國立臺灣大學生命科學院植物科學研究所 博士論文
Institute of Plant Biology College of Life Science
National Taiwan University Doctoral Dissertation
不同阿拉伯芥銅鋅超氧歧化酶對其活化機制偏好之研究 Copper Chaperone-Dependent and -Independent Activation of Three Copper-Zinc Superoxide Dismutase Homologs Localized in
Different Cellular Compartments in Arabidopsis
黃建勛
Chien-Hsun Huang
指導教授:靳宗洛 博士 Tsung-Luo Jinn, Ph.D.
中華民國 101 年 3 月
致謝
CCS-independent pathway 的發表,就結果來說並不驚人,但其過程對我來說卻是一段曲 折迂迴、感觸良深的故事:
從碩班開始,我就對 SOD 的議題充滿興趣,只是當時朱瓊枝學姊的論文發表不久,實
驗剛好告了一個段落,並不清楚未來研究方向的潛力,因此便與文鈺兩人朦朦懂懂的胡亂摸
索,我本人除了CuZnSOD 的相關研究,還同時做過 MnSOD、葉形突變株、蕃茄耐熱開花系
統等研究,當時與文鈺懷抱著極大熱情,不分假日晝夜全心投入於實驗中,同時投資許多不 同議題的研究,就是希望哪一個議題能首先出現有發表價值的結果,當時比較領先的研究其
實是MnSOD 的部分,然而就在直昇博班沒多久,MnSOD 的議題就被別人搶先發表了,當時
受到不小的打擊,心灰意冷、茫然不知所措之下,決定先去當兵離開想想。這段時間,有賴
文鈺對 CuZnSOD 的議題以酵母菌異位表現的方式繼續研究,結束兵役回來,感到這些進展
是有發表價值的,因此日後就完全集中在 CuZnSOD 的研究上。只是後續的研究並不如想像
中的順利,先是發現原來前人認為的三個CuZnSOD 的活性 band 是錯誤的,再發現植物體內
的SOD 活性強弱與性狀並無關聯,加上 GSH 參與的不確定性,諸如此類在其他物種未見,
與現有發表知識產生極大衝突、難以解釋的結論,導致我們的研究結果很難被其他 reviewer
接受,時常多所質疑與否定,因此,前前後後共投了 7 次,其中有許多不足為外人所道的辛
酸。直至同樣矛盾的現象也陸陸續續的被其他研究室所揭露,才開啟了他人正視我們研究成 果的契機。
這段時間,真的要非常感謝靳宗洛老師對我們的支持,從實驗開始,老師就大膽的放手 讓我們自己去嘗試,其中因為經驗不足,或是嘗試一些異想天開的奇思妙想,免不了浪費不
少實驗室的經費,而在 paper 屢投不上、被別人批評的一文不值的時候,也真的很感謝老師
仍然相信並且肯定我們的價值,使我們在萬般沮喪之中,仍然存有信心繼續嘗試,努力突破,
也才能有今天的成果,所以在此真的要說:『老師,謝謝你!』;此外,文鈺一路走來都是一 同並肩作戰的伙伴,實驗時互相爭論思考,沮喪失意時互相扶持,每一道難關都有文鈺協同
的足跡,因此沒有文鈺可能就沒有這篇論文的存在,只是在我因為這篇 paper 而達成了畢業
標準的時候,文鈺FeSOD 的 paper 還在 reviewer 熊熊砲火的攻擊之下,致使我們無法一起畢
業,一同享受成功的喜悅,實在深感遺憾。在此要說:『女王,真的謝謝你,加油!』;至於 在我們困苦的時候,給予安慰、建議、打氣的諸多老師及同學,建勛在此表達誠摯的感謝之 意,也希望大家都能身體健康,萬事如意!
人生的許多苦難磨練著我們,在失意之中我們往往怨懟惱怒、傷心委屈,只是儘管如此,
我們必須學會放下怨恨,才能懷抱希望;必須撫平傷痛,才能興起感激。痛苦是會過去的,
成功也不是永恆的,因此只願我們一路行來,能夠忘卻曾經的不如意,只記得美好的部分,
在愛與被愛之中,滿懷感恩的待人處世,慈憂眾生。
在此以一首詩,作為這段日子的紀念:
總有一些光
會抵達明日的居所 比鳥雀比黎明 依稀更早 總有一些光
會寫信給昨天的暗影 在透明的窗前
留下枝椏扶疏的日記 在天明時刻
鐵鳥斂著羽翅 思索海的句子 事物因此柔軟 有了陰影
雲朵吹開了黑暗 不需拆卸不需抵擋 總有一些殘瓦
會被顫抖的光陰擊中 落入雪中深埋
這是冬天也不是冬天 那裡有一顆晨星 帶著亡逝的荒塚與愛 迴蕩在廢棄的昨日 在躍動的海洋
櫻花木悄悄然的盛開了 即使沒有樹葉也揚起了風帆
TABLE of CONTENTS
Page Abstract in Chinese ………I Abstract in English ………II Abbreviation ………IV
Introduction ………1
Superoxide Dismutases: Classification and Localization ………1
Evolution of Different Types of SODs ………2
Regulation of SODs Expressions by Their Promoter Sequences ………....4
Copper-Zinc SOD ………....5
Arabidopsis CuZnSODs are regulated by Copper-regulated miR398 ………...….6
SOD Activation Requires a Metallocheprone ………....7
Previous Studies on the Function of CCS ………....7
Discovery of Arabidopsis CCS ………...….8
CCS-Independent Activation of CuZnSOD ………...….8
Aims of the Dissertation ………...…10
Material and methods ...11
Plants, Yeast Strains, Media and Growth Conditions ………...11
Protein Extraction, In-gel SOD Activity Assay and Immunoblotting ………...11
RNA Extraction, RT-PCR and Gene Cloning ………...12
Constructs ………...12
Treatments for Seed Germination Rate ………...13
Monitoring Superoxide Anion Level by Nitroblue Tetrazolium ………14
Treatments for Root Length Evaluation ………14
Lysine-Independent Aerobic Growth ………15
Protoplast Preparation and Transfection ………15
Glutathione Treatment and Quantification ………....16
Apo-CSD1 Protein Preparation and In Vitro Treatments ………17
Antibodies ………18
Statistical Analysis ………18
Accession Number ………18
Results ...19
CSD Retains Partial Activity in the Absence of AtCCS in Arabidopsis ………19
Activity Signals of CSD1 and CSD2 Are Partially Overlapped ………19
CSD1 and CSD3 Show CCS-Independent Activities in Yeast ………20
CSD2 Activation in Chloroplasts Requires CCS But Not in the Cytoplasm …………...22
CCS-Independent CSD1 Activation Occurs in the Cytoplasm but Not in Chloroplasts ………22
Activities of Both Peroxisomal and Cytoplasm-directed CSD3 Can Not Be Detected ………23
Effect of Glutathione upon CCS-Independent CSD1 Activity in Yeast ………23
Effect of Glutathione upon CCS-Independent CSD Activities in Arabidopsis Flowers ………24 Effect of Glutathione upon CCS-Independent CSD Activities in Arabidopsis Flower Protein
Altering Glutathione Concentration by Drugs or Glutaredoxin Expression Affect CCS-Independent CSD Activities in Arabidopsis Protoplasts ………26 Activation of Apo-CSD1 by Cu and GSH is Greatly Enhanced When Atccs Cellular Extracts Is
Added in Vitro ………27
Superoxide Anion Level and Seed Germination Rate of WT, Atccs and Atcsd1 ………29 Root Length of Atccs and Atcsd1 under Oxidative Stress Treatments and Different Glutathione Concentrations ………30 CSD1 Variants Show Differing Activity Levels in Yeast ………32 Discussion ...34 Different Activation Preferences of CSD1 and CSD2 Are Due to an Inhibitory Effect of
CCS-Independent Activation in Chloroplasts ………34 The Inhibition of CCS-Independent Activation in Chloroplasts Could Ensure a Proper Operation
of the Photosynthesis System ………35 CSD3 Might be Activated Primarily by the CCS-Independent Pathway ………36 Different Preferences for CCS-Dependent and -Independent Activation Might Benefit Life in Its
Habitat ………37 CCS-Independent CSD Activities Are Physiologically Functional and Sufficient to Support Growth of Plant Cells ………38 CSD1 variants activated by CCS-independent way show different activation efficiency …40 GSH Is Involved in CCS-Independent Activation Pathway with the Involvement of an Essential
Factor Remain to be Discovered ………41 Models of How the Unknown Factor Involves In the CCS-Independent Pathway ………42
C-Terminal of CSD Protein May Determines Its Activation by CCS or the Unknown Factor
………43
Perspective ………45
Figures and Table ...49
Appendix ………81
References ...85
摘要
超氧歧化酶(Superoxide Dismutase;SOD),可將超氧分子轉變為過氧化氫及氧分子,具 有解除氧化逆境的功能。對於銅鋅超氧歧化酶(CuZnSOD;CSD)的活化機制,目前已知有兩 條路徑,一者是藉由銅鑲嵌輔助蛋白(Copper Chaperone of SOD1;CCS)的幫助,達到銅離子 鑲嵌與形成內生性雙硫鍵的活化型態;另一者則是在人類及老鼠中發現,在沒有CCS情況下,
CuZnSOD仍然具有少量的活性,只是其活化途徑與機制仍未明朗。本論文,將阿拉伯芥三個 不同的CuZnSOD基因表現於酵母菌及阿拉伯芥原生質體中,我們證實阿拉伯芥不同CuZnSOD 的活化具有不同的偏好:CSD1 位在細胞質中,可藉由兩條活化機制達成活化,在無CCS的情 況下,仍保有~36%的活性,類似人類的CuZnSOD;CSD2 位在葉綠體中,只能經由CCS來達 成活化,與酵母菌的CuZnSOD類似;CSD3 位在過氧化體中,只經由非CCS的路徑來達成活 化,類似線蟲的CuZnSOD。我們證實在AtCCS-knockout突變株中,此殘存的CuZnSOD活性量 就足以提供正常生理功能之所需。最後,我們也證實了還原態的榖胱甘肽(Glutathione;GSH) 參與在非CCS的活化CuZnSOD的路徑上,並且需有一未知功能的因子共同合作才能完成。我 們綜合前人之研究及本實驗之證據,提出兩種不需經由CCS而達成活化的可能作用機制,以 及提出CuZnSOD蛋白質之C端具有與未知因子交互作用,而達成促進活化的功用。綜上而言,
我們的研究提出植物體之複雜精密的抗氧化途徑,是在其他物種中從所未見的,而其詳細的 機制,則仍有待後續的研究加以釐清。
ABSTRACT
Superoxide dismutases (SODs) are enzymes that protect cells from oxidative damage. The major pathway for CuZnSOD activation involves the function of a Copper Chaperone for SOD (CCS), whereas an additional, minor CCS-independent pathway that has been observed in mammals.
Through overexpression of three Arabidopsis CuZnSOD genes (CSDs) in yeast and Arabidopsis protoplasts, we demonstrate the existence of a CCS-independent activation pathway in Arabidopsis
thaliana. Interestingly, the three Arabidopsis CSDs show strongly different preference for the two
activation pathways: the main activation pathway for CSD1 in the cytoplasm involved a CCS-dependent and -independent pathway, which was similar to that for human CSD. Activation of CSD2 in chloroplasts depended totally on CCS similar to yeast (Saccharomyces cerevisiae) CSD.
Peroxisome-localized CSD3 via a CCS-independent pathway was similar to nematode (Caenorhabditis elegans) CSD in retaining activity in the absence of CCS. The residual SOD activity detected in AtCCS knockout plants is sufficient for seed germination and root growth, confirming that this alternative pathway is physiologically functional. Through a series of glutathione manipulation experiments, we further confirmed that glutathione plays a role in CCS-independnet pathway but must cooperate with an unknown factor for SOD activation.
According to previous publications and our finding, two models of the CCS-independent mechanism are proposed. We also suggest that the CSD protein conformation at C-terminal is
important in providing a docking site for unknown factor to interact with. Our findings reveal a complex system underlying CSD activation which ensures a highly specific and sophisticated regulation of antioxidant pathways in plants and has not been reported in other organisms. However, the clear and definite mechanism needs further investigation.
ABBREVIATIONS BH : tert-butyl hydroperoxide
BSO: L-buthionine sulfoximine
CCS: copper chaperone for superoxide dismutase
CDNB: 1-chloro-2,4-dinitrobenzene
CSD: the innate Arabidopsis copper/zinc superoxide dismutase
CuZnSOD: copper/zinc superoxide dismutase
FeSOD: iron superoxide dismutase
GFP: green fluorescence protein
MV: methyl viologen
MnSOD: manganese superoxide dismutase
NiSOD: nickel superoxide dismutase
ROS: reactive oxygen species
RT-PCR: reverse transcription-polymerase chain reaction
SOD: superoxide dismutase
TEMED: N,N,N’,N’–tetramethyl-ethylenediamine
INTRODUCTION
Reactive oxygen species (ROS) are detrimental byproducts of aerobic reactions such as respiration and photosynthesis that can cause severe damage to numerous cellular constituents.
Superoxide dismutases (SODs) are a group of metalloenzymes that defend against free radical species by disproportionating O2 into H2O2 and O2 molecules (Beyer et al., 1991; Bowler et al., 1992).
Superoxide Dismutases: Classification and Localization
By a specific metal cofactor required for the superoxide scavenging activity (McCord and Fridovich, 1969), SODs are classified as Cu-Zn SOD (CuZnSOD), Fe SOD (FeSOD), Mn SOD (MnSOD) or Ni SOD (NiSOD) (Alscher et al., 2002; Zelko et al., 2002). Most eukaryotic cells contain more than 2 types of SODs, with CuZnSOD usually located in the cytoplasm, chloroplast, and probably extracellular space (Crapo et al., 1992); MnSOD located in mitochondria and peroxisome (Weisiger and Fridovich, 1973; Marres et. al., 1985); FeSOD located in chloroplast.
However, FeSOD can only be found in prokaryotes and plants but not in animals (Alscher et al., 2002).
In Arabidopsis thaliana, seven SOD genes have been identified (Kliebenstein et al., 1998).
FeSODs are located in the chloroplast, MnSODs in the mitochondrion, and CuZnSOD exists as
numerous isoforms that are distributed among various subcellular compartments (Jackson et al., 1978; Kanematsu and Asada, 1989; Bowler et al., 1992; Bueno et al., 1995). There are three CuZnSOD (CSD) genes: CSD1 which localizes to the cytoplasm, CSD2 which is found in chloroplasts, and CSD3 which is presumed to localize in peroxisomes (Kliebenstein et al., 1998;
Alscher et al., 2002). Because phospholipid membranes are impermeable to O2 (Takahashi and Asada, 1983), protection of different organelles can be achieved by the distribution of SOD to different cellular compartments. In green tissue, the greatest protein accumulation and activity are found for CSD1, CSD2, MSD1, and FSD1 in Arabidopsis (Kliebenstein et al., 1998).
Evolution of Different Types of SODs
This cladogram containing the protein sequences of all seven Arabidopsis SODs, and the protein sequences of SODs from a variety of other plant species, clearly supports the suggestion that the FeSODs are of greatest antiquity and that the CuZnSODs evolved independently of the Fe and MnSODs (Alscher et al., 2002). It implies that MnSOD and FeSOD are more ancient types, and these enzymes very probably have arisen from the same ancestral enzyme, whereas CuZnSODs have no similarity to MnSOD and FeSOD in sequence and probably have evolved separately in eukaryotes (Kanematsu and Asada, 1990; Smith and Doolittle, 1992).
Such evolution is probably related to the varied availability of soluble transition metal
compounds in the biosphere, which was due to the O2 content of the atmosphere in different geological eras (Bannister et al., 1991).When the atmosphere was fully replenished with oxygen, Fe (II) was almost fully unavailable whereas insoluble Cu (I) was converted into soluble Cu (II). At this stage, Cu (II) began to be used as the cofactor at the active sites of SODs. Transition from the use of iron to manganese only required little change in SOD protein structure since FeSODs and MnSOD show similar electrical properties. Thus, structures of MnSOD and FeSOD are very similar.
However, the electrical properties of CuZnSOD are greatly different from those of FeSOD and MnSOD. After Cu becoming a metal cofactor, a major change should occur in the structure of the protein (Bannister et al., 1991). Nowadays, CuZnSODs have been found mostly in eukaryotes while FeSOD and MnSOD are present both in prokaryotic and in eukaryotic organisms. However, CuZnSODs have been observed in some bacteria, including Caulobacter crescentus,
Photobacterium leiognathi, and pseudomonads. Three hypotheses might explain the presence of
CuZnSOD in prokaryotes: (1) CuZnSODs in prokaryotes and eukaryotes evolved independently; (2) Gene of CuZnSOD originated in the eukaryotes and was transferred into the prokaryotes. (3) CuZnSOD originated in prokaryotes, and then the prokaryotic gene was transferred to eukaryotes.
The third hypothesis depends on the unlikely requirement that prokaryotic and eukaryotic enzymes have a common ancestor that had CuZnSODs before the time that prokaryotes and eukaryotes separated. For the second hypothesis, it was first proposed by Martin and Fridovich (Martin and
Fridovich, 1981) and was support from Bannister and Parker (Bannister and Parker, 1985) by the reason of the 30% similarity between the protein sequences of the CuZnSODs in ponyfish and in its symbiont Photobacterium leiognathi. This similarity increased to 44% after taking point mutations into consideration, bringing more support to the hypothesis (Leunissen and de Jong, 1986).
However, the presence of CuZnSOD in Caulobacter crescentus and in pseudomonads that are not symbionts suggests the hypothesis requires further refinement (Steinman, 1982; Steinman, 1985).
Regulation of SODs Expressions by Their Promoter Sequences
The promoter sequences of SODs were related to the regulation of gene expression under different stresses. Four consensus sequences were revealed to have the ability to bind to four different transcription factors, respectively (Alscher et al., 2002). (1) The ABA responsive element (ABRE) appears to be associated with genes responding to osmotic stress (high osmoticum, salt, desiccation, and cold) and binds to several similar sequences of eight nucleotides (Choi et al., 2000;
Guan and Scandalios, 1998); the consensus sequence YACGTGGC was used. (2) NFκB is a transcription factor that activates immunoglobulinκ genes; the consensus sequence GGRNNYYCC was used (Smith et al., 2000). (3) The heat shock protein gene promoter consensus sequence is the palindromic sequence TTCNNGAA (Santos et al., 1996). (4) the Ybox motif has consensus sequence GATTGG and mediates redoxdependent transcription activation (Guan and Scandalios,
1998). In Table 1, diamonds (◆) summarize the exact or close matches found at upstream locations (within 1000 nucleotides of the ATG where transcription begins). The consensus sequences analysis revealed that different SODs may express under different stresses, which indicate they may have different roles in Arabidopsis.
Copper-Zinc SOD
CuZnSOD is a homodimeric copper- and zinc-containing enzyme (McCord and Fridovich, 1969). Zinc is required for the structural integrity of the protein and influences enzyme activity, whereas copper plays a catalytic role in the disproportion of superoxide (Forman and Fridovich, 1973; Beem et al., 1974). The acquirement of both metal ions was first assumed to be by passive diffusion; however, Rae et al. (1999) found that the intracellular concentration of free copper is undetectable under normal physiological conditions, which suggests that cells require copper chaperones to facilitate copper transfer to specific partners (see below).
CuZnSODs are found throughout the plant cell. There are two different groups of CuZnSODs, which are homodimeric and homotetrameric (Bordo et al., 1994). The active sites of each subunit function independently. When these subunits are separated and coupled with an inactive subunit, newly formed enzymes show full activity, which provides evidence that the interactions between the subunits are not essential for full catalytic activity (Fridovich, 1986).
There are chloroplastic and cytosolic forms of CuZnSOD. Deduced amino acid sequences of these two isoforms show approximately 68% similarity, whereas there is approximately 90%
similarity among the chloroplastic CuZnSODs (CuZnSODchl) and 80–90% similarity among the cytosolic CuZnSODs (CuZnSODcyt). The occurrence of a peroxisomal CuZnSOD from watermelon was also previously reported, which represented about 18% of the total SOD activity in the cell (Sandalio and del Río, 1987). Presence of such a peroxisomal CuZnSOD also was shown in rat liver cells (Dhaunsi et al., 1992).
Arabidopsis CuZnSODs are regulated by Copper-regulated miR398
A microRNA, miR398, is shown to regulate CSD1 and CSD2, targeting CSD1 and CSD2 mRNA for degradation during growth on low Cu condition (Sunkar et al., 2006; Yamasaki et al., 2007; Dugas and Bartel, 2008). miR398 is one of several Cu-regulated microRNAs (the Cu-microRNAs) that down-regulate transcripts for a number of Cu proteins together during Cu-limited growth (Abdel-Ghany and Pilon, 2008). The Cu-microRNAs are regulated by a transcription factor called SPL7 (Yamasaki et al., 2009), a homolog of Chlamydomonas rheinhardtii CRR1, which is possibly a Cu-sensing protein (Kropat et al., 2005). It was proposed that the mechanism of Cu-protein down-regulation during Cu-limited condition allows for preferential delivery of Cu to plastocyanin (Burkhead et al., 2009), which is essential for electron transport and
survival of higher plant (Weigel et al., 2003)
SOD Activation Requires a Metallochaperone
The metal cofactors for SODs are transition metals, whose ability to readily accept or donate an electron are utilized in the superoxide dismutation process. In free form, however, these metal ions produce potentially harmful hydroxyl radicals via the Haber-Weiss reaction (Halliwell and Gutteridge, 1989). Hence, it is generally accepted that free transition metals must exist in a complexed form and that SOD activation should require a metallochaperone. It has also been reported that in spite of the high affinity of SOD1 for copper (dissociation constant = 6 fM) and the high intracellular concentrations of both SOD1 (10 μM in yeast) and copper (70 μM in yeast), the copper chaperone for the superoxide dismutase (CCS) gene is still necessary for expression of an active, copper-bound form of superoxide dismutase (SOD1) in vivo (Rae et al., 1999).
Previous Studies on the Function of CCS
The first metallochaperone to be identified was the copper chaperone for SOD1 (CCS) in yeast Saccharomyces cerevisiae (yCCS) (Culotta et al., 1997). Orthologs of this protein have been found in many species (Abdel-Ghany et al., 2005; Chu et al., 2005). It is known that CCS consists of three protein domains (I, II, and III). The central domain II resembles SOD1 and serves to dock CCS with
SOD1 (Schmidt et al., 1999). Once the CCS-SOD1 heterodimer formed, copper insertion and disulfide oxidation may proceed via a CXC copper-binding motif at the C-terminal CCS domain III.
In the CCS-SOD1 docked complex structure, CCS domain III Cys229 forms an intermolecular disulfide with Cys57 of SOD1, which is believed to represent the intermediate in forming the SOD1 intramolecular disulfide. Domain I harbors a single CXXC copper-binding motif, it may help CCS dock with an upstream source of copper. To sum up, CCS protein physically interacts with CuZnSOD, thereby assisting in copper incorporation and the catalysis of disulfide bond formation, and thus resulting in an active SOD (Casareno et al., 1998; Lamb et al., 2001; Brown et al., 2004;
Furukawa et al., 2004).
Discovery of Arabidopsis CCS
In Arabidopsis, Pilon et al. (2005) has reported that AtCCS is a functional homolog of the yeast copper chaperone Ccs1/Lys7. After that, Chu et al. (2005) also found that a single CCS (AtCCS) is capable of activating three CSDs in different compartments. Nevertheless, the phenotype of a CCS-knockout mutant (Atccs) has been found to be normal (Cohu et al., 2009).
These data indicate the existence of additional activating factors for CSDs.
CCS-Independent Activation of CuZnSOD
A CCS-independent activation pathway for CuZnSOD had been observed in mice and further studied in a yeast expression system (Wong et al., 2000; Subramaniam et al., 2002; Carroll et al., 2004). To date, yeast SOD1 (ySOD1) activation has been found to fully depend on yeast CCS (yCCS; Carroll et al., 2004), whereas the nematode (Caenorhabditis elegans) CSD (wSod-1) is exclusively activated independently of CCS (Jensen and Culotta, 2005). Human CSD (hSOD1) is largely activated by CCS but retains about 25% to 50% of its activity in the absence of CCS (Carroll et al., 2004). However, CCS-independent pathways for SOD activation in plants are not well understood
Reduced glutathione (GSH) was required for CCS-independent activation of hSOD1 in yeast CCS-mutant strains with defective GSH metabolism, and wSod-1 was inactive in the presence of CCS when GSH was depleted in yeast (Carroll et al., 2004; Jensen and Culotta, 2005). However, a direct interaction between GSH and CSD has yet to be demonstrated. Results from mutagenesis studies showed that amino acid residues 142 and 144 near the carboxyl terminus of human and yeast CSDs were important in the CCS-independent pathway (Carroll et al., 2004; Jensen and Culotta, 2005). When these residues were replaced by those of dual prolines in ySOD1, CCS-independent activities for both hSOD1 and wSod-1 were inhibited. A recent study further confirmed that the proline at residue 144 but not 142 restricted ySOD1 disulfide formation in the absence of CCS, which played a key role in blocking CCS-independent activation (Leitch et al.,
2009a).
Besides, it has demonstrated that CCS activation of SOD1 requires molecular oxygen, whereas there is no similar oxygen dependence with CCS-independent activation (O’Halloran et al., 2004).
Hence, CCS-independent activation allows SOD1 activity to be maintained over a range of oxygen conditions. This can be particularly critical in tissues of multicellular organisms where oxygen tensions range from near atmospheric to hypoxic.
Aims of the Dissertation
In our current study, we analyzed Arabidopsis and yeast knockout strains to investigate the dependence of Arabidopsis CSD activity on CCS metallochaperone. We found both CCS-dependent and -independent activation pathways are present in Arabidopsis, and three Arabidopsis CSD proteins display unique levels of dependence on each of the two activation pathways. Moreover, we show that the chloroplast is unique in having lost CCS-independent activation ability. This phenomenon represents a novel finding as it has not been reported for any other species. We also suggest from our current data that glutathione is involved in the CCS-independent pathway in
Arabidopsis, with an additional factor cooperatively assisting in CSD activation
MATERIALS AND METHODS
Plants, Yeast Strains, Media and Growth Conditions
The Arabidopsis thaliana Atccs (At1g12520) and Atcsd1 (At1g08830) knockout lines of Columbia ecotype, Atccs (SALK_025986) and Atcsd1 (SALK_109389), were obtained from the ABRC (Ohio State University, USA) (Chu et al., 2005). Seeds were incubated at 4C for 4 d in the
dark before sowing, and then plants were grown in a growth chamber under 16-h light/8-h dark at 21 to 23C at a light intensity of 60 to 100 mol m-2 s-1. Saccharomyces cerevisiae BY4741 (MATa, his31, leu20, met150, ura30) was used as the yeast wild type (WT); sod1 (sod1::kanMX4)
and ccs (ccs::kanMX4) are derivatives of BY4741. Enriched yeast extract, peptone-based medium supplemented with 2 (w/v) Glc (YPD), and synthetic dropout leucine medium was used to propagate the yeast strains. Yeast was incubated at 30C under aerobic conditions without shaking, and G418 was used at 200 g mL-1 to maintain the ySOD1 and yCCS deletions in the yeast as required.
Protein Extraction, In-gel SOD Activity Assay and Immunoblotting
Arabidopsis crude protein was extracted with 150 mM Tris, pH7.2, as described previously (Chu et al., 2005). Yeast crude protein was extracted by the glass bead lysis protocol according to Culotta et al. (1997). Protein concentration was determined by the Bradford method (1976) with the
Bio-Rad protein assay reagent (Bio-Rad, CA, USA). In-gel SOD activity assay and immunoblotting were performed on 10 to 15 (wv) nondenaturing or denaturing gels, respectively, according to Chu et al. (2005). The SOD activities and protein signals were quantified by analyzing the activity gels and immunoblotting membranes with LAS-3000 (Fuji Film, Tokyo, Japan) and ImageQuant software (Molecular Dynamics, CA, USA).
RNA Extraction, RT-PCR and Gene Cloning
Total RNA was prepared with TRIZOL reagent (Invitrogen, CA, USA) and TURBO DNA-free Kit (Applied Biosystems, CA, USA). cDNA synthesis was performed by using high-capacity cDNA Reverse Transcription Kits (Applied Biosystems). RT-PCR and gene cloning was performed with the primers described in Supplemental Table 2 online.
Constructs
All cDNA fragments were amplified with PCR and then cloned at yTA vector (Yeastern Biotech, Taipei, Taiwan); subsequent construction was performed after DNA sequencing. For constructs used in yeast expression, all genes from ATG to stop codon were inserted at the HindIII site of the 2 LUE2 yeast shuttle vector pADNS (Colicelli et al., 1989), with the exception of CSD3, which was inserted at the NotI site. Point mutations of CSD1 were created by the
megaprining method with the primers described in Supplemental Table 2 online. For GFP and YFP fusion constructs, 326-GFPnt vector (Lee et al., 2001) and p35S-EYFP vector (see below) were used, respectively. For overexpression in protoplasts, genes were also subcloned into p35S-EYFP vector with a stop codon. A double 35S promoter from pPE1000 vector (Hancock et al., 1997) was cloned into the XhoIHindIII sites of pEYFP vector (Clontech, CA, USA), and the CSD1, CSD2 and CSD3 genes were then cloned into this p35S-EYFP vector at the EcoRIBamHI, HindIII, and EcoRI sites, respectively. For TP-CSD2, the 61 amino acids at the C terminus were deleted by using the primer described in Supplemental Table 2 online, and then inserted into p35S-EYFP at HindIII site.
Transit peptide of CSD2 was amplified using the primer described in Supplemental Table 2 online.
Then, TPCSD2 with EcoRINcoI sites, and CSD1 gene with NcoISalI sites, were ligated into p35S-EYFP at EcoISalI sites. For GFP fusion, CSD3 and CSD3-dAKL was inserted into 326-GFPnt vector at SmaISalI site. Both ROXY1 and GRXcp was inserted into SmaIBamHI sites of p35S-EYFP with stop codon. For recombinant protein, CSD1 gene was inserted into pGEX-6P-1 (Amersham Pharmacia Biotech) at EcoRI site.
Treatments for Seed Germination Rate
Sterilized seeds of the WT, Atccs and Atcsd1 were plated on 1/2 MS medium supplemented with 1% (w/v) Suc and pretreated with 4C in the dark for 3 d. Seeds were then incubated at 23C
under 16-h light8-h dark for 3 d, and seed germination rate was observed. Methyl viologen (MV;
Sigma, MO, USA) and tert-butyl hydroperoxide (BH; Fluka Chemical Co., WI, USA) was added to the 1/2 MS plates at the indicated concentrations.
Monitoring Superoxide Anion Level by Nitroblue Tetrazolium
This assay was according to Myouga et al. (2008). Three-week-old plants grown on 1/2 MS medium supplemented with 1% (w/v) Suc were harvested, weighed and then infiltrated with 6 mM nitroblue tetrazolium (NBT) in 10 mM sodium phosphate buffer (pH 7.1) at 400 to 500 mbar for 15 min. Plants were then illuminated with 800 mol m-2 s-1 for 1 h to produce color. Chlorophyll was removed by infiltration with ethanolchloroform (41, vv) at 400 to 500 mbar for 15 min for four times. Then, samples were solubilized in a mixture of 2 M KOH and DMSO at a ratio of 11.167 (vv) and vortexed at 4C for 1 h. After centrifugation at 13,000×g for 10 min, the NBT-formazan production was determined as the absorbance at 700 nm of the supernatant.
Treatments for Root Length Evaluation
Sterilized seeds of WT, Atccs and Atcsd1 were plated on 12 MS medium supplemented with 1% (w/v) Suc, and pretreated with 4C in the dark for 3 d. Plates were then incubated at 23C for 3 d under continuous light condition for germination, and the seedlings were transferred to 12 MS
plates with each treatment and incubated at 23C for another 4 d. After the root length was measured, the seedlings were collected to quantify the protein, total and reduced glutathione concentrations. Methyl viologen (paraquat, Sigma) and buthionine sulfoximine (BSO, Sigma) were added in the 12 MS plates at the indicated concentrations.
Lysine-Independent Aerobic Growth
Yeast strains were cultured in appropriate medium overnight at 30C without shaking. Then, cells were centrifuged, washed and resuspended in sterile water, and the absorbance at 600 nm was measured. For plate assay, the yeasts were serial diluted from OD 600 1 to 10-4, plated on synthetic dropout lysine medium, and incubated at 30C under aerobic conditions for 3 d. Results in Figures 3 and 8 were performed on the same plate. For the liquid assay, the yeast cultures were seeded from OD 600 0.01 in the lysine-lacking medium, incubated at 30C for 24 h under aerobic conditions, and then the cell density was determined by measuring the absorbance at 600 nm.
Protoplast Preparation and Transfection
Protoplast preparation and transfection were performed according to Yoo et al. (2007). The WT and Atccs plants grew on soil for 3 to 4 weeks, and the 5th to 8th leaves were chosen for protoplast preparation. About 106 protoplasts were transfected with 100 to 300 g plasmid DNA for each
construct by the PEG-calcium method. After transfection, the cells were incubated in WI solution at room temperature for 16 h under continuous light conditions. For protein extract of protoplasts, cells were collected, resuspended in 150 mM Tris buffer (pH 7.2), and vortexed five times for 5 s.
Cell lysate was then subjected to the SOD activity assay and immunoblotting.
Glutathione Treatment and Quantification
For treatments in yeast, L-buthionine sulfoximine (BSO; Sigma) was added to the medium at 1 mM for 16 h; 1-chloro-2,4-dinitrobenzene (CDNB, dissolved in 95 ethanol; Fluka) and reduced-form glutathione (GSH; Sigma) were added at 1 and 20 mM, respectively, for 2 h. Equal volumes of 95 ethanol or H2O were added as mock treatments. Atccs flowers were detached and immersed in CDNB or GSH solutions at room temperature for 3 or 1 h, respectively. After CDNB or GSH solutions were removed, the flowers were washed with water 3 times and then the crude protein was extracted. For treatments of protoplasts, Atccs protoplasts transfected with CSD1 were first incubated at room temperature for 16 h for CSD1 protein expression, and then 1 mM GSH or CDNB was added into the incubation medium for 15 min. Total glutathione and GSH concentrations were quantified with use of the Total Glutathione Quantification Kit (Dojindo Laboratories, Kumamoto, Japan) and the QuantiChrom Glutathione Assay Kit (BioAssay Systems, CA, USA), respectively. The resulting concentrations were normalized to the protein concentration
of the same sample, and the values of glutathione concentration are expressed relative to those of the mock treatments.
Apo-CSD1 Protein Preparation and In Vitro Treatments
Recombinant GST fusion proteins were affinity purified according to the Glutathione-Agarose user manual (Sigma). The GST tag was removed by PreScission Protease (Sigma). For preparation of inactivated Apo-CSD1 (Lepock et al., 1981), Holo-CSD1 was dialyzed against buffer containing 50 mM sodium acetate (pH 4.0) and 10 mM EDTA at 4C for 24 h, then against buffer containing 50 mM sodium acetate (pH 4.0) and 100 mM NaCl at 4C for 24 h, and finally against buffer containing 50 mM potassium phosphate buffer (pH 7.8) at 4C for 24 h. In the in vitro treatments, each reaction contained 870 ng Apo-CSD1 protein and 20 M ZnSO2 in PBS buffer (140 mM NaCl,
2.7 mM KCl, 10 mM Na2HPO4 and 1.8 mM KH2PO4, pH 7.3), and then 0.1 M CuSO4, 1 mM GSH or 15 g Atccs cellular extract were added into the mixture as indicated. After incubation at room temperature for 30 min, 1.5 mM EDTA was added into samples, and then native-PAGE was performed with 0.1 mM EDTA in both the nondenaturing-gels and running buffer. In the treatments with both Cu and GSH, the two solutions were mixed and incubated at 4C for 16 h before the experiment.
Antibodies
Anti-sera -CSD1, -CSD2 and -CSD3 were generous gifts from Dr. D. J. Kliebenstein (Kliebenstein et al., 1998), and Arabidopsis -RPN8 antibody was kindly provided by Dr. H.-Y. Fu (Yang et al., 2004). We also used antibodies of -ACT (Chemicon, CA, USA), yeast -PGK1 (Molecular Probes, OR, USA), anti-rabbit (Jackson ImmunoResearch, PA, USA) and anti-mouse IgG (Bethyl Laboratories, TX, USA) conjugated with alkaline phosphatase.
Statistical Analysis
All experiments were independently repeated at least three times. Statistical analysis was performed by using the Student’s t test (two-tailed, unpaired).
Accession Number
Sequence data from this article can be found in GenBank as described in Table 1.
RESULTS
CSD Retains Partial Activity in the Absence of AtCCS in Arabidopsis
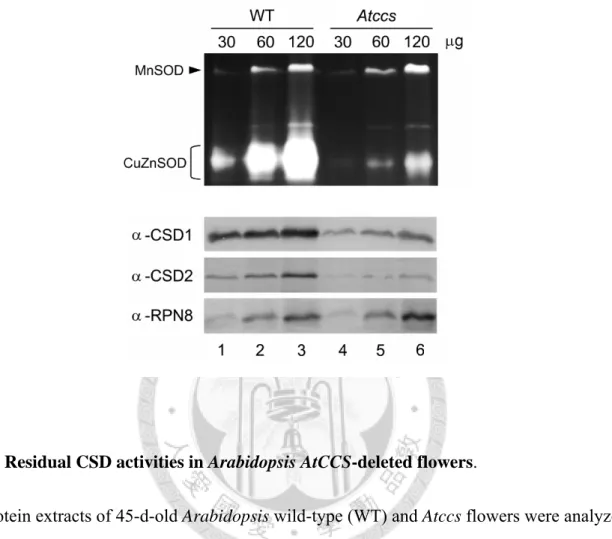
Previous studies have mapped the pattern of SOD activity among various Arabidopsis organs (Chu et al., 2005). The level of CSD activity was shown to be high in flowers and siliques but low in rosette leaves and cauline leaves. To determine whether Arabidopsis CSD can be activated in the absence of AtCCS, we analyzed CSD activities in the flowers of wild type (WT) and Atccs plants.
As shown in Fig. 1, residual CSD activity (about 6% to 30 of the WT levels) could be clearly observed in the Atccs mutant (top), indicating that a CCS-independent CSD activation pathway exists in Arabidopsis. Also of interest was our observation that the CSD1 and CSD2 protein levels in Atccs were lower than those in the WT (Fig. 1, bottom), whereas their mRNA levels were shown previously to be equivalent (Chu et al., 2005). This may indicate that the activated CSD protein is more stable than its inactivated counterpart.
Activity Signals of CSD1 and CSD2 Are Partially Overlapped
In previous study, there are two major CSD activity bands in the Arabidopsis WT protein extracts. The slow migration band was previously characterized as CSD2, whereas the fast migration band was CSD1 (Chu et al., 2005). We again characterized the 3 CSD protein and activity in this research. In the denaturing gel, the 3 overexpressed CSD proteins signals can be clearly
identified by their size (Fig. 2A). However, transient overexpression of 3 CSDs in protoplasts revealed that one of the CSD2 activity bands overlapped with that of CSD1 (Fig. 2B, lanes 1 and 2).
Thus, discriminating CSD1 and CSD2 activities by the in-gel SOD activity assay is difficult.
Notably, the CSD3 protein signal was clear when overexpressed in protoplasts, but its activity was undetectable (Fig. 2B, lane 3). We also could not detect the endogenous CSD3 protein signal by
-CSD3 antibody in the WT tissues (data not shown), which indicated that the endogenous CSD3 expression level was too low to be detected. Therefore, we cannot identify which CuZnSOD can be activated by the CCS-independent pathway through a direct observation on the SOD activity signals;
other experimental method must be taken.
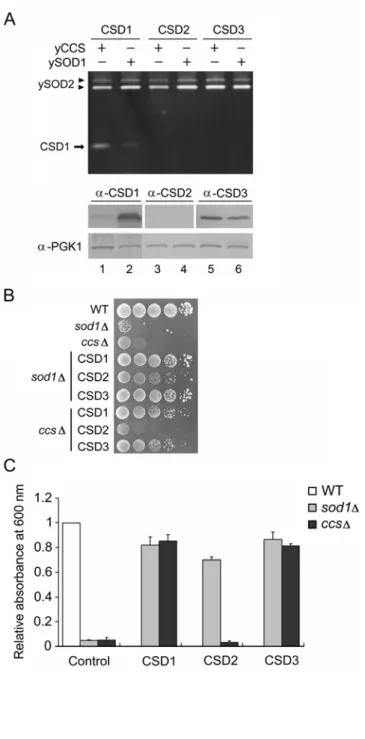
CSD1 and CSD3 Show CCS-Independent Activities in Yeast
AtCCS is a functional homolog of yCCS (Abdel-Ghany et al., 2005), implying that the
Arabidopsis CSD might be activated in a yeast system. As an additional method of determining
which CSD can be activated via the CCS-independent pathway, we expressed the three Arabidopsis CSD genes in yeast SOD1- and CCS-knockout strains (referred to as sod1 and ccs, respectively;
Fig. 3). As revealed by the in-gel SOD activity assay (Fig. 3A, top), the CSD1 activity band was strong in sod1 and was weaker, but clearly visible, in ccs (lanes 1 and 2), demonstrating a CCS-independent activation of CSD1. When CSD2 and CSD3 were overexpressed in yeast, no
detectable activity was observed (lanes 3 to 6), and the CSD2 protein was not detectable in either yeast line (Fig. 3A, lanes 3 and 4, bottom). Considering the limited sensitivity of the SOD activity and immunoblotting assays, we used lysine-independent aerobic growth as an indicator of the CSD activities in yeast in vivo because of its high sensitivity to SOD activity (Wallace et al., 2004).
Using this assay, lysine biosynthesis ability is maintained when ySOD1 activity just higher than 2
of wild type activity (Corson et al., 1998). Because ySOD1 activation is fully dependent on yCCS function, the endogenous ySOD1 activity is lost in ccs (Carroll et al., 2004), and the heterologous-expressed Arabidopsis CSDs should be exclusively responsible for the lysine biosynthesis ability in both sod1 and ccs. In our experiments, both sod1 and ccs were unable to grow under lysine-depleted conditions (Fig. 3B and 3C), which confirms the absence of ySOD1 activity in these two mutants. The lysine biosynthesis ability was recovered in both sod1 and ccs
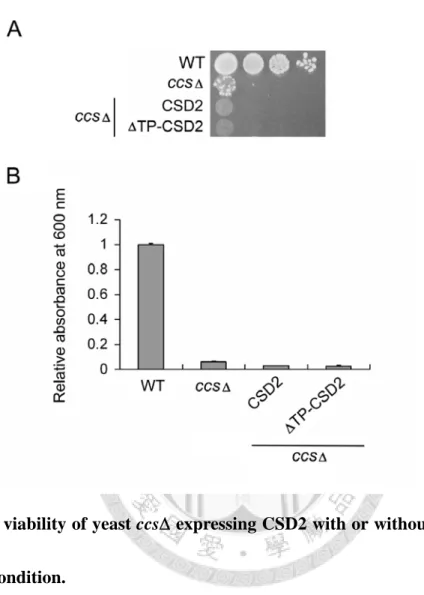
upon CSD1 and CSD3 expression. Upon the expression of CSD2, the phenotype of sod1 was partially rescued but that of ccs was not restored at all, thereby demonstrating a weak expression and activity of CSD2 in sod1. CSD2 is a chloroplast protein in Arabidopsis; because of no chloroplasts in yeast, we wondered whether the transit peptide might cause misfolding of CSD2.
Thus, we deleted the transit peptide of CSD2 (referred to as TP-CSD2) and expressed it in yeast;
however TP-CSD2 still showed no activity in ccs (Fig. 4). These results thus show clearly that CSD1 and CSD3 can both be activated via the CCS-independent pathway in yeast. However, the
failure of CSD2 to rescue lysine biosynthesis might be related to the heterologous nature of the expression system used. Thus, we sought additional means for determining whether CSD2 can be activated in a CCS-independent manner.
CSD2 Activation in Chloroplasts Requires CCS But Not in the Cytoplasm
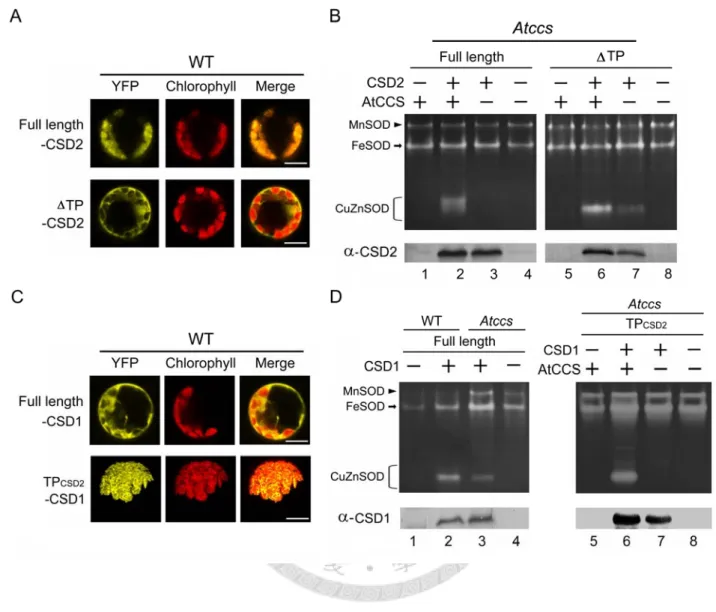
To address the question of how CSD2 is activated, CSD2 and AtCCS genes were transiently overexpressed in Atccs protoplasts (Fig. 5A and 5B). Use of yellow fluorescent protein (YFP) fused to CSD2 revealed that the full-length CSD2 localized in chloroplasts, whereas the transit peptide-deleted CSD2 (TP-CSD2) localized to cytoplasm (Fig. 5A). Full-length CSD2 was activated in the presence of AtCCS (Fig. 5B, lane 2) but was undetectable in the absence of AtCCS (lane 3). However, TP-CSD2 showed CCS-independent activity (lane 7), which indicates that CSD2 can be activated via the CCS-independent pathway when localized in cytoplasm.
CCS-Independent CSD1 Activation Occurs in the Cytoplasm but Not in Chloroplasts
To gain an understanding of CSD activation in chloroplasts, we constructed a chloroplast-directed CSD1 to observe its behavior in this compartment (Fig. 5C and 5D). We used YFP fusion proteins to confirm the localization of a CSD1 fused with the chloroplast transit peptide of CSD2 (TPCSD2-CSD1) in chloroplasts (Fig. 5C). We found CCS-independent CSD1 activity in
Atccs (Fig. 5D), and the efficiency of CSD1 activation (activity g-1 protein) in Atccs was 36 of that in wild-type (compare lanes 2 and 3). In contrast, chloroplast-localized CSD1 was active in the presence of AtCCS (lane 6), and its activity was undetectable in the absence of AtCCS (lane 7).
This result is consistent with the observation on CSD2 that CCS-independent CSD activation occurs in the cytoplasm but not in the chloroplast.
Activities of Both Peroxisomal and Cytoplasm-directed CSD3 Can Not Be Detected
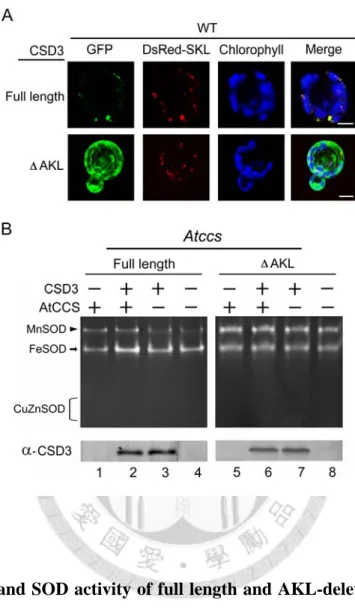
A similar experiment was also performed with CSD3 (Fig. 6), and green fluorescent protein (GFP) fused to N-terminal of CSD3 to show its localization (panel A). Our result showed that CSD3 localized in peroxisomes as predicted, but its activity were undetectable even when it was overexpressed in Atccs with AtCCS co-expression (Fig. 6B). Deletion of the peroxisome-targeting sequence AKL at its C-terminal (CSD3-AKL; Fig. 6A) leads to a cytoplasmic CSD3; however, unlike CSD1 and CSD2, there was still no activity detected (Fig. 6B).
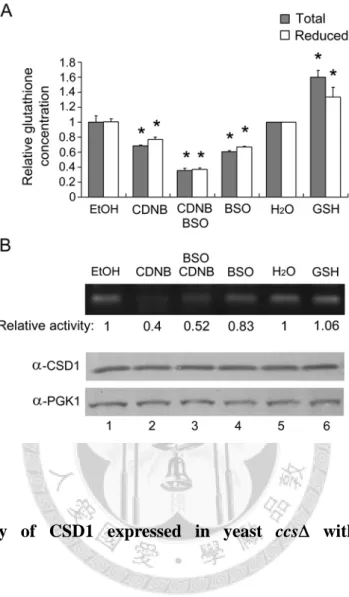
Effect of Glutathione upon CCS-Independent CSD1 Activity in Yeast
Previous studies have implicated GSH in the CCS-independent activation of human hSOD1 (Carroll et al., 2004). To investigate the possible involvement of GSH in the activation of Arabidopsis CSD, we treated a yeast ccs strain that expresses CSD1 with either GSH,
1-chloro-2,4-dinitrobenzene (CDNB, a glutathione chelator) or L-buthionine sulfoximine (BSO. a glutathione biosynthesis inhibitor), and then analyzed the effects upon CSD1 activities (Figure 7).
When treated with CDNB, CDNBBSO and BSO, the cellular concentrations of both total glutathione and GSH were significantly decreased to 0.68-, 0.36- and 0.6-fold, and 0.77-, 0.37- and 0.67-fold (Fig. 7A, treatments 2 to 4), respectively. The CSD1 activity was decreased to 0.4-, 0.52- and 0.83-fold respectively (Fig. 7B, lanes 2 to 4). Hence, decreased GSH levels correlate with a reduction in CCS-independent CSD activity. In addition, although the total glutathione and GSH concentrations were slightly enhanced upon GSH treatment by 1.6-fold and 1.33-fold (Fig. 7A, treatment 6), respectively, no corresponding increase in CSD1 activity was observed (Fig. 7B, lane 6).
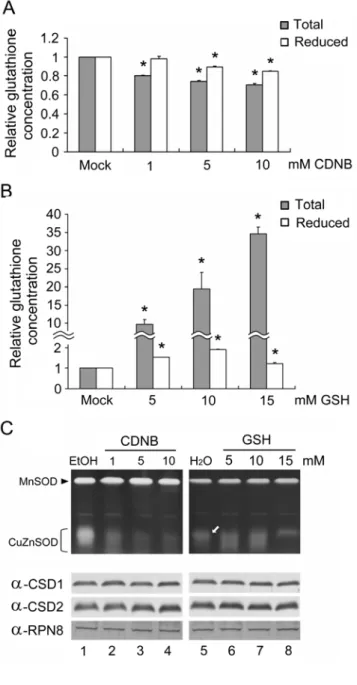
Effect of Glutathione upon CCS-Independent CSD Activities in Arabidopsis Flowers
A similar experiment was performed using Atccs flowers, which were treated with either CDNB (a glutathione chelator) or GSH, and then analyzed for CSD activity (Fig. 8). Both total glutathione and GSH concentrations in Atccs flowers decreased significantly as a result of the CDNB treatment (Fig. 8A), and the corresponding CSD activity was strongly downregulated (Fig.
8C, lanes 1 to 4). Following GSH treatment, the total glutathione concentration greatly increased, whereas the GSH concentration increased only slightly (Fig. 8B). The relatively mild change
observed in the internal GSH concentration might be due to an in vivo mechanism that maintains the homeostasis of reducing power, too much of which could be potentially damaging to cells (Lockwood, 2003; Pasternak et al., 2008). Xiang et al. (2001) have also shown previously that the maximal enhancement of GSH was only 2-fold in a -glutamylcysteine synthetase overexpression line, similar to the 2-fold increase in GSH concentration observed in our present system (Fig. 8B).
Although the bands were less well-defined, the major CSD activity band in the corresponding treatments seemed to show no enhancement (Fig. 8C, lanes 5 to 8, arrow).
Effect of Glutathione upon CCS-Independent CSD Activities in Arabidopsis Flower Protein
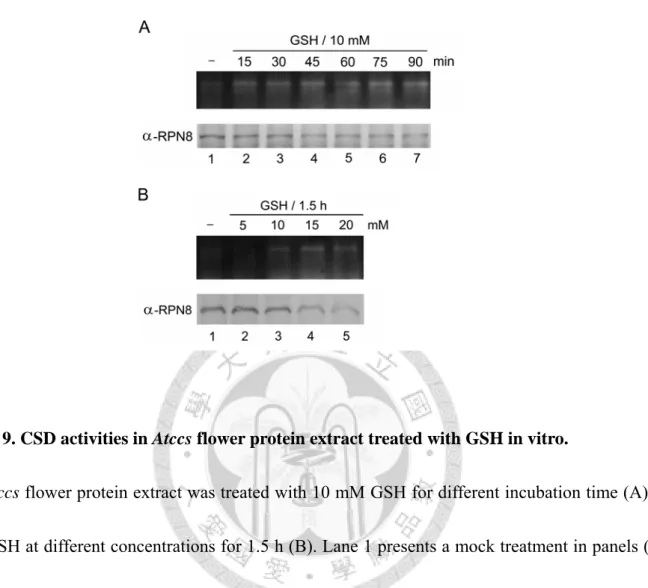
Extract
An in vitro experiment was performed to further investigate the effect of GSH on CCS-independent CSD activities. By adding GSH directly into the Atccs flower protein extracts, CSD activities were quickly enhanced in 15-min incubation with 10 mM GSH treatment (Fig. 9A), but the enhancement was still within certain limit even with 20 mM GSH treatment for 1.5 h (Fig.
9B). The limitation of the GSH-dependent activation in vitro might be due to (1) the CSD activities activated by GSH are less efficient, and thus only weak activities could be present at one time; and (2) cellular proteins showed decreasing accumulation and/or stability in cells with abundant GSH (Fig. 9A and 9B, bottom panel; Lockwood, 2003), and the lowered CSD protein levels might result
in the weak enhancement.
Additionally, we also curious about that under normal condition, whether these two pathways each account for partial activation of CSD and thus have their own activation niche. To answer this, we treated WT flowers with CDNB to see the changes of CSD activities (Fig. 10). CDNB was added directly into the flower protein extract for efficient depletion of glutathione. In this in vitro test, the CSD activities showed no significant change either treated with different CDNB concentrations (5 to 20 mM; Fig. 10A) or with longer incubation time (from 15 to 90 minutes; Fig.
10B). Hence, the activation niche accounted by CCS-independent pathway seemed negligible in WT, which implied that CSD activation by AtCCS was so efficient that a decrease in glutathione level could be ignored.
Moreover, when we treated WT flower protein extracts with 10 mM GSH for 18 h, CSD1 protein was mostly degraded, whereas CSD2 protein was still preserved (Fig. 11); compared to CSD1, CSD2 was not sensitive to GSH, and thus the effect of GSH on CSD2 seemed not efficient.
Altering Glutathione Concentration by Drugs or Glutaredoxin Expression Affect
CCS-Independent CSD Activities in Arabidopsis Protoplasts
The effects of GSH were also tested by altering its concentration in Atccs protoplasts transiently overexpressing CSD1 (Figure 12A). Cellular concentrations of total glutathione and
GSH were slightly enhanced upon GSH treatment by 1.23- and 1.13-fold, respectively, whereas the CSD1 activity levels showed no obvious increase in this system. Upon treatment with CDNB, both the concentrations of total glutathione and GSH decreased (to 0.42- and 0.87-fold), as did the amount of CSD1 and its level of activity.
In a further experiment, we manipulated the GSH concentration by overexpressing glutaredoxin, which oxidizes GSH to GSSG during its substrate-reducing reaction (Rouhier, 2010).
To this end, we overexpressed ROXY1 (a cytoplasmic glutaredoxin) or GRXcp (a chloroplastic glutaredoxin) together with CSD1 in Atccs protoplasts (Figure 12B; Xing et al., 2005; Cheng et al., 2006; Rouhier, 2010). Our results revealed that the GSH concentration decreased as a result of either ROXY1 or GRXcp overexpression, whereas the total glutathione concentration showed no significant change (Figure 12B). When ROXY1 was co-expressed, both the amount of CSD1 as well as its activity level was diminished, whereas the co-expression of GRXcp resulted in reduced CSD1 activity with no change in the amount of protein.
Taken together, our results show an overall decrease in CSD1 activity upon reduction of the intracellular GSH concentration. However the decrease in GSH concentration is not well correlated with that of CSD1 activity. Additionally, no obvious change in CSD1 activity was observed when the GSH concentration was increased.
Activation of Apo-CSD1 by Cu and GSH is Greatly Enhanced When Atccs Cellular Extracts
Is Added in Vitro
To gain a better understanding of GSH involvement in the CCS-independent pathway, we carried out in vitro analyses using purified recombinant proteins. We purified recombinant CSD1 protein as a GST fusion product (GST-CSD1), and then removed the GST tag by PreScission protease (generating Holo-CSD1). We then prepared inactivated Apo-CSD1 using an acid treatment method (Fig. 13A and 13B; Lepock et al., 1981). When Apo-CSD1 was treated with 0.1 M Cu (Fig. 14, lane 2), CSD1 itself was able to spontaneously bind copper and recover its own activity.
We then tested the effects of pH on this system (Fig. 13C), and found that the recovery of Apo-CSD1 activity was maximal at pH 7, the same pH used in our current experiments. Recovery of Apo-CSD1 activity was unsuccessful using 1 mM GSH alone, which was possibly due to the extremely low concentration of Cu in the mixture (Fig. 14, lane 3). However, treatment with both Cu and GSH did not result in greater CSD1 activity than treatment with Cu alone, indicating that GSH itself was either inefficient or possibly unable to facilitate CSD1 activation. When the same experiments were performed in the presence of Atccs cellular extracts (lanes 6 to 9), the activity of CSD1 treated with GSHextract increased slightly (lane 8) as compared with a cellular extract treatment alone (lane 6). However, CSD1 activity was greatly enhanced when GSH, extract and Cu were present in the reaction mixture (lane 9), as compared with only Cu and extract (lane 7). This
indicates that GSH can activate CSD1 efficiently under these conditions. The fact that Atccs cellular extract can greatly affect the level of CSD1 activation by GSH indicates that the interaction between GSH and CSD1 is facilitated by a yet undetermined factor.
Superoxide Anion Level and Seed Germination Rate of WT, Atccs and Atcsd1
We endeavored to determine whether these residual CSD activities were physiologically functional in vivo by using the seed germination rate as a phenotypic indicator to compare WT,
Atccs and CSD1 knockout lines (Atcsd1) after treatment with methyl viologen (MV) and tert-butyl
hydroperoxide (BH). We used MV and BH to induce oxidative stress in cells and thereby magnify the phenotypic effects due to the defects in antioxidant enzymes such as SOD. In the homologous T-DNA-inserted Atcsd1 knockout line, CSD1 mRNA and protein were absent, whereas the expression levels of CSD2 and CSD3 were unchanged (Fig. 15A to 15C). As visualized using an in-gel SOD activity assay, the total CSD activity in Atcsd1 (with functional CSD2 and CSD3) was lower than that in the WT but was higher than that in Atccs (Fig. 15D). This result correlated well with the superoxide anion accumulation level. The relative superoxide anion levels were Atccs Atcsd1 WT (Fig. 15E), indicating that the level of total SOD activity among the three strains was WT Atcsd1 Atccs.
When grown on 12 MS plates, the seed germination rates of the Atccs and Atcsd1 mutants
were similar to WT (Fig. 15F, control), indicating that the CCS-independent activities in Atccs are sufficient for normal growth. With 0.008 or 0.04 M MV treatments, the seed germination rate of
Atccs was significantly lower than that of the WT, but was higher than that of Atcsd1 (Fig. 15F).
Similar results were obtained with a 100 M BH treatment. Hence, although the total CSD activity in Atccs was found to be lower than that in Atcsd1, the phenotype of Atccs (resulting from the CCS-independent CSD activity) is less pronounced than that of Atcsd1 (resulting from total CSD2 and CSD3 activities).
Root Length of Atccs and Atcsd1 under Oxidative Stress Treatments and Different Glutathione
Concentrations
Root length was also measured to compare the phenotype of WT, Atccs and Atcsd1 on treatments with MV and BSO. When Atccs was grown on 12 MS without paraquat treatment, the root length did not differ from that of the WT (Fig. 16A, 0 M paraquat), which indicates that the residual CSD activities in Atccs were sufficient for plant growth under normal conditions. This finding might also explain why we could not find an obvious phenotype in Atccs. However, the root length of WT and Atccs was significantly longer than that of Atcsd1. Under 0.008, 0.04 and 0.2 M paraquat treatment, the root length of Atccs was shorter than that of the WT but was still longer than that of Atcsd1 (Fig. 16A). Therefore, although the CSD activities in Atccs were lower than that in
Atcsd1, the antioxidant ability resulting from these residual CSD activities in Atccs was better than
that in Atcsd1, which has CSD2 and CSD3 activities. These findings suggest that SOD activities in chloroplasts and peroxisomes cannot efficiently complement the function of the cytosolic SOD. The level of SOD activity should not be the only critical point for the resulting antioxidant ability in vivo;
whom and where the activities originated were also considerable. We concluded from Fig. 16A that although the 3 CSD activities were diminished in Atccs, the residual CCS-independent activities could provide significant protection to cells against oxidative stress. As well, even though the antioxidant ability in Atccs was weaker than that in the WT, it was sufficient for normal growth.
However, could the better antioxidant ability of Atccs (compared to Atcsd1; Fig. 16A) be weakened by glutathione depletion? To answer this, BSO was used to block glutathione biosynthesis in cells; 0.4 mM BSO effectively decreased the internal total and reduced glutathione amount to 16 and 60 (Fig. 16C and 16D), respectively. It has been reported that BSO induces root cell elongation and results in longer root length (Sánchez-Fernández et al., 1997). This can be observed in our treatments that root length became longer when BSO was treated (Fig. 16, comparing B to A), which indicates that the treatments were successful. Notably, with BSO treatment, the root lengths of Atccs and Atcsd1 became similar and were both shorter than that of the WT (Fig. 16B, 0.008 to 0.2 M paraquat). The root of Atccs was originally longer than that of
Atcsd1 with a normal cellular glutathione level (Fig. 16A) but became similar to Atcsd1 with
decreased glutathione level (Fig. 16B). These findings demonstrate that the better antioxidant ability in Atccs conferred by CCS-independent CSD activities were diminished with decreased glutathione concentration, which supported again that internal glutathione was required for CCS-independent activation.
CSD1 Variants Show Differing Activity Levels in Yeast
Previous studies by Carroll et al. (2004) have demonstrated the importance of residues 142 and 144 in the human hSOD1 protein for CCS-independent activation, and showed that substitution of these amino acids with proline prevents activation by this pathway. By aligning CuZnSOD gene sequences in A. thaliana, H. sapiens, S. cerevisiae and C. elegans (Figure 17A), the corresponding residues in Arabidopsis CSD were found to be 141G143V in CSD1, 143G145L in CSD2 and 147S149V in CSD3. We generated a series of CSD1 mutants: G141AV143A (denoted CSD1-AA, nematode form), G141SV143L (CSD1-SL, human form) and G141PV143P (CSD1-PP, yeast form) to determine the effects of these residues on the CCS-independent activation of CSD1. Each of these variants was expressed individually in yeast sod1 and ccs strains.
The activities of WT CSD1 (CSD1-GV) and CSD1-SL were detectable in both sod1 and ccs, while the activity of CSD1-AA was only measurable in sod1 (Fig. 17B, lanes 1 to 6). Under lysine-depleted conditions, CSD1-GV and CSD1-SL can restore the lysine-auxotrophic phenotype
of sod1 and ccs (Fig. 17C and 17D). Upon CSD1-AA expression, the viability of sod1 was restored but that of ccs was only partially rescued. Although the activity of CSD1-PP was undetectable in both sod1 and ccs as determined using an in-gel SOD activity assay (Fig. 17B, lanes 7 and 8), the activity of this mutant was discernable using the lysine biosynthesis assay in sod1 but not in ccs (Fig. 17C and 17D). In summary, the Arabidopsis form CSD1-GV, the human form CSD1-SL and the nematode form CSD1-AA can be activated by the CCS-independent pathway, but the yeast form CSD1-PP does not undergo CCS-independent activation.
DISCUSSION
Different Activation Preferences of CSD1 and CSD2 Are Due to an Inhibitory Effect of
CCS-Independent Activation in Chloroplasts
As characterized in both yeast and Arabidopsis protoplasts, it is clear that CSD1 can be activated by both pathways (Figs. 3 and 5D). In contrast, CSD2 could not be activated by CCS-independent pathway in yeast or Arabidopsis (Figs. 3 and 5B). Thus, ySOD1 and CSD2 might be activated by a similar mechanism. However, further investigation revealed significant differences between the 2 orthologs. The C-terminal amino acid residues 142P144P of ySOD1 prevented its activation by the alternative pathway (Carroll et al., 2004), and this effect is mediated primarily by 144P (Leitch et al., 2009a); when 144P is mutated to 144L, this ySOD1 variant showed clear activity in the absence of CCS. For CSD2, the amino acid residues corresponding to 142P144P of ySOD1 are 143G145L (Fig. 17A), which are typical for a CSD protein capable of being activated by the CCS-independent pathway (Carroll et al., 2004; Leitch et al., 2009a). The CSD2 protein can be activated by this pathway because the TP-CSD2 (cytoplasm-localized CSD2) was active in
Atccs (Fig. 5B). Thus, the lack of CCS-independent activity of CSD2 may not result from the
protein structure but rather the presence of factor(s) in the chloroplast that inhibit the CCS-independent activation pathway. This hypothesis is supported by the results obtained with the TPCSD2-CSD1 (chloroplast-directed CSD1), which also lost CCS-independent activity (Fig. 5D).
The inhibitory effect on CCS-independent CSD activity in chloroplasts may be due to the absence of chloroplastic factor(s) that are required for CCS-independent activation or to the presence of inhibitory factor(s) in the chloroplast, a novel phenomenon that has not been reported so far for any other species.
The Inhibition of CCS-Independent Activation in Chloroplasts Could Ensure a Proper
Operation of the Photosynthesis System
In contemplating the reason for the inhibition of the CCS-independent pathway in chloroplasts, oxygen is one factor worthy of consideration. In CCS-dependent activation, oxygen is required during the disulfide formation between the interacting CCS and CSD proteins (Brown et al., 2004;
Furukawa et al., 2004), such that activation cannot occur under hypoxic conditions. Thus, activation by CCS in the chloroplast implies a constant supply of oxygen, which is produced from electron transport chain of the photosynthetic machinery. Such a mechanism can ensure that CSD activation follows the presence of reactive species and oxidants which are produced from a properly functioning photosynthesis. In contrast, CCS-independent activation of CSD does not require oxygen (Leitch et al., 2009a). An additional factor might be related to the fact numerous photosynthetic enzymes require copper as their cofactor (Shcolnick and Keren, 2006). When photosynthetic enzymes are saturated with their copper cofactors, abundant oxygen will be
produced from photosynthesis reactions, which might constitute a signal for CCS to activate CSD.
An antioxidant system regulated by oxygen could guarantee that distribution of copper to photosynthetic enzymes takes priority in chloroplasts.
CSD3 Might be Activated Primarily by the CCS-Independent Pathway
Although the CSD3 activity was undetectable by the in-gel SOD activity assay in yeast, it could complement the phenotypes of both yeast sod1 and ccs (Fig. 3), which indicates the presence of a physiologically significant level of CSD activity. In these experiments, the activity in ccs could have occurred only through the CCS-independent pathway. However, sod1 contains
factors that can facilitate both CCS-dependent and -independent activation, so phenotype recovery by CSD3 in this system may have occurred by either or both pathways. Thus, these results demonstrate only that CCS-independent activation of CSD3 has occurred, not the level of CSD3 activity that can be conferred by CCS.
To further characterize the activation of CSD3 in a homologous plant system, we did not observe activation with overexpression of the full-length CSD3 in Arabidopsis protoplasts.
Considering the possible effects of localization similar to CSD2, we deleted its peroxisomal-targeting sequence but still found no activity even with AtCCS co-expression (Fig. 6).
Kliebenstein et al. (1998) demonstrated similar immunoblotting signals with the same amount (12
ng) of recombinant CSD1, CSD2 and CSD3 protein when probed with -CSD1, -CSD2 and
-CSD3 antibodies, respectively. Thus, the amount of overexpressed CSD3 protein in protoplasts should be similar to that of overexpressed CSD1 and CSD2 seen in Fig. 5. Hence, our failure to detect CSD3 activity suggests that its level of activation by CCS is much lower than that of CSD1 and CSD2. In addition, the level of CSD3 activation associated with both pathways (expression in
Atccs with AtCCS co-expression or in wild-type protoplasts) seems not to be significantly higher
than the levels associated with the CCS-independent pathway only (expression in Atccs). Therefore, CSD3 may be activated primarily by the CCS-independent pathway. Notably, the activation of nematode wSod-1 in the presence of CCS did not result in a higher activity than that by the CCS-independent pathway (Jensen and Culotta, 2005). In this respect, CSD3 seems to be similar to the nematode wSod-1.
Different Preferences for CCS-Dependent and -Independent Activation Might Benefit Life in
Its Habitat
To date, studies of CSDs in yeast, nematode, mice and human have shown different preferences for CCS-dependent and -independent activation (Carroll et al., 2004; Jensen and Culotta, 2005; Leitch et al., 2009b). However, our research find a very interesting issue that in Arabidopsis that different types of CSDs are present in a single cell (Fig. 18): cytoplasmic CSD1, like human
form, is activated mainly depending on CCS and partially by the CCS-independent pathway;
chloroplastic CSD2, like yeast form, activation depends completely on CCS, with inhibition of the alternative pathway in chloroplasts; and peroxisomal CSD3, like nematode form, is activated mainly by the CCS-independent pathway (see previous discussion).
Among the species that have been studied thus far, their habitats are highly varied, so the different preferences for CCS-dependent and -independent activation might imply that the two pathways have advantages in different types of environments. Why different organisms have evolved different preferences for CCS-dependent and -independent activation is unclear but may reflect unique lifestyle requirements for Cu and O2 (Culotta et al., 2006), which is consistent with our suggestion for the inhibition of CCS activation in chloroplasts. The functions of CCS might be limiting under certain conditions, and the existence of an alternative, CCS-independent pathway might be helpful for survival. Because plants cannot move to avoid environmental stresses, the complex activation mechanisms for plant CSDs might be a solution to cope with such varied and stressful surroundings.
CCS-Independent CSD Activities Are Physiologically Functional and Sufficient to Support Growth of Plant Cells
Upon investigating the physiological effects of the residual CCS-independent activities, we
obtained an unexpected result regarding the higher levels of superoxide anion, seed germination rate and root length of Atccs than Atcsd1. In trying to understand these results, it should be noted that in
Atcsd1, CSD1 activity is completely lost, whereas residual CSD1 activity is present in Atccs. Since
phospholipid membranes are impermeable to O2 (Takahashi and Asada, 1983), protection of different organelles can be achieved by variable distributions of SOD in different cellular compartments. In other words, the SOD activities in one compartment cannot complement functional deficiencies in these activities in another. Also, the lower germination rate and the shorter root length of Atcsd1 were very possibly due to the complete absence of CSD1 activity. In addition, current studies on SODs indicate that the level of SOD activity required for normal growth is much less than the actual activity level measured (Cohu et al., 2009). This can also be observed in yeast.
For instance, lysine biosynthesis is recovered by activity that was undetectable by the in-gel assay (Fig. 3), indicating that a small fraction of CuZnSOD activity is enough to protect the enzymes from damage by superoxide anions. In fact, Atccs showed no obvious phenotype under normal and stress conditions as compared with WT (Cohu et al., 2009). This demonstrates that loss of the bulk CSD activity did not seriously compromise plant viability. Our observations thus indicate that CCS-independent CSD activities are physiologically functional and sufficient to support the growth of plant cells. One possible reason for the high expression of CSD proteins in WT plants was suggested previously to be their role in copper buffering (Cohu et al., 2009).