Chapter 2
The vicariance and long distance dispersal of Ceriops tagal (Rhizophoraceae) on coasts of the Kra Isthmus inferred from
chloroplast DNA Abstract
The Malay Peninsula (and previously ancient Sundaland) is commonly considered to be a barrier that isolates organisms living in the South China Sea from those in the Bay of Bengal. However, during the interglacial period approximately 5 Mya sea levels rose and consequently there were leaks across the Kra Isthmus, the narrowest part of this peninsula. Thus, organisms living on both sides of the Kra Isthmus had opportunities to migrate or disperse to each other’s ranges. In the study reported here, we examined the genetic composition of Ceriops tagal, a eumangrove species that grows along the coasts of both sides of the Kra Isthmus to retrace divergence events and evaluate the probability that long distance dispersal has occurred. The gene flow among populations (Nm) on the same side of the isthmus was found to be higher than trans-isthmus flow, and differences between populations on the opposite sides of the isthmus accounted for approximately a third of the total genetic variation detected, illustrating its vicariant effects. Furthermore, the estimated time of divergence between populations on the two sides is consistent with the emergence of the Kra Isthmus, indicating that genetic continuity was disrupted by the geographic barrier.
However, recent gene flow has obscured the phylogenetic relationships between eastern and western haplotypes. In addition, evidence of pre-isthmus range expansion and subsequent restrictions of gene flow resulting from isolation by distance was obtained from nested clade analysis based on user-defined-distances corresponding to the distances across the peninsula and the sea route around it. Trans-isthmus long distance dispersal probably occurred through the southern Malay Peninsula. These findings indicate that although the Malay Peninsula separates populations on opposite sides of the isthmus, interglacial leaks at the Strait of Malacca could have provided a corridor for cross-sea-area gene flow. Thus, allopatric speciation may have been arrested due to genetic homogenization via rare long distance dispersal.
Keywords Ceriops tagal; Kra Isthmus; land barrier; dispersal; vicariance
Introduction
Geographic isolation is generally considered to be a major driver of population differentiation (Braillet et al. 2002; Liao et al. 2007; Roy et al. 2006), that can even drive allopatric speciation (Hayashi and Kawata 2006; Hoskin et al. 2005; Near and Benard 2004; Starrett and Hedin 2007; Stevens and Hogg 2004). The most well known cases are those in which marine organisms are isolated by isthmuses. For example, the Isthmus of Panama, which emerged approximately 3.5 Mya (Coates et al.
1992) appears to have caused substantial differentiation of populations and divergence of species in the Atlantic and Pacific Oceans (Smith et al. 2006; Steeves et al. 2005).
Similar barriers are found in many other places, e.g. the Malay Peninsula and the Baja California Peninsula. During short periods of geological history, organisms isolated by such isthmuses must rapidly accumulate fixed differences to drive differentiation.
In other words, their divergence must involve the absence of gene flow. However, if there has been insufficient time for fixed differences to accumulate, or genetic connections are maintained by further dispersal and migration, the allopatric speciation process will be incomplete, or cease (Berthier et al. 2006; Trakhtenbrot et al. 2005; Wiens et al. 2006). The process that can most strongly antagonize speciation is long distance dispersal (LDD) (Cain et al. 2000), although most plant seeds have poor ability to disperse over long distances (Cain et al. 2000). Thus, gene flow generally proceeds slowly, often through short incremental steps via intermediate populations over long periods of time. For instance, the seed dispersal rates of forest understory species generally only range from 0.0 to 2.5 m/yr (Cain et al. 2000).
However, the “tail” of dispersal curves can be long (Portnoy and Willson 1993), and thus much longer distance dispersal can occasionally occur, especially via the sea, since it can then be facilitated by sea currents.
However, the salinity may harm the seeds or fruits that come into prolonged contact with the sea, as noted (inter alia) by Darwin in his great work On the Origin of Species (1859). In experiments he performed on the effects of salinity on seed germination, seeds representing 64 of 87 species were able to germinate after four weeks soaking in sea water, but very few were able to germinate after soaking for 137 days. Darwin also refers to an experiment performed by Martens, who found that seeds of only 18 of 98 tested coastal plants were able to germinate after floating on the sea for six weeks. Even mangrove species, which are commonly considered to be
saline tolerant, may be harmed by prolonged soaking in saline water. Clarke et al.
(2001) found that propagules of five of 14 examined mangrove species were unable to float in fresh water, and four were unable to float in saline water. These experiments illustrate the difficulties involved in oversea LDD of plants.
Nevertheless, the globally discontinuous distribution of mangrove species (Duke et al. 2002) indicates that they have the ability to “effectively” disperse (i.e. disperse and establish) over seas (Cain et al. 2000). Thus, they appear to be able to avoid or surmount the problems associated with LDD, but their LDD processes are likely to be highly stochastic, and intriguing (Duke et al. 2002; Minchinton 2006). However, although many researchers have considered their disjunct geographic distribution (Duke 1995; Duke et al. 2002) and discontinuity of genetic composition (Chiang et al.
2001; Duke et al. 1998b; Liao et al. 2007; Su et al. 2006; Su et al. 2007; Tan et al.
2005), fewer have explored the underlying processes involved.
In the study reported here we examined populations of Ceriops tagal (Perr.) C.
B. Rob. distributed on both sides of the Kra Isthmus, the narrowest part of the Malay Peninsula, to examine the vicariant effect of the Kra Isthmus and long-distance dispersal between the two sides of the Peninsula. We addressed the following questions. Firstly, has the Kra Isthmus effectively isolated mangroves distributed on the two sides, and if so how strongly? Secondly, have there been any detectable LDD events across the isthmus, and if so did they probably involve dispersals across the Kra Isthmus or via the sea route around the Malay Peninsula? The findings provide several indications regarding the effects of the genetic differentiation imposed by the Kra Isthmus barrier. In addition to discussing these findings, possible mechanisms of trans-isthmus LDD of mangroves are considered below.
Materials and methods Sample collection and cpDNA analysis
We sampled leaves from 89 individuals in total, representing four populations of C.
tagal at various sites along the east coasts of the Kra Isthmus of the Malay Peninsula and five populations at various sites along the west coast (Table 2-1 and Fig. 2-1). All of the sampled individuals were spaced at least 10 m apart. The collected leaves were dried immediately with silica gel prior to DNA extraction. Total genomic DNA was
isolated from the samples using a commercial DNA extraction kit (BIOMAN Co.), dissolved in TE buffer (10 mM Tris, 0.1 mM EDTA, pH 8.0) and stored at -20℃ until analysis. Two chloroplast regions (atpB-rbcL and trnL-trnF) were selected for amplification and sequencing in order to trace propagule dispersal patterns. The primers, PCR conditions, and sequencing protocols used are described in Liao et al.
(2007). Sequences obtained in this study have been deposited in the NCBI database under the accession numbers: DQ983274-DQ983276, DQ983280-DQ983281 and DQ983284-DQ983307 (trnL-trnF); DQ983241, DQ983245, DQ983248, DQ983250-DQ983252, DQ983257-DQ983260 and DQ983262-DQ983273 (atpB-rbcL).
Table 2-1 Sampling sites
Population Abbreviation Longitude Latitude Geographic area
Ao Phangna National
Park AP 08°24'N 98°31'E West Kra, Bay of Bengal
Phang-Nga PN 08°24'N 98°30'E West Kra, Bay of Bengal Southern Phuket island PK 07°52'N 98°23'E West Kra, Bay of Bengal
Ranong RN 09°55'N 98°37'E West Kra, Bay of Bengal
Sirinat National Park SN 08°11'N 98°17'E West Kra, Bay of Bengal Mu Ko Chumphon
National Park MK 10°22'N 99°10'E East Kra, Gulf of Thailand Surat Thani (Korn Nan) ST 09°07'N 99° 20'E East Kra, Gulf of Thailand Thungkra-Swi Amphur
Sawi SW 10°15'N 99°05'E East Kra, Gulf of Thailand
Tumethong Amphur
Patew PT 10°02'N 99°08'E East Kra, Gulf of Thailand
Population genetic analyses
After sequence alignment, the parameters nucleotide diversity (π), haplotype diversity (Hd), and θ – based on both the total number of mutations (η) and segregating sites (S) – were estimated for each population using DnaSP 4.0 (Rozas et al. 2003).
Pairwise population differentiation was estimated by calculating FST with 1000 permutations using ARLEQUIN v. 3.0.1 (Excoffier et al. 2005) and the index of gene
flow (Nm) was calculated from Nm=(1-FST)/2FST. A neighbour joining tree, based on the matrix of pairwise FST values was drawn to visualize the magnitude of genetic differentiation and relationships among the populations using MEGA 3.1 (Kumar et al.
2004). The overall magnitude of population differentiation (ΦST) and the variance of the hierarchical structure of populations were measured by Analysis of Molecular Variance (AMOVA) using ARLEQUIN v. 3.0.1 (Excoffier et al. 2005).
Phylogeographic analyses
For phylogeographic analyses, every long-fragment indel was treated as arising from a single evolutionary event and recoded as described in our previous paper (Liao et al.
2007). The phylogenetic relationships among the haplotypes were analyzed using the unrooted neighbor-joining method with the settings of the Kimura’s (1980) two-parameter model and pairwise deletion. The statistical significance of the phylogenetic groups in the resulting tree was tested by bootstrap resampling (1000 replicates in each case) using MEGA 3.1.
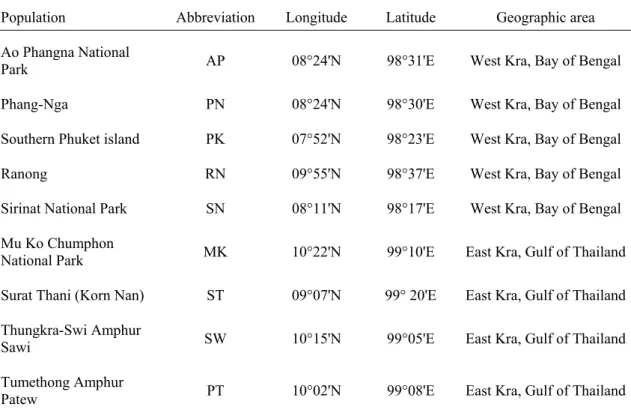
Fig. 2-1 Map of the sampling sites. The sum of the distances of dotted lines is the shortest distance between the most southern populations of the east and west Kra Isthmus around the Malay Peninsula and is roughly equal to 2,100 km (a+b+c+d+e≒2,100 km).
This distance was used for the sea-route-distance based NCA.
A haplotype network was constructed by the minimum spanning network method using ARLEQUIN v. 3.0.1 (Excoffier et al. 2005) and the genetic data were input into GEODIS v. 2.0 (Posada et al. 2000), together with information on the geographic locations of the populations to perform permutation tests to assess the
significance of associations (if any) between their genetic and geographic distributions. This was accomplished by examining the clade distances (DC) and nested clade distances (DN) generated from the population data. DC is defined as the average distance of an individual from the geographic center of all individuals within the same nesting clade, while DN measures the relative geographic distribution to other clades in the same higher-level nesting clade (Templeton 2001; Templeton et al.
1995). Two nested clade analyses (NCAs) were performed, one in which geographic (great circle) distances were used, and one in which sea-route distances were used.
The great circle distances among populations were determined from latitudinal and longitudinal coordinates (Table 2-1), and the sea-route distances were rough estimates of the shortest distances between populations by sea (Fig. 2-1). The rough sea-route distance from an eastern Kra population (pop E) to a western Kra population (pop W) is
DE-W = DE-ST + 2100 km + DW-PK,
where ST and PK are the most southerly populations of the eastern and western Kra Isthmus, respectively, DE-ST is the distance from pop E to pop ST and DW-PK is the distance from pop W to pop PK. Templeton’s (2004) inference key was then applied to the results to determine the NCA outcome. In this analysis we ignored C. tagal populations distributed in the southern Malay Peninsula, thus some answers from the inference key may be “Sampling Design Inadequate”.
Results Genetic diversity
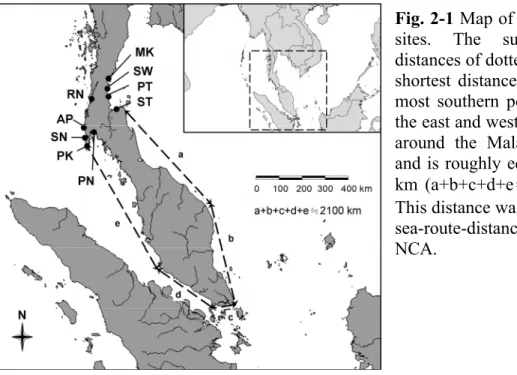
Data on sequences of both the atpB-rbcL and trnL-trnF cpDNA spacers were used in all of the genetic analyses in this study. In all, 1198 base-pair length fragments were obtained after alignment, containing 110 polymorphic sites, including 10 long-fragment indels, several substitutions, and one-base indels. When each long-fragment indel was recoded as a single mutation event (site), there were 78 variable sites, including 20 singletons and 58 informative mutations, among the sequences obtained from the investigated C. tagal populations on the east and west coasts of the Kra Isthmus. A total of 43 haplotypes (across the 89 samples) were obtained and the overall haplotype diversity was 0.974, with diversities of individual
populations ranging from 0.400 (AP) to 1.000 (RN). Overall nucleotide diversity (π) was 0.00949, and ranged for individual populations from 0.00037 (AP) to 0.01861 (RN), while estimates for θ ranged from 0.00044 (AP) to 0.01589 (MK), with an overall value of 0.01703.
The average number of nucleotide differences between populations on the east and west sides of the isthmus was 9.479, and there were 0.00918 nucleotide substitutions per site (K), on average, between them. Populations on the east and west coasts of the Kra Isthmus shared 28 mutations (excluding gaps), there were 37 mutations that were polymorphic in populations on the west coast but monomorphic in the east coast populations, and 24 mutations that were polymorphic in the east coast populations but monomorphic in the west coast populations. Thus, the west coast populations showed slightly higher genetic diversity than the east coast populations.
This finding is also supported by the π and θ estimates. The genetic diversity parameters for each population are presented in Table 2-2.
Table 2-2 Genetic diversity of the investigated Ceriops tagal populations, according to estimates of: haplotype diversity (Hd), nucleotide diversity (π) and θS. N is the sample sizes and H refers to the number of haplotypes.
Population N H Hd π θ
MK 9 8 0.972 0.01104 0.01589
SW 10 8 0.956 0.00217 0.00232
PT 8 7 0.964 0.00191 0.00181
ST 9 7 0.917 0.00410 0.00410
RN 10 10 1.000 0.01861 0.01546
AP 5 2 0.400 0.00037 0.00044
SN 21 13 0.957 0.00940 0.00940
PK 8 3 0.607 0.00215 0.00250
PN 9 5 0.806 0.00268 0.00239
E-Kra 36 22 0.967 0.00584 0.01228
W-Kra 53 26 0.966 0.01160 0.01381
Total 89 43 0.974 0.00949 0.01703
Population differentiation and gene flow
All of the pairwise FST values were significant at the P<0.05 level except those for the following four pairs of populations: RN-PN, MK-ST, SW-PT, and PT-ST. Both members of each of these pairs were located on the same side of the Kra Isthmus (Table 2-3). The pairwise Nm values, estimated from the reciprocal FST values between populations within the east Kra Isthmus were all higher than 1.0, indicating that gene flow via propagule dispersal of C. tagal has frequently occurred on the east side of the Kra Isthmus. High Nm values were also obtained in several pairwise comparisons of populations on the west side of the isthmus (AP-RN, PN-RN, AP-SN, PN-SN, and RN-SN). However, the Nm values obtained were higher than one for just two of the pairwise comparisons of populations on the east and west sides (MK-RN
and RN-ST). These results indicate that the cpDNA gene flow (propagule dispersal) was higher among mangrove populations along coasts on the same side of the isthmus than along coasts on opposite sites. Thus, the Kra isthmus does indeed appear to be a land barrier for the dispersal of mangroves, but nevertheless significant effective dispersal of maternal genes between the sides seems to have occurred.
Population genetic structure
The results of the hierarchical AMOVA indicate that within-population variation accounts for most of the observed genetic variation (51.96%), followed by variation between populations on the east and west sides of the Kra Isthmus (31.56%).
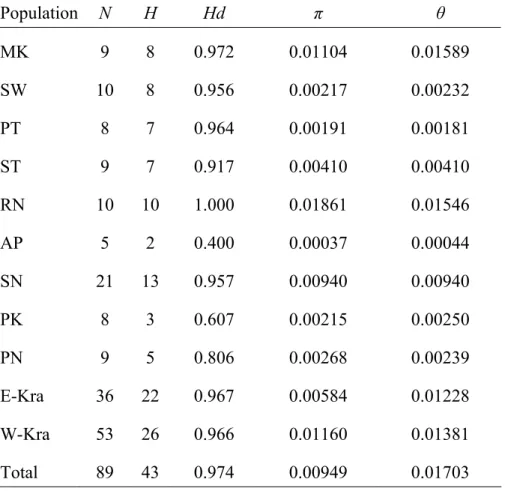
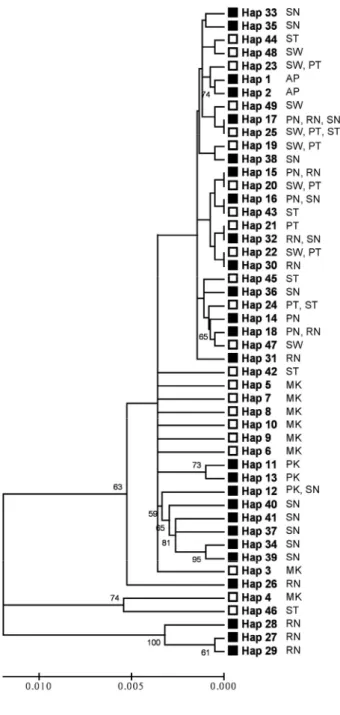
Variation among-populations-within these groups accounted for just 16.48% of the total variation. The hierarchical Φ statistics revealed the same pattern; that most of the genetic differentiation is within-populations (ΦST = 0.48040), followed by among groups (ΦCT = 0.31560) and variation among populations within groups made the least contribution (ΦSC = 0.24079). All of the Φ statistics and these differences in percentages of variation are significant (Table 2-4). The grouping of populations in the NJ tree derived from the pairwise FST values is consistent with their geographical distribution, although the branch between the east and west populations is short (Fig.
2-2). Thus, both the AMOVA results and NJ population tree indicate that the Kra Isthmus does indeed structure the C. tagal populations, despite the short genetic distance between populations on its two sides.
Table 2-4 Summary of results of molecular variance (AMOVA) for the East and West Kra areas of Ceriops tagal. The significance (P) of the variance was based on 1000 permutations.
Source of Variation d.f. % total variance Φ P
Among groups 1 31.56 0.31560 <0.005
Among populations within groups 7 16.48 0.24079 <0.0001
Within populations 80 51.96 0.48040 <0.0001
Fig. 2-2 Neighbor-Joining tree of Ceriops tagal constructed by pairwise FST. The scale bar indicates the FST differences and the divergence time. The time scale, based on the estimated time of divergence of the Gulf of Thailand and the Bay of Bengal populations, is approximately 5 Mya
Phylogeographic inferences from NCA
Due to the apparent discrepancies between the groupings in the haplotype phylogenetic tree and geographic distribution, and the unresolved, ambiguous, statistically weak relationships in the phylogenetic tree were constructed (Fig. 2-3).
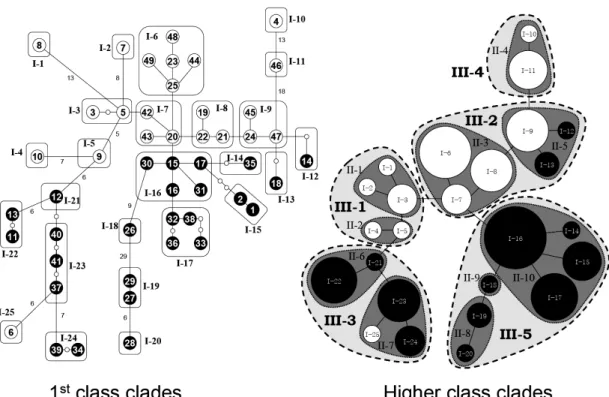
The tree provides indications of recent gene flow (Bernardi et al. 2003), but no evidence regarding the populations’ history. By contrast, the minimum spanning network splits east and west clades well, although they are only separated by just one or a few steps (Fig. 2-4). Roughly speaking, the eastern populations were located in the interior clades and the western populations in the terminal clades. The central clades, which may carry the most ancient evolutionary information, included both eastern and western haplotypes (subclades I-7 and I-16). Furthermore, several haplotypes located in the terminal clades were loosely connected to each other with long missing links (Fig. 2-3), suggesting a long evolutionary separation of these populations. Both the great-circle and sea-route distance-based analyses detected insignificant nested-clade distance associations amongst the lower level clades, indicating that the null hypothesis of no association between haplotype and geographical distributions should not be rejected for these clades.
However, both the great-circle-distance and sea-route-distance based nested clade analyses generated four clades that showed significant associations with DC, DN
or I-T values (Table 2-5), and both analyses yielded similar inferences for three of these four clades. A restricted gene flow with isolation by distance (IBD) evolutionary scenario was inferred for Clade II-10, indicating that within this central-located clade
of the western Kra Isthmus short-distance dispersal may have occurred, but only rarely. In contrast, Clade II-5 (an east Kra Isthmus clade connected to each of two western Kra Isthmus haplotypes by single missing links), was inferred to have arisen from long distance dispersal (LDD) according to the analyses based on both types of geographic-distance. Clade III-3 is a clade of west Kra Isthmus haplotypes, with many long missing links (inferred consequences of “Contiguous range expansion”) and one east Kra Isthmus haplotype.
Fig. 2-3 Unrooted Neighbor- Joining tree of Ceriops tagal haplotypes. Numbers on branches are bootstrap values of 1,000 replicates. White squares indicate haplotypes observed in the east Kra Isthmus and black squares are haplotypes observed in the west Kra Isthmus. The scale bar indicates the inferred frequency of substitutions per nucleotide site.
Fig. 2-4 Nesting scheme for one-step (1st class) and the higher (2nd and 3rd classes) clades. Small circles in the 1st class clades are inferred missing haplotypes that were not observed in the dataset. The haplotypes or clades observed in the east Kra Isthmus are presented in white and those observed in west Kra Isthmus are black. The size of circles represents the sample size of clades.
In contrast, the inferences for the total cladograms generated using the great circle and sea route distances distinctly differ. The inference for the great-circle-distance based results was that there was “inadequate sampling” to discriminate IBD from LDD. However, the sea-route-distance-based results indicated a restricted gene flow with IBD scenario. The inferences for the clades with significant haplotype versus geographical distance associations obtained from the NCA based on the two types of geographic distance are summarized in Table 2-5.
Discussion
Land barrier of the Kra Isthmus
The Malay Peninsula, and previously ancient Sundaland, is commonly considered to be a barrier for marine and coastal organisms between the Gulf of Thailand of the western Pacific Ocean and the Bay of Bengal of the eastern Indian Ocean (Benzie 1999; Imron et al. 2007; Su et al. 2006; Tan et al. 2005). However, both
oceanographic data (Woodruff 2003) and biogeographical information on the isolation of populations of the giant freshwater prawn Macrobrachium rosenbergii between northern and southern sites of the Kra isthmus imply that the Gulf of Thailand and the Bay of Bengal have been connected in the past (de Bruyn et al. 2005). Therefore, we can hypothesize that the divergence of these populations may have occurred after the formation of the isthmus, where the Malay Peninsula is thinnest.
According to Wolfe et al’s (1987) estimation of the evolutionary rate (1.0-3.0×10-9 per site per year) of chloroplast DNA, the divergence time between east and west Kra Isthmus populations was about 4.59-1.53 Mya, which roughly matches the estimated time of formation of the Malay Peninsula land barrier after the last marine transgressions during the Pliocene era (5.5-4.5 Mya; Woodruff 2003). Although the margin of error for dating due to the congenital variation of the markers is large, the consistency between the estimated time of population differentiation and the vicariant event indicates that the geographic and genetic divergence of C.
tagal were associated. The geographic isolation imposed by Kra Isthmus
contributes about one third of the present variation of the C. tagal populations.
Genetic variation amounting to ca. a quarter of the total variation detected has accumulated among populations since the Pliocene when the land barrier separating populations along the South China Sea and Bay of Bengal coasts emerged (Table 2-3).
The inferences regarding divergence drawn from the molecular data are also supported by the low gene flow (Nm) values between populations on the east and west sides of the Kra Isthmus. Thus, our data support hypothesis that the Kra Isthmus (or Malay Peninsula) has provided a vicariant barrier for marine and coastal organisms.
The haplotypes located at the centre of a network are usually considered most ancestral. Thus, clades II-3 and II-10, which are located at the centre of the network we derived may include the most ancient haplotypes of the east and west Kra Isthmus populations, and may carry the maximum amount of genetic information regarding the ancestral populations of C. tagal ~5 Mya, although their haplotypes have evolved and accumulated new variations since then. In addition, the star-like topology with short interior branches leading to long terminal clades with missing-links is indicative of exponential growth events (Anderson et al. 2003). Populations of other organisms in the Indo-Malay regions also seem to have rapidly expanded at a similar time, e.g.
populations of the mollusc Haliotis asinina (Imron et al. 2007).
Comparison of the two geographic-distance based NCAs
The two geographic-distance based NCAs gave the same inferences for the lower class network but differ in inferences obtained for the total cladograms, probably because in our network the lower class clades almost exclusively comprises populations of the same side of the Kra Isthmus, between which the great-circle and sea-route distances are very similar. However, for the highest clade there may be different implications due to the substantial differences between the great-circle and sea-route distances separating the east and west Kra Isthmus populations. The great-circle-distance based analysis failed to discriminate between IBD (short distance dispersal) and LDD. This may be due to the short great-circle distances between populations on the two sides of Kra Isthmus and between populations on the same sides. Thus, cross-isthmus dispersal of propagules cannot be distinguished from around-isthmus dispersal using great-circle distances. However, using the more realistic sea route-based geographic distances, a restricted gene flow with IBD scenario was inferred due to the absence of tight connections between haplotypes
representing populations from different sides of the Kra Isthmus within all but two clades (Clades II-5 and II-7). This pattern excludes the possibility of exchange of propagules across the Kra Isthmus mediated by changes in stream drainage patterns or sea-level rises. Indications of migration around the isthmus, i.e. via the sea-route in the 5 million years since the Malay Peninsula emerged, are also provided by the pairwise Nm values derived from the FST data (Table 2-4). Since different inferences can be drawn from analyses using the two types of distances, it has been proposed that both types should be used to obtain as much information as possible on historical and contemporary processes (Fetzner and Crandall 2003). For the clades we obtained only the inferred modes (LDD or SDD) of dispersal differed when great-circle and sea-route distances were used. Therefore, we can conclude (a) that dispersal events have occurred not only between populations located on the same side of the Kra Isthmus but also between populations on different sides of the isthmus, (b) that between-side dispersal has been far less frequent than within-side dispersal, and (c) that since its emergence the Kra Isthmus has played an important role in differentiating and structuring the C. tagal populations. According to the result of network, there are two malpositions in subclades II-5 and II-7.
Potential means of dispersal across the Kra Isthmus
Dates based on the estimated evolutionary rate of chloroplast DNA indicate that the contiguous range expansion of subclade II-7 (III-3) occurred ca. 7.04-2.35 Mya and the LDD of subclade II-5 occurred 0.49-0.17 Mya. Thus, the estimated time when contiguous range expansion of III-3 occurred matches the time when the Kra Isthmus is believed to have formed, indicating that the haplotype exchanges involved may have occurred before the isthmus formed. In other words, the NCA inferred that rapid range expansion of C. tagal approximately 5 Mya and the rising sea levels promoted frequent gene flow between mangrove populations situated on the South China Sea and Bay of Bengal coasts at this time. However, long distance dispersal events also seem to have occurred less than one million years before present, indicating that propagules have had opportunities to disperse across the Kra Isthmus either via sea routes or land bridges.
There are two possible ways in which C. tagal propagules may have dispersed from one side of the Kra Isthmus to the other. The first is directly, across the isthmus, perhaps through leaks across the Kra Isthmus during interglacial periods or through
human-assisted introductions. However, there is no evidence that sea levels have risen by more than 20 meters higher than present-day levels in the past 0.5 million years (Rohling et al. 1998): far less than the 100 meters required for such leaks.
Human-mediated dispersal also seems to have been unlikely because ancient
“Heidelberg Man” (Homo heidelbergensis) and/or relatives of H. heidelbergensis living in the area roughly 0.2 to 0.6 Mya had no apparent uses for mangrove seedlings or reasons to help them to disperse. Furthermore, although we cannot exclude the possibility of animal-mediated dispersal, there is no evidence for such dispersal. We therefore conclude that human- or animal-mediated LDD is unlikely to account for the apparent cross-isthmus gene flow we detected.
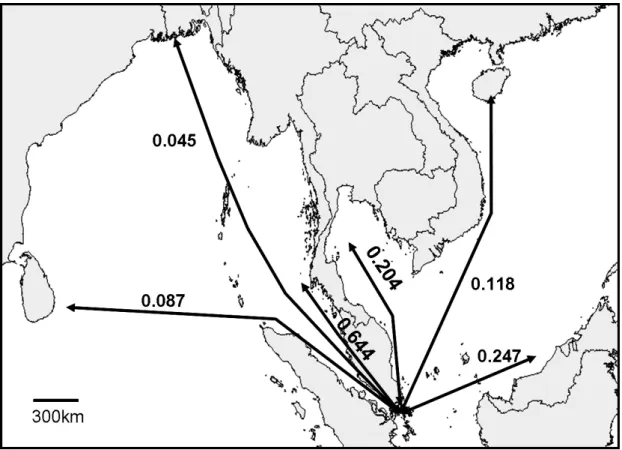
The second way in which dispersal from one side of the isthmus to the other may have occurred is by long-distance dispersal via the sea route around the Malay Peninsula. If dispersal via sea-routes has occurred, trans-sea dispersal through the Malacca Strait is the likeliest route. This hypothesis is indirectly supported by the higher Nm values obtained for gene flow through the population of the cape of Malay Peninsula between populations of the eastern and western sides of the Kra Isthmus than between the others of the South China Sea and the Bay of Bengal (Fig. 2-5, unpublished data). Straits are generally considered to provide corridors for species in the waters they connect. The Strait of Gibraltar, and the passage of organisms from the Atlantic to the Mediterranean and vice versa is often considered in studies of these phenomena (Betzler et al. 2006; Gonzalez-Wanguemert et al. 2006; Seidenkrantz et al.
2000). Similarly to the Strait of Gibraltar, the Malacca Strait plays an important role in the passage of marine organisms (and merchant vessels) between the Indian Ocean and the South China Sea. However, the two sea areas may be alternately isolated during glacial periods, when the Sunda Shelf may emerge above sea level, and reconnected during interglacial periods. The likely dates for the LDD event apparently associated with subclade II-5 of C. tagal range from 0.49-0.17 Mya, including both the last part of the Günz-Mindel (approximately 0.62-0.45 Mya) and all of the Mindel-Riss (approximately 0.3-0.2 Mya) interglacial eras. Thus, despite the large uncertainty of the molecular dating, it indicates that interglacial sea-level rises probably facilitated the trans-isthmus gene flow of mangroves.
Fig. 2-5 Values of gene flow (Nm) calculated from the cape of Malay Peninsula (Johor) to areas of the South China Sea and the Bay of Bengal.
Irrespective of the routes involved, evidence of a few LDD events has been detected, which may have hindered allopatric speciation. The unresolved nature of the haplotype tree we obtained, which makes it difficult to coalesce the time of divergence between haplotypes of the two coasts, also indicates there has been recent contact between populations on the two coasts (Fig. 2-3). This situation is very similar to that of some Baja California marine fishes (Halichoeres semicinctus, Semicossyphus pulcher, Hermosilla azurea, and Sebastes macdonaldi) described by Bernardi et al. (2003), who suggested that the lack of resolution of tree topologies they obtained was probably due to recent or ongoing dispersal with high levels of gene flow, although they could not exclude the possibility that there had been insufficient time for lineage sorting if the markers they used had evolved slowly. Thus our findings and those of the cited authors indicate that young isthmuses such as Baja California and the Kra Isthmus form barriers that inhibit migration and dispersal, but do not entirely prevent frequency LDD, and thus connections are maintained between allopatrically distributed populations in the seas they separate.
Conclusions
Our results are compatible with a vicariant scenario in which there have been few opportunities for LDD between C. tagal populations on eastern and western sides of the Kra Isthmus. The formation of land barriers is generally considered to be a common cause of allopatric speciation of marine organisms. Although ancient Sundaland and, recently, the Malay Peninsula have prevented gene exchange between South China Sea populations and Bay of Bengal populations of many organisms (Heatwole et al. 2005; Karns et al. 2000; Lessios et al. 2001; Su et al. 2006; Tan et al.
2005), leaks at points such as the Malacca Strait during interglacial periods may provide gateways for LDD of these organisms. For the C. tagal populations along the two coasts of the Kra Isthmus examined here, although genetic differentiation was detected, the differences have accumulated in the less than approximately five million year period since the formation of the Kra Isthmus, and they may have progressively homogenized by LDD around the Malay Peninsula. Such homogenization is natural, but may be accelerated by afforestation programs designed to restore populations of the species.