’s D Fu and Li’s F Fay and Wu’s H African and non-African (N = 40)
Or33a 661 -0.721 (0.133) -0.436 (0.268) -0.633 (0.204) -1.582 (0.012) Intergenic region 494 -0.649 (0.203) -0.614 (0.218) -0.744 (0.185) -0.629 (0.117) Or33b 1371 -0.419 (0.203) -0.796 (0.106) -0.789 (0.104) -0.229 (0.256) Intergenic region 482 -0.504 (0.237) -0.777 (0.150) -0.810 (0.152) -1.020 (0.036) Or33c 1177 0.511 (0.890) 0.031 (0.499) 0.237 (0.662) -0.224 (0.273) Note. —P values determined from 5000 coalescent simulations with recombination are shown in parenthesis. Significant P values (less than 5%) are shown in bold.
Discussion
Expression pattern of Or33c
The sexually dimorphic expression pattern in Or33c might be in connection with
the hypothesis that may be involved in the detection of sex-specific ligand, such as
pheromone (Goldman et al. 2005). And it can be seen as well that this pattern is shown
in both the African and the non-African populations. On the other hand, in both sexes
there is no significant difference in the expression level between populations, indicating
that even function of Or33c may diverge in different populations due to different
environmental constraints, it may not necessarily be accompanied with changes in
sensitivity (which is correlated with expression level). Nevertheless, further functional
analyses are needed to answer whether the driving force of positive selection of Or33c
is related to the potential role on the detection of CH pheromones.
Demographic effect
As mentioned in the introduction, D. melanogaster populations have undergone
complicated dispersal history, and this factor should leave footprints on the genome
sequences. The lower nucleotide diversity and the higher level of LD in non-African
population are consistent with the dispersal history leading to demographic effect of the
non-African population. The non-African population is composed of samples from
worldwide localities. As a consequence, some linkage disequilibrium actually reflects
geographical divergence, instead of true functional/physical correlations. Furthermore,
samples from the populations which underwent bottleneck phase (e.g. American
populations) would incorporate more fixation by genetic drift, thus cause more bias on
LD data.
Signature of selection
It has been shown that Or33c may be acted on by positive selection in the lineage
leading to ecologically diverse species of Drosophila (Tunstall et al. 2007). My study
further characterizes the signature of positive selection of Or33c in D. melanogaster
populations by McDonald and Kreitman test. In contrast to Or33c, neither of its tandem
duplicate genes, Or33a and Or33b (Robertson, Warr, and Carlson 2003), shows a
significant signature of positive selection. These patterns can be observed in both
African and non-African populations. Both Or33a and Or33b are expressed in antenna,
whereas Or33c is expressed in the maxillary palp (Vosshall, Wong, and Axel 2000).
The acquisition of the Dyad-1 motif, which is required for the gene expression in the
maxillary palp (Ray et al. 2007), in the 5’ upstream region of Or33c might explain the
expression pattern different from its duplicate genes. Furthermore, it has been shown
that the upstream regulatory regions of Or33c and its co-expresser Or85e contain two
shared cis-regulatory motifs that are not present in the regulatory regions of any other
Or genes (Ray et al. 2007). The acquired cis-regulatory motifs might involve the
expression shift to different sensory organs and/or positive selection. This pattern is
similar to Or59b and Or59c, which are also closely related tandemly duplicates
(Robertson, Warr, and Carlson 2003). Although Or59b and Or59c have very similar
sequence to each other, they have no shared regulatory motifs and their expression are
different (Or59b expressed on antenna and Or59c expressed on maxillary palp)
(Vosshall, Wong, and Axel 2000; Ray et al. 2007).
Mode of selection
A trend could be noticed that, values of Tajima’s D, Fu and Li’s D* and F* tests
of Or33a, Or33b and intergenic region between Or33b and Or33c are larger, though not
significant, in the non-African population than in the African one. This pattern may
reflect the fact that the non-African population has experienced a population subdivision
phase (Simonsen, Churchill, and Aquadro 1995), which is consistent with previous
studies (Schlötterer, Vogl, and Tautz 1997).
Another question is, if there exists positive selection acting on Or33c as suggested
by McDonald and Kreitman test, why don’t we find the similar signature in other
neutrality tests such as Tajima’s D, Fu and Li’s D* and F* tests? The difference
between these two kinds of tests is due to that McDonald and Kreitman test is based on
the comparison of R/S ratios between divergence in two species and polymorphism
within species, while other neutrality tests I applied in this study are based on the
frequency spectra of polymorphic sites. If selection took action only during a period in
the past, it could lead to an excess of fixation but no significant deviation from
neutrality in polymorphism data. This mode of selection suggests that an episodic
positive selection has been occurred between D. melanogaster and D. simulans
lineages.
Hitchhiking effect in Or33 gene family
The Fay and Wu’s test could be used to infer whether the neutral locus in question
is hitchhiked by a linked locus that is under positive selection (Fay and Wu 2000). It can
be seen that in both the African and the non-African populations, intergenic region
upstream to Or33c show a significant derivation from neutrality by Fay and Wu’s test,
suggesting that 5’ upstream region of Or33c may be hitchhiked by Or33c. By further
identified putative transcription factor binding site in the 5’ upstream region of Or33c
may help us to find evidence for this hypothesis.
On the other hand, although Or33a showed no sign of deviation from neutrality by
the McDonald and Kreitman test, a significant deviation from neutrality had been
detected in the synonymous sites frequency spectra and in the Fay and Wu’s test. This
result suggests that this region of Or33a may be hitchhiked by the nearly narrow region.
References
Agis M, Schlötterer C. 2001. Microsatellite variation in natural Drosophila
melanogaster populations from New South Wales (Australia) and Tasmania.
Mol Ecol. 10:1197-1205.
Baker TC. 2002. Mechanism for saltational shifts in pheromone communication
systems. Proc Natl Acad Sci USA. 99:13368-13370.
Bartelt RJ, Schaner AM, Jackson LL. 1985. cis-Vaccenyl acetate as an aggregation
pheromone in Drosophila melanogaster. J Chem Ecol. 11:1747-1756.
Baudry E, Viginier B, Veuille M. 2004. Non-African populations of Drosophila
melanogaster have a unique origin. Mol Biol Evol. 21:1482-1491.
Benton R, Sachse S., Vosshall LB. 2006. Atypical membrane topology and heteromeric
function of Drosophila odorant receptors in vivo. PLoS Biol. 4:1-18.
Couto A, Alenius M, Dickson BJ. 2005. Molecular, anatomical, and functional
organization of the Drosophila olfactory system. Curr Biol. 15:1535-1547.
Coyne J, Orr A. 2004. Speciation . Sunderland, MA: Sinauer Associates.
Coyne JA. 1996a. Genetics of differences in pheromonal hydrocarbons between
Drosophila melanogaster and D. simulans. Genetics. 143:353-364.
Coyne JA. 1996b. Genetics of a difference in male cuticular hydrocarbons between two
sibling species, Drosophila simulans and D. sechellia. Genetics. 143:1689-1698.
Coyne JA. 1996c. Genetics of sexual isolation in male hybrids of Drosophila simulans
and D. mauritiana. Genet Res. 68:211-220.
Coyne JA, Charlesworth B. 1997. Genetics of a pheromonal difference affecting sexual
isolation between Drosophila mauritiana and D. sechellia. Genetics.
145:1015-1030.
Coyne JA, Crittenden AP, Mah K. 1994. Genetics of a pheromonal difference
contributing to reproductive isolation in Drosophila. Science. 265:1461-1464.
Datta SR, Vasconcelos ML, Ruta V, Luo S, Wong A, Demir E, Flores J, Balonze K,
Dickson BJ, Axel R. 2008. The Drosophila pheromone cVA activates a sexually
dimorphic neural circuit. Nature. 452:473-477.
David JR, Capy P. 1988. Genetic variation of Drosophila melanogaster natural
populations. Trends Genet. 4:106-111.
Ejima A, Smith BP, Lucas C, van der Goes van Naters W, Miller CJ, Carlson JR,
Levine JD, Griffith LC. 2007. Generalization of courtship learning in
Drosophila is mediated by cis-vaccenyl acetate. Curr Biol. 17:599-605.
Endler JA. 1992. Signals, signal conditions, andthe direction of evolution. Am Nat.
139:125–153.
Endler JA. 1993. Some general comments onthe evolution and design of animal
communication systems. Philos Trans R SocLond Ser B. 340:215–225.
Fay JC, Wu CI. 2000. Hitchhiking under positive Darwinian selection. Genetics.
155:1405-1413.
Frith C, Beehler B. 1998. The birds of paradise: paradisaeidae . Oxford: Oxford
University Press.
Fu YX, Li WH. 1993. Statistical tests of neutrality of mutations. Genetics. 133:693-709.
Goldman AL, Van der Goes van Naters W, Lessing D, Warr CG, Carlson JR. 2005.
Coexpression of two functional odor receptors in one neuron. Neuron.
45:661-666.
Hale LR, Singh RS. 1991. A comprehensive study of genic variation in natural
populations of Drosophila melanogaster. IV. Mitochondrial DNA variation and
the role of history vs. selection in the genetic structure of geographic populations.
Genetics. 129:103-117.
Hallem EA, Ho MG, Carlson JR. 2004. The molecular basis of odor coding in the
Drosophila antenna. Cell. 117:965-979.
Hill WG, Robertson A. 1968. Linkage disequilibrium in finite populations. Theor Appl
Genet. 38:226–231.
Hoikkala A, Päällysaho S, Aspi J, Lumme J. 2000. Localization of genes affecting
species differences in male courtship song between Drosophila virilis and D.
littoralis. Genet Res. 75:37-45.
Hollocher H, Ting CT, Wu ML, Wu CI. 1997. Incipient speciation by sexual isolation in
Drosophila melanogaster: extensive genetic divergence without reinforcement.
Genetics. 147:1191-1201.
Kreitman M. 1983. Nucleotide polymorphism at the alcohol dehydrogenase locus of
Drosophila melanogaster. Nature. 304:412-417.
Kreitman M, Aguadé M. 1986. Genetic uniformity in two populations of Drosophila
melanogaster as revealed by filter hybridization of four-nucleotide-recognizing
restriction enzyme digests. Proc Natl Acad Sci USA. 83:3562-3566.
Kurtovic A, Widmer A, Dickson BJ. 2007. A single class of olfactory neurons mediates
behavioural responses to a Drosophila sex pheromone. Nature. 446:542-546.
Lachaise D, Cariou M-L, David JR, Lemeunier F, Tsacas L, Ashburner. M. 1988.
Historical biogeography of the Drosophila melanogaster species subgroup. Evol
Biol. 22:159-225.
Lande R. 1981. Models of speciation by sexual selection on polygenic traits. Proc Natl
Acad Sci USA. 78:3721-3725.
Leal M, Fleishman LJ. 2002. Evidence for habitat partitioning based on adaptation to
environmental light in a pair of sympatric lizard species. Proc Biol Sci.
269:351-359.
Librado P, Rozas J. 2009. DnaSP v5: a software for comprehensive analysis of DNA
polymorphism data. Bioinformatics. 25:1451-1452.
Marcillac F, Grosjean Y, Ferveur JF. 2005. A single mutation alters production and
discrimination of Drosophila sex pheromones. Proc Biol Sci. 272:303-309.
Marshall DC, Cooley JR. 2000. Reproductive character displacement and speciation in
periodical cicadas, with description of new species, 13-year Magicicada
neotredecem. Evolution. 54:1313-1325.
McDonald JH, Kreitman M. 1991. Adaptive protein evolution at the Adh locus in
Drosophila. Nature. 351:652-654.
Mouritzen P, Noerholm M, Nielsen PS, Jacobsen N, Lomholt C, Pfundheller HM,
Tolstrup N. 2005. ProbeLibrary: A new method for faster design and execution
of quantitative real-time PCR. Nat. Meth. 2: 313–316.
Nolte V, Schlötterer C. 2008. African Drosophila melanogaster and D. simulans
populations have similar levels of sequence variability, suggesting comparable
effective population sizes. Genetics. 178:405-412.
Pool JE, Aquadro CF. 2006. History and structure of sub-Saharan populations of
Drosophila melanogaster. Genetics. 174:915-929.
Ray A, van Naters WG, Shiraiwa T, Carlson JR. 2007. Mechanisms of odor receptor
gene choice in Drosophila. Neuron. 53:353-369.
Robertson HM, Warr CG, Carlson JR. 2003. Molecular evolution of the insect
chemoreceptor gene superfamily in Drosophila melanogaster. Proc Natl Acad
Sci USA. 100 Suppl 2:14537-14542.
Ryan MJ. 1998. Sexual selection, receiver biases, and the evolution of sex differences.
Science. 281:1999-2003.
Schlötterer C, Vogl C, Tautz D. 1997. Polymorphism and locus-specific effects on
polymorphism at microsatellite loci in natural Drosophila melanogaster
Populations. Genetics. 146:309-320.
Schlief ML, Wilson RI. 2007. Olfactory processing and behavior downstream from
highly selective receptor neurons. Nat Neurosci. 10:623-630.
Seehausen O, Terai Y, Magalhaes I, et al. 2008. Speciation through sensory drive in
cichlid fish. Nature. 455:620-626.
Shanbhag SR, Müller B, Steinbrecht RA. 1999. Atlas of olfactory organs of Drosophila
melanogaster: 1. Types, external organization, innervation and distribution of
olfactory sensilla. Int J Insect Morphol Embryol. 28:377-397.
Shanbhag SR, Müller B, Steinbrecht RA. 2000. Atlas of olfactory organs of Drosophila
melanogaster: 2. Internal organization and cellular architecture of olfactory sensilla. Arthropod Struct Devel. 29:211-229.
Simonsen KL, Churchill GA, Aquadro CF. 1995. Properties of statistical tests of
neutrality for DNA polymorphism data. Genetics. 141:413-429.
Singh RS, Rhomberg LR. 1987. A comprehensive study of genic variation in natural
populations of Drosophila melanogaster. I. Estimates of gene flow from rare
alleles. Genetics. 115:313-322.
Sokal RR, Rohlf FJ. 1981. Biometry: the principles and practice of statistics in
biological research. New York: W. H. Freeman and Co.
Spyropoulos IC, Liakopoulos TD, Bagos PG, and Hamodrakas SJ. 2004. TMRPres2D:
high quality visual representation of transmembrane protein models.
Bioinformatics. 20:3258-3260.
Stocker RF. 1994. The organization of the chemosensory system in Drosophila
melanogaster: a review. Cell Tissue Res. 275:3-26.
Tajima F. 1989. Statistical method for testing the neutral mutation hypothesis by DNA
polymorphism. Genetics. 123:585-595.
Takahashi A, Tsaur SC, Coyne JA, Wu CI. 2001. The nucleotide changes governing
cuticular hydrocarbon variation and their evolution in Drosophila melanogaster.
Proc Natl Acad Sci USA. 98:3920-3925.
Ting CT, Takahashi A, Wu CI. 2001. Incipient speciation by sexual isolation in
Drosophila: concurrent evolution at multiple loci. Proc Natl Acad Sci USA.
98:6709-6713.
Tunstall NE, Sirey T, Newcomb RD, Warr CG. 2007. Selective pressures on
Drosophila chemosensory receptor genes. J Mol Evol. 64:628-636.
Tusnády GE, Simon I. 2001. The HMMTOP transmembrane topology prediction server.
Bioinformatics. 17:849–850.
Vosshall LB, Wong AM, Axel R. 2000. An olfactory sensory map in the fly brain. Cell.
102:147-159.
Watterson GA. 1975. On the number of segregating sites in genetical models without
recombination. Theor Popul Biol. 7:256-276.
Wu CI, Hollocher H, Begun DJ, Aquadro CF, Xu Y, Wu ML. 1995. Sexual isolation in
Drosophila melanogaster: a possible case of incipient speciation. Proc Natl
Acad Sci USA. 92:2519-2523.
Xu P, Atkinson R, Jones DN, Smith DP. 2005. Drosophila OBP LUSH is required for
activity of pheromone-sensitive neurons. Neuron. 45:193-200.
Zeng ZB, Liu J, Stam LF, Kao CH, Mercer JM, Laurie CC. 2000. Genetic architecture
of a morphological shape difference between two Drosophila species. Genetics.
154:299-310.
52
Appendix
Fig. S1.—Location of PCR primers. Red indicates the modified nucleotides in primers.
53
Fig. S1.—(Continued)