行政院國家科學委員會專題研究計畫 成果報告
培養 Aspergillus awamori IFO4033 以生產 Feruloyl Esterase 酵素(第 3 年)
研究成果報告(完整版)
計 畫 類 別 : 個別型
計 畫 編 號 : NSC 98-2221-E-011-046-MY3
執 行 期 間 : 100 年 08 月 01 日至 101 年 07 月 31 日 執 行 單 位 : 國立臺灣科技大學化學工程系
計 畫 主 持 人 : 朱義旭
計畫參與人員: 博士班研究生-兼任助理人員:范陽 博士後研究:申娜薇
博士後研究:黃蓮香
報 告 附 件 : 出席國際會議研究心得報告及發表論文
公 開 資 訊 : 本計畫可公開查詢
中 華 民 國 101 年 07 月 28 日
中 文 摘 要 : 作用於植物細胞壁聚合物之 feruloyl esterase (FAE- II)為裂解植物細胞壁之重要酵素。此酵
素代表催化植物細胞壁聚醣與酚酸間酯鍵之合成或切斷之一 群水解酵素。這類酵素廣泛分布於植
物及微生物中。
在本計畫第一部分研究中,先在以麥稈、玉米芯或麥胚芽製 作成之粗半纖維素中培養
Aspergillus awamori strain IFO4033,再純化並定性所 生產之 FAE-II。生質量則以乾重法量
測。本實驗也決定 FAE-II 之最適酸鹼值及溫度 並有系統的 探討培養時間、碳及氮源等因素對
酵素生產之影響。FAE-II 先利 用離心分離,再以透析、離 子交換層析及微過濾得到均質酵素並
以 SDS-PAGE 及 IEF- PAGE 印證。在每一萃取純化步驟中測 量酵素之蛋白質含量及活性。不同
儲存條件對酵素活性之影響也是此部分實驗工作之一。
本計畫第二部分研究方面,先探討在不同溫度及不同阿魏酸 乙基酯濃度下 FAE-II 酯化反
應之動力學,然後求得動力學參數(最大速度 V max、
Michaelis 常數 K m 及二階速率常數 V/K);並 求出及討論溫度對此三動力學參數之影響。
在本計畫第三部分研究中,先探討在 25-85 o C 溫度範 圍內一經冷凍乾燥自然態之純化 A.
awamori FAE-II 及一交聯 FAE-II 聚集酵素(CLEAs)之熱穩 定性,再探討某些金屬離子及化學品
對 FAE-II 及 CLEAs 之影響。 藉由 DSC、TG 及 TGA,本研究 也探討酵素之熱失活現象。透過 FT-
IR 及 SEM 對 FAE-II 及 CLEAs 之結構分析並嘗試了解交聯是 否對 FAE-II 之粉末結構造成任何影響。
在第四部分研究中探討從 A. awamori 所得天然及活化之 FAE-II 酵素對甘蔗渣水解活性。
天然 FAE-II 酵素與鈣離子結合時會有活化效果,使得 V max 增加兩倍 並提高酵素對產物及
GdnHCl 去自然態(denaturation)之敏感度,對阿魏酸之 Km 則幾乎 無影響。本研究計算 FAE-II 酵
素與鈣離子結合所產生活化之相關動力學及熱力學參數 以 了解鈣離子如何作用,並經由 UV-
circular dichroism 光譜探討 GdnHCl 對 FAE-II 酵 素之 影響。此外,在不同溫度及酸鹼值下,本
研究也探討經鈣離子活化之 FAE-II 酵素 之密度及溶解度。
在最後一部分之研究中,成功的複製 AwfaeA cDNA、並 表現
於 E. coli 中•得到且純化重組
酵素。所得之重組 AwfaeA 酵素經結晶,取 x-ray 晶 相數 據,以 molecular replacement 可得到
1.2 Å; 之解析度。重組 AwfaeA 酵素與阿魏酸複合物 之分子結構也可決定到 1.0 Å; 之解析度。
中文關鍵詞: 阿魏酸,阿魏酸酯解酵素,酵素動力學
英 文 摘 要 : In part 1, the production, purification of FAE- II of A. awamori IFO4033 were carried out. FAE-II protein content and activity of the enzyme were measured. Biomass, pH and temperature optima of FAE-II were determined. Enzyme activities under different storage conditions were examined. In part 2, kinetic studies for the esterification of FAE-II were determined. Kinetic parameters (V max, K m, and V/K) were determined. In part 3, thermal stabilities of freeze dried native FAE-II and cross-linked FAE-II aggregate (CLEAs) were evaluated. Effects of metal ions and chemicals on the activity of both native FAE-II and CLEAs were examined. In part 4, the activity of native and activated FAE-II, which catalyzes the sugarcane bagasse hydrolysis, was studied.In the last part, AwfaeA cDNA was cloned and expressed in E. coli and the recombinant enzyme was purified and characterized. The recombinant AwfaeA enzyme was crystallized and its x-ray
crystallographic data were collected and explained.
The molecular structure of the recombinant AwfaeA enzyme-ferulic acid complex was also determined.
英文關鍵詞: Aspergillus awamori, Feruloyl esterases, Ferulic acid, Enzyme kinetics
行政院國家科學委員會補助專題研究計畫 ▓成果報告 □ 期 中 進 度 報 告
培 養
Aspergillus awamori
IFO4033 以 生 產 Feruloyl Esterase 酵素計畫類別:▓ 個別型計畫 □ 整合型計畫 計畫編號:NSC 98-2221-E-011 -046 -MY3 執行期間:2009 年 8 月 1 日至 2012 年 7 月 31 日
計畫主持人:朱義旭 共同主持人:
計畫參與人員:范陽(Ahmed E.Fazary),申娜薇(Novy S. Kasim),黃蓮香 (Lien Huong Hyunh)
成果報告類型(依經費核定清單規定繳交):□精簡報告 ▓完整報告
本 成 果 報 告 包 括 以 下 應 繳 交 之 附 件 :
□ 赴 國 外 出 差 或 研 習 心 得 報 告 一 份
□ 赴 大 陸 地 區 出 差 或 研 習 心 得 報 告 一 份
▓ 出 席 國 際 學 術 會 議 心 得 報 告 及 發 表 之 論 文 各 三 份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計畫、
列管計畫及下列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,▓一年□二年後可公開查詢
執行單位:台灣科技大學化工系
中 華 民 國 101 年 8 月 1 日
摘要
作用於植物細胞壁聚合物之 feruloyl esterase (FAE-II)為裂解植物細胞壁之重要酵素。此酵 素代表催化植物細胞壁聚醣與酚酸間酯鍵之合成或切斷之一群水解酵素。這類酵素廣泛分布於植 物及微生物中。
在本計畫第一部分研究中,先在以麥稈、玉米芯或麥胚芽製作成之粗半纖維素中培養 Aspergillus awamori strain IFO4033,再純化並定性所生產之FAE-II。生質量則以乾重法量 測。本實驗也決定FAE-II之最適酸鹼值及溫度 並有系統的探討培養時間、碳及氮源等因素對 酵素生產之影響。FAE-II先利 用離心分離,再以透析、離子交換層析及微過濾得到均質酵素並 以 SDS-PAGE 及 IEF- PAGE 印證。在每一萃取純化步驟中測量酵素之蛋白質含量及活性。不同
儲存條件對酵素活性之影響也是此部分實驗工作之一。
本計畫第二部分研究方面,先探討在不同溫度及不同阿魏酸乙基酯濃度下FAE-II酯化反
應之動力學,然後求得動力學參數(最大速度Vmax、Michaelis常數Km及二階速率常數V/K);並
求出及討論溫度對此三動力學參數之影響。
在本計畫第三部分研究中,先探討在 25-85 oC 溫度範圍內一經冷凍乾燥自然態之純化A.
awamori FAE-II及一交聯FAE-II聚集酵素(CLEAs)之熱穩定性,再探討某些金屬離子及化學品 對FAE-II及CLEAs之影響。 藉由DSC、TG及TGA,本研究也探討酵素之熱失活現象。透過FT- IR及SEM對FAE-II及CLEAs之結構分析並嘗試了解交聯是否對FAE-II之粉末結構造成任何影響。
在第四部分研究中探討從 A. awamori 所得天然及活化之 FAE-II 酵素對甘蔗渣水解活性。
天然 FAE-II 酵素與鈣離子結合時會有活化效果,使得 Vmax 增加兩倍 並提高酵素對產物及
GdnHCl 去自然態(denaturation)之敏感度,對阿魏酸之 Km 則幾乎 無影響。本研究計算 FAE-II 酵 素與鈣離子結合所產生活化之相關動力學及熱力學參數 以了解鈣離子如何作用,並經由 UV- circular dichroism 光譜探討 GdnHCl 對 FAE-II 酵 素之影響。此外,在不同溫度及酸鹼值下,本 研究也探討經鈣離子活化之 FAE-II 酵素 之密度及溶解度。
在最後一部分之研究中,成功的複製 AwfaeA cDNA、並 表現於 E. coli 中‧得到且純化重組 酵素。所得之重組 AwfaeA 酵素經結晶,取 x-ray 晶 相數據,以 molecular replacement 可得到 1.2 Å 之解析度。重組 AwfaeA 酵素與阿魏酸複合物之分子結構也可決定到 1.0 Å 之解析度。
關鍵詞:阿魏酸,阿魏酸酯解酵素,酵素動力學,
ABSTRACT
Feruloyl esterases (FAE-II) represent key tools for degradation of plant cell wall. It represents a diverse group of hydrolases catalyzing the cleavage and formation of ester bonds between plant cell wall polysaccharide and phenolic acid.
In the first part, the production, purification and characterization of FAE-II of Aspergillus awamori strain IFO4033 in a culture medium from crude hemicellulose preparation were carried out. FAE-II was extracted and purified by ion exchange chromatography and micro filtration.
Protein content and activity of the enzyme were measured in each step of extraction and purification. Biomass was determined. pH and temperature optima of FAE-II were also determined. The effects of culturing time, carbon and nitrogen sources on enzyme production were investigated. Enzyme activities under different storage conditions were examined.
In the second part, kinetic studies for the esterification reaction of FAE-II at different temperatures and various ethyl ferulate concentrations were determined. Kinetic parameters (Vmax, Km, and V/K) were determined. The effects of temperature on those 3 kinetic parameters were presented and discussed.
In the third part, thermal stabilities of freeze dried native A. awamori FAE-II and cross- linked FAE-II aggregate (CLEAs) were evaluated and discussed. Effects of metal ions and chemicals on the activity of both native FAE-II enzyme and CLEAs were examined. By means of DSC, TG, and TGA, thermal denaturation process was observed and explained. Structural analyses were made by using FT-IR and SEM to investigate whether the cross-linking had any effect on the powder structure of native FAE-II.
In the fourth part, the activity of native and activated FAE-II, which catalyzes the sugarcane bagasse hydrolysis, was studied. The activation of native FAE-II, by its binding with calcium ion, resulted in an increase of Vmax of two times, and a higher sensitivity of the enzyme to product and guanidine hydrochloride (GdnHCl) denaturation, with almost no change in the Km for ferulic acid. The kinetic and thermodynamic parameters associated with the activation of FAE-II enzyme by the interaction with calcium ions were calculated and explained. Densities and solubility of FAE-II activated by calcium ions were determined and discussed.
In the last part, AwfaeA cDNA was cloned and expressed in E. coli and the recombinant enzyme was purified and characterized. The recombinant AwfaeA enzyme was crystallized and its x-ray crystallographic data were collected and explained. The molecular structure of the recombinant AwfaeA enzyme-ferulic acid complex was also determined.
Keywords: Aspergillus awamori, Feruloyl esterases, Ferulic acid, Enzyme kinetics
TABLE OF CONTENTS
Page
摘要 I
ABSTRACT II
TABLE OF CONTENTS III
LIST OF FIGURES V
LIST OF TABLES VI
CHAPTER
1 INTRODUCTION 1
2 LITERATURE STUDIES 2
2.1 Nature, chemistry and biochemistry of ferulic acid 2
2.2 Carbohydrate esterases 2
2.3 Microbial production of feruloyl esterases 2
3 EXPERIMENTAL 4
3.1 Production, partial purification and characterization of FAE-II 4
3.1.1 Microorganism 4
3.1.2 Treatment of substrates 4
3.1.3 Submerge fermentation 4
3.1.4 Enzyme extraction 4
3.1.5 Enzyme purification 5
3.1.6 Enzyme activity assay 5
3.1.7 Biomass estimation 5
3.2 Biochemical studies of FAE-II 5
3.2.1 Cross-linked FAE-II aggregates 5
3.2.2 Enzyme activities of native and CLEAs of FAE-II 6 3.3 Expression of FAE-II in Escherichia coli 6
3.3.1 Strains and plasmids 6
3.3.2 PCR amplification and cloning 6
3.3.3 Expression in E. coli 6
3.3.4 AwfaeA protein extraction and purification 7
3.3.5 AwFaeA crystallization 7
4 RESULTS AND DISCUSSION 8
4.1 Production, partial purification and characterization of FAE-II 8
4.2. Studies on Kinetics of FAE-II 10
4.3. Studies on native and cross-linked aggregates of FAE-II 11 4.4 Stability and solubility studies of native and activated FAE-II 14
4.5 Expression of FAE-II in E. coli 18
4.5.1 AwfaeA cDNA cloning, expression, purification and characterization
18
5 CONCLUSIONS 20
REFERENCES 22
LIST OF FIGURES
FIGURE NAME Page
4-1 Vo (μmol/min), versus 10-4×[S] (mol.dm-3), for the esterification reaction of FAE-II
10
4-2 Effect of Ca2+ on the relative activity of FAE-II for the sugarcane bagasse hydrolyzed reaction in water solution. (□) with GdnHCl = 0.5 mol.dm-3; (■) without GdnHCl
15
4-3 Temperature stability of native and activated FAE-II for the sugarcane bagasse hydrolyzed reaction in water solution. (■) Native enzyme; (□) activated enzyme with [Ca2+] = 0.3 mol.dm-3
15
LIST OF TABLES
TABLE NAME Page
4-1 Purification of FAE-II in culture medium containing different carbon
source (substrate) 9
4-2 Kinetic parameters of esterification reaction of FAE-II 11 4-3 Effect of temperature on the relative activities of both native FAE-II
and CLEAs. Thermal stabilities were evaluated on a freeze dried purified FAE-II enzyme and CLEAs preincubated in 25-85 oC, pH~5.5
12
4-4 Effect of metal ions on the relative activities of FAE-II and CLEA.
Enzyme activities was evaluated on a freeze dried purified FAE-II enzyme and CLEAs preincubated for 60 min, with metal ions (5 mM, 10 mM and 15 mM) at 55 oC, pH~5.5
13
4-5 Effect of chemical additives on the relative activities of native FAE-II and CLEA. Enzyme activities were evaluated on a freeze dried purified FAE-II enzyme and CLEAs preincubated for 60 min, with chemicals (5 mM, 10 mM and 15 mM) at 55 oC, pH~5.5
14
4-6 Kinetic parameters of native FAE-II and activated FAE-II 16 4-7 Densities (g · cm-3) and solubilities of activatedFAE-II in water
solutions
17
4-8 Purification of AwfaeA recombinant protein from E. coli BL21 cells 19 4-9 Enzymatic properties of purified AwfaeA recombinant enzyme
expressed in E. coli. 19
CHAPTER 1: INTRODUCTION
Filamentous fungi, including the genus Aspergillus, are major producers of enzymes that have important applications. Aspergillus awamori is a common fungus used widely in koji- making for awamori, a traditional Japanese spirits (shochu), and simultaneously produces various enzymes including several xylanases and cellulases.
Feruloyl esterases (FAE) are promising enzymes to have potential contributions in research and industry. Microbial FAE acting on plant cell wall polymers represent key tools for degradating cell wall polysaccharides, modifing physical and chemical properties of plant cell walls and components. There has recently been interest in potential applications of FAE due to their roles in many biotechnological processes and in their potential applications in obtaining ferulic acid from agro-industrial waste. Two of the problems to commercialize the use of FAE are:
to date there is no commercial full purified FAE and the lack of sufficient enzyme stocks. Thus, efforts have to be made in order to achieve cheap overproduction of FAE in heterologous hosts and also their modification by chemical means or protein engineering to obtain more robust and active purified FAE-II enzymes.
One of the more satisfactory criteria for FAE enzyme purification is the nature of its solubility curves as the data obtained from this curves will be very useful for determining the degree of purity of FAE-II enzyme. The solubility studies would be of great help both for research and for the large scale production of the enzyme. Results obtained from the biophysico- chemical and thermo kinetic studies on FAE will help advancing the use of that enzyme.
Although recombinant enzymes and native enzymes are manufactured to meet the same rigorous quality control standards, it is recombinant enzymes that produce a purer product with less processing time. So considering the remarkable impact of FAEs in various industrial and medical applications, it is anticipated that the structural studies will provide an initial framework for the rational design of novel enzymes with improved biotechnological potential.
This work describes: (1) Production, partial purification and characterization of FAE by A.
awamori in submerged fermentation. (2) Studies on temperature dependent kinetics of native A.
awamori FAE-II in water solutions. (3) Biochemical studies on native and cross-linked aggregates of A. awamori FAE-II. (4) Stability and solubility studies of native and activated A.
awamori FAE-II. (5) Expression of FAE-II from A. awamori in Escherichia coli; characterization and crystal studies of the recombinant enzyme.
CH C HA AP PT TE ER R 2 2: : LI L IT TE ER RA AT TU UR RE E S ST TU UD DI IE ES S
2.1 Nature, chemistry and biochemistry of ferulic acid
Ferulic acid (FA) is the most abundant hydroxycinnamic acid derived from phenolic compounds (Ou and Kwok, 2004), distributed widely throughout the plant kingdom. It is said that FA supplies hydrogens to free radicals with phenolic-OH groups to provide the antioxidation effect. FA also has an active oxygen erasing function and the effect has been reported to be similar to superoxide dismutase, known as the enzyme that protects living bodies from the toxicity of active oxygen. FA is believed to suppress melanin generation by antagonizing tyrosine because their chemical structures are similar. It is also believed to powerfully absorb the harmful UV. It is listed as an “oxidation inhibitor” in the “food additive list”.
Today, commercial natural FA is mainly produced from γ-oryzanol in rice bran oil, although plant cell wall materials contain more FA. One of the main reasons is that FA cross-links with polysaccharides, and it is not easy to release FA from polysaccharides and purify it.
Fortunately, two methods were developed (Faulds and Williamson, 1999) to break the cross-link and release FA from plant cell wall materials. One is an enzymatic method using FAEs. Although there was extensive research on the preparation of FA using FAEs, this is not a practical way to produce commercial FA because of high cost of the enzyme. The similarity of chemical structures between FA and vanillin has led to the development to transform FA into vanillin (Zheng et al., 2007; Brunati et al., 2004; Mathew et al., 2007).
2.2 Carbohydrate esterase
Two enzymes, feruloyl and cinnamoyl esterases (FAE-B), purified from A. niger strains, have different physicochemical characteristics and catalytic properties against cinnamoyl model substrates (Ralph et al., 1994). These enzymes are exocellular and the corresponding expression is inducible. A new strain of A. niger I-1472 was shown to produce numerous polysaccharide- degrading enzymes as well as esterases that released FA from natural feruloylated oligosaccharides, when grown on sugar beet pulp and maize bran (Szwajgier et al., 2005).
2.3 Microbial production of feruloyl esterase
Most research on the microbial production of FAE to date involve the isolation, purification, and characterization of FAE derived from a wide range of microorganisms, as well as the enzymatic release of the products from cell wall degradation.
Esterases like FAE and cinnamic acid esterase are produced from Aspergillus sp. using either solid state or submerged fermentations. Solid-state fermentation (SSF) is defined as a fermentation process in which microorganisms grow on solid materials without the presence of
free liquid. In SSF, productivity per reactor volume is much higher than that in submerged culture (Roche et al., 1994). Operational cost is lower (Donaghy and McKay, 1995). Solid culture processes are practical for complex substrates including agricultural, forestry, and food processing residues and wastes, used as inducing carbon sources for the production of FAEs. A.
niger (Asther et al., 2002; Panagiotou et al., 2006), different Penicillium sp. (Asther et al., 2002;
Panagiotou et al., 2006; Donaghy and McKay, 1997; Castanares et al., 1992) and the thermophilic fungus Sporotrichum thermophile (Topakas et al., 2003) were capable of producing FAEs when grown on agricultural residues such as sugar beet pulp, wheat bran, wheat straw and brewer’s spent grain under SSF. Penicillium brasilianum (Panagiotou et al., 2006) was the more effective FAE producer under SSF.
CHAPTER 3: EXPERIMENTAL
3.1 Production, partial purification and characterization of FAE-II 3.1.1 Microorganism
A. awamori strain IFO4033 used in this study was obtained from Food Research and Development Institute, Bioresource Collection and Research Centre (BCRC Number: 31509), Hsinchu, Taiwan. The stock culture was maintained on potato-dextrose-agar (PDA) slants and stored at 4oC.
3.1.2 Treatment of substrates
The row material as carbon source (wheat straw, corn cobs and wheat germ), supplemented by mineral solution prior to heat sterilization (120°C, 30 min) were prepared as follows: 1) Wheat straw (WS) and corn cobs (CC) were washed, dried, milled and sieved (50 mesh) before use. Each substrate was given alkali treatment with 5% (w/v) 1 mol.dm-3 NaOH for 18 h at room temperature. After treatment, the substrates were thoroughly washed with water to neutral pH, dried and reground to 50 mesh sizes; 2) De-esterified wheat straw (DEWS) and de- esterified corn-cobs were prepared by incubating wheat straw and corn-cobs (0.75 g, containing 0.2% fatty acid (w/w), alkali-extractable fatty acid) overnight in 100 ml NaOH (0.1 mol.dm-3), in a covered flask, under gentle agitation. The whole mixture was dialyzed extensively against water;
3) De-starched wheat straw (DSWS) and de-starched corn-cobs were obtained by incubation wheat straw and corn-cobs in 0.25% (w/v) potassium acetate for 10 min at 95°C. The treated straw was then washed and dried prior to use.
3.1.3 Submerge fermentation
A 3-stage cultivation process was employed: 1) A. awamori was grown on PDA slants for 3 days at 30oC; 2) Next, 25 ml de-ionized water was added to a PDA slant and aliquot (5 ml) of the mixture was used to inoculate Erlenmeyer flask (500 ml) containing 250 ml of enzyme production medium [0.1 % bactotryptone + 0.5% yeast extract + 0.3 % sucrose + 0.1 % NaNO3
(nitrogen source) + 0.001 % FeSO4.7H2O + 2% treated substrate (carbon source), adjusted to pH 5.5]. The 500 ml-baffled Erlenmeyer flask was incubated at 30oC for 3 days in an orbital shaker (250 rpm.) for mycelium production; 3) Finally, inoculum of mycelial suspension (5% v/w) was added again to the enzyme production medium and the culture was grown for 5 days in 500 ml- baffled Erlenmeyer flask with a working volume of 250 ml.
3.1.4 Enzyme extraction
After ca 5 days, FAE-II was extracted from the fermented culture broth with 5-fold (v/w) distilled water by shaking (150 rpm) at room temperature for 60 min. The suspended material and fungal biomass were separated by centrifugation (15,000 rpm for 15 min). Tthe supernatant was re-centrifuged at 20,000 rpm for 30 min. The resulting clarified supernatant solution was collected as the source of crude enzyme and its protein content and enzyme activity were
estimated as described below.
3.1.5 Enzyme purification
The crude cell-free extract was equilibrated by microfiltration (0.45 μl) with 50 mmol.dm-
3 tris-HCl buffer, pH6 and applied onto a DEAE cellulose column (2.5 cm × 25 cm), equilibrated with the same buffer. The column was first washed with 1500 ml of 20 m mol.dm-3 tris-HCl buffer, pH~6, and then eluted with 200 ml 100 mmol.dm-3 NaCl solution prepared in the same buffer. Fractions (50 ml) were collected and two active fractions were collected (the first two DEAE-cellulose fractions). The concentrated second fraction was reloaded onto a second DEAE column (2.5 cm × 25 cm), equilibrated with the same buffer and a linear gradient (200 ml.h−1) of 30 mmol.dm-3 NaCl in 1200 ml of the same buffer was applied. Aliquots of the pooled active fractions (2nd DEAE-cellulose fraction) were concentrated by microfiltration, equilibrated with 20 mmol.dm-3 tris-HCl (pH~6). Saturated ammonium sulphate solution (0.8 mol.dm-3)was run slowly (about 10 ml/min) from a burette with stirring to a portion (10 ml) of the second DEAE- cellulose fraction to obtain the desired final salt concentration (Di Jeso et al., 1968). pH of the solution was readjusted if necessary. The two phases were separated by centrifugation at 25,000 rpm and 6°C for 30 min.
The results were expressed as total protein or activity from 10 ml extract. This purification step yielded one (NH4)2SO4 feruloyl active fraction.
3.1.6 Enzyme activity assay
The activity of FAE was assayed by the analysis of FA released from ethyl derivative of ethyl ferulate. One unit of FAE activity was defined as the amount of enzyme that liberates 1 μmol FA per minute.
3.1.7 Biomass estimation
Indirect method was adopted for biomass determination (Terebiznik and Pilosof, 1999;
Mitchell, 1992). In this work, we measured the fungal biomass gravimetrically. Culture medium was filtered through a Millipore filter (0.45 μm). The mycelium retained was washed with a saline solution (NaCl, 0.9%) and samples were dried to constant weight at 60oC for 48 h.
Biomass concentration was reported as g dry weight per mL culture medium (mg/mL).
3.2 Biochemical studies of FAE-II 3.2.1 Cross-linked FAE-II aggregates
FAE-II (100 mg per ml) was dissolved in phosphate buffer (0.15 M, pH ~ 6.5). Aggregation was achieved with the addition of (NH4)2SO4. Different concentration of (NH4)2SO4 was added stepwise at regular intervals with stirring at 10°C. Then the super saturated solution was kept undisturbed for 48 h. Enzyme activity of the precipitate was measured. The aggregates formed were separated by centrifugation (10,000 rpm for 15 min). One gram of FAE-II aggregates was cross-linked by using glutaraldehyde solution at two different concentrations (3 and 5%) in 1- propanol for 30 min at room temperature. After cross-linking, it was washed several times with
phosphate buffer (pH~6.5) containing 150 mM NaCl, to remove excess glutaraldehyde and stored in the same buffer. Enzyme activity was then measured. The preparation of FAE-II aggregates was carried out in the presence of triton X-100 to improve the aggregation stability.
3.2.2 Enzyme activities of native and CLEAs of FAE-II
The effect of metal ions, detergents, alcohols, inhibitors and chelating agent EDTA on the activity of purified FAE-II enzyme and cross-linked feruloyl esterase aggregates were determined by incubating the enzyme at 55°C for 1 h in 50 mM sodium phosphate buffer (pH~6) containing 150 mM NaCl of each compound mentioned. After incubation, relative activity was measured.
3.3 Expression of FAE-II in Escherichia coli 3.3.1 Strains and plasmids
AwfaeA cDNA gene was provided by Professor Takuya Koseki (Koseki et al., 1998).
Escherichia coli (strain BL21), supplied by Invitrogen, USA, grown at 37oC in Luria-Bertani broth containing kanamycine (50 μg/ml), as the strain carries a kanamycin-resistance plasmid were used for cloning expression and protein expression. pTZ57R/T vector was purchased from Fermentas, Novagen (InsT/AcloneTM PCR product cloning kit).
3.3.2 PCR amplification and cloning.
AwfaeA cDNA gene was amplified by PCR. A set of specific oligonucleotide primers;
sense primer (forward) 5’- AAgAATTCATgAAgCAATTCTCCgCAAA-3’ and antisense primer (reverse) 5’-AAAAAgCTTTCACCATgTACAggCTCCgC-3’, based on the nucleotide sequence information for the target AwfaeA gene, was used for specific amplification. PCR was carried out in 50 μl using a Thermocycler (Bio- Rad, Hercules, CA, USA) under the following conditions:
After an initial 5 min denaturation at 94°C, each cycle consisted of 60 s at 94°C, 30 s at 62°C and 60 s at 72°C. At the end of the 32 cycles of amplification, a final extension was continued for 30 min at 72°C. The PCR products were analyzed by electrophoresis on a 1% agarose gel and photographed. The size markers used to estimate PCR products were the 1 kbp DNA ladders (Fermentas). The forward and reverse primers were designed using GenRuner Software according to the published full-length cDNA sequence of the AwfaeA cDNA gene in Gene Bank database on NCBI (National Center for Biotechnology Information; http://www.ncbi.com) with accession No. AB032760 and 846 bp.
3.3.3 Expression in E. coli
Amplified PCR products were purified using a QIAquickTM gel extraction kit following the manufacturer's recommendations. The purified PCR products were ligated to pTZ57R/T cloning vector using T4 DNA ligase (InsT/AcloneTM PCR product cloning kit, Fermentas), following the manufacturer's protocol. For high expression of AwfaeA, the recombinant plasmid (pT-AwfaeA) was used to transform competent E. coli TBL21 strain. The cloned AwfaeA was confirmed by PCR, restriction enzymes (HindIII and EcoRI) digestion and nucleotide sequencing. E. coli
BL21carrying pT-AwfaeA was grown at 37°C with 200 rpm overnight in LB medium containing 50 μg/mL kanamycin. When absorbance (OD) at 600 nm reached 0.6, 0.03 mM IPTG was added to the culture, which was then incubated for an additional 3 h at 30oC. Next, the expressed recombinant cells were harvested from a culture broth by centrifugation (10000g, 20 min) and washed twice with 10 mM Tris–HCl buffer (pH 6.0). The cells were resuspended in phosphate- buffered saline (PBS), pH 7.4, and stored at −70°C.
3.3.4 AwfaeA protein extraction and purification
BugBuster Master Mix Protein Extraction Reagent (Novagen) was used to extract the expressed proteins from bacterial lysates. The extraction was carried out according to a standard procedure (Novagen, User protocol TB245 Rev. E 0304, P.8). The inclusion bodies containing AwFaeA in insoluble pellets (precipitate) were washed twice with buffer containing 50 mM Tris–
HCl (pH 8.0), 50 mM NaCl and 0.2% (v/v) Triton X-100. Solublization and refolding of the washed inclusion bodies was carried out using the Protein Refolding Kit (Novagen). The solublization buffer was supplemented with folding buffer, 0.5 mM dithiothreitol (DTT) for correct folding of disulfide bonds and with 1% N-lauroylsarcosine. The solubilized proteins were dialyzed against 20 mM Tris–HCl pH 8.0 four times at 4°C. After solublization and refolding, the recombinant protein was purified by nickel–nitrilotriacetic acid (Ni–NTA) metal-affinity chromatography matrices. Ni-NTA superflow resin (Qiagen, Hilden, Germany) was packed and equilibrated with buffer (20 mM Tris pH 8.0, 200 mM NaCl). The target protein (AwFaeA) was eluted with a stepwise gradient of imidazole from 20 to 200 mM. After concentration of the purified target protein (5 mg/ml), refolded AwFaeA was further purified by size exclusion chromatography. The concentrated purified protein was stored at −70°C for subsequent analysis.
Protein concentration was estimated using the Bradford method with BSA (Sigma Co., St Louis, MO) as standard (Bradford, 1976). The activity of AwfaeA was assayed by the analysis of free FA released from ethyl ferulate.
3.3.5 AwFaeA crystallization
A 5 mg/ml solution of the purified recombinant AwfaeA enzyme was used for crystallization trials. Setups were made using the hanging drop vapor diffusion method, in 24- well lnbro culture plates with drops on siliconized glass cover slides. The wells were sealed with high-vacuum laboratory grease. Drops contained 5 µl AwfaeA enzyme solution and 5 µl precipitant solutions from the corresponding 750 µl reservoir. The temperature was 20oC. A standard, coarse matrix crystallization screen was used. AwfaeA enzyme crystals appeared under the following conditions: 2 M (NH4)2SO4, 0.25 M NaAc, 0.1 M Tris-Cl pH 8.6, 5 days. The complex with FA was obtained by soaking crystal in 2 M (NH4)2SO4, 0.15 M tris-HCl, pH 8.6, 25 mM FA and 10% ethanol for about 2 h.
C C HA H A PT P TE ER R 4 4: : R RE ES SU UL LT TS S A AN ND D D DI IS SC CU US SS SI IO ON N
4.1 Production, partial purification and characterization of FAE-II
Submerged fermentation was used to produce FAE-II. The estimated biomass contents (mg/ml) for wheat straw (WS), de-esterified wheat straw (DEWS), de-starched wheat straw (DSWS), corn cobs (CC), de-esterified corn cobs (DECC), de-starched corn cobs (DCC) and wheat germ (WG) are 58.41 ± 3, 60.02 ± 1, 62.46 ± 2, 57.17 ± 2 56.39 ± 4, 58.12 ± 3 and 58.34 ± 2, respectively . The results show that the biomass contents were found to decrease in the order:
DSWS > DEWS > WS > WG > DSCC > CC > DSCC; 3) generally, total biomass decreases as the submerged fermentation proceeds. These results are in good agreements with previous data reported (Andrews, 1965; Dorta et al., 1994; Foster et al., 1971; Burbidge and Collier, 1968;
Kulkarni et al., 1999). The dry weight dropping of a submerged fermentation process is explained in terms of mass balance including oxygen consumed, carbon dioxide evolved, volatile products, evaporation of metabolically generated water and water consumed in polysaccharide hydrolysis (Andrews, 1965; Foster et al., 1971).
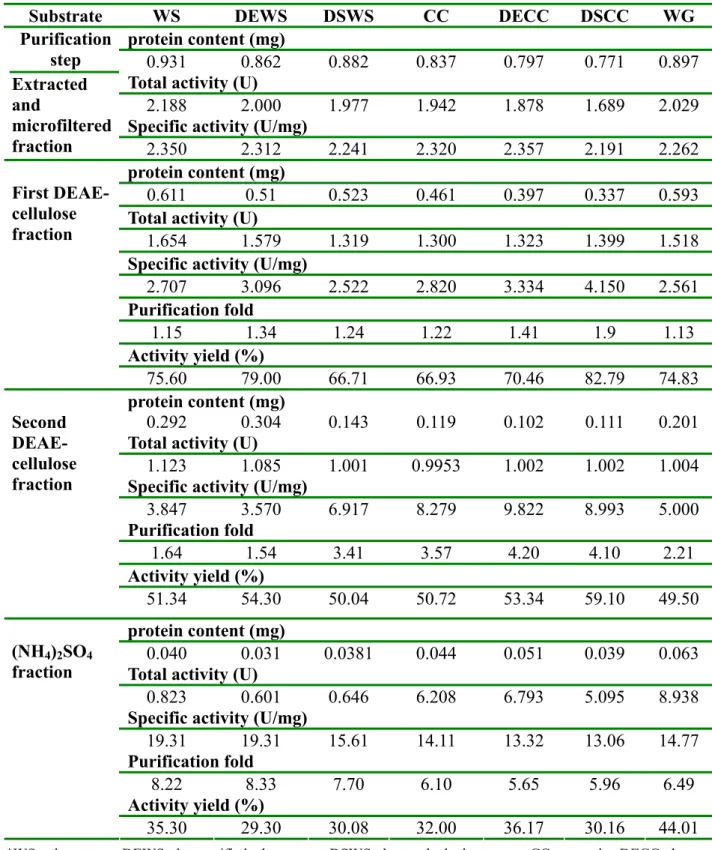
Centrifugation and microfiltration were partly successful in removing colored material and reducing the viscosity of the enzyme culture broth. DEAE cellulose gradient elution chromatography was used as the second step of purification. A distinct main peak of enzyme activity was eluted and well separated. In this step, enzyme activity yield was only ca. 85%-94%.
A second gradient-elution anion exchange DEAE cellulose column was applied and failed to produce a greater purification than that achieved in the first one. The average purification fold after the first step was ca. 1.5, as can be seen from Table 4-1. In the two DEAE trials, there was less purification achieved than initially expected. The enzyme activity yield (50%-65%) also decreased significantly after this two DEAE separation steps (Table 4-1). Salting-out of FAE-II is ideal as the final step in their purification. (NH4)2SO4 fractionation (ca. 1 mol.dm-3) of the second eluted DEAE cellulose fraction yielded 30% of the enzyme with 9-fold purification (Table 4-1).
Studies on the effect of temperature on FAE-II production showed that FAE-II production increased with temperature and reached maximum at 30oC. At 37oC a significant decline in activity was evident. Growth and FAE-II production almost completely stopped at 45oC.
Initial culture pH affects the production of FAE-II. Maximal FAE-II was obtained at pH~6. Growth and FAE-II production almost completely stopped at pH~10. A decrease in enzyme production was observed at lower pH (2-4).
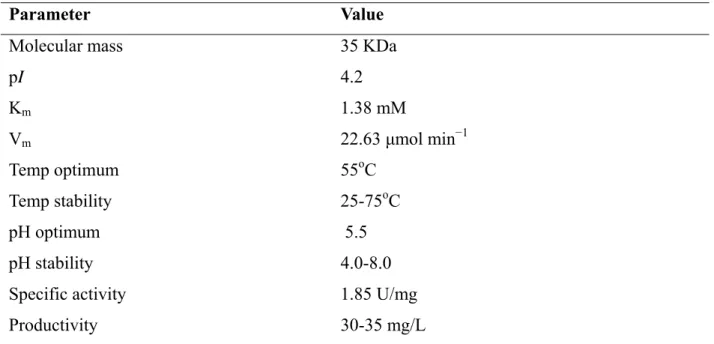
Gels of active FAE-II fractions from SDS-PAGE showed that a progressive purification has been achieved. FAE-II ammonium sulphate fraction appeared as a single protein band on a SDS-PAGE gel with a molecular mass about 31 KDa. This fraction was then concentrated and used in the subsequent characterization.
Table 4.1. Purification of FAE-II in culture medium containing different carbon source (substrate*)
*WS; wheat straw, DEWS; de-esterified wheat straw, DSWS; de-starched wheat straw, CC; corncobs, DECC; de- esterfied corn cobs, DSCC, de-starched corn cobs, WG; wheat germ.
Studies on the effect of temperature on activity of FAE showed that the enzyme possesses optimal activity at 55oC. Activities of the partially purified enzyme were measured at different pH, and the results show that FAE-II activity possesses pH optimum at 5.5. The enzyme shows
Substrate WS DEWS DSWS CC DECC DSCC WG
protein content (mg)
0.931 0.862 0.882 0.837 0.797 0.771 0.897
Total activity (U)
2.188 2.000 1.977 1.942 1.878 1.689 2.029
Specific activity (U/mg) Purification
step Extracted and
microfiltered
fraction 2.350 2.312 2.241 2.320 2.357 2.191 2.262 protein content (mg)
0.611 0.51 0.523 0.461 0.397 0.337 0.593
Total activity (U)
1.654 1.579 1.319 1.300 1.323 1.399 1.518
Specific activity (U/mg)
2.707 3.096 2.522 2.820 3.334 4.150 2.561
Purification fold
1.15 1.34 1.24 1.22 1.41 1.9 1.13 Activity yield (%)
First DEAE- cellulose fraction
75.60 79.00 66.71 66.93 70.46 82.79 74.83
protein content (mg)
0.292 0.304 0.143 0.119 0.102 0.111 0.201
Total activity (U)
1.123 1.085 1.001 0.9953 1.002 1.002 1.004 Specific activity (U/mg)
3.847 3.570 6.917 8.279 9.822 8.993 5.000
Purification fold
1.64 1.54 3.41 3.57 4.20 4.10 2.21
Activity yield (%) Second
DEAE- cellulose fraction
51.34 54.30 50.04 50.72 53.34 59.10 49.50
protein content (mg)
0.040 0.031 0.0381 0.044 0.051 0.039 0.063 Total activity (U)
0.823 0.601 0.646 6.208 6.793 5.095 8.938
Specific activity (U/mg)
19.31 19.31 15.61 14.11 13.32 13.06 14.77
Purification fold
8.22 8.33 7.70 6.10 5.65 5.96 6.49
Activity yield (%) (NH4)2SO4
fraction
35.30 29.30 30.08 32.00 36.17 30.16 44.01
relatively stable activity in pH 4 to 9. The high stability in acidic or alkaline conditions of A.
awamori strain IFO4033 may be advantageous in many biotechnological applications.
At room temperature, FAE-II loss 15% its initial activity after 2 days and only about 1/3 remained after one week. At 4oC no apparent loss of activity up to 4 weeks but after 4 weeks a marginal decrease in activity (5-10%) was found. The enzyme retained almost its full activity when stored at -20oC for up to 4 months
4.2. Studies on Kinetics of FAE-II
Initial rates (Vo) for esterification of FAE-II are shown in Figure 4-1. From Lineweaver- Burk plot, kinetic parameters (Table 4-2) were determined.
Guanidine hydrochloride (GdnHCl) and urea disrupt 3-D structure of protein and are used in the studies of protein unfolding and refolding. The kinetics of GdnHCl denaturation of FAE-II suggests partially unfolding of the protein structure. Structural changes produced by denaturants run in parallel to decreases in enzyme activity.
Figure 4-1 Vo (μmol/min), versus 10-4×[S] (mol.dm-3), for the esterification reaction of FAE-II; (□) 288 K; (■) 298 K; (Δ) 308 K; (▲) 318 K; (○) 328 K; (●) 338 K; (+) 348 K; (×) 358 K. (a) without guanidine hydrochloride (GdnHCl) (b) with GdnHCl
From findings shown in Table 4-2, we see that Vo increases from 288 K to 328 K and then decreases a little from 338 K to 358 K. However, the enzyme is still active even at 368K. As the substrate concentration increases, Vo increases up to a certain concentration, and then remains constant. Vm increases 7-fold from the minimal temperature (288 K) to the maximal temperature (328 K) and almost 2-fold from physiological temperature to 50 K. The Michaelis constant, Km, also shows a similar trend but with only a 2-fold increase from the minimal to maximal temperature with no significant increase at any given temperature as seen with Vm values. As with the first order rate constant, the second order rate constant, V/K, shows a similar rate of increase, with the values increasing 5-fold from minimal to maximal temperature. The kinetic parameters for each temperature are listed in Table 4-2.
4.3. Studies on native and cross-linked aggregate of FAE-II
Thermal stability on a freeze dried purified FAE-II enzyme and on a CLEAs in 25-85oC is reported in Table 4-3. FAE-II and CLEAs have optimum temperature at 55oC and optimum pH at 5.5 (Table 4-3). The thermal stability profile of both native and CLEAs of feruloyl esterase clearly shows that both enzymes were stable at 55oC, showing a half life of 24 h. While at 65oC the enzyme in both forms retained almost 60% of the initial activity, after 3 h of incubation at 85oC more than 80% of enzyme activity was lost.
Table 4-2 Kinetic parameters of esterification reaction of FAE-II GdnHCl absent*
T(K) Vm (min-1) Km (m mmol.dm-3) V/K (min-1/ mmol.dm-3)
288 3.64±0.46 4.13±0.19 0.88±0.04
298 9.82±0.35 6.32±0.16 1.55±0.02
308 12.73±0.71 7.42±0.16 1.72±0.03
318 15.98±0.64 9.12±0.17 1.75±0.04
328 20.52±0.76 9.28±0.18 2.21±0.07
338 17.41±0.54 10.12±0.15 1.72±0.06
348 16.62±0.56 9.32±0.13 1.78±0.06
358 13.63±0.49 9.10±0.16 1.50±0.07
GdnHCl present*
T(K) Vm (min-1) Km (mmol.dm-3) V/K (min-1/ mmol.dm-3)
288 2.03±0.39 8.21±0.16 0.25±0.07
298 4.33±0.45 8.83±0.17 0.50±0.06
308 5.72±0.45 9.13±0.18 0.63±0.07
318 6.93±0.43 9.78±0.15 0.71±0.06
328 7.41±0.47 9.89±0.13 0.75±0.06
338 7.10±0.60 7.32±0.21 0.97±0.07
348 6.71±0.63 7.43±0.16 0.90±0.06
358 6.72±0.58 6.84±0.18 0.98±0.06
*Guanidine hydrochloride (GdnHCl) (1.5 mol.dm-3 in 50 mmol.dm-3 phosphate buffer (pH~5.5) containing 0.15 mol.dm-3 NaCl)
The effect of metal ions on FAE-II (native and CLEAs) activity is summarized in Table 4- 4. Ca(II) and Mn(II) increase the activity of enzyme by almost 170 % and 120 %, respectively.
While other metal ions [K(II), Na(II), Fe(II), Zn(II), Mg(II)] caused a decrease of enzyme activity by 30%. Yet Cu(II) andCo (II) at the same concentration led to decrease of enzyme activity by abouut 70%. Hg(II) reduces the enzyme activity by almost 85 %. Our results agree well with those reported that Hg(II) inhibited the esterases of Arthrobacter viscosus NRRL B-1973 (Cui et al., 1999), Propionibacterium freudenreichii ssp. freudenreichii ITG 14 (Kakariari et al., 2000), Bacillus subtilis NRRL 365 (Meghji et al., 1990) and Arthrobacter nicotianae 9458 (Smacchi et al., 2000).
The results shown in Table 4-5 indicate that both ionic (SDS) and non ionic surfactants (tween 80 and triton X-100) inhibit enzyme activity by 20 to 40 %. No significant difference was Table 4-3 Effect of temperature on the relative activities of both native FAE-II and CLEA.
Thermal stabilities were evaluated on a freeze dried purified FAE-II enzyme and CLEA preincubated at 25-85 oC, pH~5.5.
Relative Activity (%)
FAE-II Temperature (oC)
Time (min) 25 35 45 55 65 75 85
0 100 100 100 100 100 100 100
30 94.21 91.25 93.24 97.21 91.12 83.70 76.24
60 88.32 83.31 86.41 91.22 79.32 64.72 61.51
90 83.42 76.46 77.35 87.30 68.42 53.82 50.31
120 77.51 70.52 71.31 80.51 64.61 40.64 37.23
150 73.32 66.25 68.65 76.43 58.41 31.76 27.43
180 64.40 62.36 61.98 70.61 47.52 23.91 20.68
210 58.87 57.46 53.74 64.52 41.61 19.83 16.81
240 52.15 53.85 49.86 59.34 38.63 12.64 12.90
270 49.37 46.32 46.74 58.30 32.70 9.76 6.76
CLEA Temperature (oC)
Time (min) 25 35 45 55 65 75 85
0 100 100 100 100 100 100 100
30 105.6 102 104.4 108.9 102.2 93.21 85.35
60 98.82 93.20 96.58 102.2 88.72 71.87 68.5
90 93.21 85.30 86.47 97.72 76.36 59.52 56.15
120 86.47 78.60 79.73 89.84 71.87 44.92 41.55
150 81.98 74.13 76.36 85.35 65.13 34.81 30.32
180 71.87 69.62 68.50 78.61 52.78 25.83 22.46
210 65.13 64.04 59.52 71.87 46.04 21.34 17.97
240 58.40 59.50 55.03 66.26 42.67 13.48 13.48
270 55.03 51.73 51.66 65.13 35.94 10.11 6.738
observed between the effect of ionic detergents and non ionic one, even the ionic detergents should typically modify protein structure to a greater extent than the non ionic one. A zwitterionic detergent, CHAPS, caused a slight inhibition on esterase, which is more effective in protein- protein interaction than non-ionic detergents and their extent of protein denaturation are less than those of ionic detergents. The degree of modification varies with the individual protein and the particular detergent. Neugebauer et al. (1990) proved that the non-ionic detergents stimulated the esterase enzyme and most of the non-ionic detergents are non-denaturing, but are less effective at disrupting protein aggregation.
Data presented previously for the inhibition effect of EDTA indicate that enzyme activity did not drop to zero in the presence of EDTA (Dosanjh ad Kaur, 2002; Nawani et al., 1998). In our results, enzyme activity decreased by 45-50 % (Table 4-5). Enzyme activity was almost completely restored by adding any metal ion studied in this work as metal ions can form very strong and stable binary complexes (Holleman and Wiberg, 2001). However, we can not suggest that the enzyme requires metal ions for its activity.
From Table 4-5, we observe that enzyme activities lost by nearly 50% and 25%, when 30- 70% (v/v) of glycerol and 1-propanol were used, respectively. Urea is an important solute found in the intra- and extracellular compartments of marine elasmobranchs (Perlman and Goldstein, 1989) and in the mammalian kidney (Wolff and Balaban, 1990). The findings in this study show that after the addition of urea, 60-70% of the activity of the enzyme (native FAE-II and CLEAs) was lost (Table 4-5).
In the present work, we found that after the addition of GdnHCl, the relative activities were about 70% (Table 4-5). The GdnHCl-induced denaturation of the holo proteins is generally found to be highly reversible, enabling quantitative analysis of the effects of the mutations on protein stability. Because H2O2 is widely regarded as a cytotoxic agent, levels must be minimized Table 4-4 Effect of metal ions on relative activities of FAE-II and CLEA. Enzyme activities was evaluated on FAE-II and CLEA preincubated with metal ions at 55 oC, pH5.5 for 60 min.
Metal Ion Relative Activity (%)
5 mM 10 mM 15 mM
FAE-II CLEAs FAE-II CLEAs FAE-II CLEAs Ca2+ 162.21±6 183.14±3 171.32±3 188.13±6 173.11±3 192.23±3 Mg2+ 108.32±4 128.26±5 117.45±5 134.25±4 119.36±5 130.33±5 K + 56.57±6 62.23±4 65.32±6 69.64±6 63.64±4 73.11±6 Na+ 62.32±6 68.24±5 73.16±4 74.39±6 71.43±5 71.66±4 Fe2+ 72.91±4 78.36±5 80.65±5 84.48±4 78.33±5 88.34±5 Zn2+ 68.46±7 71.46±4 76.55±6 78.69±4 79.21±4 77.13±6 Co2+ 30.32±4 31.94±3 37.23±4 36.51±5 41.03±3 34.68±4 Cu2+ 27.21±5 29.85±4 33.64±3 35.44±4 37.36±4 37.87±3 Mn2+ 73.23±4 82.67±4 78.33±7 88.64±4 77.43±4 84.67±7 Hg2+ 17.21±4 15.32±5 18.37±5 19.94±4 12.99±5 11.46±5
by the action of antioxidant defense enzymes. In this work, we study the effect of H2O2 on the enzyme activity and the experimental findings listed in Table 4-5 show that the enzyme activity reduced by 70-80%. The thermal stability of FAE-II and CLEA was also experimentally evaluated by DSC The denaturation temperature (Td) and the denaturation enthalpy for pure enzyme FAE-II and CLEAs are 158.24oC and 188.26oC, respectively.
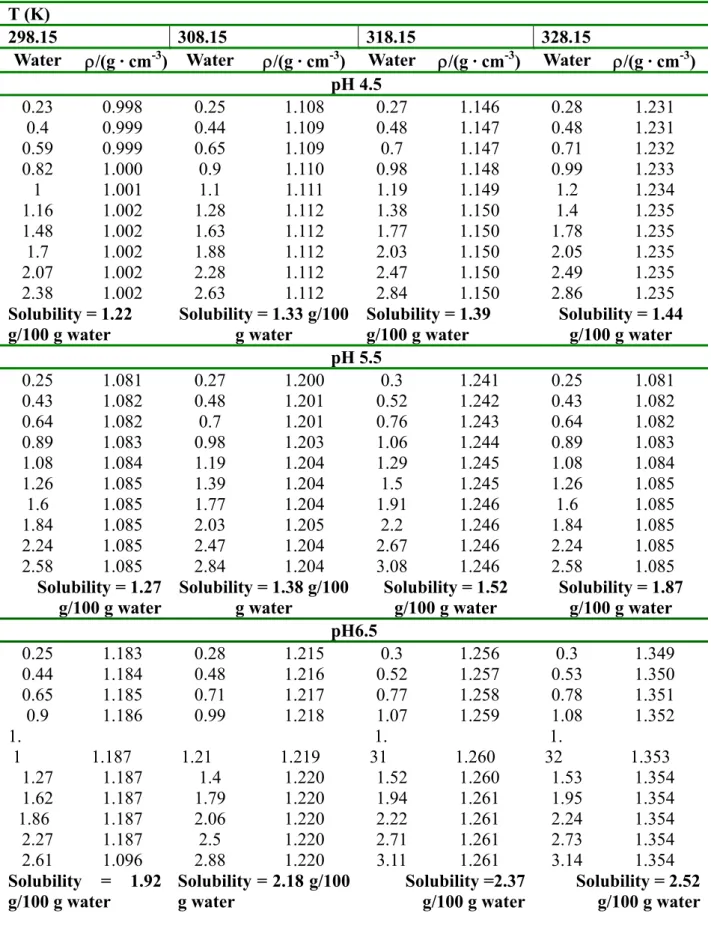
4.4 Stability and solubility studies of native and activated FAE-II
Experimental data obtained for the effect of calcium ions [Ca2+] on the relative activity of FAE-II for sugarcane bagasse hydrolyzed reaction at 328 K and pH 5.5 in water solution in the presence and absence of GdnHCl as enzyme chemical denaturant are presented in Figure 4-2. The thermal stability of native FAE-II and activated FAE-II by calcium ions are presented in Figure 4- 3. From Lineweaver-Burk double reciprocal and Eadie-Hofstee plots the kinetic parameters were determined (Table 4-6). The density and solubility of activated FAE-II are illustrated in Table 4-7.
As shown in Figure 4-2, one can conclude that Ca2+ has a significant effect on the activity of native FAE-II. The relative activity of FAE-II enzyme in the absence of GdnHCl increases with increasing Ca2+ concentration added to the native enzyme till a maximum relative activity of 257% is obtained at [Ca+2] = 0.27 mol.dm-3, and then starts to decrease with increasing Ca2+
concentration. The same behavior happens in the case with the presence of GdnHCl, as the relative activity increases to 131 % at [Ca+2] = 0.27 mol.dm-3, and then starts to decrease with increasing Ca2+ concentration.
Table 4-5 Effect of additives on relative activities of native FAE-II and CLEA. Enzyme activities were evaluated on FAE-II and CLEA preincubated with chemicals at 55 oC, pH5.5 for 60 min
Compound Relative Activity (%)
5 mM 10 mM 15 mM
FAE-II CLEAs FAE-II CLEAs FAE-II CLEAs Triton X-100 80.12±2 87.14±4 84.14±2 90.71±4 85.64±4 92.31±2
EDTA 54.32±3 58.23±3 58.34±2 63.64±3 56.75±2 66.23±2
SDS 63.13±3 67.45±4 69.46±4 72.31±4 71.91±4 76.51±4
H2O2 18.21±3 22.67±2 24.68±4 24.13±2 26.87±3 28.61±2
Urea 67.15±4 70.94±4 68.97±3 74.18±4 71.64±2 77.74±4
GdnHCl 24.34±2 28.64±3 31.64±2 33.25±3 34.31±2 37.69±3
CHAPS 70.40±4 76.34±4 74.49±2 80.28±4 77.18±4 83.97±3
glycerol 53.61±2 57.45±3 54.53±3 61.63±3 56.64±2 64.64±2 Tween 80 61.72±4 66.51±3 68.84±2 69.67±3 72.76±4 72.87±4 Propanol 73.82±3 78.37±4 76.67±2 82.94±4 79.31±2 85.36±2
Figure 4-2 Effect of Ca2+ on the relative activity of FAE-II for the sugarcane bagasse hydrolyzed reaction in water solution. (□) with GdnHCl = 0.5 mol.dm-3; (■) without GdnHCl.
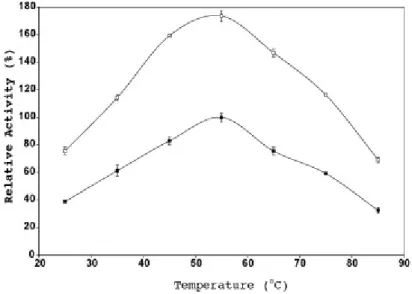
Thermal inactivation results plotted in Figure 4-3 indicates that calcium ions [Ca+2] = 0.2 mol.dm-3 increased the stability of the native FAE-II enzyme. However, the addition of a higher amount of calcium ions (≥0.3 mol.dm-3) induced a decrease in enzymatic thermostability, independent of the amount of Ca+2added.
Figure 4-3 Temperature stability of native and activated FAE-II for sugarcane bagasse hydrolyzed reaction in water solution. (■) Native enzyme; (□) activated enzyme with [Ca2+] = 0.3 mol.dm-3.
Vmax shown in Table 4-6 indicates that the incubation of native FAE-II in 0.2 mM Ca2+
resulted in about two-fold increase of Vmax (22.2 - 54.05) μmol.min-1 for native FAE-II and
activated FAE-II, respectively in the absent of GdnHCl; Vmax were (10.44, 25.25) μmol.min-1 for FAE-II and activated FAE-II, respectively in the presence of GdnHCl). KCat for activated FAE-II is two times higher than that for the native FAE-II with or without GdnHCl.
KCat/Km is an apparent second order rate constant that refers to the property and the reactions of free enzyme and substrate. Table 4-6 also show that KCat/Km for activated FAE-II is two times higher than that for the native FAE-II with or without GdnHCl.
In the present investigation, the stability of enzymes is defined by their half-life. The stabilization factor is the ratio of the half-life after treatment relative to that of the untreated enzyme. Stabilization for the activated FAE-II was higher in the absence of GdnHCl than with GdnHCl.
It is well known that protein solubility in a spiritual medium depends on solvent environment and relates to surface hydrophobic (protein–protein) and hydrophilic (protein–
solvent) interaction. We showed that the minimum solubility of FAE-II was always near the pI, as has been observed previously (Arakawa and Timasheff, 1985; Cohn and Edsall, 1943). The effect of temperature on the protein solubilities in different pH mediums is given in Table 4-7. From these data, we see the solubility increases with temperature. When solution temperature is raised high enough for a given time, the protein is denatured.
Table 4-6 Kinetic parameters of native FAE-II and activated FAE-II Without GdnHCl*
Vmax (μmol.min-1) KCat (min-1) Km (mM) KCat/Km (min-1/mM) Native FAE-II 22.2±0.46 4.44±0.46 2.50±0.15 1.78±0.14 Activated FAE-II 54.05±0.26 10.81±0.46 2.33±0.10 4.64±0.31
With GdnHCl*
Vmax (μmol.min-1) KCat (min-1) Km (mM) KCat/Km ( min-1/mM) Native FAE-II 10.44±0.42 2.09±0.46 2.49±0.10 0.84±0.09 Activated FAE-II 25.25±0.24 5.05±0.46 2.33±0.06 2.17±0.20
*GdnHCl: Guanidine hydrochloride (1.5 M in 50 mM phosphate buffer (pH5.5) containing 0.15 M NaCl). The Vmax, KCat, and Km values were determined from Lineweaver-Burk plots. The data given are average values ± S.D.
(Standard Deviations) of n = 3 independent experiments.
![Figure 4-1 Vo (μmol/min), versus 10 -4 ×[S] (mol.dm -3 ), for the esterification reaction of FAE-II; (□) 288 K; (■) 298 K; (Δ) 308 K; (▲) 318 K; (○) 328 K; (●) 338 K; (+) 348 K; (×) 358 K](https://thumb-ap.123doks.com/thumbv2/9libinfo/9127670.411835/20.892.181.591.540.1021/figure-vo-μmol-min-versus-esterification-reaction-fae.webp)