行政院國家科學委員會專題研究計畫 成果報告
戴奧辛引發人類滋養層細胞缺氧及細胞功能缺損之分子機 制
計畫類別: 個別型計畫
計畫編號: NSC94-2320-B-038-028-
執行期間: 94 年 08 月 01 日至 95 年 07 月 31 日 執行單位: 臺北醫學大學生物醫學技術研究所
計畫主持人: 高淑慧 共同主持人: 曾啟瑞 計畫參與人員: 趙湘台
報告類型: 精簡報告
處理方式: 本計畫可公開查詢
中 華 民 國 95 年 10 月 31 日
行政院國家科學委員會補助專題研究計畫
█ 成 果 報 告□期中進度報告
戴奧辛引發人類滋養層細胞缺氧及細胞功能缺損之分子機制
計畫類別:▉ 個別型計畫 □ 整合型計畫 計畫編號:NSC 94-2320-B-038-028
執行期間: 94 年 08 月 01 日至 95 年 07 月 31 日
計畫主持人:高 淑 慧 共同主持人:曾 啟 瑞 共同主持人:
計畫參與人員:趙 湘 台
成果報告類型(依經費核定清單規定繳交):▉精簡報告 □完整報告
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計畫、列管計畫及下 列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,□一年□二年後可公開查詢
執行單位:臺北醫學大學醫學檢驗暨生物技術系研究所
中 華 民 國 95 年 10 月 31 日
中文摘要
戴奧辛(dioxin)為環境污染物因具高毒性及廣泛性,已經被許多國家列為人類確定致癌 物。由戴奧辛所引發的人類生殖系統的相關疾病有日漸增加的趨勢,如子宮內膜異位症及著 床前後的受精卵流失被認為與戴奧辛的傷害有關。戴奧辛其結構和荷爾蒙相似,可嚴重影響 人體的新陳代謝﹐干擾荷爾蒙的平衡等稱為荷爾蒙干擾物(hormone disruptor)。在懷孕婦女,
常發現戴奧辛會增加胚胎流產及胎兒過小的機率。本計畫以人類胚胎滋養層細胞為細胞模 式,研究 TCDD 對胎盤細胞調控的影響及其可能對胚胎所引發的傷害。研究結果顯示,經 TCDD 處理人類胚胎滋養層細胞會引發細胞產生缺氧(hypoxia),同時細胞內有大量 reactive oxygen species (ROS)產生及胚胎滋養層細胞凋亡。細胞中的氧化性傷害(oxidative damages, 如 DNA 氧化損傷的分子標記-8-羥基去氧鳥糞嘌呤(8-hydroxy-2’-deoxyguanosine) 及脂質過氧化 物(lipid peroxide))隨 TCDD 的劑量增加,分別呈 6.5 倍與 2.58 倍的增加。此外,在戴奧辛 48 小時刺激下,人類胚胎滋養層細胞粒線體 DNA 7599bp 斷損突變(deletion)增加及粒線體 DNA 拷貝數(copy number)的減少為 52%。我們進一步分析基因表現的變化,戴奧辛刺激可誘發人 類滋養層細胞產生缺氧誘發因子-1(hypoxia inducible factor-1,HIF-1α)大量表現。綜合上述結 果,我們初步證實 TCDD 可經由引發細胞缺氧及氧化性傷害並造成粒線體功能缺損而促使人 類胚胎滋養層細胞凋亡。當我們以 Wortamnin, Ly294002 抑制劑此來研究 PI 3-K/PDK-1/ Akt 訊息傳遞路徑對 HIF1-基因之表現關聯性,或以 PD98059 及環境荷爾蒙來分析 MEK signaling pathway 是否參與 HIF1-基因表現之調控,我們發現 TCDD 處理人類胚胎滋養層細胞會引發 細胞產生缺氧誘發因子(HIF-1)是經由 PI 3-K/PDK-1/ Akt 訊息傳遞路徑。當以抗氧化劑 L-N-acetylcysteine 發現可以減緩由戴奧辛誘發細胞凋亡及 HIF1-的蛋白表現。綜合以上實驗 結果,我們推論由戴奧辛引發的缺氧及 ROS 大量產生可能是造成人類胚胎滋養層細胞凋亡的 主要因素。
關鍵詞:TCDD、胚胎滋養層細胞、缺氧因子、氧化性自由基、細胞凋亡
Abstract
In the recent years, considerable attention has been focus on understanding the potential adverse reproductive effects of 2, 3, 7, 8-tetrachlorodibenzo-p-dioxin (TCDD) in humans. Some studies have demonstrated that TCDD has the ability to cause endometriosis and pre- and/or post-implantation losses of fertilized eggs. Exposure to TCDD during pregnancy causes fetal loss in
many animal species. Exactly how TCDD may affect the reproductive capacity of humans is not well understood. In this study, we plan to distinguish the actions of TCDD on fetal death and placenta development. We established the human trophoblastic cells and plan to identify the molecular mechanisms of TCDD-induced cellular damages. In preliminary data, we found the increased reactive oxygen species (ROS) and hypoxia in the TCDD-treated trophoblast cells. Our central hypothesis proposes that ROS and hypoxia might be anticipated in the TCDD-induced damages. We set several experiments to explore the hypoxia effects and ROS related responses including ROS generation, oxidative damages including lipoperoxides and oxidized DNA bases (8-OH-dG), mitochondrial DNA mutations, cell viability, and cell invasion assay. We found the increased ROS generation and hypoxia inducing factor-1α (HIF-1α) in the TCDD-treated trophoblast cells as the dose-response and time-dependent manners. We also examined the 6.5 folded-increase of 8-OH-dG and 2.58-folded increase of lipoperoxides in the TCDD-treated cells by time- and dose-response manners. Furthermore, the content of mitochondrial DNA copy numbers was reduced to 52 % vs control. The 7599 bp mtDNA deletions were found in the TCDD treated cells. The increased apoptotic fraction was found in the TCDD-manipulated cells. Moreover, there were approximately two-folded invasion was detected in the TCDD-resistant cells. Hypoxia is identified in the cause of placenta hypoplasia, tissue hypoxia and reducing placental blood supply.
Moreover, we plan to explore the molecular mechanisms and signal transduction pathway(s) of the TCDD-induced hypoxia and cell apoptosis. By using the inhibitors Wotmannin, Ly294002 (PI-3K/PDK-1/Akt pathway), and PD98059 (MEK pathway), the PI-3K/PDK-1/Akt signaling pathway was identified to be involved in the TCDD induced cytotoxicity. Additionally, the reduced cellular damages were found in the supplementation of ROS scavengers, L-N-acetylcysteine.
According to our findings, the enlarged ROS generations, HIF-1αaccumulation, increased cell damages, and reduced viability might be contributed in TCDD-induced fetal death.
Key words: TCDD, trophoblastic cells, HIF-1,reactive oxygen species, apoptosis
研究計畫之背景及目的
近年來於自然界環境中發現具有干擾內分泌調節系統的物質,這些物質被稱為”endocrine disruptors”。這些物質可以干擾或破壞荷爾蒙的合成、分泌、運送、鍵結能力及作用機制 (1)。
內分泌系統與體內發育、生長及生殖功能等重要調控息息相關。而日益增加的內分泌調節異 常所引發的疾病如子宮內膜異位症(endometriosis)、乳癌、睪丸及前列腺癌及精子生成等皆被 認為與 endocrine disruptors 有關 (1,2)。這些物質中以戴奧辛(dioxin)的毒性被廣泛研究。戴奧 辛約有 17 種不同的化合物,其中以 2,3,7,8-四氯戴奧辛(2,3,7,8-tetrachloro- dibenzo-p- dioxin, TCDD)其毒性最強,且其作用機制較被廣泛探討(3)。
目前已知戴奧辛對人體健康的影響包括:增加癌症可能性、孕婦接觸戴奧辛可能流產、
體重過輕的胎兒、產下畸形兒或學習障礙兒、雄性荷爾蒙降低、糖尿病以及影響免疫系統的 功能。此外,並且造成著床前及著床後的的胚胎死亡,被推測干擾 placenta-uterine 的功能 (1,4,5)。其所造成胎兒發育的傷害被證實與imprinted gene的甲基化有關(6)。文獻也指出TCDD 易引發胎兒流產、畸形及死亡。並且於體外試驗中,發現TCDD會降低 bioavailable HCG的生 成進而造成脅迫性流產(threaten abortion) (7)。
本研究計畫以人類胚胎滋養層細胞(trophoblast, 3-A-sub E cell lines)進行體外細胞培養。人 類胚胎滋養層細胞為懷孕初期胎盤構造的必需組成細胞,藉由滋養層的活性,促使囊胚 (blastocyst)約在第11天可完全深植於子宮內膜組織中。並影響胎兒及母體的循環系統。本計畫 研究結果顯示,經TCDD處理人類胚胎滋養層細胞會引發細胞產生缺氧(hypoxia),同時細胞內 有大量 reactive oxygen species (ROS)產生。文獻指出,缺氧及氧化自由基兩者之間有相互的 影響(8-11)。此外文獻也指出人類胚胎滋養層細胞若長期處於缺氧下會促使胎盤發育不全、組 織缺氧、常伴隨胎兒-母體血液供應不足,並影響胚胎細胞生長,甚或造成細胞凋亡(12,13)。
我們發現隨TCDD的劑量增加或處理時間的增加,細胞中的氧化性傷害及細胞凋亡有漸增的 趨勢,即呈現dose-responsive及time-dependent的影響。並且成粒線體功能缺損而促使人類胚 胎滋養層細胞凋亡。本研究計畫擬進一步探討戴奧辛引發細胞缺氧及氧化性傷害其分子訊息 機制。
實驗材料與方法
人類胚胎滋養層細胞(human placenta trophoblast cell) 培養
人類胚胎滋養層細胞(human placenta trophoblast cell) 需培養於含有 10% 胚胎牛血 清(fetal bovine serum , FBS)、100unit/ml penicillin、0.1mg/ml streptomycine、0.25ug/ml
amphotericin B 之 DMEM 培養液,將此細胞培養於 10 cm 培養皿中,置於 5% CO2、37℃
恆溫之培養箱中。
細胞蛋白質製備(Preparation of cell lysate)
Lysis buffer:50mM Tris-HCl (pH 7.4), 150mM NaCl, 1% PMSF, 4% protease inhibitor cocktail 及 0.1% NP-40。將培養於 10cm 培養皿中的細胞分別給與藥物刺激一段時間之 後,吸去培養液,再以冰的 PBS 反覆清洗三次,以減緩細胞本身的生理作用,最後加入 200μl 的 lysis buffer 於培養皿中,將細胞刮下移至 1.5ml 離心管中,置於冰上 30 分鐘待 細胞膜溶解後,以 12000 rpm、4℃,離心 30 分鐘,收集上清液為蛋白質萃取液,經蛋 白定量後取出等量蛋白進行西方墨點法測定蛋白質表現的變化。
蛋白質定量法(Protein assay)
Bio-Rad protein assay dye (Bio-Rad Laboories,Hercules,CA)先以去離子水以 1:4 之比例稀釋五倍後,用 Whatman No.1 濾紙過濾於試管中,分別加入 1ml 過濾液。並以 0.2mg/ml、0.4mg/ml 及 0.8mg/mll 之 Bovine serum albumin ( BSA )作為標準液,用以製定
標準曲線。需取 2μl的待測物加入裝有過濾液之試管中,經震盪後,待測物之顏色深淺
須介於 standard 0.2~0.8 mg/ml BSA 標準液間,並測定波長 595 nm 之吸光值,測得及計 算出蛋白質含量。
西方墨點法(western blotting) 1. SDS-PAGE gel preparation
Gel 分為兩層,上層為 stacking gel,配置方法為 3.94ml ddH2O 中,分別加入 0.5ml 40%的 Acylamide、0.5ml 3M 的 Tris-HCl (pH8.9)、50μl的 10% SDS(sodium dodecyl sulfate)、50μl的 10% APS 及 10μl的 TEMED 即完成。下層為 10%separating gel,配置方 法為在 12.2ml ddH2O 中,分別加入 5ml 的 40% Acylamide、2.5ml 的 3M Tris-HCl (pH8.9)、
200μl的 10% SDS、100μl的 10% APS 及 20μl的 TEMED 即完成。
2. 電泳
將電泳膠片置於電泳槽中,並填滿電泳緩衝液(running buffer),並取以定量的待測 蛋白質萃取液(50μg),加入 3 倍的 SDS protein loading buffer 混合,並於 95℃下處理 5 分鐘後,隨及置於冰上冷卻,再將處理好的蛋白質萃取液加到每個 well 中,使用 90 voltage 電壓跑 10 % SDS-PAGE gel。約 3 小時後,停止電泳並進行蛋白質的轉印。
3. 轉印 (transfer)
剪裁適當大小之 PVDF membrane,以 methanol 潤濕後,以去離子水洗淨浸泡備用。
電泳結束後將 SDS-PAGE gel 由玻片上取下放在 3M paper 上,再將 PVDF membrane 貼 於 SDS-PAGE gel 上,再蓋上一層 3M papper,以塑膠棒趕走其間之氣泡,然後置於轉印 槽,置於 4℃冰箱中以 100 mA 之電流通電隔夜或 300 mA 之電流轉印 3 小時。
4. Immunoblot
轉印完成後,將 PVDF membrane 以含 5 % 脫脂奶粉的 TBST (tris buffered saline with Tween-20)blocking buffer 作用 30 分鐘後,倒掉 blocking buffer,加入 primary antibody 作 用 2 小時,再以 1× TBST 清洗三次,每次 5 min,然後加入 secondary antibody 如:
anti-mouse IgG conjugated alkaline phosphatase 或 anti-rabbit IgG conjugated horse radish peroxidase 等 特 異 性 抗 體 作 用 1 小 時 , 再 以 TBST 清 洗 三 次 , 最 後 以 enhanced chemiluminescence (ECL)做呈色。ECL 呈色指 PVDF membrane 加上 1 ml ECL 作用 3 分 鐘後以水漂洗,包入保鮮膜中放入含有 X-film 的卡匣,再以底片曝光呈色。
流式細胞儀分析粒線體膜電位 1. 細胞處理
使 用 1 μM 5,5',6,6'-tetrachloro-1,1',3,3'-tetraethylbenzimidazolylcarbocyanine iodide (JC-1, Molecular Probe, Inc., Europore BV)偵測細胞中粒線體膜電位的變化。粒線體在呼 吸氧化過程中,將產生的能量以電化學位能儲存於粒線體內膜(約 250 mV/5~10 nm),稱 之為 mitochondrial membrane protential (△),為外正內負的形式,並進行電子傳遞鏈,
產生 ATP 供細胞使用。JC-1 為親脂性的陽離子化合物可結合至粒線體內膜,當 △小 於 100 mV 時會以 monomer 的形式存在,以 488 nm 激發光下會放出 525 nm 的綠色螢 光;而當 △逐漸升高,JC-1 會聚集在粒線體內膜形成橘色的 J-aggregates (590 nm)。
JC-1 目前因較傳統粒線體膜電位有更高的敏感性和解析分辨能染劑,被廣泛使用於共軛 焦顯微鏡或流式細胞儀(flow cytometry)細胞來觀察粒線體的形態、移動與膜電位之改 變。在細胞處理好後,避光環境下加入 1 μM JC-1 於 medium 中,放入 37℃培養箱中作 用 15 分鐘,以 PBS 沖洗二次,覆蓋蓋玻片時,載玻片中央滴上,以指甲油封片後,立 即以共軛顯微鏡觀察。
結果
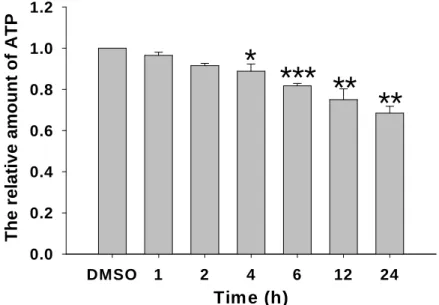
1 TCDD影響細胞粒線體的ATP形成能力
本實驗進一步觀察經TCDD處理後分析細胞ATP含量的變化。首先以TCDD處理細胞,在
不同時間點2小時、4小時、6小時、8小時、12小時、24小時收集其細胞標本。以冷光測定儀 的方法,來分析細胞內ATP產生的情形。依照不同收集細胞的方式,實驗結果Fig. 1顯示,在 20 nM的TCDD處理細胞24小時後,若僅將仍貼附在培養皿的細胞做分析,細胞ATP的產生減 少為0.68倍(p<0.01)。
2 TCDD處理影響細胞粒線體膜電位改變
本實驗以JC-1染色來觀察細胞粒線體膜電位的變化。實驗結果如Fig. 2所示,圖A, C, 是 將以0.1% DMSO處理的控制組細胞,可觀察到細胞粒線體呈現黃色並有聚集的現象,圖B, D 是將以20 nM TCDD處理的實驗組細胞,可觀察到細胞粒線體螢光呈現綠色平滑的現象。觀察 結果可證明經TCDD處理後,人類滋養層細胞粒線體膜電位有下降的情形發生。
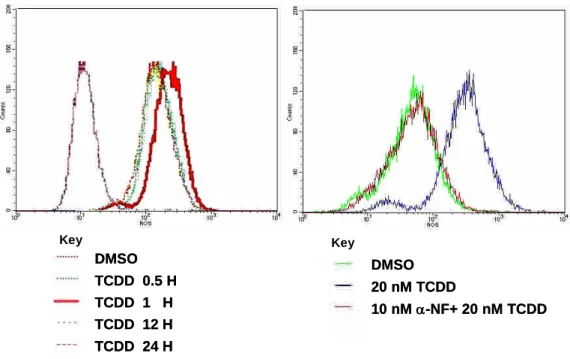
3 TCDD引發人類滋養層細胞(3A-sub-E cells)中活性氧類(ROS)增加
觀察四氯戴奧辛是否會對人類滋養層細胞3A-sub-E cells造成細胞內粒線體及細胞氧化性 傷害,預先將sub-3AE cells以20 nM TCDD處理,並於37 ℃、5% CO2 環境下進行培養,待反 應時間到達後,立即將培養皿置於冰浴,以終止反應。收集之細胞將立即進行實驗流程與分 析。將人類滋養層細胞以20 nM的TCDD在0小時、5分鐘、15分鐘、0.5小時、1小時、2小時、
4小時、6小時、8小時、12小時、24小時、48小時時間點處理細胞後,收集細胞,並以H2DCFDA 進行染色,以流式細胞儀的方法,來分析細胞內ROS產生的情形。我們可發現在20 nM的TCDD 處理細胞1小時後,ROS的產生增加1.50倍(p<0.05),如Fig. 4所示。另外,爲觀察細胞內ROS 分布情形,實驗將細胞以H2DCFDA染色,並以共軛顯微鏡觀察和紀錄。觀察結果如Fig. 11所 示,在TCDD處理的實驗組細胞ROS螢光表現較以0.1% DMSO處理的控制組細胞為高。顯示 經TCDD刺激後,細胞內ROS有增加的情形。
4. TCDD引發缺氧誘發因子(Hypoxia inducing factor-1; HIF-)的表現
為確定TCDD是否會對人類滋養層細胞3A-sub-E cell造成組織缺氧的情形,將欲測試之 TCDD(溶劑DMSO)加入3A-sub-E cells中與之反應,並於37 ℃、5% CO2 環境下進行培養,將 20 nM的TCDD加入3A-sub-E cell,在0.5小時、1小時、2小時、4小時、6小時、8小時不同時 間點處理後,收集其cell lysate以西方墨點法分析HIF-1的表現。實驗結果如Fig. 8所示,當20 nM TCDD與3A-sub-E cell反應時間達1小時,與控制組相較,TCDD能誘發1.88倍於控制組之
HIF-1產生(1.174 ± 0.044 vs 2.207 ± 0.091 HIF-1per 50 μg protein)(p<0.005))。而HIF-1
protein隨表現在不同濃度2 nM、10 nM、20 nM、60 nM、100 nM的TCDD處理1小時,其HIF-1
依濃度增加而有遞增的現象。由實驗結果可發現,以20 nM TCDD處理一小時,與控制組相較,
可觀察到HIF-1的表現有3.51倍的增加 (Fig. 3)。
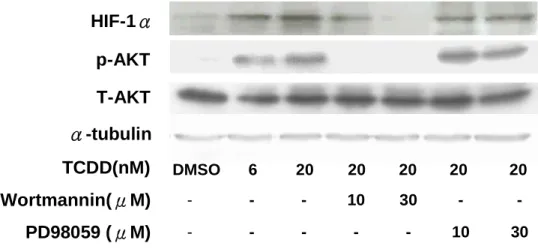
5. TCDD藉由PI3K/Akt訊息傳遞調控路徑誘導HIF-1蛋白質表現
我們為了進一步確認TCDD能誘導HIF-1蛋白表現,是經由哪一條訊息傳遞路徑來誘 導。依2003年Mottet研究指出HIF-1可能經由PI3K-kinase及其下游蛋白,包括serine-threonine kinases、Akt (protein kinase B) pathway等訊息傳遞路徑刺激HIF-1蛋白產生。實驗以PI3K (Phosphatidylinositol 3-kinase)抑制物wortmannin與MEK (Mitogen-activated ERK kinase)抑制物 PD98059,來探討TCDD誘發HIF-1表現之分子訊息傳遞。預先加入10 μM、30 μM wortmannin 或10μM、30 μM PD98059分別與細胞共同培養30分鐘,再加入20 nM TCDD 1小時後,收集其 cell lysate以西方墨點法分析HIF-1與PI3K下游phospho-Akt的表現,實驗結果如Fig. 5所示,
預先加入wortmannin的組別、預先加入PD98059的組別與只加入TCDD控制組結果相較,
wortmannin抑制約1.53倍於控制組之phospho-Akt產生,同時wortmannin也抑制HIF-1的表 現。而預先加入PD98059的組別則沒有抑制phospho-Akt及HIF-1表現。運用PI3K抑制劑 wortmannin的添加觀察HIF-1的表現,我們確認TCDD藉由PI3K/Akt訊息傳遞調控路徑誘導 HIF-1蛋白質表現。
6. ROS參與在TCDD所誘導的HIF-1蛋白質表現訊息傳遞調控路徑
因TCDD會誘發細胞ROS與NO的產生,我們進一步探討由TCDD引發的HIF-1表現是否 與ROS、NO有關,因此實驗另以ROS抑制物L-NAC (L-N-acetylcysteine)與NOS (NO synthase) 抑制劑L-NAME (N-nitro-L-arginine- methyl ester),來探討TCDD誘發HIF-1表現之分子訊息傳 遞。首先利用100、300 μM L-NAME與100、300 μM L-NAC預先處理細胞30分鐘,之後再分 別以6、20 nM TCDD刺激細胞,1小時後收集其cell lysate以西方墨點法分析HIF-1與PI3K下 游phospho-AKT的表現,實驗結果如Fig. 6所示。預先加入L-NAC的組別、預先加入L-NAME 的組別與只加入TCDD控制組結果相較,300 μM L-NAC抑制phospho-AKT產生,同時也抑制 HIF-1的表現,另外,L-NAME則沒有明顯抑制的現象。運用ROS抑制劑的添加觀察HIF-1
的表現,我們進一步確認ROS參與在TCDD所誘導的HIF-1蛋白質表現訊息傳遞調控路徑中。
討論
本實驗證明TCDD刺激人類滋養層細胞造成細胞內ROS增加及8-oxo-dG增加,與粒線體 DNA斷損突變。觀察人類胚胎滋養層細胞,可發現在TCDD刺激下,HIF-1蛋白增加,證明 TCDD使細胞處於化學性的缺氧狀態(chemical hypoxia)下,並伴隨有ROS的產生。推測人類胚 胎滋養層細胞遠較其他細胞依賴氧氣的需求(14),所以在由TCDD引起細胞缺氧環境下,對於 人類胚胎滋養層細胞粒線體易造成傷害,進而產生大量的ROS,促使細胞走向凋亡。
本實驗證實TCDD會引發人類胚胎滋養層細胞產生缺氧狀態。已知完整的胎盤發育是胚胎 發育的重要因子之一,目前已發現許多的外在因子包括genetic factors、immunological factors、
environmental factors皆影響胎盤的發育,甚至進而影響胚胎的發展。先前已有研究報告顯示,
缺氧會誘導人類細胞株發生細胞凋亡,但其真正的機轉到現在仍然不是很清楚。由以上累積 實驗結果,推論在體內,若長期暴露於TCDD環境下,可能有類似的情形,婦女早期懷孕時 可能造成子宮胎盤組織缺氧及ROS大量產生,促使胎盤細胞凋亡現象增加,影響胚胎的發育 或胚胎死亡(15,16)。在動物模式實驗,TCDD會誘導C57BL/6 mice氧化壓力增加,使組織中氧 化DNA增加。
HIF-1在TCDD-induced toxic reaction是扮演怎樣的角色?許多文獻指出,HIF-1蛋白表 現量與活性會隨著細胞種類(cell type)與刺激物(types of stimuli)的不同而有差異,而其中所參 與的訊息傳遞路徑也有不同,包括了MAPK pathway和PI3K/Akt pathway皆被證實,除了參與 HIF-1蛋白translation,亦參與HIF-1 activity的調節。同時亦有文獻指出,在這些kinase pathway中,ROS為一個中間訊息傳遞的步驟影響HIF-1蛋白表現與活性。ROS可能經由 ligand–independent 活化過程活化 growth-factor之receptors , 並聚集 (recruit)Akt pathway活 化 HIF-1。綜合實驗結果顯示,當細胞受到TCDD的刺激而引發細胞凋亡的過程中,產生ROS 進而引發HIF-1轉錄因子的活化。而細胞內ROS大量產生會造成細胞內脂質過氧化、8-OH-dG 增加,並造成粒線體功能缺陷及基因突變。我們推論由TCDD造成的胚胎死亡等臨床現象,
可能與TCDD所誘導產生的大量ROS及HIF-1缺氧因子有密切的關係。
圖表
Fig. 1 Declined ATP production in TCDD-treated trphoblast cells. ATP production were determined by luciferin reagent as described in Methods. Plots are presented as meanS.D. (n=3).
*, p<0.05 compared with the control groups; **, p<0.01 compared with the control groups; ***,
p<0.001 compared with the control groups.
Fig. 2 Fluorescence images of TCDD-treated trophoblast cells by JC-1 staining. Human trophoblast were treated without (A, C) or with (B, D) 20 nM TCDD for 16 h. Illustrated the JC-1 accumulated preferentially in polarized mitochondria, exisiting as green fluorescent monomer at low membrane potentials and as orange fluorescent aggregates at high membrane potentials.
Manification, 100X in A and B ; 200X in C and D.
Time (h)
DMSO 1 2 4 6 12 24
T h e re la ti v e a m o u n t o f A T P
0.0 0.2 0.4 0.6 0.8 1.0 1.2
** **
* ***
0.1% DMSO TCDD
A B
C D
Fig 3 Time-dependent and dose-responsive effect of dioxin on HIF-1α expression in human trophoblast cell. Cell were incubated with 100nM TCDD at 37℃for various time period, and then immunodetect with HIF-αspecificantibody as described in Methods.Equal loading in each lanewasdemonstrated by thesimilarintensitiesofα-tubulin.The mean densitometry data from independent experiments werenormalized to the result obtain in cells in the absence of TCDD(control).Plots are mean + S.E. values(n=3);*,p<0.05 compared with the control;**,p<0.01 compared with the control;***,p<0.001 compared with the control.
Fig 4 Effect of TCDD on reactive oxygen species production in human trophoblast cell.
Cell were incubated with 100nM TCDD at 37℃for various time period. A, the generation of reactive oxygen species production were shown as the shift of the fluorescent peak. B, the reduced generation was shown in the supplementation of Ahr inhibitor-NF.
DMSO TCDD 0.5 H TCDD 1 H TCDD 12 H TCDD 24 H
DMSO 20 nM TCDD
10 nM-NF+ 20 nM TCDD Key Key
DMSO TCDD 0.5 H TCDD 1 H TCDD 12 H TCDD 24 H
DMSO 20 nM TCDD
10 nM-NF+ 20 nM TCDD Key Key
HIF-1/-tubulin
0.0 0.5 1.0 1.5 2.0 2.5
DMSO 0.5 1 2 4 6 8
Tim e (h)
** **
*
**
***
DMSO 0.5 1 2 4 6 8 20 nM TCDD (h)
HIF-1α α-tubulin
0 1 2 3 4
HIF-1/-tubulin
DMSO 2 6 20 60 100
Time (h)
**
***
DMSO 2 6 20 60 100 TCDD (nM)
HIF-1α α-tubulin
**
***
***
Fig 5 Effect of inhibitors of PI3K and MEK on HIF-1 expression in TCDD-treated trophoblast cells. Immunoblot detection of HIF-1αprotein in trophoablast cells that pretreated with Wortmannin or PD98059, in the presence or absence (-) of TCDD(same induction as in Fig. 5).
Control cell exposed to 0.1% DMSO are shown in lane 1. Blots are representative of results obtain in three separate experiments. Equal loading in each lane was demonstrated by the similar intensitiesofα-tubulin.
Fig 6 Effect of NO and ROS inhibitors mediated accumulation of HIF-1αexpression in human trophoblast cell. Immunoblot detection of HIF-1αprotein in trophoablast cells that pretreated with or without L-NAME or NAC, in the presence or absence (-) of TCDD. Control cell exposed to 0.1% DMSO are shown in lane 1. Blots are representative of results obtain in three separate experiments. Equal loading in each lane was demonstrated by the similar intensities of α-tubulin.
參考文獻
1. Whitlock, J.P., Jr. (1990) Genetic and molecular aspects of 2,3,7,8-tetrachlorodibenzo-p- dioxin action. Annu Rev Pharmacol Toxicol, 30, 251-277.
2. Gregoraszcuzuk EL. (2002) Dioxin exposure and porcine reproductive hormone activity. ARTIGO, 18, 453-462.
3. Ishimura R,Ohsako S, Kawakami T, Sakaue M, Aoki Y, Tohyama C. (2002) Altered protein 300
100 -
- -
-
L-NAC(μM) -
- -
300 100
- -
L-NAME(μM) -
TCDD(nM) α-tubulin T-Akt p-Akt HIF-1α
20 20
20 20
20 6
DMSO
30 10
- -
- -
PD98059 (μM) -
- -
30 10
- -
Wortmannin(μM) -
TCDD(nM) α-tubulin T-AKT p-AKT HIF-1α
20 20
20 20
20 6
DMSO
profile and possible hypoxia in the placenta of 2,3,7,8-tetrachlorodibenzo-p-dioxin- exposed rats.
Toxic. Appl. Pharm., 185, 197-206.
4 Sakurai K, Todaka E, Saito Y, Mori C. (2004) Pilot study to reduce dioxins in the human body.
Intern Med., 43, 792-795.
5. Miller KP, Borgeest C, Greenfeld C, Tomic D, and Flaws JA. (2004) In utero effects of chemicals on reproductive tissues in females. Toxic. Appl. Pharm. 198, 111-131.
6. Wu Q, Ohsako S, Ishimura R, Suzuki JS, and Tohyama C. Exposure of mouse peimplantation embryos to 2,3,7,8-tetacholorodibenzo-p-dioxin (TCDD) alters the methylation status of imprinted genes H19 and Igf2. Biol. Reprod. 70, 1790-1797.
7. Schroedl, C., McClintock, D.S., Budinger, G.R. and Chandel, N.S. (2002) Hypoxic but not anoxic stabilization of HIF-1alpha requires mitochondrial reactive oxygen species. Am J Physiol Lung Cell Mol Physiol, 283, L922-931.
8. Camejo MJ, Proverbio F, Febres F, and Casart YC. (2003) Bioactive to immunoreactive ratio of circulating human chorionic gonadotropin as possible evaluation for the prognosis of threatened abortion. Europ. J. Obs. Reprod. Biol., 109, 181-184.
9. Chandel, N.S., Maltepe, E., Goldwasser, E., Mathieu, C.E., Simon, M.C. and Schumacker, P.T.
(1998) Mitochondrial reactive oxygen species trigger hypoxia-induced transcription. Proc Natl Acad Sci U S A, 95, 11715-11720.
10. Chandel, N.S., McClintock, D.S., Feliciano, C.E., Wood, T.M., Melendez, J.A., Rodriguez, A.M.
and Schumacker, P.T. (2000) Reactive oxygen species generated at mitochondrial complex III stabilize hypoxia-inducible factor-1alpha during hypoxia: a mechanism of O2 sensing. J Biol Chem, 275, 25130-25138.
11. Berra, E., Benizri, E., Ginouves, A., Volmat, V., Roux, D. and Pouyssegur, J. (2003) HIF prolyl-hydroxylase 2 is the key oxygen sensor setting low steady-state levels of HIF-1alpha in normoxia. Embo J, 22, 4082-4090.
12. Kaufmann, P., Black, S. and Huppertz, B. (2003) Endovascular trophoblast invasion: implications for the pathogenesis of intrauterine growth retardation and preeclampsia. Biol Reprod, 69, 1-7.
13. Bae S, Xiao Y, Li G, Casiano CA, and Zhang L. (2003) Effect of maternal chronic hypoxic exposure during gestation on apoptosis in fetal rat heart. Am. J. Physiol. Heart. Circ. Physiol. 285, H983-H990.
14. Mackova, M., Kilani, R. T., Davidge, S. T., and Guilbert, L. J. (2003). The effect of oxygen tension on intracellular survival signalling in primary villous trophoblasts. Placenta 24, 627-637.
15. Miller KP, Borgeest C, Greenfeld C, Tomic D, and Flaws JA. (2004) In utero effects of chemicals on reproductive tissues in females. Toxic. Applied Pharm. 198, 111-131.
16. Burton GJ, Jauniaux E.(2004) Placental oxidative stress: from miscarriage to preeclampsia J Soc.
Gynecol. Investig. 11:342–52.