中國醫藥大學機構典藏 China Medical University Repository, Taiwan:Item 310903500/32506
72
0
0
全文
(2) 目錄. 目錄.................................................................................................................................. i 圖目錄............................................................................................................................iii 表目錄............................................................................................................................ iv 中文摘要......................................................................................................................... v 英文摘要........................................................................................................................ vi 第一部分......................................................................................................................... 1 前言 ......................................................................................................................... 1 文獻回顧 ................................................................................................................. 2 一. 動脈粥狀硬化 (Atherosclerosis)................................................................. 2 1. 動脈粥狀硬化的成因................................................................................ 2 2. 細胞黏著分子與動脈粥狀硬化的關係:白血球徵募 (leukocyte recruitment)… .......................................................................................... 4 3. 腫瘤壞死因子 (tumor necrosis factor-α,TNF-α) ................................ 14 二. Heme Oxygenase-1 (HO-1)......................................................................... 15 1. HO-1 介紹................................................................................................ 15 2. HO-1 的生理角色.................................................................................... 16 3. HO-1 代謝產物對細胞的保護作用........................................................ 17 3-1 Biliverdin and bilirubin..................................................................... 17 3-2 Carbon monoxide (CO).................................................................... 17 3-3 Fe2+ and ferritin ................................................................................ 18 4. 調控HO-1 基因表現的訊息傳遞路徑.................................................... 19 5. 參與HO-1 表現的轉錄因子................................................................... 20 三. 穿心蓮 ........................................................................................................ 24 1. 簡介......................................................................................................... 24 2. 生理功效................................................................................................. 26 2-1 抗發炎作用 ...................................................................................... 26 2-2 抗癌作用 .......................................................................................... 26 2-3 肝臟保護作用 .................................................................................. 27 3. 使用風險................................................................................................. 28 研究目的 ............................................................................................................... 29 文獻參考 ............................................................................................................... 30 第二部分....................................................................................................................... 39 Induction of Heme Oxygenase 1 and Inhibition of Tumor Necrosis Factor-α-Induced Intercellular Adhesion Molecule Expression by i.
(3) Andrographolide in EA.hy926 Cells ..................................................................... 39 參考文獻 ............................................................................................................... 61 附錄 ............................................................................................................................. 65. ii.
(4) 圖目錄 第一部分 圖 2-1. 動脈粥狀硬化形成過程 ................................................................................... 3 圖 2-2. 血管腔內動脈粥狀硬化進展剖面圖 ............................................................... 3 圖 2-3. 白血球黏附作用的連續步驟 ........................................................................... 4 圖 2-4. 參與白血球黏附作用的機制 ........................................................................... 6 圖 2-5. 細胞激素調控 ICAM-1 的訊息傳遞路徑 ..................................................... 11 圖 2-6. 血基質分解路徑 ............................................................................................. 15 圖 2-7. 調控 HO-1 表現的轉錄因子及其上游激酶.................................................. 20 圖 2-8. Nrf2/ARE 的活化路徑 ................................................................................... 23 圖 2-9. Nrf2 的調控網路 ............................................................................................ 23 圖 2-10. 穿心蓮及穿心蓮內酯結構............................................................................ 25. 第二部分 Fig. 1. Chemical structure of andrographolide. ............................................................ 53 Fig. 2. Effect of andrographolide (AP) on the cell viability of EA.hy926 cells in the presence of TNF-α. ........................................................................................... 53 Fig. 3. Andrographolide (AP) inhibits TNF-α-induced ICAM-1 expression in EA.hy926 cells….............................................................................................. 55 Fig. 4. Effect of andrographolide (AP) on TNF-α-induced HL-60 cell adhesion. ....... 56 Fig. 5. Andrographolide (AP) induces HO-1 expression in EA.hy926 cell. ................ 57 Fig. 6. Effect of andrographolide (AP) on Nrf2 nuclear translocation and ARE-luciferase reporter activity. ...................................................................... 58 Fig. 7. Effect of HO-1 siRNA on the inhibition of ICAM-1 expression by andrographolide (AP). ...................................................................................... 59 Fig. 8. Model showing pathways that mediate the inhibition of expression of ICAM-1 and HL-60 cell adhesion to EA.hy926 cells by andrographolide under inflammatory conditions. ...................................................................................60. iii.
(5) 表目錄 第一部分 表 2-1. 表 2-2. 表 2-3. 表 2-4. 表 2-5. 表 2-6.. 黏著分子參與動脈粥狀硬化的白血球徵募作用 ........................................... 9 黏著分子及其配體 ......................................................................................... 10 不同類型的細胞可受到不同刺激而引發 ICAM-1 的表現 ......................... 11 整合素和其配體 ............................................................................................. 13 細胞激素誘發細胞黏著分子(CAM)的表現 ................................................. 14 穿心蓮的傳統醫療作用 ................................................................................. 25. iv.
(6) 中文摘要. 穿心蓮內酯(Andrographolide)是穿心蓮中含量最多的二帖類,穿心蓮是東南亞 國家廣為流傳的傳統中草藥,常用於治療感染、發炎、感冒、發燒和腹瀉。血基 質氧化酶 Heme oxygenase (HO)-1 是一個壓力反應的基因,也是抗氧化酵素的一 種。過去文獻指出 HO-1 會藉由產生 bilirubin 和減少細胞內游離鐵而抑制內皮細 胞黏著分子表現。ICAM-1 是一種發炎生理指標,會使單核球黏附至內皮細胞上 進而滲入細胞。在本研究中,我們探討在 EA.hy926 內皮細胞中,穿心蓮內酯對 TNF-α 誘發 ICAM-1 表現的影響以及可能參與的機制。隨著處理劑量增加(0-7.5 μM),穿心蓮內酯抑制 TNF-α 所誘發的 ICAM-1 表現及內皮細胞與白血球的黏附 作用,具有顯著的抑制效果(P<0.05)。同時,處理穿心蓮內酯誘發 HO-1 表現呈現 劑量效應。穿心蓮內酯會增加轉錄因子 Nrf2 的核轉移作用和誘導 antioxidant response element (ARE)-luciferase reporter 活性。利用轉染 HO-1 specific small interfering RNA (siRNA)壓制細胞中 HO-1 表現,顯著破壞穿心蓮內酯抑制 ICAM-1 的表現。這些結果顯示,穿心蓮內酯藉由增加 Nrf2 細胞核轉移、誘發 ARE-luciferase reporter 活性而刺激 HO-1 表達,進而抑制 TNF-α 所誘發的 ICAM-1 表現。. 關鍵詞:穿心蓮內酯‧EA.hy926 cells‧血基質氧化酶-1‧細胞間黏著分子-1‧腫瘤壞 死因子. v.
(7) 英文摘要. Andrographolide is the most abundant diterpene lactone in Andrographis paniculata, which is widely used as a traditional medicine in Southeast Asia. Heme oxygenase 1 (HO-1) is an antioxidant enzyme encoded by a stress-responsive gene. HO-1 has been reported to inhibit the expression of adhesion molecules in vascular endothelial cells (EC). Intercellular adhesion molecule (ICAM-1) is an inflammatory biomarker that is involved in the adhesion of monocytes to EC. In this study, we investigated the effect of andrographolide on the expression of ICAM-1 induced by tumor necrosis factor R (TNF-α) in EA.hy926 cells and the possible mechanisms involved. Andrographolide (2.5-7.5 μM) inhibited the TNF-α-induced expression of ICAM-1 in a dose-dependent manner and resulted in a decrease in HL-60 cell adhesion to EA.hy926 cells (p < 0.05). In parallel, andrographolide significantly induced the expression of HO-1 in a concentration-dependent fashion (p < 0.05). Andrographolide increased the rate of nuclear translocation of nuclear factor erythroid 2-related 2 (Nrf2) and induced antioxidant response element-luciferase reporter activity. Transfection with HO-1-specific small interfering RNA knocked down HO-1 expression, and the inhibition of expression of ICAM-1 by andrographolide was significantly reversed. These results suggest that stimulation of Nrf2-dependent HO-1 expression is involved in the suppression of TNF-α-induced ICAM-1 expression exerted by andrographolide.. Key words : Andrographolide; EA.hy926 cells; heme oxygenase 1; ICAM-1; TNF-α. vi.
(8) 第一部分 前言. 根據衛生署統計,心臟疾病與腦血管疾病一直高居國內十大死因的第 二、三名,心血管疾病本身就是一種發炎反應,動脈粥狀硬化更是導致心血 管疾病的重要病因,黏著分子的表現對於動脈硬化疾病的發展扮演重要角 色,因此了解動脈粥狀硬化形成並加以預防是目前預防醫學重要的課題之 一。在開發中或已開發中國家使用傳統中草藥的人口比例或市場金額都逐年 上升,加上現代人注重健康養生,認為中草藥應用在膳食和疾病療養上會有 不錯效果,因此,中草藥在飲食及醫學領域也逐漸受到西方國家的重視。 穿心蓮,拉丁學名為 Andrographis paniculata (Burm. F.) Nees,主要分 布於東南亞熱帶地區,而中國大陸與台灣各地也有零星栽培,穿心蓮味苦性 寒,具清熱解毒、涼血及消腫止痛之效,過去研究發現,穿心蓮具有廣泛的 生理功能,包括抗發炎、抗腹瀉、抗病毒、保護肝臟、抗癌等。因此本研究 將探討穿心蓮對於內皮細胞的抗發炎效果及其機制。. 1.
(9) 文獻回顧. 一. 動脈粥狀硬化 (Atherosclerosis) 1.動脈粥狀硬化的成因 心臟疾病與腦血管疾病一直高居國內十大死因的第二、三名,根據衛生署統 計,九十八年的心臟與腦血管疾病分別占所有死亡原因的 10.6%及 7.3% (行政院 衛生署, 2010)。動脈粥狀硬化是一種慢性發炎疾病,會造成血管內壁脂肪堆積及 纖維斑塊形成,是導致心血管疾病的主要原因之一(de Winther et al., 2005)。Ross (1999) 提出動脈粥狀硬化在致病初期血管內壁的內皮細胞有受損現象,造成內皮 細胞受損的原因包括: 抽菸、高血壓、高膽固醇飲食、糖尿病等。動脈粥狀硬化 的致病機轉十分複雜,主要是因為血漿中低密度脂蛋白(low-density lipoprotein, LDL)濃度的增加,高低密度脂蛋白濃度會增加轉變成氧化型低密度脂蛋白 (oxidized-low-density lipoprotein, ox-LDL)的機會,當血管壁內層長期受到大量的 氧化型低密度脂蛋白刺激時,會使內皮細胞的通透性增加,促進前發炎細胞激素或 趨化因子(chemokines)的活化與黏著分子表現,進而吸引循環中的白血球包括單 核球黏附至內皮細胞,單核球會進一步穿透血管壁,增加對ox-LDL的攝入 (uptake) 而分化成巨噬細胞,此階段稱為血管內膜發炎。接著,巨噬細胞會吞噬大量的 ox-LDL,形成泡沫細胞(foam cell),進而在血管內膜形成脂肪紋(fatty streak),這 種富含脂質的脂肪紋除了巨噬細胞也含有膠原蛋白及蛋白多醣,這些脂肪條紋會 緩慢的形成纖維斑塊(fibrous plaque)。纖維斑塊會分泌生長因子及細胞激素使得 斑塊增厚,造成斑塊表面纖維化,形成更緊密的粥狀物,而內部堆積更多壞死的 細胞與脂質,進而使斑塊往周圍蔓延。動脈粥狀硬化晚期,纖維性的胞外間質被 分解導致斑塊不穩定而破裂,形成血栓,增加心肌梗塞或是腦中風發生率 (de Winther et al., 2005; Dzau et al., 2002)。. 2.
(10) 圖 2-1. 動脈粥狀硬化形成過程 (de Winther et al., 2005). 圖 2-2. 血管腔內動脈粥狀硬化進展剖面圖 (Rader and Daugherty, 2008). 3.
(11) 2. 細胞黏著分子與動脈粥狀硬化的關係:白血球徵募 (leukocyte recruitment) 細胞黏著分子表現與動脈粥狀硬化形成初期關係密切,動脈粥狀硬化的早期 階段涉及到發炎細胞會從血液循環中徵募(recruitment)並遷移至內皮細胞發炎 處。此現象是經由一連串的白血球與血管壁內皮細胞黏附作用所調控,主要包括 下列步驟(圖 2-3):(1)滾動(rolling):當組織受到損害時,會大量表現選擇素 (selectin),與循環中的白血球產生交互作用,白血球進而在血管壁上滾動,這項 作用是由白血球的 L-selectin 和血管內皮細胞上的 P-selectin、E-selectin 所調控; (2)黏著(adhesion):受傷的組織會釋放趨化因子來活化滾動中的白血球,使它們 停止滾動並黏附至內皮細胞上,此階段的血管內皮細胞黏著分子(ICAM-1、 VCAM-1)會受到細胞激素(如: TNF-α)的活化而增加其表現,細胞激素同時也會活 化 integrin,活化的 integrin (β1、β2)則會增強白血球與血管內皮細胞結合的力量; (3)遷移(transmigration):白血球黏附至內皮細胞後會開始變形,在這個步驟中, 參與的黏著分子 PECAM-1 會使白血球的型態產生改變進而穿透血管內皮細胞 層,從血管內遷移至血管外的發炎組織處,促使後續動脈粥狀硬化的形成 (Blankenberg et al., 2003;Price and Loscalzo, 1999)。. 圖 2-3. 白血球黏附作用的連續步驟 (Blankenberg et al., 2003). 4.
(12) 2-1 細胞黏著分子 在動脈粥狀硬化初期,研究發現在細胞黏附的過程中,有許多細胞表面分子 參與,表2-1為各種黏著分子參與動脈粥狀硬化初期的過程(Galkina and Ley, 2007)。根據 Gene Ontology Consortium定義,細胞黏著分子為表現於細胞表面之 分子且能使細胞黏著到其他細胞或胞外基質的分子,這些細胞黏著分子(cellular adhesion molecules, CAMs)依其不同的結構特性可分為以下三大類(表2-2):分別 為選擇素(selectins)、類免疫球蛋白分子(immunoglobulin-like molecules)及整合素 (integrins) (Blankenberg et al., 2003;Danese et al., 2005)。. 2-1-1 選擇素(selectin) 選擇素是一個含有三個不同醣蛋白(glycoproteins)的結構,其蛋白的 N 端有一 個類似 lectin 的區域,在血管層內扮演調控細胞和細胞間黏附作用的角色,主要 有三個成員,分別以發現細胞的位置來命名,例如表現在白血球表面的 L-selectin (CD62L)、表現於內皮細胞的 E-selectin (CD62E)及儲存在血小板內的 P- selectin (CD62P)(Blankenberg et al., 2003)。 L-selectin 表現在所有的白血球細胞,文獻指出若淋巴球上的 L-selectin 產生 缺陷,會減少 50%淋巴球在主動脈上的移行作用,另外,淋巴球移行到主動脈的 粥狀硬化處也受到 L-selectin 的調控(Galkina et al., 2006)。L-selectin 的配體(ligands) 是含有碳水化合物的分子,PSGL-1 是 L-selectin 的配體之一,當 PSGL-1 與 L-selectin 結合時,會讓循環中的白血球與在內皮細胞上滾動的白血球產生交互作 用,造成 secondary capture (圖 2-4)。因此,研究認為在白血球徵募到動脈血管內 皮發炎處可能是透過 L-selectin 和 PSGL-1 的交互作用產生 secondary capture 來達 成(Eriksson, 2001)。. 5.
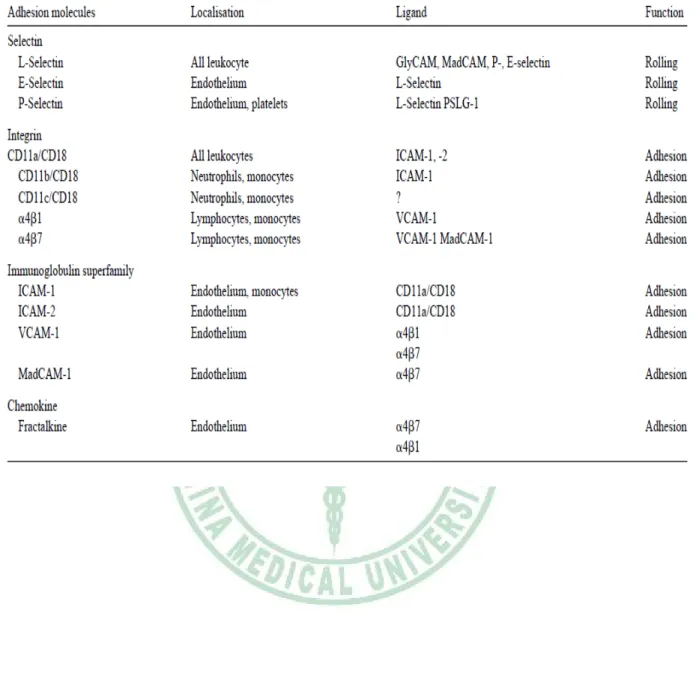
(13) 圖 2-4. 參與白血球黏附作用的機制 (Eriksson, 2001). E-selectin 只表現在內皮細胞,當內皮細胞受到發炎激素刺激時,例如: tumor necrosis factor (TNF)-α、interleukin (IL)-1 或 platelet factor 4 (PF4),則會誘發 E-selectin 表現,人類動脈粥狀硬化的內皮細胞處,也會表現 E-selectin (Yu et al., 2005)。 P-selectin主要儲存於血小板內,在生物體中,動脈粥狀硬化的斑塊內可以檢 測到P-selectin (Johnson et al., 1994)。為了瞭解P-selectin在白血球徵募至動脈粥狀 硬化過程中所扮演的角色,利用頸動脈的體外模式,在 knockdown P-selectin或 PSGL-1 的 Apoe-/-小鼠模式中發現,單核球滾動和黏附至頸動脈內皮細胞的速度 明顯降低。因此推論在單核球的黏附過程中,P-selectin及PSGL-1 的相互作用扮 演重要的角色(Ramos et al., 1999)。. 2-1-2 類免疫球蛋白分子 (Immunoglobulin-like molecules) Immunoglobulin (Ig) superfamily 細胞黏著分子是表現於細胞表面的醣蛋白, 包括 ICAM-1, ICAM-2, VCAM-1, platelet–endothelial cell adhesion molecule (PECAM)-1 and the mucosal addressin MAdCAM-1(表 2-2)。. 6.
(14) Intercellular adhesion molecules (ICAMs)為此superfamily中的一員,ICAMs含有 五個成員,ICAM-1 表達在白血球細胞、纖維細胞、上皮細胞及內皮細胞,具有 增加白血球黏附作用,ICAM-2 受到發炎物質的負向調控,表現於白血球、血小 板與內皮細胞,ICAM-3 除了表現在內皮細胞及白血球細胞外,也是ICAM分子中 唯一表現在嗜中性白血球的黏著分子。另外,ICAM-4 表現於紅血球,ICAM-5 則表現在大腦(Briscoe et al., 1992;Blankenberg et al., 2003) 。這些ICAM分子的 配體主要包括白血球特異性β2整合素,每個ICAM分子和ligand間的結合,促進白 血球的黏附作用,ICAM-1、2、3 可加強白血球牢固的黏附於血管表面(Blankenberg et al., 2003)。 ICAM-1 (CD54)是一個跨膜醣蛋白,含有 505 個胺基酸,因不同程度的醣基化 修飾,其分子量介於 80-114 kDa 之間,主要和表現於白血球表面的整合素 LFA-1 結合,ICAM-1 表現於許多細胞表面,包括上皮細胞、角質細胞、肝臟細胞、淋 巴細胞及纖維母細胞,這些細胞平時只會表現少量的 ICAM-1,但是當受到促發 炎細胞激素(TNF-α、IL-1、IL-6 及 interferon-γ 等)、氧化壓力、病毒感染等刺激 後,會大量增加 ICAM-1 的表現(表 2-3) (Roebuck and Finnegan, 1999)。ICAM-1 對於白血球穿透血管內層的移行作用扮演重要角色,同時也與許多發炎疾病發生 有關,除了動脈粥狀硬化外,還包括發炎性腸道疾病(Fiocchi, 1998)及氣喘等疾病 (Stanciu and Djukanovic, 1998)。 人類 ICAM-1 基因位於第十九號染色體上,包括七個 exons 與六個 introns (Voraberger et al., 1991)。ICAM-1 promoter 上有許多特定轉錄因子的結合位,包 括 Sp1、NF-κB、AP-1、AP-2、interferon-stimulated response element (ISRE 或稱 為 STAT),因此 ICAM-1 表現會受到許多轉錄因子的調控 (Roebuck and Finnegan, 1999)。細胞內調控 ICAM-1 基因表現的訊息傳遞路徑包括 MAPK/AP-1 (Karin, 1996)、IKK/NF-κB (Mercurio et al., 1997)、PKC (Sippy etal., 1996)、JAK/STAT (Song et al., 1997; Look et al., 1995)。 以 NF-κB 訊息傳遞路徑為例,TNF-α 和 IL-1β 都會透過活化 NF-κB 來調控 7.
(15) ICAM-1 表現,NF-κB 以 Rel A 的 homodimers 或 heterodimers 形式結合至 NF-κB 的結合區域,啟動 ICAM-1 基因表現(圖 2-5) (Roebuck and Finnegan, 1999)。 Vascular cell adhesion molecule 1 (VCAM-1)是一個分子量 110 Kda的蛋白質,主 要表現在內皮細胞、巨噬細胞、肌肉細胞及樹突狀細胞。VCAM-1 表現於活化的 內皮細胞時,會藉由和白血球表面的整合素α4β1 (very late antigen 4,VLA-4)或α4β7 結合來促使白血球滲出血管壁,特別是單核球、嗜鹼性球、嗜酸性球以及淋巴球 需要有VCAM-1 來產生黏附作用(Zapolska et al., 2001)。 另外,PECAM-1 也是 Ig family 成員之一,主要表現在白血球、血小板與內皮 細胞,PECAM-1 在內皮細胞連接處有密集的表現量,會參與細胞和細胞間的鍵 結,與內皮細胞完整性及白血球滲出血管壁有關(Blankenberg et al., 2003)。. 8.
(16) 表 2-1. 黏著分子參與動脈粥狀硬化的白血球徵募作用 (Galkina and Ley, 2007). Vascular Adhesion Molecules in Atherogenesis Migration Step Tethering,. Adhesion Molecules L-selectin. rolling. Model Sell-/- vs C57BL/6 and Apoe-/lymphocytes (adoptive transfer). P-selectin. C57BL/6 and Apoe-/- mice on western diet. Effect 50% reduction in lymphocyte homing Reduced rolling and attachment with anti-P-selectin or PSGL-1 Abs. E-selectin. Adhesion. VCAM-1. Sele-/- and Apoe-/- double. Slightly reduced. Knockout(DKO) mice. lesion area. C57BL/6 and Apoe-/- mice on western diet. Increased rolling velocity with anti-VLA-4 or VCAM-1 Abs. VLA-4. Apoe-/- mice on western diet. Decrease in neointimal growth and neutrophil and Mψ recruitment with anti-VLA-4 Abs. ICAM-1. Icam1-/- mice (on western diet). Slightly decreased lesionsize compared to C57BL/6 controls. 9.
(17) 表 2-2. 黏著分子及其配體 (Danese et al., 2005). 10.
(18) 表 2-3. 不同類型的細胞可受到不同刺激而引發ICAM-1 的表現 (Roebuck and Finnegan, 1999). 圖 2-5.細胞激素調控ICAM-1 的訊息傳遞路徑(Roebuck and Finnegan, 1999). 11.
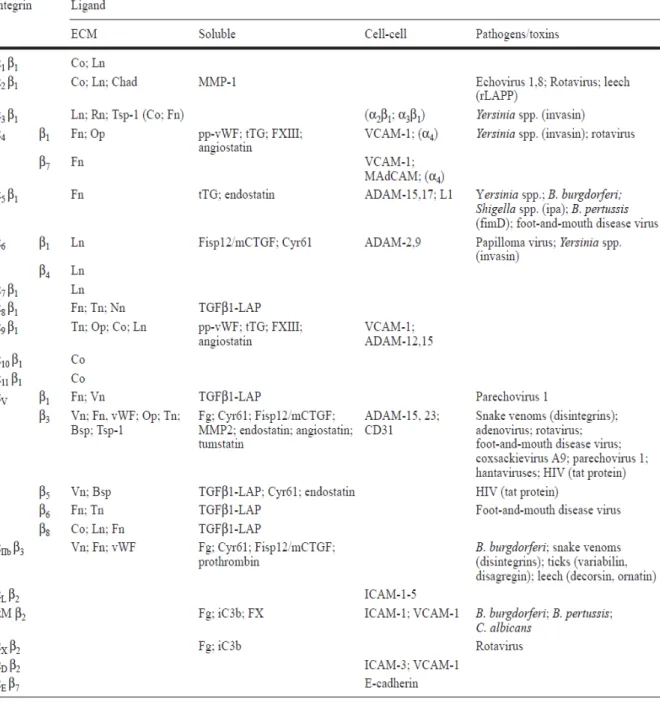
(19) 2-1-3 整合素 (integrins) 整合素家族含有 24 個次家族,為 αβ 異質二聚體 (heterodimer),由 18 種 α 次單元與 8 種 β 次單元所組成,藉由不同的組合配對可得到 24 種的 integrin (表 2-4)(van der Flier and Sonnenberg, 2001),其主要功能為形成細胞間或胞外基 質黏著的橋梁,調控細胞與細胞、細胞與胞外基質及細胞與病原體的接觸(Galkina and Ley, 2007)。LFA-1 (αLβ2) (lymphocyte function-related antigen)是細胞表面抗 原,會表現在所有的白血球,也稱為CD11a/CD18, 屬於整合素的β2次家族;VLA-4 (α4β1) 整合素則是α4的次家族成員,主要表現在單核球及淋巴球,另外,VLA-4 也會與免疫球蛋白家族的VCAM-1 及淋巴球細胞相互作用並且黏附至內皮細胞 發炎處(Danese et al., 2005)。. 12.
(20) 表 2-4. 整合素和其配體 (van der Flier and Sonnenberg, 2001). 13.
(21) 3. 腫瘤壞死因子 (tumor necrosis factor-α,TNF-α) 免疫系統是生物體的防禦系統之一,生物體藉由辨別並殺死病原體及腫瘤細 胞的過程,來保護宿主免於疾病的傷害。發炎現象是血管組織在傷害刺激下(例如, 病原體、受損細胞或刺激物)所產生的一種生物反應(Calder and Kew, 2002)。前發 炎細胞激素(pro-inflammatory cytokines)例如:TNF-α、IL-1α、IL-1β、IFN-γ 及 gram-negative bacterial endotoxins (lipopolysaccharide, LPS)等都已被證實可以刺激 內皮細胞並增加白血球的黏附作用(表 2-5) (Meager, 1999)。TNF-α 屬於 TNF/TNFR cytokine superfamily 的成員,會引發局部發炎反應,活化的巨噬細胞 會分泌 TNF-α,產生活性氧物質(ROS),另外,TNF-α 也會透過活化轉錄因子 NF-κB 刺激內皮細胞,增加 ICAM-1、VCAM-1、MCP-1 等發炎基因的表現,並促使白 血球黏附於內皮細胞上,因此 TNF-α 在動脈粥狀硬化過程中也扮演重要角色 (Meager, 1999)。. 表 2-5. 細胞激素誘發細胞黏著分子(CAM)的表現 (Meager, 1999). 14.
(22) 二. Heme Oxygenase-1 (HO-1). 1. HO-1 介紹 文獻指出血基質氧化酶-1 (HO-1) 對心血管疾病包括動脈粥狀硬化有重要的 影響(Immenschuh and Schroder, 2006)。HO-1為一種具有保護作用的蛋白質,許多 研究證實它在內皮細胞、上皮細胞、平滑肌細胞中扮演抗發炎、抗凋亡、抗增生 的作用(Brouard et al., 2000;Lee et al., 1996;Peyton et al., 2002;Tobiasch et al., 2001)。HO-1是血基質代謝的速率限制酵素(Maines, 2005; Maines and Gibbs, 2005),它將血基質代謝成膽紅素(biliverdin)、一氧化碳 (CO)和游離鐵(free iron) (圖2-6)。HO-1的表現受到氧化壓力的調控,可藉由增加膽紅素和促進CO產生來 達到抗發炎和保護細胞的作用(Ryter et al., 2006)。. 圖2-6. 血基質分解路徑 (Farombi and Surh, 2006). 15.
(23) 哺乳動物的血基質氧化酶(HO)有三種異構物,分別命名為HO-1、HO-2和HO-3 (Maines, 1988; McCoubrey et al., 1997)。HO-1又稱為熱休克蛋白32 (heat-shock protein 32, Hsp 32),人類 HO-1是由288個胺基酸所組成,分子量為32.8 kDa,其 中約80%的胺基酸序列和大鼠 HO-1相同(Yoshida e al., 1988),人類 HO-2分子量 為36 kDa,由316個胺基酸所組成(Ishikawa et al., 1995),HO-3分子量為33 kDa, 研究指出 HO-3對血基質的催化作用不強,且無法受誘發而大量表現(Farombi and Surh., 2006)。相反地,HO-1容易被血基質或其他非血基質物質所誘發,例如:植 化物(phytochemicals),以蘿蔔硫素(sulforaphane)為例,sulforaphane為含硫配醣體 (glucosinolate)之水解物,廣存於十字花科蔬菜,如青花菜、甘藍菜及花椰菜等。 根據文獻指出,在人類血管內皮細胞(human vascular endothelial cell),sulforaphane 會透過誘發HO-1來抑制lipopolysaccharide (LPS)誘發的發炎損傷(Shan et al., 2010)。另外,ultraviolet (UV)、過氧化氫、重金屬以及一氧化氮也會誘發HO-1 ( Motterlini et al., 2000)。. 2. HO-1 的生理角色 透過各種刺激,被誘發的HO-1扮演保護的角色。文獻指出,在誘導肝損傷的 實驗中,大鼠經過缺血再灌注的損傷模式,HO-1的mRNA及蛋白質表現增加,這 和心臟、腎臟及肝臟的器官功能改善具有相關性(Masini et al., 2003)。Pannen等人 (1998)證實HO-1在大鼠出血性休克和復甦的模式中具有保護肝臟的作用。近期研 究發現,誘發HO-1表現可以保護人類肝臟缺氧所帶來的傷害,同時誘發HO-1表 現也被認為可以修復發炎性損傷(Tuzuner et al., 2004)。HO-1在調控肝損傷的角色 也同樣被證實,研究發現向上調控HO-1表現可以挽救小鼠肝損傷所造成的細胞凋 亡(Sass et al., 2003)。另外,HO-1過度表現也會改善肝臟損傷造成的細胞凋亡及 組織發炎反應併發症狀(McCarter et al., 2004),因此調控HO-1表現可以做為肝臟 疾病發炎的治療方式。雖然HO-1對於肝臟保護作用的機制還未完全被釐清,但可 16.
(24) 以確定的是和其代謝產物biliverdin/ bilirubin、CO和ferritin有關。. 3. HO-1 代謝產物對細胞的保護作用. 3-1. Biliverdin and bilirubin Biliverdin、bilirubin和CO是heme的代謝產物,在過去常被認為是沒有用或有 毒的產物,而今它們被認為是重要的內生性抗氧化劑(圖2-6)。在體內及體外試驗 中均已證實biliverdin和bilirubin具有抗氧化的活性(Llesuy and Tomaro, 1994)。最近 研究顯示,biliverdin對大鼠肝臟局部缺血再灌流所造成的傷害具有保護作用 (Fondevila et al., 2004);10 nM bilirubin也被發現可以回復子宮頸癌細胞(HeLa cells)在200 μM過氧化氫誘導下造成的傷害(Baranano et al., 2002)。HO-1和 bilirubin的抗氧化活性在大鼠肝臟暴露於UVA模式中也獲得證實(Ossola and Tomaro, 1998)。另外,在大鼠AH 136B肝癌細胞中,HO-1會透過增加bilirubin濃 度、降低細胞內促氧化劑濃度,而達到抗氧化效果(Tanaka et al., 2003; Fang et al., 2004)。. 3-2 Carbon monoxide (CO) 一氧化碳(CO)是血基質代謝的副產物之一,在組織損傷時會透過誘導HO-1表 現而代謝生成較多的CO,幫助減緩發炎反應。一氧化碳具有抗發炎的保護角色 最初是在急性肺損傷的模式中被發現,另外在小鼠心臟移植的模式中也被證實 (Sato et al., 2001)。文獻指出,一氧化碳在急性和慢性發炎反應中,具有保護的 角色(Moore et al., 2005),一氧化碳可以抑制前發炎反應並提高巨噬細胞的抗發炎 作用(Yachie et al., 1999)。在RAW 264.7鼠科動物的巨噬細胞中,CO被證實會減 弱受lipopolysaccharide (LPS)所誘發的免疫反應,並且可調控大鼠肝臟中誘發型 一氧化氮合成酵素(inducible nitric oxide synthase) (iNOS)表現和一氧化氮(nitric. 17.
(25) oxide) (NO)產生(Sarady et al., 2004) (Sawle et al., 2005)。CO在體內和體外試驗中 也被證實會影響其它細胞的增生作用,例如,CO在體內試驗中會抑制平滑肌細 胞的增生(Otterbein et al., 2003)。大鼠預處理250 ppm的CO 1小時,可以明顯降低 球囊損傷後的內膜增生作用(Gerard et al., 1993)。. 3-2-1 參與一氧化碳(CO)作用的訊息傳遞路徑 有研究發現 guanylyl cyclase-cyclic (c)GMP 或 p38 mitogen-activated protein kinase (MAPK)在 CO 的抗增生及抗凋亡作用中扮演重要角色。舉例來說,在纖維 母細胞和 BTC3 細胞中,藉由誘導 cGMP 的產生來達到 CO 的抗細胞凋亡作用; 相反地,在內皮細胞中 CO 則需透過活化 p38 MAPK,才具有抗細胞凋亡的作用 (Brouard et al., 2000)。文獻指出,CO 抗平滑肌細胞增生作用和抗血小板凝集作用 皆需要 cGMP 和 p38 的參與(Petrache et al., 2000)。CO 會經由活化 p38 和產生 cGMP 的 guanylyl cyclase 來抑制巨噬細胞 IL-10 和 TNF-α 的生成(Yachie et al., 1999)。在一些含有血基質的蛋白質中,有可能透過 CO 來調控訊息傳遞,例如: 血紅素(hemoglobin)、肌紅蛋白(myoglobin)、鳥苷酸環化酶(guanylyl cyclase)、環 氧合酶(cyclooxygenase)、cytochrome P450 oxidase、inducible nitric oxide synthase (iNOS)等 (Otterbein et al., 2003)。CO 會透過與血基質內部構造中的鐵結合,改變 這些蛋白質的構形而調控其生理活性,例如,當 CO 與 iNOS 結合時,iNOS 的活 性會降低。CO 也會和含有其他金屬離子的蛋白質例如:Zinc(鋅)、Copper(銅)、 Manganese(錳)產生結合作用(Otterbein et al., 2003)。. 3-3 Fe2+ and ferritin 血基質經HO-1催化產生游離鐵,一般來說,Fe2+在低濃度時會被羥自由基 (hydroxyl radical,.OH)催化而對細胞產生毒性,但有研究顯示,當HO-1被誘發時, 鐵蛋白(ferritin)濃度和Fe2+-sequestering protein量會被提升;而當HO-1活性被抑制 時,則會減少(Eisenstein et al., 1991)。雖然誘發HO-1而產生ferritin對細胞保護作 18.
(26) 用這部分仍有爭議,但最近有些文獻對ferritin和HO-1間的關係皆持認同看法,例 如:過氧化氫會對人類內皮細胞(ECV 304)造成毒性,但它也會誘發HO-1和ferritin 合成(Grosser et al., 2004);Gonzales等人(2005)研究證實在大鼠肝臟中鈷氯化物 (cobalt chloride)誘發HO-1表現後會增加鐵蛋白和含鐵儲鐵蛋白的濃度;另外,也 有文獻證實在非酒精性脂肪肝病症中,HO-1增加和鐵蛋白的濃度有顯著相關性 (Malaguanera et al., 2005)。. 4. 調控 HO-1 基因表現的訊息傳遞路徑 胞內調控HO-1基因表現的訊息傳遞路徑如圖2-7所示,例如 mitogen activated protein kinases (MAPKs)、phosphatidyl inositol 3-kinase PI3K/Akt pathway及 protein kinase C(PKC)皆被認為與調控HO-1基因表現有關。MAPKs pathways包含 extracellular signal-regulated kinase 1/2 (ERK1/2)、c-Jun-N-terminal kinase (JNK)和 p38,它們會透過磷酸化下游轉錄因子而影響標的基因表現。亞砷酸鈉在肝癌細 胞株中會透過ERK1/2和p38訊息傳遞路徑來誘發HO-1基因轉錄(Elbirt et al., 1998)。最近研究證實當angiotensin II誘發HO-1表現及增加CO釋放時,PKC參與 angiotensin II (Ang II)調控HO-1基因表現(Li et al., 2004)。此外與HO-1表現有關的 訊息傳遞路徑還包括protein kinase A (PKA),舉例來說,賀爾蒙和外在刺激會增 加胞內cAMP濃度進而活化PKA,Immenschuh等人(1998)研究發現,大鼠初代肝 細胞給予PKA活化劑,例如Bt2cAMP和glucagon會誘發HO-1表現,並呈現劑量關 係。另外,活性氧分子被證實會透過PI3K/Akt訊息傳遞路徑來誘發大鼠腎上腺嗜 鉻細胞(pheochromocytoma 12 cell ) HO-1表現(Salinas et al., 2003)。另外也有研究 指出,在人類乳腺癌細胞(MCF-7)中,15-deoxy-Δ12,14-prostaglandin J2 (15d-PGJ2) 會透過PI3K/Akt訊息傳遞路徑來誘發HO-1基因表現(Kim et al., 2004)。在人類神經 母細胞(SH-SY5Y)實驗中證實,HO-1基因表現是透過PI3K調控nuclear factor E2-related factor-2 (Nrf2)來達成(Nakaso et al., 2003)。. 19.
(27) 5. 參與 HO-1 表現的轉錄因子 目前文獻已證實activator protein (AP)-1、NF-κB、Nrf2等轉錄因子會參與調控 HO-1基因表現(Farombi and Surh, 2006)。. 圖2-7. 調控HO-1表現的轉錄因子及其上游激酶 (Farombi and Surh, 2006). * Activator protein (AP)-1 AP-1是一個由fos、Jun或ATF家族所組成的homodimer或heterodimer,藉由和 基因啟動區結合啟動基因轉錄作用。透過活化AP-1可提升HO-1表現,細胞激素、 生長因子、氧化壓力及細菌和病毒感染等外來刺激會活化AP-1進而刺激基因表 現,AP-1同時也參與細胞分化、增生和凋亡過程(Karin et al., 1997)。有研究證實 在外來刺激下,例如:血基質、亞砷酸鈉、氯化鈷和cobalt protoporphyrin (CoPP), 它們誘發 HO-1表現是透過AP-1轉錄因子(Shan et al., 2004) (Lu et al., 2000) (Shan et al., 2004)。. * Nuclear factor E2-related factor-2 (Nrf2) 20.
(28) Nrf2是一個分子量66-kDa的蛋白質,屬於 basic region-leucine zipper (bZIP)家 族的轉錄因子之一,對活化帶有 antioxidant response element (ARE) 的基因表現 扮演調控者的角色(Mann et al., 2007) (圖2-8)。 ARE 位於許多phase II 解毒酵素(phase II detoxifying enzymes) 與抗氧化酵 素的promoter中,例如: HO-1、NQO1、glutathione S-transferase (GST)。Nrf2有六 個 domains,從Neh 1到 Neh 6 (Nrf2-ECH homology),在C端終點,Nrf2有一段 basic leucine zipper structure;在N端終點,Nrf2有一個高活化區的domain,Neh 2, Keap-1和Nrf2結合時,是結合至Nrf2上的Neh 2 domain (Itoh et al., 2004) (圖2-9)。 在C端位置上的basic leucine zipper(bZip),bZip區域中的basic region主要負責與 DNA結合,而leucine zipper則是會與其它的bZip轉錄因子結合形成dimmer。Nrf2 調控基因轉錄時必須先和其它的 bZip蛋白質包括Jun (c-Jun、Jun-D、Jun-B) 及 small Maf (MafG、MafK、MafF)等,形成 heterodimerization (Kaspar et al., 2009)。 在無刺激條件下,Nrf2和抑制蛋白 Kelch-like ECH-associated protein-1 (Keap-1) 結合形成複合體,存在細胞質中,呈不活化狀態,當細胞受到氧化壓力刺激時, 會促使 Nrf2與 Keap-1分離,此時Nrf2轉移至細胞核內並且結合到含有ARE的基 因序列上,進而啟動目標基因的轉錄作用,例如: HO-1 (Itoh et al., 2003)。文獻指 出 Nrf2的核轉錄作用會受到 mitogen activated protein kinases (e.g., ERK1/2, JNK, p38 MAPK)、PI3K或 protein kinase C的磷酸化作用所調控(Huang et al., 2002)。以 人類主動脈內皮細胞(human aortic endothelial cells, HAECs)為實驗模式,當Nrf2 在細胞內的蛋白質及mRNA表現增加,可抑制促發炎細胞激素TNF-α誘發p38 MAPK的磷酸化、單核球黏附作用與發炎基因,例如 VCAM-1及MCP-1的表現, 進而達到預防發炎疾病發生的作用(Chen et al., 2006)。 Keap-1是一個含有624個胺基酸的多胜肽,具有五個domains: (1) the N-terminal region (NTR); (2) the BTB domain; (3) the intervening region (IVR); (4) the double glycine repeat (DGR) or Kelch domain; and (5) the C-terminal region (CTR) (Itoh et al., 2004)。Nrf2的Neh 2 domain會跟Keap-1上的DGR domain結合,當Keap-1 21.
(29) 與Nrf2結合時會抑制Nrf2的活化,將Nrf2隔離在細胞質內(Zipper and Mulcahy., 2002)。Keap-1是Nrf2的抑制蛋白,在無刺激環境下,Keap-1會與Nrf2結合並增加 Nrf2的蛋白質降解作用(McMahon et al., 2003)。有文獻指出Keap-1含有好幾個具 有高反應性的cysteine residues,在活化ARE基因序列的過程中,會包含一個具有 高反應性的cysteine residues之胞內氧化還原接受器(redox receptor),此氧化還原接 受器可能是Keap-1 (Dhakshinamoorthy et al., 2001)。有研究發現,在Keap-1上有四 個硫氫基(sulfhydryl)會受到細胞內氧化壓力的影響,當硫氫基受到氧化壓力修飾 後,造成Nrf2與Keap1分開,進而促使Nrf2結合至ARE基因序列(Dinkova-Kostova et al., 2002)。因此,當細胞受到氧化壓力刺激後,調控活化ARE基因序列的過程中, Keap1及Nrf2兩者同時扮演重要角色。. * Nuclear factor-κB (NF-κB) 除了Nrf2和AP-1,NF-κB也被證實會向上調控HO-1基因表現。在刺激物如血 紅素(hemin)、鎘(cadmium)及LPS刺激下,會活化NF-κB而向上調控HO-1基因表現 (Liu et al., 2004;Chen et al., 2004;Wijayanti et al., 2004)。人類肝癌母細胞株(human hepatoblastoma-derived HepG2 cells)過度表現NF-κB,使得HO-1 mRNA表現也增加 (Lavrovsky et al., 2000)。. 22.
(30) 圖2-8. Nrf2/ARE的活化路徑 (Mann et al., 2007). 圖2-9. Nrf2的調控網路 (Itoh et al., 2004). 23.
(31) 三. 穿心蓮. 1. 簡介 穿心蓮,拉丁學名為Andrographis paniculata (Burm. f.) Nees,屬於唇形目 (Lamiales)、爵床科 (Acathaceae)、穿心蓮屬(Andrographis),為一年生草本植 物,別名一見喜、苦膽草、印度草、斬蛇劍、金香草、欖核蓮等,主要分布於 東南亞熱帶地區如印度、斯里蘭卡、巴基斯坦、爪哇、緬甸、泰國、越南、印 尼等地,非洲及南美洲也有其蹤跡,中國大陸與台灣各地也有零星栽培。植株 直立、多枝,花期 9 月~10 月,採收時視生長旺盛欲開花前,割取地上部分或 全株拔起,洗淨,曬乾,可內服(煎湯)及外用(煎汁塗抹或研末調敷)。穿心蓮 味苦性寒,具清熱解毒、涼血及消腫止痛之效,在傳統治療上常用於感冒發熱、 咽喉腫痛、口舌生瘡、上呼吸道感染、氣管炎及毒蛇咬傷等。穿心蓮的葉子及 根部在傳統藥理上已被使用數百年,在亞洲和歐洲民間更廣泛用來作為疾病治 療或預防保健 (表 2-6) ( Jarukamjorn and Nemoto, 2008)。穿心蓮的葉子和莖部 可萃取出具有活性的植物化學成份(phytochemicals),研究指出穿心蓮的活性成 分中含有豐富的diterpenoids and 2′-oxygenated flavonoids,包括andrographolide, neoandrographolide, 14-deoxy-11, 12-didehydroandrographolide, 14-deoxyandrographolide, isoandrographolide, 及 14-deoxyandrographolide, homoandrographolide, andrographan, andrographosterin, and stigmasterol等 (Pholphana et al., 2004)。在這些diterpenoids中,穿心蓮內酯(andrographolide) 是最重要的活性成分,其結構式為 (3-[2-{decahydro-6-hydroxy-5-(hydroxymethyl)-5,8α-dimethyl-2-methylene-1-nap thalenyl}ethylidene]dihydro-4-hydroxy-2(3H)-furanone);分子式C20H30O5;分子 量為 350.5,它存在植株的各部分,在全株乾燥的穿心蓮中,其莖內穿心蓮內 酯約含 0.8~1.2%,而葉子內的含量最多(>2%)。根據研究發現,穿心蓮具有廣. 24.
(32) 泛的生理功效,包括抗發炎 (Chiou et al., 2000;Sheeja et al., 2006)、抗腹瀉 (antidiarrhoeal) (Gupta et al., 1990)、抗病毒(Wiart et al., 2005)、保護肝臟(Trivedi and Rawal, 2001)、抗癌(Kumar et al., 2004;Li et al., 2007;Zhou et al., 2006)等。. 表 2-6. 穿心蓮的傳統醫療作用 (Jarukamjorn and Nemoto, 2008). 圖 2-10. 穿心蓮及穿心蓮內酯結構 (Jarukamjorn and Nemoto, 2008;Singha et al., 2007). 25.
(33) 2. 生理功效 2-1 抗發炎作用 許多文獻指出穿心蓮及穿心蓮內酯皆具有抗發炎的功效,過去研究發現穿心 蓮萃取物可用來預防或治療上呼吸道感染(Poolsup et al., 2004),Shen 等人(2002) 提出,穿心蓮內酯的抗發炎作用機制可能是藉由預防 ROS 的產生,以 N-formyl-methionyl-leucyl-phenylalanine (fMLP)誘導人類嗜中性白血球為模式,給 予穿心蓮內酯 (0.1~10 μM),可以透過調控 protein kinase C (PKC) 路徑減少 ROS 的產生,進而降低 Mac-1 表現而抑制白血球的 adhesion 和 transmigration。也有研 究指出,穿心蓮內酯可以直接影響 NF-κB 和 DNA 的結合而減少促發炎激素的表 現,例如 cyclooxygenase 2 (Cox-2) 和 nitric-oxide synthase (NOS) (Hidalgo et al., 2005;Iruretagoyena et al., 2005)。穿心蓮內酯在小鼠巨噬細胞株 RAW 264.7 中, 會透過抑制 inducible NO synthase (iNOS)表現來減少 LPS 所誘發的 NO,另外, 研究也發現以 LPS 刺激小鼠腹腔內巨噬細胞,穿心蓮內酯會透過抑制 ERK1/2 的 活化而減少 tumor necrosis factor-α (TNF-α) 及 interleukin-12 (IL-12)產生(Qin et al., 2006)。 此外,Bao 等人(2009)證實,在 BALB/c 小鼠以 ovalbumin (OVA)誘發 Th2 細胞過敏導致氣喘的模式中,給與穿心蓮內酯可以抑制 nuclear factor-κB (NF-κB)的活化,而達到抗發炎的作用。另外,Chao 等人(2009)在最近的研究也 發現,以 LPS 刺激 Raw264.7 巨噬細胞株的模式中,穿心蓮的乙酸乙酯萃出物可 以透過抑制 NF-κB 的活化而減少 TNF-α、IL-6、MIP-2、NO 的產生;穿心蓮的乙 酸乙酯萃出物也會抑制實驗小鼠的初代巨噬細胞產生 TNF-α、IL-12p40、MIP-2 及 NO 進而抑制發炎作用。. 2-2 抗癌作用 在現代醫學中,大多數的抗癌藥物透過直接抑制癌細胞增生、或是誘導癌細. 26.
(34) 胞凋亡壞死、以及造成細胞週期停滯來達到抗癌的作用,穿心蓮被證實也可透過 類似的機制,直接或間接調控癌細胞,而達到抗癌的功效(Vojdani and Erde, 2006)。 Kumar 等人(2004)研究發現,穿心蓮的甲醇萃取物可以顯著抑制人類結腸癌細胞 (HT-29 cell)的增生,在人類急性骨髓白血病癌細胞(human acute myeloid leukemic HL-60 cells)中,穿心蓮乙醇萃出物及穿心蓮內酯會誘導細胞週期停止,並且透過 調控促細胞凋亡(pro-apoptotic)分子的表現來影響癌細胞進行細胞凋亡反應 (Cheung et al., 2005)。另外,穿心蓮也可以抑制肺癌細胞,研究發現穿心蓮內酯 會藉由抑制 PI3K/Akt/AP-1 訊息傳遞路徑來降低 matrix metalloproteinase-7 (MMP-7) 表現,進而減少非小細胞肺癌細胞(A549)的侵襲性(Lee et al., 2010)。血 管內皮生長因子(vascular endothelial growth factor, VEGF) 是血管內皮細胞生長和 存活重要的因素之一,它會調控血管內皮細胞增生、血管新生和血管通透性, VEGF 會被腫瘤細胞大量利用以利其存活,有研究證實穿心蓮內酯可以抑制 VEGF 和 nitric oxide (NO)產生,並提高細胞中抗血管新生因子表現,如 IL-2 和組 織金屬蛋白酵素抑制劑(tissue inhibitor of metalloproteinase ,TIMP-1) (Sheeja et al., 2007)。這些研究皆顯示穿心蓮內酯具有抑制腫瘤生長的潛力。. 2-3 肝臟保護作用 過去文獻指出穿心蓮具有保護肝臟的功效,研究顯示在小鼠肝臟中,穿心蓮 可以預防 hexachlorocyclohexane (BHC)刺激下所造成的γ-glutamyl transpeptidase (γ-轉胺基胜肽酶)、glutathione-S-transferase (GST)及脂質過氧化的增加,顯示穿 心蓮具有潛在的抗氧化和保肝作用(Trivedi and Rawal, 2001)。在動物實驗中給予 穿心蓮內酯,對半乳醣胺(galactosamine)誘發的肝損傷具有保護的功能(Handa and Sharma, 1990)。另外,小鼠在以四氯化碳(carbon tetrachloride)誘發肝損傷模式下, 穿心蓮內酯會藉由降低malondialdehyde (MDA)、glutamate pyruvate transaminase (GPT)及alkaline phosphatase (ALP) 並維持血漿中高濃度的glutathione (GSH)而達 到保護肝臟的功效(Kapil et al., 1993)。 27.
(35) 3. 使用風險 穿心蓮在傳統醫學上常用於治療感冒,因此評估穿心蓮萃取物是否具有毒性 值得注意。急性口服毒性數據測試可用來評估人類在使用藥物上的風險。 Chandrasekaran 等人(2009)以 8-12 週的大鼠來做急性口服毒性試驗,研究發現每 公斤體重的大鼠給予穿心蓮萃出物 5000 mg,研究結束大鼠仍存活,並在 14 天的 實驗期間,並沒有顯示出任何的不良症狀,也沒有任何不正常的體重增加情形。 但是,有文獻指出,每天給予雄性小白鼠乾燥的穿心蓮葉磨成的粉末 20 mg,持 續 60 天會停止精子的產生(Akbarsha et al., 1990)。廣州中醫藥大學徐等人(2005), 以小鼠來做急性毒性試驗,每公斤體重管灌穿心蓮軟膠囊內容物 15 ml(每粒膠囊 含有 2.1 g 的穿心蓮生藥),一天兩次,一日總給藥量為 110.25 g/kg,並連續觀察 14 天,結果發現給藥後的小鼠無躁動不安、呼吸困難、突眼等急性毒性表現,給 藥後連續觀察 14 天並無小鼠死亡,且藥後兩周內小鼠的正常生長週期並未顯著 受影響,此急性毒性試驗發現,小鼠一日最大耐受量為 110.25 g/kg,相當於成人 臨床推薦用量的 525 倍,顯示穿心蓮在臨床上的應用是安全的。此外,在中草藥 被廣泛應用的現代社會,中草藥與藥物間的交互作用也是一個嚴重的問題,以動 物模式利用 HPLC/UV 來分析穿心蓮萃出物的成分,實驗發現預先給予穿心蓮內 酯可以增加血液內茶鹼(theophylline)的清除,若長期使用穿心蓮則會提高茶鹼在 血液內的濃度,研究中提出穿心蓮萃出物和其中的草藥成分會和 CYP1A2 產生交 互作用,因此在使用上應多加留意(Chien et al., 2010)。. 28.
(36) 研究目的. 動脈粥狀硬化是一種慢性發炎疾病,會造成血管內壁脂肪堆積及纖維斑塊形 成,是導致心血管疾病的主要原因之一。許多文獻指出穿心蓮及穿心蓮內酯皆具 有抗發炎的藥理作用,而穿心蓮內酯更是穿心蓮中的重要活性成分,因此本實驗 將利用內皮細胞株 EA. hy926 以腫瘤壞死因子 TNF-α 誘發其發炎反應,之後探討 穿心蓮內酯對於內皮細胞發炎反應的影響及其作用機制。. 29.
(37) 文獻參考 徐志勇.(2005). 穿心蓮軟膠囊與穿心蓮片的藥理作用及急性毒性實驗研究. 廣州中 醫藥大學. Arjan van der Flier, Arnoud Sonnenberg.(2001). Function and interactions of integrins. Cell Tissue Res 305, 285–298. Akihiro Yachie, Yo Niida, Taizo Wada, Noboru Igarashi, Hisashi Kaneda, Tomoko Toma, Kazuhide Ohta, Yoshihito Kasahara and Shoichi Koizumi. (1999). Oxidative stress causes enhanced endothelial cell injury in human heme oxygenase-1 deficiency. European Journal of Clinical Investigation. 103, 129-135. Akbarsha, M.A., Manivannan, B., Hamid, K.S., Vijayan, B., (1990). Antifertility effect of Andrographis paniculata (Nees) in male albino rat. Indian Journal of Experimental Biology. 28, 421–426. By Sophie Brouard, Leo E. Otterbein, Josef Anrather, Edda Tobiasch, Fritz H. Bach, Augustine M.K. Choi, and Miguel P. Soares. (2000). Carbon monoxide generated by heme oxygenase 1 suppresses endothelial cell apoptosis. The Journal of Experimental Medicine. 192,1015–1026 Baranano, D. E., Rao, M., Ferris, C. D. and Snyder, S. H. (2002). Biliverdin reductase: a major physiologic cytoprotectant. Proceedings of the National Academy of Sciences of the United States of America 99, 16093-16098. Briscoe DM, Cotran RS, Pober JS. (1992) Effects of tumor necrosis factor, lipopolysaccharide, and IL-4 on the expression of vascular cell adhesion molecule 1 in vivo: correlation with CD3+ T-cell infiltration. Journal of Immunology. 149, 2954–2960. Claudio fiocchi (1998). Inflammatory Bowel Disease: Etiology and Pathogenesis. Gastroenterology. 115, 182–205. Chiou,W. F., Chen, C. F. and Lin, J. J. (2000). Mechanisms of suppression of inducible nitric oxide synthase (iNOS) expression in RAW 264.7 cells by andrographolide. British Journal of Pharmacology. 129, 1553–1560.. 30.
(38) Chao-Feng Chiena, Yu-Tse Wua, Wen-Chuan Leea, Lie-Chwen Linb, Tung-Hu Tsai. (2010). Herb–drug interaction of Andrographis paniculata extract and andrographolide on the pharmacokinetics of theophylline in rats. Chemico-Biological Interactions. 184, 458–465. C.V. Chandrasekaran , P. Thiyagarajan, K. Sundarajan, Krishna S. Goudar, M. Deepak, B. Murali, J. Joshua Allan, Amit Agarwal.(2009). Evaluation of the genotoxic potential and acute oral toxicity of standardized extract of Andrographis paniculata (KalmColdTM). Food and Chemical Toxicology. 47, 1892–1902.. D. Zapolska-Downar, A. Zapolski-Downar , M. Markiewski , A. Ciechanowicz , M. Kaczmarczyk , M. Naruszewicz. (2001). Selective inhibition by probucol of vascular cell adhesion molecule-1 (VCAM-1) expression in human vascular endothelial cells. Atherosclerosis. 155,123–130. Eriksson EE, Xie X, Werr J, Thoren P, Lindbom L.(2001) Importance of primary capture and L-selectin-dependent secondary capture in leukocyte accumulation in inflammation and atherosclerosis in vivo. The Journal of Experimental Medicine. 194, 205–218. Fondevila, C., Shen, X. D., Tsuchiyashi, S., Yamashita, K.,Csizmadia, E.,Lassman, C., Busuttil, R. W., Kupiec-Weglinski, J. W. and Bach, F. H. (2004). Biliverdin therapy protects rat livers from ischemia and reperfusion injury. Hepatology. 40, 1333-1341. Grosser, N., Oberle, S., Berndt, G., Erdmann, K., Hemmerle, A. and Schroder, H.(2004). Antioxidant action of L-alanine: heme oxygenase-1 and ferritin as possible mediators. Biochem. Biophys. Res. Commun. 314, 351-355. Gonzales, S., Polizio, A. H., Erario, M. A. and Tomaro, M. L.(2005). Glutamine is highly effective in preventing in vivo cobalt-induced oxidative stress in rat liver. World J. Gastroenterol. 11, 3533-3538. Galkina E, Kadl A, Sanders J, Varughese D, Sarembock IJ, Ley K.(2006).Lymphocyte recruitment into the aortic wall before and during development of atherosclerosis is partially L-selectin dependent. The Journal of Experimental Medicine. 203,1273–1282.. 31.
(39) Gunter voraberger, renate schafer, and christian stratowa.(1991). Cloning of the human gene for intercellular adhesion molecule and analysis of its 5”regulatory region Induction by Cytokines and Phorbol Ester. The Journal of Immunology. 147, 2777-2786. Gahmberg CG, Tolvanen M, Kotovuori P. (1997). Leukocyte adhesion structure and function of human leukocyte beta2-integrins and their cellular ligands. European Journal of Biochemistry/FEBS. 245, 215-232. Guan JL, Hynes RO.( 1990). Lymphoid cells recognize an alternatively spliced segment of fibronectin via the integrin receptor alpha 4 beta 1. Cell. 60, 53– 61. Giovanni E. Mann, Jörg Niehueser-Saran, Alan Watson, Ling Gao, Tetsuro Ishii, Patricia de Winter, Richard C. M. Siow.(2007). Nrf2/ARE regulated antioxidant gene expression in endothelial and smooth muscle cells in oxidative stress: implications for atherosclerosis and preeclampsia. Acta Physiologica Sinica. 59, 117-127. Gupta, S., Choudhry, M. A., Yadava, J. N. S., Srivastava, V. and Tandon, J. S. (1990). Antidiarrhoeal activity of diterpenes of Andrographis paniculata (Kal-Megh) against Escherichia coli enterotoxin in in vivo models. International Journal of Crude Drug Research. 28, 273–283.. Huang HC, Nguyen T, Pickett CB. (2002). Phosphorylation of Nrf2 at Ser-40 by protein kinase C regulates antioxidant response element -mediated transcription. The Journal of Biological Chemistry.; 277: 42769-42774. Hidalgo MA, Romero A, Figueroa J, Cortes P, Concha II, Hancke JL, Burgos RA. (2005). Andrographolide interferes with binding of nuclear factor-kB to DNA in HL-60-derived neutrophilic cells. British Journal of Pharmacology. 144, 680–686. Hon-Yeung Cheung, Sau-Ha Cheung, Jieliang Li, Chui-Shan Cheung, Wai-Ping Lai, Wang-Fun Fong, Fo-Man Leung.(2005). Andrographolide isolated from Andrographis paniculata induces cell cycle arrest and mitochondrial-mediated apoptosis in human leukemic HL-60 cells. Planta medica. 71, 1106-1111. Handa, S. and Sharma, A. (1990). Hepatoprotective activity of andrographolide against galactosamine & paracetamol intoxication in rats. The Indian Journal of Medical Research. 92, 284–292. 32.
(40) Immenschuh, S., Schroder, H.(2006). Heme oxygenase-1 and cardiovascular disease. Histology and histopathology. Cellular and Molecular Biology. 21, 679–685. Ishikawa, K., Takeuchi, N., Takahashi, S., Matera, K. M., Sato, M., Shibahara, S., Rousseau, D. L., Ikeda-Saito, M. and Yoshida, T. (1995). Heme oxygenase-2. Properties of the heme complex of the purified tryptic fragment of recombinant human heme oxygenase-2. The Journal of Biological Chemistry. 270, 6345-6350. Ishii T, Itoh K, Takahashi S, Sato H, Yanagawa T, Katoh Y, Bannai S, Yamamoto M. (2000). Transcription factor Nrf2 coordinately regulates a group of oxidative stress-inducible genes in macrophages. The Journal of Biological Chemistry. 275, 16023-16029. Iruretagoyena MI, Tobar JA, Gonzalez PA, Sepulveda SE, Figueroa CA, Burgos RA. (2005). Andrographolide interferes with T Cell activation and reduces experimental autoimmune encephalomyelitis in the mouse. The Journal of Pharmacology and Experimental Therapeutics. 312, 366–372. Johnson-Tidey RR, McGregor JL, Taylor PR, Poston RN. (1994). Increase in the adhesion molecule P-selectin in endothelium overlying atherosclerotic plaques. Coexpression with intercellular adhesion molecule-1. The American Journal of Pathology. 144, 952–961. James W. Kaspar, Suryakant K. Niture, Anil K. Jaiswal. (2009). Nrf2:INrf2 (Keap1) signaling in oxidative stress. Free Radical Biology & Medicine. 47, 1304–1309. Kanokwan Jarukamjorn, and Nobuo Nemoto. (2008). Pharmacological aspects of Andrographis paniculata on health and its major diterpenoid constituent Andrographolide. Journal of Health Science. 54, 370–381. Kumar RA, Sridevi K, Kumar NV, Nanduri S, Rajagopal S. (2004). Anticancer and immunostimulatory compounds from Andrographis paniculata. Journal of Ethnopharmacology. 92, 291-295. Kapil, A., Koul, I. B., Banerjee, S. K. and Gupta, B. D. (1993). Antihepatotoxic effects of major diterpenoid constituents of Andrographis paniculata. Biochemical Pharmacology. 46, 182–185.. 33.
(41) Karin, M. (1996). The regulation of AP-1 activity by mitogen-activated protein kinases. Philosophical transactions of the Royal Society of London. Series B, Biological Sciences. 351, 127–134. Koichiro Sato, Jozsef Balla, Leo Otterbein, R. Neal Smith, Sophie Brouard, Yuan Lin, Eva Csizmadia, Jean Sevigny, Simon C. Robson, Gregory Vercellotti, Augustine M. Choi, Fritz H. Bach and Miguel P. Soares. (2001). Carbonmonoxide generated by heme oxygenase-1 suppresses the rejection of mouse-to-rat cardiac transplants. American Journal of Immunology. 166, 4185–4194. Kelly J. Peyton, Sylvia V. Reyna, Gary B. Chapman, Diana Ensenat, Xiao-ming Liu, Hong Wang, Andrew I. Schafer, and William Durante.(2002). Heme oxygenase-1-derived carbon monoxide is an autocrine inhibitor of vascular smooth muscle cell growth. Blood . 99, 4443–4448. L.A. Stanciu, R. Djukanovic (1998). The role of ICAM-1 on T-cells in the pathogenesis of asthma. The European respiratory journal : official journal of the European Society for Clinical Respiratory Physiology. 11, 949–957. Leo E. Otterbein, Miguel P. Soares, Kenichiro Yamashita and Fritz H. Bach. (2003). Heme oxygenase-1: unleashing the protective properties of heme. Trends in Immunology. 24, 449-455. Lin-Hua QIN, Ling KONG, Guo-Jun SHI, Zheng-Tao WANG, and Bao-Xue G. (2006). Andrographolide Inhibits the Production of TNF-α and Interleukin-12 in Lipopolysaccharide-Stimulated Macrophages: Role of Mitogen-Activated Protein Kinases. Biological & Pharmaceutical Bulletin. 29, 220-224. Llesuy, S. F and Tomaro, M. L. (1994). Heme oxygenase and oxidative stress. Evidence of involvement of bilirubin as physiological protector against oxidative damage. Biochimica et Biophysica acta. 1223, 9-14. Li, J., Cheung, H. Y., Zhang, Z., Chan, G. K. L. and Fong,W. F.(2007). Andrographolide induces cell cycle arrest at G2/M phase and cell death in HepG2 cells via alteration of reactive oxygen species. European Journal of Pharmacology. 568, 31–44. Look, D. C., Pelletier, M. R., Tidwell, R. M., Roswit, W. T., Holtzman, M. J. (1995). Stat1 depends on transcriptional synergy with Sp1. The Journal of Biological Chemistry. 270, 30264–30267. 34.
(42) Leo E. Otterbein, Brian S. Zuckerbraun, Manabu Haga, Fang Liu, Ruiping Song, Anny Usheva ,Christina Stachulak, Natalya Bodyak, R. Neal Smith, Eva Csizmadia, Shivraj Tyagi, Yorihiro Akamatsu, Richard J. Flavell, Timothy R. Billiar, Edith Tzeng, Fritz H. Bach, Augustine M.K. Choi & Miguel P. Soares .(2003). Carbon monoxide suppresses arteriosclerotic lesions associated with chronic graft rejection and with balloon injury. Journal of Natural Medicines. 9, 183–190. Maines, M. D. and Gibbs, P. E. (2005). 30 some years of heme oxygenase: from a “molecular wrecking ball” to a “mesmerizing” trigger of cellular events. Biochemical and Biophysical Research Communications. 338, 568-577. Maines, M. D. (1988). Heme oxygenase: function, multiplicity, regulatory mechanisms, and clinical applications. The Fede ration of American Societies for Experimental Biology. 2, 2557-2568. McCoubrey, W. K. Jr., Huang, T. J. and Maines, M. D. (1997). Isolation and characterization of a cDNA from the rat brain that encodes hemoprotein heme oxygenase-3. European Journal of Biochemistry/FEBS. 247, 725-732. Motterlini, R., Foresti, R., Bassi, R., Calabrese, V., Clark, J. E. and Green, C. J. (2000). Endothelial heme oxygenase-1 induction by hypoxia. Modulation by inducible nitric-oxide synthase and S-nitrosothiols. The Journal of biological chemistry. 275, 13613-13620. Moore, B. A., Overhaus, M., Whitcomb, J., Ifedigbo, E., Choi, A. M., Otterbein, L. E. and Bauer, A. J. (2005). Brief inhalation of low-dose carbon monoxide protects rodents and swine from postoperative ileus. Critical Care Medicine. 33, 1317-1326. Malaguarnera, L., Madeddu, R., Palio, E., Arena, N. and Malaguarnera, M. (2005). Heme oxygenase-1 levels and oxidative stress-related parameters in non-alcoholic fatty liver disease patients . Journal of Hepatology. 42, 585-591. Menno P.J. de Winther, Edwin Kanters, Georg Kraal, Marten H. Hofker. (2005). Nuclear Factor B Signaling in Atherogenesis. Arteriosclerosis, Thrombosis, and Vascular biology. 25, 904-914. Mercurio, F., Zhu, H., Murray, B. W., Shevchenko, A., Bennett, B. L., Li, J., Young, D. B., Barbosa, M., Mann, M., Manning, A., Rao, A. (1997). IKK-1 and IKK-2: cytokine-activated IkappaB kinases essential for NF-kappaB activation. Science. 278, 35.
(43) 860–866. Mahin D Maines.(2001). The heme oxygenase sytem: Update 2005. Antiox Redox Signal.7, 1761-1766. N. Poolsup PhD, C. Suthisisang PhD, S. Prathanturarug PhD, A. Asawamekin BPharm and U. Chanchareon BPharm.(2004). Andrographis paniculata in the symptomatic treatment of uncomplicated upper respiratory tract infection: systematic review of randomized controlled trials. Journal of Clinical Pharmacy and Therapeutics. 29, 37–45. Ossola, J. O. and Tomaro, M. L.(1998). Heme oxygenase induction by UVA radiation. A response to oxidative stress in rat liver. The International Journal of Biochemistry & Cell Biology. 30, 285-292. Pholphana, N., Rangkadilok, N., Thongnest, S., Ruchirawat, S., Ruchirawat, M. and Satayavivad. (2004). Determination and variation of three active diterpenoids in Andrographis paniculata (Burm.f.) Nees. Phytochemical Analysis. 15, 365–371. Pannen, B. H., Kohler, N., Hole, B., Bauer, M., Clemens, M. G. and Geiger, K. K. (1998). Protective role of endogenous carbon monoxide in hepatic microcirculatory dysfunction after hemorrhagic shock in rats. European Journal of Clinical Investigation. 102, 1220-1228. P J Lee, J Alam, G W Wiegand, and A M Choi. (1996). Overexpression of heme oxygenase-1 in human pulmonary epithelial cells results in cell growth arrest and increased resistance to hyperoxia. Proceedings of the National Academy of Sciences of the United States of America. 93, 10393–10398. Ryter, S.W., Alam, J., Choi, A.M.K. (2006). Heme oxygenase-1/carbon monoxide: from basic science to therapeutic applications. Physiological Reviews. 86, 583–650. Ramos CL, Huo Y, Jung U, Ghosh S, Manka DR, Sarembock IJ, Ley K. (1999). Direct demonstration of P-selectin- and VCAM-1-dependent mononuclear cell rolling in early atherosclerotic lesions of apolipoprotein E-deficient mice. Circulation Research. 84, 1237–1244. Ross R (1999). Atherosclerosis- an inflammatory disease. The New England journal of medicine. 340(2):115-26. Sarady, J. K., Zuckerbraun, B. S., Bilban, M., Wagner, O., Usheva, A., Liu, F., Ifedigbo, E., Zamora, R., Choi, A. M. and Otterbein, L.E. (2004). Carbon monoxide protection 36.
(44) against endotoxic shock involves reciprocal effects on iNOS in the lung and liver. The Federation of American Societies for Experimental Biology. 18, 854-856. Sawle, P., Foresti, R., Mann, B. E., Johnson, T. R., Green, C. J. and Motterlini, R. (2005). Carbon monoxide-releasing molecules (CO-RMs) attenuate the inflammatory response elicited by lipopolysaccharide in RAW264.7 murine macrophages. British Journal of Pharmacology. 145, 800-810. Sheeja, K., Shihab, P. K. and Kuttan, G.(2006). Antioxidant and anti-inflammatory activities of the plant Andrographis Paniculata Nees. Immunopharmacology and Immunotoxicology. 28, 129–140. Singha, P. K.; Roy, S. Dey, S. (2007). Protective activity of andrographolide and arabinogalactan proteins from Andrographis paniculata Nees. against ethanol-induced toxicity in mice. J. Ethnopharmacol. 111, 13–21. Sheeja K, Guruvayoorappan C, Kuttan G. (2007). Antiangiogenic activity of Andrographis paniculata extract and andrographolide. International Immunopharmacology. 7, 211–221. Stefan Blankenberg , Sandrine Barbaux , Laurence Tiret. (2003). Adhesion molecules and atherosclerosis. Atherosclerosis. 170, 191-203. Song, S., Ling-Hu, H., Roebuck, K. A., Rabbi, M. F., Donnelly, R. P., Finnegan, A.(1997). Interleukin-10 inhibits interferon-gamma-induced intercellular adhesion molecule-1 gene transcription in human monocytes. Blood. 89, 4461–4469. Tobiasch, Edda; Günther, Lukas; Bach, Fritz H. (2001). Heme oxygenase-1 protects pancreatic beta cells from apoptosis caused by various stimuli. Investigación Médica Internacional. 49, 566–571. Trivedi, N. and Rawal, U. M. (2001). Hepatoprotective and antioxidant property of Andrographis paniculata (Nees) in BHC induced liver damage in mice. Indian Journal of Experimental Biology. 39, 41–46. Vojdani A, Erde J. (2006) Regulatory T cells, a potent immunoregulatory target for CAM researchers: modulating tumor immunity, autoimmunity and alloreactive immunity (III). Evidence-based complementary and alternative medicine. 3, 309-316. 37.
(45) Victor J. Dzau, Ruediger C. Braun-Dullaeus & Daniel G. (2002). Vascular proliferation and atherosclerosis: New perspectives and therapeutic strategies. Nature Medicine. 8, 1249-1256. Wiart, C., Kumar, K.,Yusof, M. Y., Hamimah, H., Fauzi, Z. M. and Sulaiman, M. (2005). Antiviral properties of ent-labdene diterpenes of Andrographis paniculata Nees, inhibitors of herpes simplex virus type 1. Phytotherapy Research. 19, 1069-1070. Wen-Wan Chao, Yueh-Hsiung Kuo, Shie-Liang Hsieh and Bi-Fong Lin. (2009). Inhibitory effects of ethyl acetate extract of Andrographis paniculata on NF-κB trans-activation activity and LPS-induced acute inflammation in mice. Evidence-based Complementary and Alternative Medicine. 10, 1-9. Yuh-Chiang Shen, Chieh-Fu Chen & ,Wen-Fei Chiou. (2002). Andrographolide prevents oxygen radical production by human neutrophils: possible mechanism(s) involved in its anti-infammatory effect. British Journal of Pharmacology. 135, 399-406. Yi-Chieh Lee , Hui-Hsuan Lin , Chin-Hsun Hsu, Chau-Jong Wang , Tai-An Chiang , Jing-Hsien Chen.(2010). Inhibitory effects of andrographolide on migration and invasion in human non-small cell lung cancer A549 cells via down-regulation of PI3K/Akt signaling pathway. European Journal of Pharmacology. 632, 23-32. Yu G, Rux AH, Ma P, Bdeir K, Sachais BS. (2005). Endothelial expression of E-selectin is induced by the platelet-specific chemokine platelet factor 4 through LRP in an NF-kappaB-dependent manner. Blood. 105, 3545–3551. Zipper, L. M.; Mulcahy, R. T. (2002). The Keap1 BTB/POZ dimerization function is required to sequester Nrf2 in cytoplasm. The Journal of Biological Chemistry. 277, 36544-36552. Zhou, J., Zhang, S., Ong, C. and Shen, H. (2006). Critical role of pro-apoptotic Bcl-2 family members in andrographolide-induced apoptosis in human cancer cells. Biochemical Pharmacology. 72, 132-144. Zhang Bao1, Shouping Guan, Chang Cheng, Songlian Wu, Siew Heng Wong, D. Michael Kemeny, Bernard P. Leung, and W. S. Fred Wong. (2009). A Novel Antiinflammatory Role for Andrographolide in Asthma via Inhibition of the Nuclear Factor-kB Pathway. American Journal of Respiratory and Critical Care Medicine. 179, 657-665. 38.
(46) 第二部分. Induction of Heme Oxygenase 1 and Inhibition of Tumor Necrosis Factor α-Induced Intercellular Adhesion Molecule Expression by Andrographolide in EA.hy926 Cells. INTRODUCTION. Current epidemiologic predictions suggest that cardiovascular disease (CVD) is reaching pandemic proportions (1), and CVD is the leading cause of death worldwide, accounting for ~16.7 million deaths each year (2). This number is predicted to reach approximately 25 million by 2020, if current trends continue (3). The reasons for the increased global incidence of CVD include the aging of the world’s population and lifestyles in lower- and middle-income countries becoming more akin to those of wealthier nations (1). CVD is partially characterized by chronic inflammation and an increased level of expression of inflammatory biomarkers, such as intercellular adhesionmolecule (ICAM-1), vascular cell adhesion molecule (VCAM-1), and E-selectin, on the surface of the activated endothelial cells (EC) and in the blood circulation (4). Increased circulating levels of these adhesion molecules are considered to be predictive of CVD risk because they indicate a proinflammatory state in the vasculature (4). In vivo inflammatory processes are mediated by the involvement of proinflammatory mediators, including tumor necrosis factor α (TNF-α), interleukin 6 (IL-6), IL-12, and interferon γ (INF-γ). EC play a key role in coordinating leukocyte trafficking to specific tissues that 39.
(47) regulate their activation (5). When confronted by an inflammatory stimulus, e.g., bacterial lipopolysaccharide (LPS) (6), TNF-α(7), or IL-1β (8), EC become activated as a result of modification of their phenotype. This is accompanied primarily by the upregulation of a series of proinflammatory genes, e.g., E-selectin, P-selectin, VCAM-1, and ICAM-1 (6-8). The expression of these genes is regulated primarily by the activation of transcription factor NF-κB (9). The proper production of proinflammatory cytokines aids in innate immune responses; however, overproduction causes undesirable side effects such as tissue injury, septic shock, and even death (10). To avoid the consequences of uncontrolled production, the expression of proinflammatory genes must be strictly regulated (11). One of the mechanisms by which inflammation is counteracted is the expression of the anti-inflammatory genes of activated EC. Because of their dual ability to inhibit the expression of proinflammatory genes associated with EC activation and to protect EC from undergoing apoptosis, Bach and colleagues (11) referred to these genes as protective genes. Soares et al. (12) have hypothesized that the stress-responsive gene encoding heme oxygenase 1 (HO-1) acts in such a manner. HO-1 is an inducible enzyme that catalyzes the rate-limiting step in the oxidative degradation of free heme into Fe2+ , carbon monoxide, and biliverdin, which is subsequently catabolized into bilirubin by biliverdin reductase (13). Induction of the gene encoding HO-1 is primarily regulated at the transcriptional level, and transcription factor Nrf2 plays a critical role in the inducibility of the gene (14). Under unstimulated conditions, Nrf2 is sequestered in the cytoplasm by binding to Klech-like ECHassociated protein 1 (Keap1) (15). This complex is disrupted by several electrophilic antioxidants, and Nrf2 is freed and translocated to the nucleus where it binds to antioxidant response element (ARE) sequences in the HO-1 gene promoter (16, 17). HO-1 can also be induced by its substrate, free heme, and by a diversity of proinflammatory stimuli, which suggests that HO-1 is involved in heme degradation as 40.
(48) well as in resolution of inflammation (18). Andrographis paniculata (Burm. f) Nees, a Chinese herb, is a member of the Acanthaceae family. It is widely cultivated in Southeast Asia and is widely used as a traditional medicine in India, China, Thailand, and Scandinavia (19). Andrographolide (Figure 1) is the most abundant diterpene lactone in the leaves and stems of A. paniculata and has high biological activity and therapeutic potential (20). Andrographolide has been studied for its anti-inflammatory (21-23), chemopreventive (24), antiangiogenic (25), antiproliferative (26), and antiapoptotic (27) activities. Because of the significant role of inflammation in CVD development, the cardiovascular benefits of andrographolide as the result of its anti-inflammatory activity cannot be ignored. Therefore, on the basis of the well-known anti-inflammatory activity of andrographolide, this study was designed to explore the effect of andrographolide on the TNF-α-induced expression of ICAM-1 in EA hy.926 cells and the mechanism by which andrographolide acts to influence ICAM-1 expression.. MATERIALS AND METHODS. Materials.. Monocytic HL-60 cells were obtained from Bioresources Collection and. Research Center (BCRC, Taiwan). Dulbecco’s modified Eagle’s medium (DMEM), RPMI 1640, 25% trypsin-EDTA, penicillinstreptomycin, RPMI-1640 without phenol red, and OPTI-MEM were from GIBCO/BRL (Grand Island, NY). Fetal bovine serum (FBS) was from HyClone (Logan, UT). 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), sodium 41.
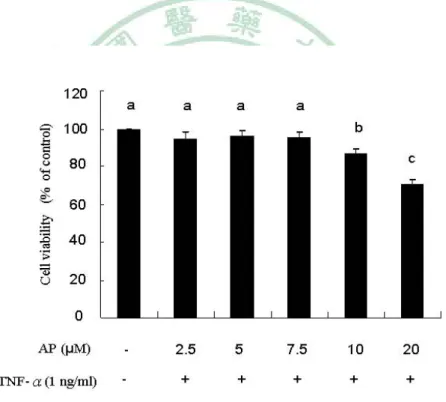
(49) bicarbonate, sodium phosphate dibasic heptahydrate, TNF-R, and all other chemicals were from Sigma (St. Louis, MO). TRIzol reagent was from Invitrogen (Carlsbad, CA). Andrographolide and antibody against HO-1 were from Calbiochem (Darmstadt, Germany). The antibody raised against ICAM-1 was from Cell Signaling Technology (Boston, MA). The antibody raised against Nrf2 was from Santa Cruz Biotechnology (Santa Cruz, CA). The luciferase assay kit was from Promega (Madison, WI). Bis(carboxyethyl) carboxyfluorescein acetoxymethyl ester (BCECF-AM) was from Molecular Probes (Eugene, OR). Cell Cultures. The human endothelial cell line EA.hy926 was cultured in DMEM supplemented with 3.7 g/L NaHCO3, 10% FBS, 100 units/mL penicillin, and 100 μg/mL streptomycin at 37℃ in a 5% CO2 humidified incubator. The HL-60 cells were cultured in T-75 tissue culture flasks in RPMI 1640 medium supplemented with 10% fetal bovine serum, 100000 units/L penicillin, and 100 mg/L streptomycin. Cell Viability Assay. Cell viability was assessed by the MTT assay. The MTT assay measures the ability of viable cells to reduce a yellow 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide to a purple formazan by mitochondrial succinate dehydrogenase.EA.hy926 cells were grown to 70-80% confluence and were then treated with different concentrations of andrographolide (0-20 μM) for 16 h followed by incubation with TNF-α (1 ng/mL) for an additional 6 h. Finally, the DMEM medium was removed, and the cells were washed with PBS. The cells were then incubated with MTT (0.5 mg/mL) in DMEM medium at 37 ℃for an additional 3 h.The mediumwas removed, and 2-propanolwas added to dissolve the formazan. After centrifugation at 20000g for 5 min, the supernatant of each sample was transferred to 96-well plates, and the absorbance was read at 570 nm in an ELISA reader. The absorbance in cultures treated with 0.1% DMSO was regarded as 100% cell viability. 42.
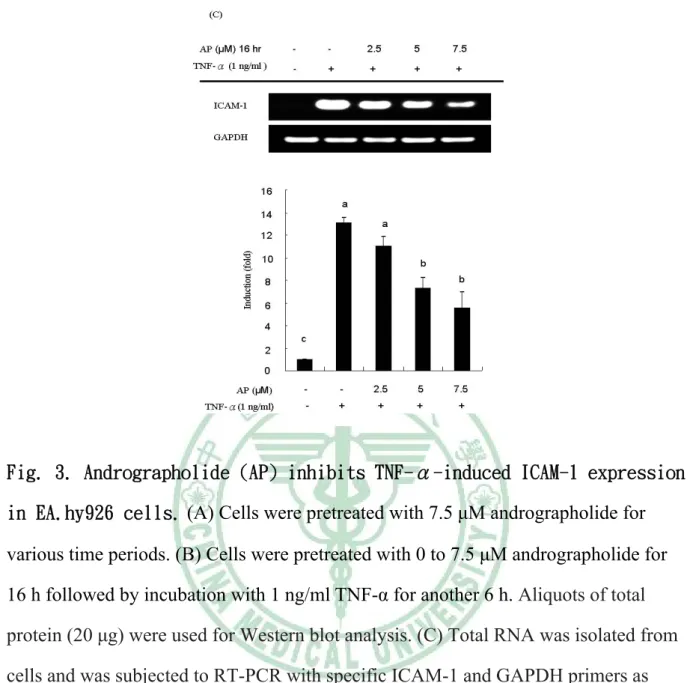
(50) Monocyte Adhesion Assay. EA.hy926 cells in 12-well plates were allowed to grow to 80% confluence and were then pretreated with andrographolide for 16 h followed by incubation with 1 ng/mL TNF-R for an additional 6 h. The humanmonocytic HL-60 cells cultured in RPMI 1640 medium with 10% FBS were labeled with 1 μM 2,7-bis(2-carboxyethyl)- 5(6)-carboxyfluorescein acetoxymethyl ester (BCECF-AM). At the end of the andrographolide and TNF-α treatment, a total of 1×106 BCECF-AM-labeled HL-60 cells were added to each well, and the cells were co-incubated with EA.hy926 cells at 37 ℃ for 30 min. The wells were washed and filled with cell culture medium, and the plates were sealed, inverted, and centrifuged at 100g for 5 min to remove nonadherent HL-60 cells. Bound HL-60 cells were lysed in a 1% SDS solution, and the fluorescence intensity was determined in a fluoroscan ELISA plate reader (FLX800, Bio-Tek, Winooski, VT) with an excitation wavelength of 480 nm and an emission wavelength of 520 nm. A control study showed that fluorescence is a linear function of HL-60 cell density in the range of 3000-80000 cells/well. The results are reported on the basis of the standard curve obtained. Western Blotting Analysis. After each experiment, cells were washed twice with cold PBS and were harvested in 150 μL of lysis buffer [10 mM Tris-HCl (pH 8), 0.1% Triton X-100, 320 mM sucrose, 5 mM EDTA, 1 mM PMSF, 1 mg/L leupeptin, 1 mg/L aprotinin, and 2 mM dithiothreitol]. Cell homogenates were centrifuged at 14000g for 20min at 4 ℃. The resulting supernatant was used as a cellular protein for Western blot analysis. The total protein was analyzed by use of the Coomassie Plus protein assay reagent kit (Pierce Biotechnology Inc.,Rockford, IL). Equal amounts of cellular proteins were electrophoresed in an sodium dodecyl sulfate-polyacrylamide gel, and proteins were then transferred to polyvinylidene fluoride membranes (Millipore Corp., Bedford, MA). Nonspecific binding sites on themembranes were blocked with 43.
數據




+7


相關文件
Wang, Solving pseudomonotone variational inequalities and pseudocon- vex optimization problems using the projection neural network, IEEE Transactions on Neural Networks 17
volume suppressed mass: (TeV) 2 /M P ∼ 10 −4 eV → mm range can be experimentally tested for any number of extra dimensions - Light U(1) gauge bosons: no derivative couplings. =>
For pedagogical purposes, let us start consideration from a simple one-dimensional (1D) system, where electrons are confined to a chain parallel to the x axis. As it is well known
The observed small neutrino masses strongly suggest the presence of super heavy Majorana neutrinos N. Out-of-thermal equilibrium processes may be easily realized around the
Define instead the imaginary.. potential, magnetic field, lattice…) Dirac-BdG Hamiltonian:. with small, and matrix
incapable to extract any quantities from QCD, nor to tackle the most interesting physics, namely, the spontaneously chiral symmetry breaking and the color confinement..
(1) Determine a hypersurface on which matching condition is given.. (2) Determine a
• Formation of massive primordial stars as origin of objects in the early universe. • Supernova explosions might be visible to the most
