科技部補助專題研究計畫報告
利用基因轉殖小鼠研究凝血調節素在血管新生內膜及動脈硬化 中的角色(第3年)
報 告 類 別 : 成果報告 計 畫 類 別 : 個別型計畫
計 畫 編 號 : MOST 106-2314-B-006-045-MY3 執 行 期 間 : 108年08月01日至109年07月31日 執 行 單 位 : 國立成功大學醫學系內科學科
計 畫 主 持 人 : 李貽恆
計畫參與人員: 碩士級-專任助理:鍾幸君
本研究具有政策應用參考價值:■否 □是,建議提供機關
(勾選「是」者,請列舉建議可提供施政參考之業務主管機關)
本研究具影響公共利益之重大發現:□否 □是
中 華 民 國 109 年 10 月 28 日
中 文 摘 要 : 正常的動脈血管,凝血調節素(thrombomodulin, TM)只會表現在內 膜(intima)上的血管內皮細胞表面,負擔抗凝血的功能;血管損傷 後,中層和新生內膜(neointima)中的平滑肌細胞會表現出大量的 TM。本計畫的主要目的在探討血管平滑肌細胞所表現的TM的生理意 義。
培養人的主動脈SMCs顯示具有菱形形狀的合成表型並表達TM。TM組 合式誘導形態的紡錘形變化與收縮表型標誌物的表達增加和合成表 型標誌物的表達減少。 TM敲除不僅減弱了SMCs的增殖,而且降低了 腫瘤壞死因子-α誘導的核因子-κB激活和白介素6的產生。在頸動 脈結紮模型中,與野生型小鼠(SM22-cretg / TM + / +)相比,具 有SMC特異性TM缺失的轉基因小鼠(SM22-cretg / TMflox /
flox)在動脈壁的細胞增殖明顯更少。結紮後4週,SM22-cretg / TMflox / flox小鼠的新內膜面積和新內膜/培養基面積比較小。
我們的結果表明血管SMC綁定TM在SMC表型的變化中發揮作用。它還 會影響SMC細胞行為和損傷誘導的新內膜形成。
中 文 關 鍵 詞 : 新生內膜;平滑肌細胞;凝血酶調節素
英 文 摘 要 : Thrombomodulin (TM) is an endothelial cell membrane-bound anticoagulant protein
expressed in normal arteries. After vascular injury, medial and neointimal smooth muscle cells (SMCs) exhibit large amounts of TM. The purpose of this study was to investigate the physiological significance of vascular SMC-bound TM.
Cultured human aortic SMCs displayed a synthetic phenotype with a rhomboid-shaped morphology and expressed TM. TM knockdown induced a spindle-shaped change in morphology with an increased expression of contractile phenotype marker and decreased expression of synthetic phenotype marker. TM knockdown not only attenuated the proliferation of SMCs but also reduced tumor necrosis factor-α-induced nuclear factor-κB activation and interlukin-6 production.
In a carotid artery ligation model, transgenic mice with SMC-specific TM deletion (SM22-cretg/TMflox/flox) had
significantly less cellular proliferation in arterial walls compared with wild type mice (SM22-cretg/TM+/+). The
neointima area and neointima/media area ratio were smaller in SM22-cretg/TMflox/flox mice at 4 weeks after ligation.
Our results indicate that vascular SMC-bound TM plays a role in changes of the SMC phenotype. It also influences SMC cell behavior and injury-induced neointima formation.
英 文 關 鍵 詞 : Neointima; Smooth muscle cell; Thrombomodulin
科技部補助專題研究計畫成果報告
(□期中進度報告/■期末報告)
利用基因轉殖小鼠研究凝血調節素在血管新生內膜及動脈硬化中的角色
計畫類別:■個別型計畫 □整合型計畫
計畫編號:MOST 106-2314-B-006-045-MY3 執行期間:2017 年 8 月 1 日至 2020 年 7 月 31 日 執行機構及系所:
國立成功大學醫學系內科學科計畫主持人:李貽恒 共同主持人:吳華林
計畫參與人員:
鍾幸君/陳柏升/馮文翰本計畫除繳交成果報告外,另含下列出國報告,共 ___ 份:
□執行國際合作與移地研究心得報告
□出席國際學術會議心得報告
□出國參訪及考察心得報告
本研究
具有政策應用參考價值:
■否 □是,建議提供機關_______(勾選「是」者,請列舉建議可提供施政參考之業務主管機關) 本研究具影響公共利益之重大發現:■否 □是
中 華 民 國 109 年 10 月 28 日
附件一
目錄
中文摘要... II Abstract... III
1. Introduction ... 1
2. Materials and Methods ... 2
3. Results ... 7
4. Discussion... 10
5. Reference...13
Figure …... 18
Figure legends ...25
中文摘要
正常的動脈血管,凝血調節素(thrombomodulin, TM)只會表現在內膜
(intima)上的血管內皮細胞表面,負擔抗凝血的功能;血管損傷後,中層和新生內 膜(neointima)中的平滑肌細胞會表現出大量的TM。本計畫的主要目的在探討 血管平滑肌細胞所表現的TM的生理意義。
培養人的主動脈SMCs顯示具有菱形形狀的合成表型並表達TM。TM組合式 誘導形態的紡錘形變化與收縮表型標誌物的表達增加和合成表型標誌物的表達 減少。 TM敲除不僅減弱了SMCs的增殖,而且降低了腫瘤壞死因子-α誘導的核 因子-κB激活和白介素6的產生。在頸動脈結紮模型中,與野生型小鼠(SM22- cretg / TM + / +)相比,具有SMC特異性TM缺失的轉基因小鼠(SM22-cretg / TMflox
/ flox)在動脈壁的細胞增殖明顯更少。結紮後4週,SM22-cretg / TMflox / flox小鼠的
新內膜面積和新內膜/培養基面積比較小。
我們的結果表明血管SMC綁定TM在SMC表型的變化中發揮作用。它還會影 響SMC細胞行為和損傷誘導的新內膜形成。
關鍵字: 新生內膜;平滑肌細胞;凝血酶調節素
Abstract
Thrombomodulin (TM) is an endothelial cell membrane-bound anticoagulant protein
expressed in normal arteries. After vascular injury, medial and neointimal smooth muscle cells (SMCs) exhibit large amounts of TM. The purpose of this study was to investigate the physiological significance of vascular SMC-bound TM.
Cultured human aortic SMCs displayed a synthetic phenotype with a rhomboid- shaped morphology and expressed TM. TM knockdown induced a spindle-shaped change in morphology with an increased expression of contractile phenotype marker and decreased expression of synthetic phenotype marker. TM knockdown not only attenuated the proliferation of SMCs but also reduced tumor necrosis factor-α-induced nuclear factor-κB activation and interlukin-6 production. In a carotid artery ligation model, transgenic mice with SMC-specific TM deletion (SM22-cretg/TMflox/flox) had significantly less cellular proliferation in arterial walls compared with wild type mice (SM22-cretg/TM+/+). The neointima area and neointima/media area ratio were smaller in SM22-cretg/TMflox/flox mice at 4 weeks after ligation.
Our results indicate that vascular SMC-bound TM plays a role in changes of the SMC phenotype. It also influences SMC cell behavior and injury-induced neointima formation.
Keywords: Neointima; Smooth muscle cell; Thrombomodulin
1. Introduction
The pathophysiology of atherosclerosis and vascular injury-induced neointima involves endothelial injury as the initiating event followed by complex inflammatory and immune responses [1, 2]. Vascular smooth muscle cells (SMCs) are intimately involved during this process [3]. In the media of normal arteries, vascular SMCs assume a contractile phenotype and express a variety of SMC-specific contractile markers, such as α-smooth muscle actin (α-SMA) and calponin. Upon stimulation, the medial SMCs transform into a synthetic, proliferative and proinflammatory phenotype which is associated with decreased expression of contractile markers and increased expression of synthetic markers, such as vimentin and non-muscle myosin heavy chain (NM-MHC). Activated vascular SMCs are capable of robust proinflammatory responses by producing multiple cytokines, such as interleukin (IL)-6, which is the major regulator of downstream inflammatory cascades [4, 5]. The switch of vascular SMC phenotype can be induced by a wide range of factors [6], however, the definite molecular mechanisms underpinning vascular SMC phenotype transition remain to be fully elucidated.
Thrombomodulin (TM) isa cell membrane-bound glycoprotein which is only expressed on the vascular endothelium in normal arteries. TM binds thrombin, and the TM-thrombin complex accelerates the rate of protein C activation which acts as a physiological anticoagulant [7]. Binding of TM to thrombin also prevents thrombin- mediated protease-activated receptor-1 (PAR-1) activation and inhibits its
downstream proinflammatory effects on the endothelium [8]. In normal arterial walls, TM is only expressed on endothelial cells, but not medial vascular SMCs. Medial vascular SMCs have been shown to express large amounts of TM during
atherosclerosis and neointima formation after vascular injury [9-11]. Proinflammatory
cytokines and growth factors such as thrombin, cyclooxygenase-2-derived
prostaglandin and platelet-derived growth factor (PDGF)-BB have been shown to stimulate the expression of TM on cultured vascular SMCs [12-14]. Although the presence of TM on vascular SMCs in diseased arteries has been known for years, the physiological and pathological significance of SMC-bound TM remains largely undetermined. The expression of TM has been shown to be decreased on the
endothelium in atherosclerotic lesions and in vascular injury-induced neointima [11, 15]. Many researchers have speculated that the expression of TM on vascular SMCs is only a compensatory mechanism to balance endothelial TM downregulation, thereby maintaining vascular wall homeostasis. In recent years, TM has been shown to possess multiple direct cellular effects beyond its anticoagulant effects [8, 16-18]. We hypothesized that the presence of TM on activated SMCs may alter vascular SMC behaviors and influence vascular injury-induced neointima formation. In the current study, we observed the differences in cell morphology and behaviors between cultured aortic SMCs with and without TM expression. A complete loss of TM in TM
knockout transgenic mice causes embryonic lethality [19]. Therefore, we generated vascular SMC-specific TM-deficient mice using the Cre-loxP system to explore the role of vascular SMC membrane-bound TM in vivo.
2. Materials and methods 2.1. TM knockdown
Human aortic SMCs (Invitrogen, Carlsbad, CA) were maintained in medium 231 (Invitrogen) containing 5% smooth muscle growth supplement (Invitrogen) at 37°C in a humidified atmosphere of 5% CO2 and 95% air. Cultured human aortic SMCs were grown to 60-70% confluence and transfected with TM siRNA or control siRNA (25 nM) using DharmaFECT transfection reagents (Thermo Scientific, Lafayette, CO)
according to the manufacturer’s instructions. Cells were treated with siRNA for 48 hours before the following experiments. After transfection, the cells were harvested and the knockdown efficiency of TM was evaluated using immunoblotting.
2.2. Microarray analysis
Agilent microarray analysis was carried out according to Agilent Technologies’
guidelines. Briefly, RNA samples from cells treated with TM siRNA or control siRNA were collected, amplified and labeled using an Agilent Quick Amp Labeling Kit. The samples were hybridized to an Agilent whole genome oligo microarray and then scanned using an Agilent DNA microarray scanner. The results were imported into Agilent GeneSpring GX software (version 11.0) for further analysis. Differentially expressed genes were identified and the p values were calculated using the t-test.
2.3. TM overexpression
The pEGFPN1 vector was obtained from BD Biosciences Clontech (Palo Alto, CA) and the fluorescent protein-tagged TM expression vector, pEGFPN1-TM, was
constructed as previously described [16]. In brief, the human TM gene encoding TM amino acid residues 1–575 in chromosomal DNA was amplified by polymerase chain reaction (PCR) using a BamHI forward primer, TM719 (5′-
CGGGATCCCGGAATGCTTGG
GGTCCTGGTCCTTG-3′) and an EcoRI reverse primer (5′-GGAATTCGGAGTCT CTGCGGCGTCCGCT-3′). The 1.7-kb PCR product was digested with BamHI and EcoRI. The fragment was ligated to the expression vector pEGFPN1 and named pEGFPN1-TM. The pEGFPN1-TM vector was transfected into aortic SMCs using Polyjet transfection reagent (SignaGen Laboratories, Rockville, MD) for 24 hours and the transfection efficiency was analyzed by immunoblotting.
2.4. Immunoblotting
Nuclear and cytoplasmic fractions of aortic SMCs were separated using NE-PER Nuclear and Cytoplasmic Extraction Reagents (Thermo Fisher Scientific) following the manufacturer's instructions. Cell lysates were extracted from aortic SMCs and subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), followed by electroblotting. They were then probed with specific antibodies against human TM, IB-, p50, p65, α-tubulin, GAPDH (all from Santa Cruz Biotechnology) and SMC markers, including α-SMA, calponin, NM-MHC and vimentin (all from Abcam).
2.5. Proliferation assay
Cell proliferation in 96-well plates (3×103/well) was assessed using a WST-1 assay according to the manufacturer’s instructions (Roche, Madison, WI). The medium was replaced every 2 days. Ten L of WST-1 was added to the wells followed by
incubation for 1 hour. The absorbance was directly recorded at a wavelength of 450 nm in a microplate reader.
2.6. Quantification of IL-6 and MMP-2
IL-6 mRNA was evaluated by reverse transcription (RT)-PCR, and release of IL-6 into culture supernatants was analyzed using an enzyme-linked immunosorbent assay (ELISA) kit. In brief, total RNA was extracted and reverse transcribed using TaqMan Reverse Transcription Reagents. The gene specific primers were as follows: IL-6 (sense) 5’ –AGTTGTGCAATGGCAATTCTG-3’; IL-6 (antisense) 5’-
GGAAATTGGGGTAGGAAGGAC-3’; TM (sense) 5’- TACGGGAGACAACAACACCA-3’; TM (antisense) 5’- AAGTGGAACTCGCAGAGGAA-3’. -actin (sense) 5’- TGTTACCAACTGGGACGACA-3’; -actin (antisense) 5’-
GGGGTGTTGAAGGTCTCAAA-3’. Quantitative RT–PCR was performed and
analyzed as previously described [20]. ELISA was used to measure the concentrations of IL-6 (R&D Systems, Minneapolis, MN) in the supernatant of cultured aortic SMCs treated with TM siRNA or control siRNA based on the manufacturers’ instructions as previously described [21]. Matrix metalloproteinase-2 (MMP-2) activity in the condensed supernatants was examined by gelatin zymography. Protein (0.5 µg) was subjected to electrophoresis with 10% SDS-PAGE containing 0.1% gelatin. The gels were stained with Coomassie blue, and the band intensity was quantitatively
determined using Gel-Pro Analyzer software.
2.7. Migration assay
We evaluated the migration ability of aortic SMCs with and without TM
knockdown using a 48-well Boyden chamber assay. In the Boyden chamber (Neuro Probe, Bethesda, MD), 50 μL SMC suspension (5x104/mL) was added tothe upper chamber, and PDGF (20 ng/mL) was added tothe lower chamber. The chambers were incubatedat 37°C for 6 hours with 5% CO2. SMCs that did not migrate were scraped off the membrane, and the cells that migrated were fixedand visualized by Liu’s staining. The migrated cells were counted using an optical microscope (Leica) at 100X magnificationwith MetaMorph imaging software (Universal Imaging Corp).
Five random fields in each well were counted.
2.8. Vascular SMC-specific TM-deficient mice
Vascular SMC-specific TM-deletion mice were generated using the Cre-loxP system as previously described [20]. In brief, the whole TM gene was designed to flank with two loxP sites, and TMflox/+ chimera mice were generated from the
Transgenic Mouse Models Core, National Research Program for Genome Medicine, Taiwan (Supplemental figure 1A). The TMflox/+ mice were bred with wild type C57BL/6 mice for at least 5 generations. We used SM22-cre transgenic mice in which
cre recombinase was driven by the Tagln promoter and mainly expressed in vascular SMCs [22]. To generate vascular SMC-specific TM-deficient mice, TMflox/flox mice were crossbred to transgenic mice carrying a SM22-cre transgene (SM22-cretg, The Jackson Laboratory, stock number: 004746) to generate SM22-cretg/TMflox/+ mice, which were subsequently interbred to yield SM22-cretg/TMflox/flox mice and their wild- type controls, SM22-cretg/TM+/+ mice. All animal experiments were approved by the Institutional Animal Care and Use Committee, National Cheng Kung University (IACUC approval number:104215) and conformed to the Guide for the Care and Use of Laboratory Animals published by the National Institutes of Health (NIH
Publication #85-23, revised 1996).
2.9. Mouse carotid ligation model
Carotid artery ligation was performed according to our previous study [9]. In brief, the mice were anesthetized by intraperitoneal injection of pentobarbital.Neck incision was performed and the left common carotid artery was ligated completely with a 6-0 silk suture near the carotid bifurcation. At 2 and 4 weeks after ligation, the mice were sacrificed and the segment of left common carotid artery distal to the ligation site was excised for further analysis.
2.10. Neointima severity evaluation
Carotid artery was excised and 5 transverse sections (5 μm thick) of each carotid artery were cut at 100 μm intervals. After staining with hematoxylin-eosin, the
borders of the internal lumen, internal elastic lamina (IEL), and external elastic lamina (EEL) were traced on a digitizing board with Meta Imaging Series 5.0 (Adobe Inc).
The luminal, IEL, and EEL areas were measured, and the neointima area was calculated by subtracting the luminal area from the IEL area. The media area was calculated by subtracting the IEL area from the EEL area, and the ratio of neointima
to media area (N/M ratio) was calculated. The areas of the 5 sections were analyzed and averaged.
2.11. Staining of carotid artery
For immunofluorescence staining, carotid arteries isolated from the mice were embedded in optimum cutting temperature compound. Frozen sections (5 μm thick) were fixed and blocked with 5% goat serum. The sections were then stained with antibodies for TM (clone 1009, 1:150, Dako, Glostrup, Denmark), α-SMA (1:200), or Ki-67 (1:200, all from Abcam), followed by incubation with FITC-conjugated rabbit anti-mouse IgG (1:50, Millipore, Darmstadt, Germany) as the secondary antibody.
DAPI (Sigma) was used to identify the nuclei.
2.12. Statistical analysis
Data were presented as mean ± standard error. Comparisons between 2 groups were made using the Mann-Whitney test. For multiple comparisons of groups, one-way ANOVA was used, followed by Bonferroni post hoc analysis. All statistical analyses were performed using Prism 6 (GraphPad Software, San Diego, CA). A p value < 0.05 was considered to be statistically significant.
3. Results
3.1. Changes in aortic SMCs after TM knockdown
As expected, cultured human aortic SMCs expressed TM as shown by Western blot of the cell lysates and ELISA of the cultured media (Figure 1A). Most of the cultured aortic SMCs displayed a rhomboid-shaped morphology (Figure 1B). When the cells were transfected with TM siRNA, they redifferentiated back into a spindle-shaped morphology (Figure 1B). The expression of contractile phenotype marker, calponin, increased, while those of the synthetic phenotype markers, vimentin and NM-MHC, decreased after TM knockdown in the aortic SMCs (Figure 1C). In cells without TM
knockdown, PDGF-BB treatment decreased the expression of calponin and increased vimentin. In addition, an increased expression of calponin and decreased expression of vimentin were noted in the TM knockdown cells with or without PDGF-BB treatment compared to the cells without TM knockdown (Figure 1D).
3.2. Aortic SMC behaviors
Microarray analysis demonstrated that the most prominent effects of TM knockdown on the aortic SMCs were changes in gene expressions related to
proliferation (cell cycle) and inflammation (cytokine-cytokine receptor interaction) compared to the non-targeting siRNA controls (Figure 2A). Proliferation assays demonstrated that the aortic SMCs with TM knockdown had lower proliferation ability than the cells without TM knockdown (Figure 2B). We then evaluated the cytokine-producing ability of the aortic SMCs. Treatment with tumor necrosis factor (TNF)-α induced greater IL-6 mRNA expression and IL-6 protein production in the aortic SMCs, whereas the IL-6 producing ability was significantly reduced in the aortic SMCs with TM knockdown (Figure 2C). In contrast, the forced expression of TM in the aortic SMCs with TM knockdown increased IL-6 production after TNF-α stimulation (Figure 2D). These results suggested that membrane-bound TM might be responsible, at least in part, for TNF-α-mediated inflammation in aortic SMCs.
Since NF-κB transcription factor is the most important signal integration step in vascular inflammation, we further investigated the influence of TM knockdown on NF-κB activation in the aortic SMCs. Exposure of the human aortic SMCs to TNF-α significantly decreased the expression of NF-kB inhibitor, IκBα, and TM knockdown augmented the expression of IkBα (Figure 3A). Treating the aortic SMCs with TNF-α caused significant increases in p50 and p65 protein levels in the nuclear fraction of the aortic SMCs without TM knockdown, whereas TM knockdown decreased the levels
of p50 and p65 (Figure 3B) in the aortic SMCs.
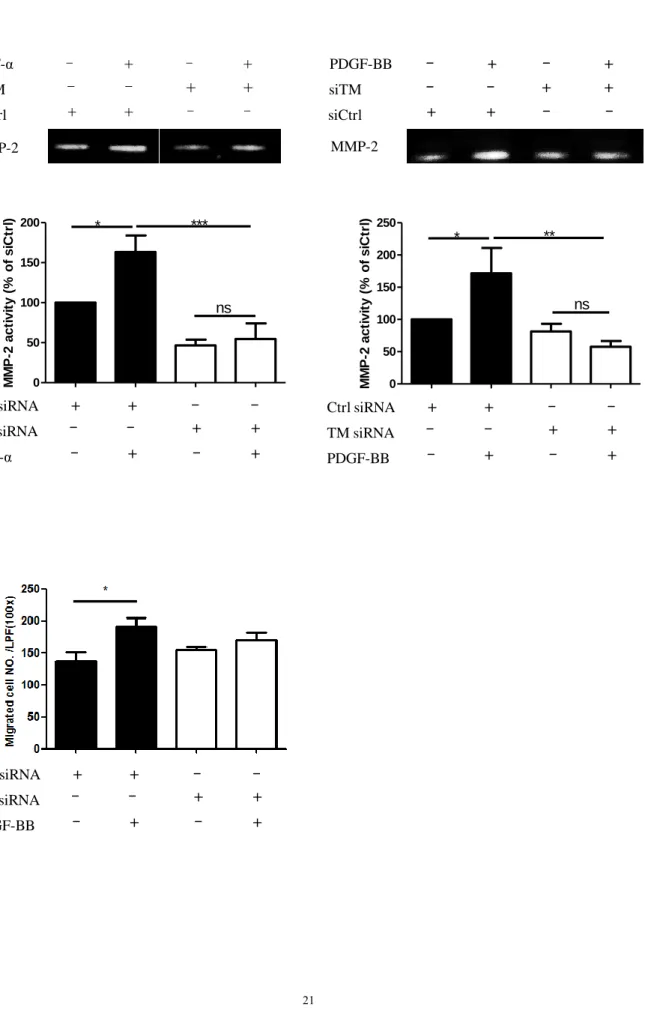
Although microarray analysis did not show significant changes in migration-related genes in the aortic SMCs with TM knockdown, we still examined the influence of TM expression on aortic SMCs migration. TNF-α or PDGF-BB stimulation increased MMP-2 activity in the aortic SMCs, and TM knockdown significantly decreased MMP-2 producing activity of the aortic SMCs (Figure 4A). The Boyden chamber assay showed that aortic SMC migration in response to PDGF-BB stimulation was slightly attenuated in cells with TM knockdown (Figure 4B).
3.3. Vascular SMC-specific TM-deletion mice
Transgenic mice with TM specifically deleted in vascular SMCs (SM22- cretg/TMflox/flox mice) and their wild-type controls (SM22-cretg/TM+/+ mice) were generated. Immunoblotting confirmed that TM was eliminated from vascular SMCs isolated from the SM22-cretg/TMflox/flox mice (Supplemental figure 1B). The blood pressure and body weight were similar between the SM22-cretg/TMflox/flox mice and their wild-type controls (Supplemental figure 1C and 1D). Carotid ligation caused neointima formation in the mice. Immunofluorescence staining showed high TM expression in the medial and neointimal cells at 4 weeks in the SM22-cretg/TM+/+
mice after carotid ligation, whereas no TM staining was found in the SM22-
cretg/TMflox/flox mice (Figure 5A). There was a progressive increase in the neointima area from 2 to 4 weeks after carotid ligation in both the SM22-cretg/TMflox/flox and SM22-cretg/TM+/+ mice (Figure 5B). The neointima area and neointima/media area ratio were significantly smaller in the SM22-cretg/TMflox/flox mice than in the SM22- cretg/TM+/+ mice (Figure 5B). Immunofluorescence staining showed fewer Ki67- positive cells in the media and neointima in the SM22-cretg/TMflox/flox mice compared to the SM22-cretg/TM+/+ mice (Figure 5C). The α-SMA positive staining intensity
was also stronger in the SM22-cretg/TMflox/flox mice (Figure 5D), suggesting that TM deficiency in the vascular SMCs in medial lesions resulted in a more contractile status after carotid ligation.
4. Discussion
In this study, we found that aortic SMC membrane-bound TM played a role in SMC phenotype and changes in behavior. TM affects cell behavior differently in different cell types. In epidermal epithelial A431 cells, cells expressing TM were shown to maintain compact cell colonies with an epithelial morphology, whereas TM knockdown induced the formation of cell protrusions at the edges of the colonies and increased cell migration [23]. TM expressed on monocyte works as an adhesion molecule and mediates monocytes adhesion to vascular endothelium [24]. With regards to SMCs, Tohda et al. demonstrated that exogenous recombinant TM protein treatment significantly increased the proliferation of rat vascular SMCs [10].
However, the effects of endogenous expression of TM on vascular SMCs have rarely been studied. Several studies have demonstrated that cultured vascular SMCs express TM, and this was associated with the down-regulation of contractile phenotype markers such as α-actin and tropomyosin [25, 26]. In addition, Ramachandran et al.
found that knockdown of the endogenous expression of TM in cultured human urinary bladder SMCs, a visceral SMC, did not affect bladder SMC growth but significantly attenuated PDGF-induced bladder SMC migration [27]. In the current study, we demonstrated that the TM expression on aortic SMCs was mainly associated with proliferative and proinflammatory phenotype transition, and also mildly increased aortic SMC migration. SMCs have extensive functional diversity depending on the specific demand within a given organ. Phasic and rhythmic contraction is the characteristic of visceral SMCs, while vascular SMCs in arteries usually maintain
continuous contraction to preserve vascular tone [28]. The mechanisms underlying the different effects of TM expression on different cell types, including vascular and visceral SMCs, are unclear and need further exploration.
Our in vivo experiments showed that neointima formation was more severe in the wild-type mice with TM expression compared to vascular SMC-specific TM-deficient mice. Previous studies have shown that systemic injections of exogenous recombinant TM protein or local incubation of recombinant adenoviral constructs overexpressing TM within injured arterial lumen decreased vascular injury-induced neointima formation [9, 29, 30]. The anti-thrombotic and anti-inflammatory effects of
recombinant TM protein on the endothelium are the major mechanisms underlying the anti-atherosclerosis effect of TM. Recombinant TM has been shown to decrease thrombin-induced PAR-1 activation and inflammation on the endothelium after vascular injury [31]. Recent studies have further demonstrated that recombinant TM protein has a thrombin-independent effect on the endothelium and can directly decrease the stress-induced apoptosis of endothelial cells [32, 33] and reduce leukocyte transmigration to the endothelium [34]. The accumulation of vascular SMCs in the subintima is a major feature of atherosclerosis and neointima. Co-
culturing SMCs with endothelial cells has been shown to cause a shift in SMCs to the synthetic phenotype and induced the expressions of inflammatory genes in endothelial cells [35]. Although recombinant TM has beneficial effects on the endothelium, the potential impact of vascular SMC-bound TM on endothelial cells is unknown.
Monocyte-expressed TM enhances adhesion of monocytes to endothelium and myeloid-specific TM-deficient mice had less neointima formation after carotid ligation [24]. The interplay between TM expression on monocytes, vascular SMCs and endothelial cells is unclear. The effects of TM on individual vascular cell types
are different and further investigations are needed to elucidate its role in cross talk between different cell types during atherosclerotic process. Another limitation of our study is the specificity of SM22 as a SMC-specific deletion Cre. Although commonly used to study smooth muscle specific expression, it is not the most specific marker and could express in non-muscular cells [36]. This could influence the experimental results of our study.
In conclusion, although previous studies have documented the expression of TM on neointimal SMCs and hypothesized its effect in protecting injured arteries from thrombosis, the present study showed that vascular SMC-bound TM had an important physiological effect on vascular SMC biology beyond its traditional anti-thrombotic effect.
Conflict of Interest None
Financial Support
This work was supported by grants 104-2314-B-006-083-MY2 and 106-2314-B-006- 045-MY3 from the Ministry of Science and Technology, Taipei, Taiwan.
Author contributions
Wang KC, Chung HC, Tseng SY, Huang TY, and Lin YL performed the experiments and collected the data. Wang KC, Chen PS, Chao TH, and Li YH designed the study and analyzed the data. Li YH and Wang KC drafted the manuscript. Luo CY, Shi GY, and Wu HL revised the manuscript for important intellectual content.
References
1. Libby P, Ridker PM, Maseri A. Inflammation and atherosclerosis. Circulation 2002;105:1135–1143.
2. Hansson GK, Hermansson A. The immune system in atherosclerosis. Nat Immunol 2011;12:204–212.
3. Orr AW, Hastings NE, Blackman BR, et al. Complex regulation and function of the inflammatory smooth muscle cell phenotype in atherosclerosis. J Vasc Res 2010;47:168–180.
4. Tellides G, Pober JS. Inflammatory and immune responses in the arterial media.
Circ Res 2015;116:312-322.
5. Hartman J, Frishman WH. Inflammation and atherosclerosis: a review of the role of interleukin-6 in the development of atherosclerosis and the potential for targeted drug therapy. Cardiol Rev 2014;22:147-151.
6. Gomez D, Owens GK. Smooth muscle cell phenotypic switching in atherosclerosis. Cardiovasc Res 2012;95:156–164.
7. Esmon CT. The roles of protein C and thrombomodulin in the regulation of blood coagulation. J Biol Chem 1989;264 4743-4746.
8. Li YH, Kuo CH, Shi GY, et al. The role of thrombomodulin lectin-like domain in inflammation. J Biomed Sci 2012;19:34.
9. Li YH, Liu SL, Shi GY, et al. Thrombomodulin plays an important role in arterial remodeling and neointima formation in mouse carotid ligation model. Thromb Haemost 2006;95:128-133.
10. Tohda G, Oida K, Okada Y, et al. Expression of thrombomodulin in
atherosclerotic lesions and mitogenic activity of recombinant thrombomodulin in
vascular smooth muscle cells. Arterioscler Thromb Vasc Biol 1998 18 1861- 1869.
11. Yoshii Y, Okada Y, Sasaki S, et al. Expression of thrombomodulin in human aortic smooth muscle cells with special reference to atherosclerotic lesion types and age differences. Med Electron Microsc 2003;36:165-172.
12. Ma SF, Garcia JG, Reuning U, et al. Thrombin induces thrombomodulin mRNA expression via the proteolytically activated thrombin receptor in cultured bovine smooth muscle cells. J Lab Clin Med 1997;129:611-619.
13. Rabausch K, Bretschneider E, Sarbia M, et al. Regulation of thrombomodulin expression in human vascular smooth muscle cells by COX-2-derived
prostaglandins. Circ Res 2005;96:e1-6.
14. Lo IC, Lin TM, Chou LH, et al. Ets-1 mediates platelet-derived growth factor- BB-induced thrombomodulin expression in human vascular smooth muscle cells.
Cardiovasc Res 2009;81:771-779.
15. Laszik ZG, Zhou XJ, Ferrell GL, et al. Down-regulation of endothelial expression of endothelial cell protein C receptor and thrombomodulin in coronary atherosclerosis. Am J Pathol 2001;159:797-802.
16. Huang HC, Shi GY, Jiang SJ, et al. Thrombomodulin-mediated cell adhesion:
involvement of its lectin-like domain. J Biol Chem 2003;278:46750-46759.
17. Ikezoe T, Yang J, Nishioka C, et al. Thrombomodulin protects endothelial cells from a calcineurin inhibitor-induced cytotoxicity by upregulation of extracellular signal-regulated kinase/myeloid leukemia cell-1 signaling. Arterioscler Thromb Vasc Biol 2012;32:2259-2270.
18. Chen PS, Wang KC, Chao TH, et al. Recombinant thrombomodulin exerts anti- autophagic action in endothelial cells and provides anti-atherosclerosis effect in apolipoprotein E deficient mice. Sci Rep 2017;7:3284.
19. Isermann B, Hendrickson SB, Zogg M, et al. Endothelium-specific loss of
murine thrombomodulin disrupts the protein C anticoagulant pathway and causes juvenile-onset thrombosis. J Clin Invest 2001;108:537-546.
20. Wang KC, Li YH, Shi GY, et al. Membrane-bound thrombomodulin regulates macrophage inflammation in abdominal aortic aneurysm. Arterioscler Thromb Vasc Biol 2015;35:2412-2422.
21. Lai CH, Wang KC, Kou CH, et al. Recombinant adeno-associated virus vector carrying the thrombomodulin lectin-like domain for the treatment of abdominal aortic aneurysm. Atherosclerosis 2017;262:62-70.
22. Boucher P, Gotthardt M, Li WP, et al. LRP: role in vascular wall integrity and protection from atherosclerosis. Science 2003;300:329-332.
23. Hsu YY, Shi GY, Kuo CH, et al. Thrombomodulin is an ezrin-interacting protein that controls epithelial morphology and promotes collective cell migration.
FASEB J 2012;26:3440-3352.
24. Lin WL, Chen CC, Shi GY, et al. Monocytic thrombomodulin promotes cell adhesion through interacting with its ligand, Lewisy. Immunol Cell Biol 2017;95:372-379.
25. Soff GA, Jackman RW, Rosenberg RD. Expression of thrombomodulin by smooth muscle cells in culture: different effects of tumor necrosis factor and cyclic adenosine monophosphate on thrombomodulin expression by endothelial cells and smooth muscle cells in culture. Blood 1991;77:515-518.
26. Chamley-Campbell J, Campbell GR, Ross R. The smooth muscle cell in culture.
Physiol Rev 1979;59:1-61.
27. Ramachandran A, Ranpura SA, Gong EM, et al. An Akt- and Fra-1-dependent pathway mediates platelet-derived growth factor-induced expression of
thrombomodulin, a novel regulator of smooth muscle cell migration. Am J Pathol 2010;177:119-131.
28. Fisher SA. Vascular smooth muscle phenotypic diversity and function. Physiol Genomic. 2010;42A:169-187.
29. Li JM, Singh MJ, Itani M, et al. Recombinant human thrombomodulin inhibits arterial neointimal hyperplasia after balloon injury. J Vasc Surg 2004;39:1074- 1083.
30. Waugh JM, Li-Hawkins J, Yuksel E, et al. Thrombomodulin overexpression to limit neointima formation. Circulation 2000;102:332-337.
31. Wei HJ, Li YH, Shi GY, et al. Thrombomodulin domains attenuate
atherosclerosis by inhibiting thrombin-induced endothelial cell activation.
Cardiovasc Res 2011; 92: 317-327.
32. Ikezoe T, Yang J, Nishioka C, et al. Thrombomodulin protects endothelial cells from a calcineurin inhibitor-induced cytotoxicity by upregulation of extracellular signal-regulated kinase/myeloid leukemia cell-1 signaling. Arterioscler Thromb Vasc Biol. 2012;32:2259-2270.
33. Chao TH, Tsai WC, Chen JY, et al. Soluble thrombomodulin is a paracrine anti- apoptotic factor for vascular endothelial protection. Int J Cardiol 2014;172:340- 349.
34. Lin WL, Chang CF, Shi CS, et al. Recombinant lectin-like domain of thrombomodulin suppresses vascular inflammation by reducing leukocyte
recruitment via interacting with Lewis Y on endothelial cells. Arterioscler Thromb Vasc Biol 2013;33:2366-2373.
35. Chiu JJ, Chen LJ, Chang SF, et al. Shear stress inhibits smooth muscle cell- induced inflammatory gene expression in endothelial cells: role of NF-kappaB.
Arterioscler Thromb Vasc Biol 2005;25:963-969.
36. Chakraborty R, Saddouk FZ, Carrao AC, et al. Promoters to study vascular smooth muscle. Arterioscler Thromb Vasc Biol 2019;39:603-612.
A.
B.
Figure 1
siCtrl siTM TM
α-SMA Calponin Vimentin
NM-MHC GAPDH
C.
TM
Vimentin Calponin
GAPDH siCtrl siTM PDGF-BB
+ +
+
+ +
+ –
–
–
– –
–
D.
siCtrl siTM
Normal
24 48 72 Time (hr)
TM β-actin
siCtrl siTM
24 48 72
0 500 1000 1500 2000 2500
hr
TM ELISA conc. (pg/mL)
A.
Figure 2
B.
D.
C.
siCtrl siTM
0 1 2 3 4 Days
0 1 2
Ctrl siRNA TM siRNA
TNF- - + - + + + - - - - + +
** *** ###
ns
IL-6 ELISA conc.(ng/ml)
TNF-α TM siRNA Ctrl siRNA
– – + +
– + – +
+ + – –
0 1 2
Ctrl siRNA TM siRNA
TNF- - + - + + + - - - - + +
** * ###
ns
Relative expression of IL-6
TNF-α TM siRNA Ctrl siRNA
– – + +
– + – +
+ + – –
1 2 3
4
***
A conc.(ng/g)
1.0 1.5 2.0
2.5
*
M protein levels
A.
Figure 3
B.
siCtrl
siCtrl TNF-α siTM
TNF-α siTM
TNF-α TM siRNA Ctrl siRNA
– – + +
– + – +
+ + – –
TNF-α TM siRNA Ctrl siRNA
– – + +
– + – +
+ + – –
0.0 0.5 1.0 1.5
Ctrl siRNA TM siRNA
TNF- - + - + + + - - - - + +
***
n.s. *** *
Relative protein levels of
I B
P50 P65 Lamin A/C TM IκBα α-Tubulin
0 .0 0 .5 1 .0 1 .5 2 .0
Relative protein levels of p50
*
n .s . n .s .
*
0 1 2 3 4
Relative protein levels of p65
***
**
**
n .s .
Figure 4 A.
siCtrl siTM TNF-α
– – + +
+ + – –
– + – +
MMP-2 MMP-2
siCtrl siTM PDGF-BB
– – + +
+ + – –
– + – +
B.
0 50 100 150 200
Ctrl siRNA TM siRNA
TNF- - + - + + + - - - - + +
*
ns
***
M M P -2 a c ti v it y ( % o f s iCt rl )
TNF-α TM siRNA Ctrl siRNA
– – + +
– + – +
+ + – –
0 50 100 150 200 250
Ctrl siRNA TM siRNA
PDGF-BB - + - + + + - - - - + +
* **
ns
M M P -2 a c ti v it y ( % o f s iCt rl )
PDGF-BB TM siRNA Ctrl siRNA
– – + +
– + – +
+ + – –
PDGF-BB TM siRNA Ctrl siRNA
– – + +
– + – +
+ + – –
Figure 5
A. DAPI α-SMA TM Merge SM22-cre
tg/TM
+/+SM22-cre
tg/TM
flox/floxSM22-cretg/TM+/+
★★★
N/M ratio 2 Weeks (N=6) 4 Weeks (N=9)
★★★
SM22-cretg/TM+/+ SM22-cretg/TMflox/flox
N /M r ati o
B.
U n li gate d
SM22-cre
tg/TM
flox/flox2 We e k s 4 We ek s
SM22-cre
tg/TM
+/+4 We ek s
L L
L
L L
M L
M
M M
A A
A
A
N
N
N N
M M
A A
4 Weeks SM22-cretg/TM+/+
SM22-cretg/TMflox/flox
★★★
A r e a (μ m
2)
C.
D.
SM22-cre
tg/TM
+/+SM22-cre
tg/TM
flox/flox2 week
Unligated
α- S M A
★
SM22-cretg/TMflox/flox 2 weeks
SM22-cretg/TMflox/flox SM22-cretg/TM+/+
Ki67+positive cell / Total neointimacell (%)
4 weeks
★★
★★
★★
★★★
α-SMAstaining intensity
Medial cells SM22-cretg/TM+/+
SM22-cretg/TMflox/flox
★★
★
Cell number
SM22-cretg/TM+/+SM22-cretg/TM+/+SM22-cretg/TMflox/flox
2 w e e k s 4 w e e k s
Ki67+positive cell / Total neointimacell (%)
SM22-cretg/TM+/+
DAPI α-SMA Ki67 Merge
SM22-cretg/TMflox/flox
M M
M
M M M
M M
1 M
M M
N N N
N N N
N N
M N
Supplemental Figure 1
A.
C.
Mouse Blood Pressure
1 2 3
60 90
120
SM22CreTM
wt/wt(n=5)
SM22CreTM
flox/flox(n=6)
Day
Bl o o d p re s s u re ( m m Hg )
SM22-cretg/TM+/+
B lo o d p r e ss u r e (m m H g)
1 2 3 Day
SM22-cretg/TMflox/flox
B.
TM GAPDH
D.
0 5 10 15 20 25 30
Mouse Body Weight ns
(n=10) (n=11)
SM22CreTM
wt/wt
SM22CreTM
flox/flox
Weight(g)
We igh t (g)
ns
Figure legends
Figure 1. Phenotype changes of aortic SMCs after TM knockdown. (A) Cultured aortic SMCs expressed TM as shown by Western blot (left panel) of the cell lysates and ELISA of the cultured media (right panel). (B) Cultured aortic SMCs treated with TM-targeting siRNA (siTM) underwent morphological changes from a rhomboid (arrows) to spindle shape (arrowheads) compared with non-targeting siRNA controls (siCtrl). Scale bars, 20 μm. (C) Representative Western blot illustrating expression levels of TM, contractile and synthetic markers in aortic SMCs with (siTM) and without (siCtrl) TM knockdown. GAPDH was used as a loading control.
Quantification of protein expression (n=6) was normalized to GAPDH and expressed as mean ± SEM. *p < 0.05; **p < 0.01 and ***p < 0.001. α-SMA, α-smooth muscle actin; NM-MHC, non-muscle myosin heavy chain. (D) Representative Western blot illustrating the expressions of calponin and vimentin in aortic SMCs with the indicated treatment. GAPDH was used as a loading control.
Figure 2. Functional changes of aortic SMCs after TM knockdown. (A) Microarray results showed significant changes in cell cycle and cytokine-cytokine receptor interaction pathway in aortic SMCs with TM knockdown (siTM). *p<0.05 ,
***p<0.001 compared with controls (siCtrl). (B) Cell proliferation was measured by WST-1 cell proliferation assays. The number of cells was expressed in units of optical density (OD450). All experiments were performed at least 3 times. ***p< 0.001. (C) IL-6 mRNA expression levels in aortic SMCs and IL-6 concentrations in conditioned media were measured. Data were expressed as mean ± SEM. *p<0.05, **p<0.01,
***p<0.001, ###p<0.001, n.s. indicates not significant (n=3). (D) Expression and quantification of TM protein in pEGFPN1 vector and pEGFPN1-TM transfected
aortic SMCs with TM knockdown (n=3). IL-6 concentrations in the conditioned media from TNF-α-treated pEGFPN1 vector and TNF-α-treated pEGFPN1-TM transfected cells were determined by ELISA. Data were expressed as mean ± SEM.
*p<0.05, ***p<0.001. GAPDH was used as a loading control. Results were normalized to GAPDH and expressed as mean ± SEM.
Figure 3. TM knockdown attenuated TNF-α-induced NFκB activation in aortic SMCs. (A) Cell lysates with the indicated treatment were analyzed for TM and IκBα expressions. α-tubulin was used as a loading control. Quantification of IκBα protein expression (n=3) was shown as the bar graph. *p<0.05, **p<0.01, ***p<0.001. n.s.
indicates not significant. (B) Western blot analysis of p50 and p65 protein
expressions. Equality of nuclear sample loading was confirmed with control lamin A/C, a nuclear protein marker. Quantification of p50 and p65 protein expressions (n=3) were shown as the bar graph. Data were expressed as mean ± SEM. *p<0.05,
**p<0.01, ***p<0.001. n.s. indicates not significant.
Figure 4. Aortic SMC migration after TM knockdown. (A) Aortic SMCs received the indicated treatment and MMP-2 activity was analyzed by zymography. Data were expressed as mean ± SEM. *p<0.05, **p<0.01, ***p<0.001, n.s. indicates not significant (n=3). (B) Migration of aortic SMCs with the indicated treatment was evaluated by Boyden chamber assay. The cells were allowed to migrate for 6 hours in response to medium with or without PDGF (20 ng/mL) in the lower chamber. The number of migrated cells was counted in 5 random fields for each well. Data were shown as mean ± SEM; n=3. *p<0.05.
Figure 5. Neointima formation in mice after carotid artery ligation. (A) Double immunofluorescent staining for α-SMA and TM in the ligated carotid arteries. Nuclei were counterstained with DAPI. TM expression was detected in the media and neointima in SM22-cretg/TM+/+ mice, but only minimal TM staining was observed in the endothelium of the ligated carotid arteries of SM22-cretg/TMflox/flox mice. Scale bar, 100 μm. (B) Hematoxylin–eosin staining of carotid arteries after carotid ligation.
The dashed lines indicated borders of neointima/media and media/adventitia. A, adventitia; L, lumen; M, media. Scale bar, 100 μm. Quantification of the neointima was calculated and shown as the bar graph. M, media; N, neointima. ***p<0.001. (C) Double immunofluorescent staining for α-SMA and Ki67 (arrows) in the carotid arteries after ligation. The dashed lines indicated borders of the neointima and media.
Scale bar, 50 μm. M, media; N, neointima. The ratio of Ki positive cells to total cells and the absolute number of Ki67 positive cells were calculated and shown as the bar graphs. *p<0.05; **p<0.01; ***p<0.001. (D) Immunohistochemical staining of α- SMA in the carotid arteries at 2 weeks after carotid ligation. Scale bar, 100 μm. The α- SMA positive staining intensity was quantitated and shown as the bar graph.
**p<0.01. ***p<0.001.
Supplemental Figure 1. (A) Schematic description of Cre recombination leading to TM gene inactivation in cells expressing SM22. (B) Western blotting of TM protein in SMCs isolated from SM22-cretg/TM+/+ and SM22-cretg/TMflox/flox mice. (C) The blood pressure and (D) body weight were similar between the SM22-cretg/TM+/+ and SM22- cretg/TMflox/flox mice.
106年度專題研究計畫成果彙整表
計畫主持人:李貽恆 計畫編號:106-2314-B-006-045-MY3 計畫名稱:利用基因轉殖小鼠研究凝血調節素在血管新生內膜及動脈硬化中的角色
成果項目 量化 單位
質化
(說明:各成果項目請附佐證資料或細 項說明,如期刊名稱、年份、卷期、起 訖頁數、證號...等)
國
內 學術性論文
期刊論文 0
研討會論文 0 篇
專書 0 本
專書論文 0 章
技術報告 0 篇
其他 0 篇
國
外 學術性論文
期刊論文 1
篇
Atherosclerosis 2019;287: 54–63
研討會論文 2
1. Wang KC, Li YH, Chen PS, Chung HC, Wu HL. Thrombomodulin expressed on vascular smooth muscle cell influences the behaviors of smooth muscle cell. 2018 Annual Scientific Session of the American Heart
Association, Chicago, Illinois, USA, November 10-12, 2018.
2. Li YH, Wang KC, Chen PS, Chung HC, Wu HL. Thrombomodulin expressed on vascular smooth muscle cell influences arterial injury-induced neointima formation in mouse. 2019 Annual Scientific Session of the European Society of Cardiology, Paris, France, August 31 – September 4, 2019.
專書 0 本
專書論文 0 章
技術報告 0 篇
其他 0 篇
參 與 計 畫 人 力
本國籍
大專生 0
人次
碩士生 1 馮文翰
博士生 1 陳柏升
博士級研究人員 0
專任人員 1 鍾幸君
非本國籍
大專生 0
碩士生 0
博士級研究人員 0
專任人員 0
其他成果
(無法以量化表達之成果如辦理學術活動
、獲得獎項、重要國際合作、研究成果國 際影響力及其他協助產業技術發展之具體 效益事項等,請以文字敘述填列。)