行政院國家科學委員會補助專題研究計畫成果報告
※※※※※※※※※※※※※※※※※※※※※※※※※
※ ※
※ 利用 Cunninghamella blakesleeana 及 Actinoplanes sp. ※
※ 進行微生物轉換 ent-kauranes 及 ent-beyeranes ※
※ ※
※※※※※※※※※※※※※※※※※※※※※※※※※
計畫類別: 個別型計畫 □整合型計畫 計畫編號:NSC 89-2320-B-038-057 執行期間: 89 年 8 月 1 日至 91 年 7 月 31 日
計畫主持人:林淑娟 共同主持人:楊禮明
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
執行單位:台北醫學大學藥學系
中 華 民 國 91 年 10 月 15 日
行政院國家科學委員會專題研究計畫成果報告
國科會專題研究計畫成果報告 計畫編號:NSC 89-2320-B-038-057
執行期限:89 年 8 月 1 日至 91 年 7 月 31 日 主持人:林淑娟 台北醫學大學藥學系 共同主持人:楊禮明 國立中國醫藥研究所
一、中文摘要
Bacillus megaterium, Aspergillus niger 已被用於轉換具 ent-kaurene 結構的 steviol (ent-13-hydroxy-kaur-16-en-19-oic acid)(1) 及具 ent-beyerane 結構的 ent-16β-hydroxy- beyeran-19-oic acid (3). B. megaterium 轉換 此二種受質得到 19-O-β-D-glucopyranosyl (5,9)及 7β-hydroxy (4,8)衍生物,以及受 質 3 的 7β-hydroxy-19-O-β-D-glucopyrano- syl (10) 衍生物;此外受質 1 與 A. niger 進行培養可得到 7β-hydroxy (4),7-oxo (6),以及 7β,11α-dihydroxy (7)衍生物;受 質 3 與 A. niger 進行培養可得到 1α, 7β,16α-trihydroxy (11)及 1α,7β-dihydroxy- 16-keto (12)衍生物;其中代謝物 7, 9, 10 及 11 為首次發表的化合物,分離的代謝物經 由各種光譜分析以建立其結構。
關鍵詞:微生物轉換,四環二 類, steviol, ent-16β-hydroxybeyeran-19-oic acid
Abstr act
The microbial transformations of the tetracyclic diterpenes steviol (ent-13-hy- droxy-kaur-16-en-19-oic acid) (1) and ent-16β-hydroxybeyeran-19-oic acid (3) by Bacillus megaterium and Aspergillus niger have been studied. Incubation of 1 and 3 with B. megaterium afforded 19-O-β-D- glucopyranosyl (5 and 9) and 7β-hydroxy (4 and 8) derivatives, and 7β-hydroxy-19-O- β-D-glucopyranosyl derivative (10) for 3.
Incubation of 1 with A. niger afforded 7β- hydroxy (4), 7-oxo (6), and 7β,11α-dihy- droxy (7) derivatives. Incubation of 3 with A.
niger afforded 1α,7β,16α-trihydroxy (11) and 1α,7β-dihydroxy-16-keto (12) deriva- tives. Among them, the metabolites 7, 9, 10, and 11 are the first reported. Metabolite
structures were established on the basis of HRFABMS, 1D and 2D NMR spectral data, and enzymatic hydrolysis.
Keywor ds: microbial transformations, tetracyclic diterpenoids, steviol, ent-16β- hydroxybeyeran-19-oic acid
二、緣由與目的
The microbial transformation of multi- functional substrates can give rise to mixtures of products due to the presence of numerous enzymatic activities in the whole cells biocatalyst.1 Alternatively, microorgan- isms have already been successfully applied for the selective oxygenation of organic compounds, especially the unactivated sites in hydrocarbons.2 Thus, microorganisms have been used to transform a variety of organic compounds, such as steroids, alkaloids, terpenoids, and antibiotics.3,4 Stevioside is the major sweet component isolated from the leaves of Stevia rebaudiana.5,6 Steviol (ent-13-hydroxykaur- 16-en-19-oic acid) (1), the aglycone part of stevioside, is one of the major metabolite of stevioside during its enzymatic hydrolysis.7 It has been known that stevioside is converted to steviol by intestinal bacteria when stevioside is orally administered to rats.8,9 Steviol can also be obtained by oxidation of stevioside with NaIO4, and then hydrolysis with KOH.10 On the other hand, hydrolysis of stevioside with mineral acid affords isosteviol (2),11,12 which is then reduced with NaBH4 to yield ent-16β- hydroxybeyeran-19-oic acid (3).13 Steviol has been reported to be a toxic substance with mutagenic and bactericidal activities in Salmonella typhimurium TM 677.14 In contrast, steviol and stevioside have also been reported to have therapeutic value as
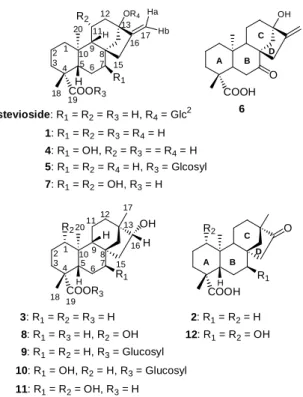
diuretic drugs15 and also as diabetic drugs by stimulating insulin secretion from pancreas.16 Ent-16β-hydroxybeyeran-19-oic acid (3) has the ability to lower blood pressure in spontaneously hypertensive rats.17 Although the A/B ring junction of the steviol (1) and ent-16β-hydroxybeyeran- 19-oic acid (3) is the same, the C- and D-rings are different (Figure 1). There is very little data available on the biotransformations of steviol and ent-16β-hydroxybeyeran- 19-oic acid, apart from that on Gibberella fujikuroi.13,18−22 As a part of an ongoing program to study the bioconversion of diterpenoids by microorganisms,12,23,24 and attempt to find out whether there exists a parallel between the action of the microorganisms on ent-kaurene and ent-beyerane of tetracyclic diterpenoids with similar chemical functions, the microbial metabolisms of 1 and 3 were investigated.
The isolation and structure elucidation of the metabolites are described herein.
三、結果與討論
A total of 27 microorganisms including fungi, yeast and bacteria were tested for their ability to metabolize steviol (1) and ent-16β-hydroxybeyeran-19-oic acid (3).
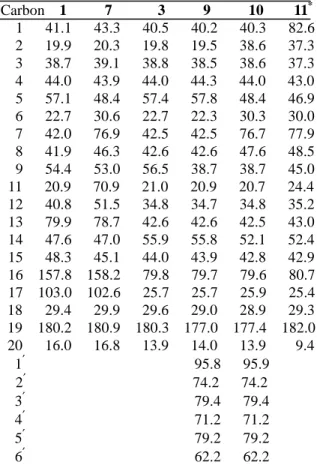
Bacillus megaterium and Aspergillus niger were the cultures capable of reproducibly bioconversion of 1 and 3 to different metabolites. The bioconversion of steviol (1) by B. megaterium and A. niger led to the isolation of the metabolites 4−7. Metabolites 4 and 6 have been prepared from the incubation of 1 with G. fujikuroi.19,20 Metabolite 5 has been isolated from the biotransformation of steviol by cultures cells of Eucalyptus perriniana.25 Metabolite 7 was obtained as white needles. Its HRFABMS (negative-ion mode) showed a [M – H]− at m/z 349.2021, indicating a molecular weight of 350, compatible with a molecular formula of C20H29O5 (calcd 349.2015). The 13C NMR spectrum displayed resonances for 20 carbons, while the DEPT spectrum showed the presence of the two methyl, eight methylene, four methine, and six quaternary carbons. The HMQC spectrum, compared to
that of 1, showed new resonances at δH 4.78 (δC 70.9) and δH 3.96 (δC 76.9). It indicated that metabolite 7 contains two more oxygen atoms than does steviol (1). In the DEPT spectrum, the resonance of C-6 has shifted downfield from δ 22.7 to 30.6, and the resonances of C-5 and C-9 have shifted upfield, from δ 57.1 to 48.4 and from δ 54.4 to 53.0, respectively. In the HMBC spectrum, δH 3.96 exhibits cross-peaks with δC 47.0 (C-14), 48.4 (C-5) and 53.0 (C-9). Thus, hydroxylation occurs at C-7, to which the resonance at δ 76.9 is attributed. The configuration at C-7 follows from the multiplicity of the H-7 signal in the 1H NMR spectrum, which is a broad singlet, indicating that the proton is in the equatorial (α) position.12,22,27 Furthermore, the NOESY spectrum also shows cross-peaks between δ 3.96 (H-7α) and H-6 (δ 2.47 and 2.69), H-14ax (δ 1.74) and H-15eq (δ 2.75).
Accordingly, the hydroxyl group is in the axial (β) position. The location of the second hydroxyl group at the C-11 position of 7 was deduced by HMBC correlations between δH
4.78 (H-11) and δC 41.4 (C-10) and 51.5 (C-12). The relative stereochemistry of the hydroxyl group at C-11, in the axial position (α), was suggested from the cross-peaks between H-11 (δ 4.78) and δ 2.52 (2H, H-9β and H-12) and 2.75 (H-12 and H-15) in the NOESY spectrum. Comparison of 1H NMR spectrum of 7 with methyl ent-7β,11α,13- trihydroxykaur-16-en-19-oate21 found that H-11 was located at δ 4.78 as a doublet triplet (J = 13.0, 6.5 Hz) in 7 and H-11α was located at δ 4.48 as a doublet (J = 6.0 Hz) in methyl ent-7β,11α,13-trihydroxykaur-16-en- 19-oate. Thus, the configuration of the hydroxyl group at C-11 was established to be α. On the basis of the above evidence, the structure of 7 is determined to be ent-7α, 11β,13-trihydroxykaur-16-en-19-oic acid.
Incubation of 3 with B. megaterium for 6 days led to the formation of metabolites 8-10. Metabolite 8 is ent-7α,16β-dihydroxy- beyeran-19-oic acid by comparison of NMR data with literature.13Metabolite 9 showed a quasi-molecular ion [M – H]− at m/z 481 and
COOR3 H 1 4
20 17
7
18 19 10
15 2
3 5
6 98 1112
16 13
H
OH H
A B
C D
R1 R2
COOHH R1 R2
3: R1= R2= R3= H 8: R1= R3= H, R2= OH 9: R1= R2= H, R3= Glucosyl 10: R1= OH, R2= H, R3= Glucosyl 11: R1= R2= OH, R3= H
O
2: R1= R2= H 12: R1= R2= OH OR4
2 1 20
34 5
18 11
12 13
6 7 15 98 10
1617 H
Ha Hb
COORH 3 19
R2
R1
stevioside: R1= R2= R3= H, R4= Glc2 1: R1= R2= R3= R4= H 4: R1= OH, R2= R3= = R4= H 5: R1= R2= R4= H, R3= Glcosyl 7: R1= R2= OH, R3= H
OH
COOH O
6
A B
C D
Figure 1. Structures of stevioside and compounds 1-12
the aglycone peak at m/z 319 [M – H]− due to loss of a hexosyl moiety in the negative-ion FABMS. It also gave a quasi-molecular ion peak at m/z 481.2809 corresponding to the molecular formula C26H41O8 (calcd 481.2801) in the negative HRFABMS. The 1H and 13C NMR spectra were similar to the substrate, except for the additional proton and carbon signals for the sugar moiety. The DEPT experiment of sugar moiety displayed one CH2 group at δ 62.2 and five CH groups at δ 95.8, 79.4, 79.2, 74.2, and 71.2 ppm. These signals were similar to those glucose with the anomeric proton resonating at δ 6.23 with J = 8.0 Hz.
The coupling constant agreed with a β-D configuration for the sugar moiety in metabolite.27 The HMQC and 1H−1H COSY spectra allowed for the assignment of all the protons of the sugar moiety of the metabolite, including the readily discernible cross-peak corresponding to the coupling of the anomeric proton H-1′ to H-2′. Comparison of the 13C NMR spectra of 3 and 9 indicated that 9 is a glucopyranosyl ester of 3 at C-19, based on the upfield shift of C-19 from δ 180.3 to 177.0.27 The sugar moiety was also confidently linked at C-19 of the aglycone basing on the HMBC correlation between the C-19 (δ 177.0) and H-1′ (δ 6.23). The β
configuration at C-1′ was further confirmed by enzymatic hydrolysis of the glucosidic linkage in 9 using β-D-glucosidase enzyme.
Based on the above evidence, metabolite 9 was determined as ent-16β-hydroxybeyeran- 19-β-D-glucopyranosyl ester. Metabolite 10 displayed a quasi-molecular ion peak at m/z 497.2755 corresponding to the molecular formula C26H41O9 (calcd 497.2751) in the negative HRFAMS. The 1H NMR spectrum shows close similarity to those of 9 and displays additional downfield signal for oxygen-bearing methine proton at δ 3.88 (br s). The characteristic anomeric proton of the sugar was seen as a doublet at δ 6.24 with the coupling constant, J = 8.0 Hz, indicating the glucosidic linkage to have β configura- tion.27 The DEPT spectrum shows the disappearance of one CH2 signal relative to 9 and the presence of one new CH signal at δ 76.7. By comparison of 13C NMR spectrum with 9, the resonance of C-6 has shifted downfield from δ 22.3 to 30.3, and the resonances of C-5 and C-9 have shifted upfield, from δ 57.8 to 48.3 and from δ 56.4 to 50.7, respectively. In the HMBC spectrum, δH 3.88 exhibits correlations with δ 48.3 (C-5) and 50.7 (C-9). Thus, hydroxylation occurs at C-7. The configuration of the hydroxyl group at C-7 was established to be β on the basis of NOESY data. The NOESY spectrum shows cross-peaks between δ 3.88 (H-7) and H-6 (δ 2.68 and 2.42), H-15 (δ 2.34), and H-14 (δ 1.59). The H-7 signal in the 1H NMR spectrum is a broad singlet, indicating that the proton is in the equatorial (α) position.12,26 Accordingly, the β-orienta- tion of 7-OH was established. On the basis of the 1H and 13C NMR spectra with the aid of HMQC, HMBC, NOESY, and 1H−1H COSY experiments and by comparison of NMR spectral data with 9, metabolite 10 is assigned the structure ent-7α,16β-hydroxy- beyeran-19-β-D-glucopyranosyl ester.
Metabolites 11 and 12 were obtained in 9.3 % and 11.5 % yields, respectively, by preparative-scale incubations of ent-16β- hydroxybeyeran-19-oic acid (3) with A. niger after 144 h incubation. Following solvent
Table 1. 13CNMR assignments for steviol (1) and 3, and metabolites 7, 9-11 (C5 D5 N, δ values)
Carbon 1 7 3 9 10 11* 1 41.1 43.3 40.5 40.2 40.3 82.6
2 19.9 20.3 19.8 19.5 38.6 37.3 3 38.7 39.1 38.8 38.5 38.6 37.3 4 44.0 43.9 44.0 44.3 44.0 43.0
5 57.1 48.4 57.4 57.8 48.4 46.9 6 22.7 30.6 22.7 22.3 30.3 30.0 7 42.0 76.9 42.5 42.5 76.7 77.9 8 41.9 46.3 42.6 42.6 47.6 48.5
9 54.4 53.0 56.5 38.7 38.7 45.0
11 20.9 70.9 21.0 20.9 20.7 24.4 12 40.8 51.5 34.8 34.7 34.8 35.2
13 79.9 78.7 42.6 42.6 42.5 43.0 14 47.6 47.0 55.9 55.8 52.1 52.4 15 48.3 45.1 44.0 43.9 42.8 42.9 16 157.8 158.2 79.8 79.7 79.6 80.7 17 103.0 102.6 25.7 25.7 25.9 25.4 18 29.4 29.9 29.6 29.0 28.9 29.3 19 180.2 180.9 180.3 177.0 177.4 182.0 20 16.0 16.8 13.9 14.0 13.9 9.4 1′ 95.8 95.9
2′ 74.2 74.2 3′ 79.4 79.4 4′ 71.2 71.2 5′ 79.2 79.2 6′ 62.2 62.2
* in CD3OD
extraction and column chromatographic purification, samples of metabolites were subjected to spectral analyses. The HRFABMS of metabolite 11 exhibited a quasi-molecular ion peak at m/z 351.2168 corresponding to C20H31O5 (calcd 351.2172), indicating a metabolite structure containing two more oxygen atoms than 3. In C5D5N as the solvent, the DEPT spectrum shows the disappearance of two CH2 signals and the presence of two new CH signals at δ 81.8 and 76.8, confirming that 11 is a dihydro- xylated metabolite of 3. However, δC 81.8 and 76.8 are correlated to δH 3.94 (2H, singlet and shoulder like) in the HMQC spectrum. By changing the solvent to CD3OD, these two signals are cleanly separated and showed at δC 82.6 and 77.9, and δH 3.33 and 3.42, respectively, in the HMQC spectrum. In the HMBC spectrum, the chemical shift of δH 3.33 showed connectivities with CH3-20 (δ 9.4), C-2 (δ 30.6), C-10 (δ 45.0), and C-9 (δ 51.7). In
1H−1H COSY spectrum, δH 3.33 showed cross peak with H-2 (δ 1.48). Thus, it
suggested that one of hydroxyl group was at C-1. The relative stereochemistry of 1-OH was deduced from the NOESY experiment as well as the coupling constant in the 1H NMR spectrum. The coupling constant of the methine proton at C-1 exhibited double- doublet (J = 11.5, 4.5 Hz) due to coupling with the protons of the neighboring C-2. The NOESY spectrum also showed NOE effects between H-1 (δ 3.33) and H-2 (δ 1.48), H-5β (δ 1.63), H-9β (δ 1.57), and CH3-18 (δ1.13).
Thus, the hydroxyl group at C-1 was in an equatorial (α) configura- tion.26 The location of the second hydroxyl group at C-7 position was deduced by 13C NMR signals. In spite of in C5D5N or CD3OD, the resonance of C-6 has shifted downfield from δ 22.7 to 30.0, and the resonances of C-5 and C-9 have shifted upfield, from δ 57.4 to 46.9 and from δ 56.5 to 51.7, respectively. In the HMBC spectrum, δH 3.42 exhibits connectivities with δ 46.9 (C-5) and 51.7 (C-9). Therefore, hydroxylation occurs at C-7. The configura- tion of hydroxyl group at C-7 was assigned to be β due to the presence of a broad singlet proton signal at δH 3.42.12,26 On the basis of the above evidence, the structure of 11 is determined to be ent-1β,7α,16β-trihydroxy- beyeran-19-oic acid. The difference between 11 and 12 is the lack of the H-16β and C-16 signals in the 1H and 13C NMR spectra of 12, which is replaced by a carbonyl group at δ 221.0. Thus, metabolite 12 is ent-1β,7α- dihydroxy-16-ketobeyeran-19-oic acid. This metabolite has been isolated as methyl derivative in biotransformation of isosteviol by Aspergillus niger CMI 17454.26
四、計畫成果自評
Two selected substrates, steviol and ent-16β-hydroxybeyeran-19-oic acid, are with different carbon skeletons of tetracyclic diterpenoids. In this investigation, the selected microorganisms have the ability to introduce hydroxyl group into 7β position for both substrates. This is the most common reaction for introduction of oxygen function at C-7 in the microbial hydroxylation of tetracyclic diterpenoids and steroids. In addition, this study revealed that A. niger has
the ability to hydroxylate at 1α,7β- positions of ent-16β-hydroxybeyeran-19-oic acid (3) followed by oxidation at 16α-hydroxy group to yield 12. However, the dihydroxylation was occurred at 7β,11α-positions of steviol (1) instead of 1α,7β-positions. The configu- rations of 7 were opposite to the metabolite obtained from G. fujikuroi. Thus, the results obtained in this study indicated that the hydroxylation of both A- and B-rings’ of ent-beyerane could be accompli- shed by A.
niger. On the other hand, conjugation of metabolized xenobitics is generally considered to be a detoxication mechanism.
Previous studies demonstrated that B.
megaterium could serve as a prokaryotic model for mammalian drug metabolism. Our study demonstrated that B. megaterium could catalyze not only in phase I hydroxylation but also in phase II conjugation for both substrates. Thus, this study provides us to understand the relationship between the ent-kaurene and ent-beyerane and the microbial enzymes responsible for their hydroxylation and glucosidation. Also, elucidation of the metabolic mechanism of the glucosidation in tetracyclic diterpenoids by B. megaterium may provide new insights in the field of enzymology and glucosidic chemistry. The antidiabetic and antihyper- tension activity tests will be evaluated later.
五、參考文獻
1. Fuganti, C.; Minut, J.; Fantoni, P.; Servi, S. J.
Mol. Catal. B: Enzym. 1998, 4, 47-52.
2. Adam, W.; Luckacs, Z.; Kahle, C.;
Saha-Möller, C. R.; Schreier, P. J. Mol. Catal. B: Enzym.2001, 11, 377-385.
3. Kieslich, K.; 1976. Microbial Transformation of Non-Steroid Cyclic Compounds. Georg Thieme Verlag, Stuttgart, Germany.
4. Hanson, J. R. Nat. Prod. Rep. 1992, 10, 139-151.
5. Hanson, J. R.; de Oliveira, B. H. Nat. Prod. Rep. 1993, 10, 301-309.
6. Wood, H. B.; Allerton, R.; Diehl, H. W.;
Fletcher, H. G. J. Org. Chem.1995, 20, 875-883.
7. Hupatae A. M.; Toskulakao, C.; Buddhaasukh, D.; Wilairat, P.; Glinsukon, T. J. Clin.
Biochem. Nutr. 1997, 23, 177-186.
8. Nakayama, K.; Kasahara, D.; Yamamoto, F. J.
Food Soc. Japan 1986, 27, 1-8.
9. Wingard, R. E. Jr.; Brown, J. P.; Enderlin, F.
E.; Dale, J. A.; Seiz, C. T.
Experientia1980, 36, 519-520.
10. Ogawa, T.; Nozaki, M.; Matsui, M.
Tetrahedron1980, 36, 2641-2648.
11. Avent, A. G.; Hanson, J. R.; de Oliveira, B. H. Phytochemistry1990, 29, 2712-
2713.
12. Hsu, F. L.; Hou, C. J.;Yang, L. M.; Liu, B. C.; Lin, S. J. J. Nat. Prods.2002, 65,
273-277.
13. Ali, H. S.; Hanson, J. R.; de Oliveira, B.
H. Phytochemistry, 1992, 31, 507-510.
14. Pezzuto, J. M.; Compaadre, C. M.;
Swansion, S. M.; Nanayakkara, P. D.;
Kinghorn, A. D. Proc. Natl. Acad. Sci.
USA 1985, 82, 2478-2482.
15. Melis, M. S. Phytomedicine1997, 3, 349-352.
16. Jeppesen, P. B.; Gregersen, S.; Poulsen, C. R.; Hermansen, K. Metabolism 2000, 49, 208-214.
17. Cheng, J. T., personal communication, Department of Pharmacology, College of Medicine, National Cheng-Kung University, Tainan, Taiwan.
18. Hanson, J. R.; White, A. F. Tetrahedron, 1968, 24, 6291-6293.
19. Bearder, J. R.; Frydman, V. M.; Gaskin, P.;
MacMillan, J.; Wels, C. M. J. Chem. Soc.
Perkin I, 1976, 173-183.
20. Murofushi, N.; Shigematsu, Y.; Nagura, S.;
Takahashi, N. Agric. Biol. Chem.1982, 46, 2305-2311.
21. Shigematsu, Y.; Murofushi, N.; Takahashi, N. Agric. Biol. Chem.1982, 46, 2313-2318.
22. Hanson, J. R.; de Oliverira, B. H.
Phytochemistry1990, 31, 3805-3809.
23. Lin, S. J.; Short, R. E.; Ford, S. P.; Grings, E.
E.; Rosazza, J. P. N. J. Nat. Prods.1998, 61, 51-56.
24. Lin, S. J.; Rosazza, J. P. N. J. Nat. Prods.
1998, 61, 922-928.
25. Orihara, Y.; Saiki, K.; Furuya, T.
Phytochemistry, 1991, 30, 3989-3992.
26. de Oliveira, B. H.; dos Santos, M. C.; Leal, P. C. Phytochemistry1999, 51,
737−741.
27. Chatterjee, P.; Pezzuto, J. M.; Kouzi, S. A. J.
Nat. Prod, 1999, 62, 761-763.