行政院國家科學委員會專題研究計畫 成果報告
Sildenafil 和葡萄柚汁在 Cytochrome P450 的體外代謝交互 作用之動力學模式探討(2/2)
計畫類別: 個別型計畫
計畫編號: NSC94-2320-B-038-012-
執行期間: 94 年 08 月 01 日至 95 年 07 月 31 日 執行單位: 臺北醫學大學藥學系(所)
計畫主持人: 何秀娥
計畫參與人員: 林盈谷,黃嘉惠
報告類型: 完整報告
處理方式: 本計畫可公開查詢
中 華 民 國 95 年 6 月 28 日
目錄
中文摘要 ... II
Abstract ...III
Introduction ...1
Materials and Methods ...3
Results ...6
Discussion...7
References ...10
Figures ...13
中文摘要
Sildenafil 經過人體吸收後最主要是經過肝臟酵素 CYP450 3A4 代謝成 同樣具有藥理活性的代謝物 UK-103,320。實驗結果顯示,Sildenafil 在 microsome 的酵素動力學是遵循 Michaelis - Menten kinetic model,其 Vmax= 1.96 μMmin-1, Km= 27.31 μM.。利用 Testosterone 和 Carbamazepine 評估 Sildenafil 在 CYP3A4 的活性結合區的實驗,結果顯示 Sildenafi 與 Testosterone 和 Carbamazepine 進行 代謝作用是屬於互相抑制的現象,彼此之間會相互影響,結果也顯示,Sildenafil 與 Testosterone 和 Carbamazepine,在 CYP3A4 的鍵結牽涉到一個以上活性位置 的動力學模式。
關鍵詞: sildenafil, testosterone, carbamazepine, drug interaction, CYP3A4
Abstract
The biotransformation of sildenafil to its major circulating metabolite, UK-103,320, was studied in male rat liver microsomes. The conversion of sildenafil to UK-103,320 by rat microsomes followed Michaelis-Menten kinetics, for which the parameters were Vmax = 1.96 μM min-1and Km = 27.31 μM. Using substrates of CYP3A4 of testosterone and carbamazepine, the active sites on CYP3A4 responsible for metabolizing sildenafil were also evaluated. Sildenafil biotransformation was inhibited in the individual presence of testosterone and carbamazepine. The most unusual results were the individual interactions of sildenafil with testosterone and carbamazepine. Although testosterone and carbamazepine can inhibit sildenafil demethylation in concentration- and incubation time-dependent manners, sildenafil did not inhibit testosterone hydroxylation or carbamazepine epoxidation. These results may be explained by a model in which multiple substrates or ligands can concurrently bind to the active site of a single CYP3A4 molecule. However, the contribution of separate allosteric sites and conformational heterogeneity to the atypical kinetics of CYP3A4 cannot be ruled out in this study.
Key words: sildenafil, testosterone, carbamazepine, drug interaction, CYP3A4
Introduction
Sildenafil (Viagra), a selective inhibitor of cGMP phosphodiesterase type 5, is an orally active treatment for erectile dysfunction (ED) (Langtry et al., 1999). A study using human liver microsomes demonstrated that 79% of sildenafil's biotransformation to UK-103,320 (N-demethylsildenafil) is attributable to CYP3A.
UK-103,320 has been identified as a metabolite of sildenafil in the mouse, rat, dog, and human (Walker et al., 1999). A small percentage of metabolite formation is due to CYP2C9 (20%), CYP2D6, and CYP2C19 (collectively < 2%) activity (Hyland et al., 2001). Current numbers indicate that approximately 30 million men in the US are affected by ED, with half of all men aged > 40 years experiencing some degree of ED.
Due to the high frequency of ED in the US as well as to the possible role of a variety of diseases or pharmacological agents in the pathogenesis of sexual dysfunction, sildenafil is likely to be co-administered with other drugs. In general, ED and depression frequently co-occur with other conditions, including diabetes, hypertension, cardiovascular disease, neurologic disorders (e.g., parkinsonism and multiple sclerosis), and endocrine disorders (e.g., adrenal thyroid and gonadal disorders) (Ashton and Bennett, 1999; Nurnberg et al., 1999). It can be expected that sildenafil will be co-administered with agents that either directly cause sexual dysfunction or are used to treat disease associated with sexual dysfunction. Thus drug-drug interactions have become an important clinical issue due to the effects of one drug on the efficacy, toxicity, or disposition of another drug. The estimated incidence of clinically significant drug-drug interactions is as high as 20% in patients receiving multiple drugs. The majority of cases of drug-drug interactions are a result of pharmacokinetic or pharmacodynamic alterations. Although interacting agents can affect all aspects of drug disposition, including absorption, distribution, metabolism, and excretion through a variety of mechanisms, the most common drug interactions can be understood in terms of alterations in metabolism, which are associated primarily with changes in the activities of cytochrome P450. Co-administration of sildenafil and CYP3A4 inhibitors (ketoconazole and ritonavir) may lead to increased plasma concentrations of sildenafil (Warrington et al., 2000). This may, in turn, lead to an increase in adverse effects commonly associated with sildenafil such as headaches, flushing, dyspepsia, and visual changes. Therefore, it is important to understand the potential for drug interactions with sildenafil.
To better understand drug interactions with sildenafil, the present study was conducted to investigate the effects of carbamazepine and testosterone on sildenafil metabolism and evaluate the possibility of drug-drug interactions with sildenafil.
Carbamazepine is widely used for the treatment of epileptic seizures, trigeminal neuralgia, and psychiatric disorders. Carbamazepine is usually administered at daily oral doses ranging from 200 to 1200 mg or higher, which give rise to drug plasma levels of 4~12 μg ml-1. The relationship between the carbamazepine dose and its plasma concentration is often unpredictable and depends on the patient’s metabolic state and age and might be affected by cotherapy. The drug is principally metabolized by CYPs to 10,11-epoxy carbamazepine (Segelman et al., 1985). At the same time, carbamazepine metabolism can be induced or inhibited by other drugs. The addition of felbamate to carbamazepine monotherapy results in a decrease in carbamazepine plasma concentrations. It is suggested that in vivo heteroactivation of CYP3A4 is a possible mechanism of clinically observed drug interactions between felbamate and carbamazepine (Egnell et al., 2003). In the human male, testosterone is the major
circulating androgen. Testosterone is essential for the development and maintenance of specific reproductive tissues as well as for other characteristic male properties such as control of spermatogenesis, and many aspects of sexually dimorphic behavior.
Indeed, in subsequent studies, Webb et al. demonstrated that an intracoronary infusion of physiological concentrations of testosterone increased coronary artery diameter and coronary blood flow in male patients with cardiovascular disease. Testosterone hydroxylation, yielding the 6ß-hydroxy metabolite, is considered to be a relatively specific index reaction for human CYP3A-mediated metabolism. In testosterone interaction studies, testosterone inhibited 1-OH-triazolam formation, but significantly activated 4-OH-triazolam formation in human liver microsomes. The interaction patterns between compounds with CYP3A4 were found to be substrate-dependent.
Mutual inhibition, partial inhibition, and activation were observed in testosterone-terfenadine, testosterone-midazolam, and terfenadine-midazolam interactions. As a result, drug-drug interactions associated with modulation of CYP3A-mediated metabolism can be of substantial clinical importance (Wang et al., 2000).
Kinetic techniques that adequately describe the interaction between multiple drugs that bind simultaneously to a single enzyme active site are needed to aid in the understanding of clinically observed drug-drug interactions and in assessing their clinical significance. In many cases, CYP3A exhibits unusual kinetic characteristics that result from the metabolism of multiple substrates which coexist at the active site (Shou et al., 1994; Ueng et al., 1997; Korzekwa et al., 1998; Hosea et al., 2000;
Kenworthy et al., 2001). It is hoped that application of kinetic characteristics can aid in our understanding of drug-drug interactions of sildenafil with carbamazepine and testosterone.
Materials and Methods Drugs and chemicals
Standard sildenafil citrate was obtained from Trans American Chemical Co. (Upland, CA, USA). Nicotinamide adenine dinucleotide phosphate (NADPH)-CYP reductase, potassium dihydrogen phosphate, di-potassium hydrogen phosphate, trifluoroacetic acid, dimethyl sulfoxide, carbamazepine, 10,11-epoxide carbamazepine, and hydroxyl testosterone were purchased from Sigma Aldrich Chemical Co. (St. Louis, MO, USA).
Testosterone was obtained from Fluka Chemical Co. (Buchs, Steinheim, Switzerland, USA). Methanol and acetonitrile for liquid chromatography were HPLC grade and were obtained from Merck (Darmstadt, Germany). All other reagents used were of reagent grade or better.
Animals and preparation of liver microsomes
Male adult Wistar rats (300±50 g) were obtained from the Laboratory Animal Center of National Taiwan University (Taipei, Taiwan). All animal use protocols were approved by the Laboratory Animal Research Committee of Taipei Medical University (Taipei, Taiwan). Rats were housed in stainless steel cages with five animals per cage in a temperature-controlled (24~26 °C) room with a 12-h light/dark cycle. Rats were allowed free access to water and food for 1 week before the experiments. They were starved overnight before the experiments. Each animal was anesthetized with ether. Microsomes were prepared from pooled (six to eight) fresh livers by homogenization and differential centrifugations, as previously described (Guengerich et al., 1989). All procedures were carried out at 4 °C. The microsomes were stored at –80 °C until use in 0.1 M phosphate buffer (pH 7.4) containing 20%
(w/v) glycerol. Protein concentrations were determined using the Bio-Rad protein assay kit (Bio-Rad, Hercules, CA, USA) with BSA as the standard.
Sildenafil kinetics in rat liver microsomes
Varying quantities of sildenafil ranging from 2.5 to 103 μM were added to hepatic microsomes from rat preparations in the presence of NDPH and 0.1 M potassium phosphate buffer (pH 7.4). The mixtures were incubated for different times (5, 10, 20 and 40 min) at 37 °C and terminated with the addition of methanol and exposure to ice. Furthermore incubation mixtures were centrifuged at 14,000 rpm for 10 min to remove the protein. The supernatants were transferred to autosampling vials for high-performance liquid chromatography (HPLC) analysis. All samples were incubated in triplicate. Samples were chromatographed on a 25-cm Biosil Aqu-ODS-W column, with a mobile phase consisting of acetonitrile and water solution (containing 0.1% trifluoroacetic acid) at a 40:60 (v/v) ratio. The eluent was detected with a Jasco (Tokyo, Japan) UV-975 UV-VIS detector at 230 nm. The flow rate was set at 1.0 ml min-1. This method was found to be selective, precise, and linear over a concentration range of 0.5 to 103 μM sildenafil (Sheu et al., 2003). All mobile-phase solutions were filtered and degassed ultrasonically before use. The HPLC system was controlled by a PC workstation using Chromatography Data Station software (SISC, Taipei, Taiwan) installed on it.
Liquid chromatography-tandem mass spectrometry (LC-MS)
Identification of UK-103,320 as the principal metabolite of sildenafil in incubation mixtures was performed by liquid chromatography-tandem mass spectrometry (LC-MS) (Hyland et al., 2000). For liquid chromatography-mass spectrometry (LC-MS) studies, chromatography was conducted on a Hewlett-Packard HP 1100 gradient system. The system was controlled and data were processed using ChemStation Software revision A.06.01 (Hewlett Packard, Palo Alto, CA, USA). The MS system was operated using the API-ES mode in the positive ion mode. The gas used was nitrogen, the temperature was 325 °C, and the fragment voltage was optimized. Separation was carried out on a 50 × 4.6-mm C8, 3.5-μm analytical column (Zorbax XDB, C8; Hewlett Packard). The mobile phase consisted of 0.1%
trifluoroacetic acid in water (solvent A) and acetonitrile (solvent B). The following gradient was run: at time 0 min, 100% solvent A and at 8 min, 80% solvent B. The flow rate was 1.0 ml min-1. The analytical column was also maintained at 40 °C. One minute after sample injection, the mass-selective detector was activated.
Metabolic interactions of sildenafil with carbamazepine and testosterone in rat liver microsomes
The effects of carbamazepine and testosterone on the metabolism of sildenafil (0.5 to 103 uM) by CYP enzymes were investigated in a pool of rat liver microsomes. The incubation mixture consisted of hepatic microsomes from rat preparations in the presence of NADPH, 0.1 M potassium phosphate buffer (pH 7.4), and carbamazepine or testosterone. The concentrations of carbamazepine and testosterone were varied from 20 to 600 and 10 to 200 μM, respectively. The incubation mixture was preincubated for 5 min at 37 °C in a shaking water bath, and sildenafil (0.5 to 103μM) was added to further incubation for different times (5, 10, 20, and 40 min) at 37 °C.
The reactions were terminated with the addition of methanol and exposure to ice.
Furthermore incubation mixtures were centrifuged at 14,000 rpm for 10 min to remove the protein. The supernatants were analyzed immediately by HPLC. An HPLC system equipped with a pump (Jasco PU-980 Intelligent HPLC pump) and an autosampler (Jasco AS-950-10 Intelligent Sampler) were used. The supernatants of carbamazepine and testosterone were detected with a Jasco UV-975 UV-VIS detector at 236 and 254 nm, respectively. A 4.6 x 250-mm (I.D.) Biosil Aqu-ODS-W column (Biotic Chemical Co., Taipei, Taiwan) with a particle size of 5 μm and a mobile phase consisting of water, acetonitrile, and methanol (containing 0.1% trifluoroacetic acid) at 60: 20: 20 and 50: 20: 30 (v/v) ratios were used in separating the metabolites of carbamazepine and testosterone, respectively. The flow rate was set at 1.0 ml min-1. These methods for analyzing testosterone and carbamazepine were found to be selective, precise, and linear over concentration ranges of 0.1~10 and 0.25~10 μM, respectively. All mobile-phase solutions were filtered and degassed ultrasonically before use. The HPLC system was controlled by a PC workstation using Chromatography Data Station software (SISC) installed on it.
Data analysis
The formation of UK-102,320 by liver microsomes was consistent with Michaelis–Menten kinetics with uncompetitive substrate inhibition. The following equation was fitted to UK-103,320 data points:
S K
S V V
m
max
0 ;
in which V represents the velocity of UK-103,320 formation and S is the concentration of the substrate, sildenafil. Vmax (representing the maximum reaction velocity) and Km(indicating the substrate concentration corresponding to 50% Vmax) were calculated by nonlinear regression.
Results
Identification of UK-103,320 (N-demethylsildenafil) and metabolism of sildenafil Upon LC-MS analysis, a major metabolite (with a retention time of 5.7 min) of the CYP-catalyzed metabolism of sildenafil exhibited an MH+ion at m/z 461, suggestive of the demethylation of sildenafil. By comparing the HPLC profile and mass spectral information, the metabolite was identified as UK-103,320. Incubation of sildenafil with microsomes in the presence of NADPH led to the formation of the major metabolite, UK-103,320, which was identified by comparing the HPLC retentions and LC-MS spectra. Formation of UK-103,320 (μg ml-1) in the metabolic reaction of various concentration of sildenafil incubated with rat microsomes for different incubation times is shown in Fig. 1. Results indicate that the formation of UK-103,320 was proportionally enhanced by an increasing incubation time and concentration of sildenafil to 200 μM. Figure 2 shows UK-103,320 formation in rat liver microsomes was consistent with single-enzyme Michaelis-Menten kinetics with uncompetitive substrate inhibition. The estimated Km and Vmax were 27.31 μM and 1.96 μM min-1.
Metabolic interaction of sildenafil with carbamazepine and testosterone in rat liver microsomes
Sildenafil was metabolized by CYP3A4 to UK-103,320, and we investigated the effects of testosterone and carbamazepine on the metabolic pathways of sildenafil. In the metabolism of sildenafil by microsomes, the presence of testosterone (10~200 μM) or carbamazepine (20~600 μM) was found to inhibit the formation of UK-103,320. As shown in Figs. 3 and 4, testosterone and carbamazepine inhibited the major sildenafil metabolic pathway. When the concentration of testosterone was increased from 10 to 200 μM, a similar extent of inhibition (of about 70%~85%) of the metabolism of sildenafil for different incubation times was noted. However, inhibition of sildenafil metabolism increased in parallel with an increase in the carbamazepine concentration and incubation time. At higher concentrations (150 and 600 μM), carbamazepine showed greater inhibition. Maximum inhibition was observed at approximately 600 μM carbamazepine with 40 min of incubation time. Both testosterone and carbamazepine produced almost complete inhibition of UK-103,320 formation at 200 μM sildenafil used. Effects of sildenafil on the formation of hydroxyl testosterone and 10,11-epoxide carbamazepine from biotransformation of testosterone and carbamazepine incubated with rat liver microsomes for different incubation times are listed in Tables 1 and 2, respectively. At fixed concentration of testosterone or carbamazepine, increasing concentrations of sildenafil produced insignificant differences in the formation of hydroxyl testosterone and 10,11-epoxide carbamazepine from the biotransformation of testosterone and carbamazepine incubated with rat liver microsomes for different incubation times. Thus, higher concentrations of sildenafil did not necessarily result in greater inhibition of testosterone 6-hydroxylation and carbamazepine epoxidation. Instead, only partial inhibition was observed. These results showed that testosterone and carbamazepine can inhibit sildenafil demethylation, and that carbamazepine has an inhibitory effect in a concentration-dependant manner, but sildenafil did not inhibit testosterone hydroxylation or carbamazepine epoxidation.
Discussion
Using rat liver microsomes, we demonstrated that sildenafil is N-demethylated to produce UK-102,320. The Km for UK-103,320 formation in rat liver microsomes (27.31 μM) was similar to the Km values for UK-103,320 formation by microsomes expressing CYP3A4 (23.1 μM) (Warrington et al., 2000). This further supports the role for CYP3A4 in UK-103,320 formation. The biotransformation of sildenafil in male rat liver microsomes is suitable as a model for sildenafil metabolism and evaluation of drug-drug interactions.
In general, substrates for CYP3A4 vary greatly in their physicochemical properties such as structure, molecular size and shape, lipophilicity, electronic characteristics, and kinetic interactions with enzyme proteins. The fact that CYP3A4 can accommodate substrates of a relatively large size, e.g., cyclosporine (Mr = 1201), suggests that multiple small or intermediate-sized molecules might be able to coexist at the active site of this enzyme. Indeed, some evidence has been obtained to support this hypothesis based on kinetic studies and NMR data (Shou et al., 1999, 2001). If an active site is capable of accommodating two substrates simultaneously, the resulting kinetic properties, e.g., binding affinity and catalytic ability, are likely to be affected differently from those observed with simple Michaelis-Menten inhibition and activation, both of which are derived from the one binding region at the active site (Domanski et al., 2000; Schrag et al., 2001). Using substrates of CYP3A4 of testosterone and carbamazepine, active sites on CYP3A4 responsible for metabolizing sildenafil were evaluated in this study. Results demonstrated that the metabolism of sildenafil via N-demethylation by CYP3A4 in the presence of its substrate of either testosterone or carbamazepine was inhibited. These results suggest that the testosterone-CYP3A4-sildenafil and cabamazepine-CYP3A4-sildenafil complexes favor testosterone 6-hydroxylation and carbamazepine epoxidation pathways rather than the sildenafil demethylation pathway. These observed kinetic changes with the interaction of two substrates with the enzyme cannot be explained by simple Michaelis-Menten kinetics. The interaction patterns between compounds with CYP were found to be substrate-dependent. A kinetic model for two substrates binding to two separate active sites was found to be optimal for describing the metabolism of sildenafil by CYP3A4 in the presence of either testosterone or carbamazepine. It was concluded that more than one site present on CYP3A4 is responsible for its metabolism.
The unusual kinetics associated with two or more substrate interactions have been documented in a number of reports. Several models have been proposed to explain the unusual kinetic characteristics with CYP3A4 involving two substrates. The atypical CYP3A4 kinetics, including activation, mutual inhibition, partial inhibition, and alteration of regiospecificity, observed in substrate oxidation and substrate-substrate interaction studies was explained by the two-substrate model (Korzekwa et al., 1998), the cooperative model (Ueng et al., 1997), and the multiple conformer model (Koley et al., 1995). The two-site model proposes that two substrates can simultaneously bind to the CYP3A4 active site (Shou et al., 1994, Wang et al., 1997). Korzekwa et al.
(1998) proposed a two-site (or multiple-site) model in which the enzyme can bind two molecules of one substrate or one molecule each of the two substrates, or one molecule each of the substrate and effector. The two-site model can describe many of the atypical CYP3A4 kinetics. The potential for binding to more than two sites on
CYP3A4 was also alluded to by Shou et al. (1994) and more recently by Domanski et al. (2000) and Hosea et al. (2000). It has been proposed that the site for metabolism and activation by an effector may be distinct since two cooperative substrates have not been shown to cause mutual inhibition. Studies with site-directed mutants of CYP3A4 also support the hypothesis that both substrate and effector sites are closely linked and may be involved in substrate and/or effector binding, depending on the molecule of study (Domanski et al., 2000). In this study, we showed that testosterone and carbamazepine inhibited sildenafil demethylation, but less so for sildenafil inhibition of testosterone 6-hydroxylation and carbamazepine epoxidation. As for the unusual sildenafil-testosterone and sildenafil-carbamazepine interactions, one can explain the results with a two-substrate model by postulating that testosterone and carbamazepine have freedom of movement and can bind to multiple sites, including the sildenafil-binding site of the CYP3A4 active site. Consequently, testosterone and carbamazepine can inhibit sildenafil demethylation, but inhibition of testosterone 6-hydroxylation and carbamazepine epoxidation cannot be demonstrated kinetically.
With the multiple conformer model, sildenafil-testosterone and sildenafil-carbamazepine interactions can be explained by assuming that testosterone and carbamazepine have high affinities for multiple CYP3A4 conformers (including the ones interacting with sildenafil), but sildenafil has only limited affinity for certain conformers. Perhaps one should consider a modified model which takes multiple binding sites, allosteric sites, and multiple conformations into account.
An unusual result was also found in the interaction between testosterone and nifedipine. Although nifedipine inhibited testosterone 6-hydroxylation, testosterone did not inhibit nifedipine oxidation (Wang, 2000). One similar, but not identical, example was the report by Ueng et al. (1997), which stated the 7,8-benzoflavone activates the metabolism of aflatoxin B1, but aflatoxin B1 does not inhibit 7,8-benzoflavone metabolism. However, it is rare to find two substrates of the same enzyme which do not show mutual inhibition kinetics in the literature. These results may be explained by a model in which multiple substrates or ligands can bind concurrently to the active site of CYP molecules. However, the contribution of separate allosteric sites and conformational heterogeneity to the atypical kinetics of CYP cannot be ruled out in this model. Because of the unique properties of CYP3A4, substrate interactions involving this enzyme do not always follow typical competitive inhibition kinetics. Regardless of the mechanisms by which CYP3A4-dependent drug interactions occur, this study provides information that the co-administration of sildenafil and testosterone or carbamazepine may lead to increased plasma concentrations of sildenafil. This may, in turn, lead to an increase in the adverse effects commonly associated with sildenafil such as headaches, flushing, dyspepsia, and visual changes. Information on in vitro drug-drug interactions at the level of cytochrome P450 enzymes can be extremely useful in the evaluation of the potential of an agent to cause drug interactions in clinic use.
In conclusion, the effects of one substrate on the metabolism of another appeared to be dependent on the substrate of use. Clinicians should therefore be aware of the possibility of drug-drug interactions when prescribing sildenafil concomitantly with known substrates of CYP3A4.
Acknowledgements
Financial support by the National Sciences Council of the ROC (NSC93-2320-B-038-049) is highly appreciated.
References
Ashton, A.K., Bennett, R.G., 1999, Sildenafil treatment of serotonin reuptake inhibitor-induced sexual dysfunction. J. Clin. Psychiatry 60, 194-195.
Domanski, T.L., He, Y.A., Harlow, G.R., Halpert, J.R., 2000, Dual role of human cytochrome P450 3A4 residue Phe-304 in substrate specificity and cooperativity. J.
Pharmacol. Exp. Ther. 293, 585-591.
Egnell, A.C., Houston, B., Boyer, S., 2003, In vivo CYP3A4 Heteroactivation is a possible mechanism for the drug interaction between felbamate and carbamazepine. J. Pharmacol. Exp. Ther. 305, 1251-1262.
Guengerich, F.P., 2001, Analysis and characterization of enzymes and nucleic acids.
In A. W. Hayes (ed.), Principles and Methods of Toxicology, 4th Edition (Philadelphia: Taylor & Francis) pp. 1625-1687.
Hosea, N.A., Miller, G.P., Guengerich, F.P., 2000 Elucidation of distinct ligand binding sites for cytochrome P450 3A. Biochemistry 39, 5929-5939.
Hyland, R., Roe, E.G., Jones, B.C., Smith, D.A., 2001, Identification of the cytochrome P450 enzymes involved in the N-demethylation of sildenafil. Br. J.
Clin. Pharmacol. 51, 239-248.
Kenworthy, K., Clarke, S., Andrews, J., Houston, J.B., 2001, Multisite kinetic models for CYP3A4: simultaneous activation and inhibition of diazepam and testosterone metabolism. Drug Metab. Dispos. 29, 1644-1651.
Koley, A.P., Buters, J.T.M., Robinson, R.C., Markowitz, A., Friedman, F.K. 1995, CO binding kinetics of human cytochrome P450 3A4: specific interaction of substrates with kinetically distinguishable conformers. J. Biol. Chem. 270, 5014-5018.
Korzekwa, K.R., Krishnamachary, N., Shou, M., Parise, R.A., Rettie, A.E., Gonzalez, F.J., Tracy, T.S., 1998, Evaluation of atypical cytochrome P450 kinetics with two-substrate models: evidence that multiple substrates can simultaneously bind to cytochrome P450 active sites. Biochemistry 37, 4137-4147.
Langtry, H.D., Markham, A., 1999, Sildenafil. Drugs 57, 967-989.
Nurnberg, H.G., Lauriello, J., Hensley, P.L., Parker, L.M., Keith, S.J., 1999, Sildenafil for iatrogenic serotonergic antidepressant medication-induces sexual dysfunction in 4 patients. J. Clin. Psychiatry 60, 33-35.
Schrag, M.L., Wienkers, 2001, Triazolam substrate inhibition: evidence of competition for heme-bound reactive oxygen within the CYP3A4 active site. Drug Metab. Dispos. 29, 70-75.
Segelman, F.H., Kelton, E., Terzi, R.M., Kucharczy, N., Sofia, R.D., 1985, The comparative potency of phenobarbital and five 1,3-propanediol dicarbamates for hepatic cytochrome P450 induction in rats. Res. Commun. Chem. Pathol.
Pharmacol. 48, 467-470.
Sheu, M.T., Wu, A.B., Yeh, G.C., Hsia, A., Ho, H.O, 2003, Development of a liquid chromatographic method for bioanalytical applications with sildenafil, J.
Chromatogr. B 791, 255-262.
Shou, M., Dai, R., Cui, D., Korzekwa, K.R., Bailliet, T.A., Rushmore, T.H., 2001, A kinetic model for the metabolic interaction of two substrates at the active site of cytochrome P450 3A4. J. Biol Chem 272, 3149-3152.
Shou, M., Grogan, J., Mancewiez, J.A., Krausz, K.W., Gonzalez, F.J., Gelboin, H.V., Korzekwa, K.R., 1994, Activation of CYP3A4: evidence for the simultaneous binding of two substrates in a cytochrome P450 active site. Biochemistry 33,
格式化: 項目符號及編號
Shou, M., Mei, Q., Ettore, M.W., Dai, R., Baillie, T.A., Rushmore, T.H., 1999, Sigmoidal kinetic model for two cooperative substrate-binding sites in a cytochrome P450 3A4 active site: an example of the metabolism of diazepam and its derivatives. Biochem J 340, 845-853.
Ueng, Y.F., Kuwabara, T., Chun, Y.J., Guengerich, F.P., 1997 Cooperativity in oxidations catalyzed by cytochrome P450 3A4. Biochemistry 36, 370-381.
Walker, D.K., Ackland, M.J., James, G.C., Muirhead, G.J., Rance, D.J., Wastall, P., Wright, P.A., 1999, Pharmacokinetics and metabolism of sildenafil in mouse, rat, rabbit, dog and man. Xenobiotica 29, 297-310.
Wang, R.W., Newton, D.J., Liu, N., Atkins, W.M., Lu, A.Y.H., 2000, Human cytochrome P-450 3A4: in vitro drug-drug interaction patterns are substrate-dependent. Drug Metab. Dispos. 28, 360-366.
Wang, R.W., Newton, D.J., Scheri, T.D., Lu, A.Y.H., 1997, Human cytochrome P450 3A4-catalyzed testosterone 6-hydroxylation and erythromycin N-demethylation:
competition during catalysis. Drug Metab. Dispos. 25, 502-507.
Warrington, J.S., Shader, R.I., von Moltke, L.L., Greenblatt, D.J., 2000, In vitro biotransformation of sildenafil (Viagra): identification of human cytochromes and potential drug interactions. Drug Metab. Dispos. 28, 392-397.
Legends:
Figure 1. Formation of UK-103,320 (μg/ml) in the metabolic reaction of various concentrations of sildenafil incubated with rat microsomes for different incubation times (,5 min; , 10 min; , 20 min; , 40 min). (n=3)
Figure 2. Michaelis-Menten plot of the conversion of sildenafil to UK-103,320.
Figure 3. Formation of UK-103,320 (μg/ml), in the metabolism of sildenafil with the addition of different concentration of testosterone for different incubation times in rat microsomes. (n=3) (A, 10 μM; B, 25 μM; C, 100 μM; D, 200 μM)
Figure 4. Formation of UK-103,320 (μg/ml), in the metabolism of sildenafil with the addition of different concentrations of carbamazepine for different incubation times in rat microsomes. (n=3) (A, 20 μM; B, 50 μM; C, 150 μM; D, 600 μM)
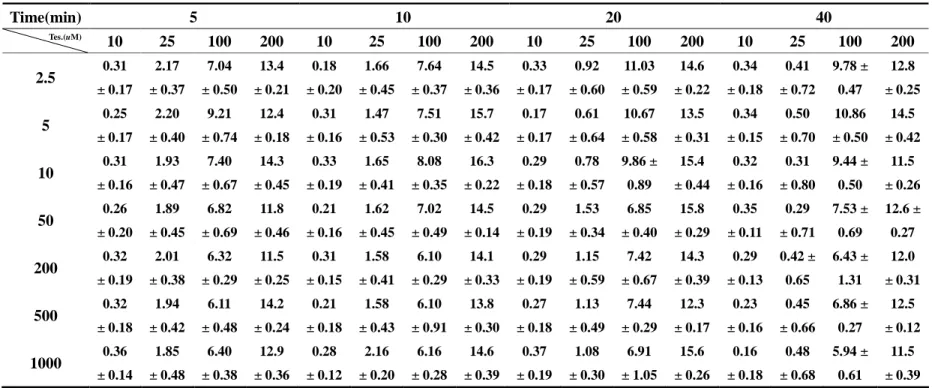
Table 1. Formation of hydroxyl testosterone (μg/ml) in the metabolism of different concentrations of testosterone with the addition of different concentrations of sildenafil for different incubation times in rat liver microsomes. (n=3)
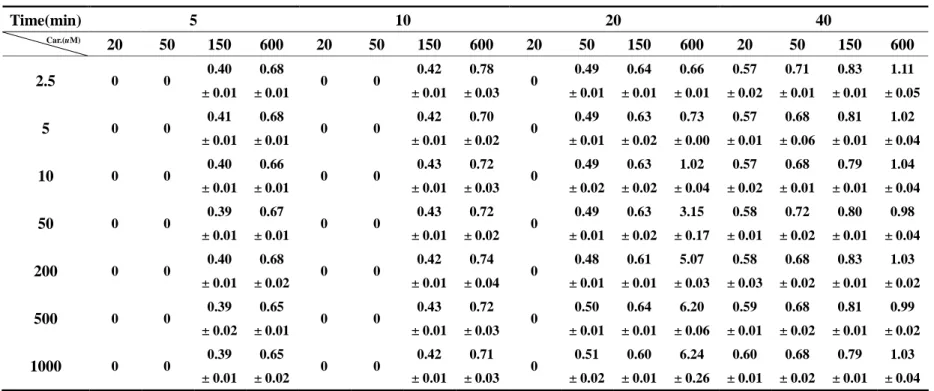
Table 2. Formation of 10-11 epoxide carbamazepine(μg/ml)in themetabolism of different concentrations of carbamazepine with the addition of different concentrations of carbamazepine for different incubation times in rat liver microsomes. (n=3)
Figures
Sildenafil concentration (uM)
0 200 400 600 800 1000 1200
U K -1 0 3 ,3 2 0 c o n ce n tr a ti o n (u M )
0 10 20 30 40 50
Figure 1
Sildenafil concentration (uM)
0 200 400 600 800 1000 1200
UK-103,320formationrate(uMmin-1 ) 0.0 0.5 1.0 1.5 2.0 2.5
Figure 2.
Sildenafil concentration(uM)
0 200 400 600 800 1000 1200
UK-103,320formation(uM)
0 2 4 6 8
10 5min
10min 20min 40min
Sildenafil concentration (uM)
0 200 400 600 800 1000 1200
UK-103,320concentration(uM)
0 2 4 6 8 10
5 min 10 min 20 min 40 min
Sildenafil concentration(uM)
0 200 400 600 800 1000 1200
UK-103,320concentration(uM)
0 2 4 6 8 10
5 min 10 min 20 min 40 min
Sildenail concentration(uM)
0 200 400 600 800 1000 1200
UK-103,320concentration(uM)
0 2 4 6 8 10
5min 10min 20min 40min
Figure 3
B
D A
C
Sildenafil concentration (uM)
0 200 400 600 800 1000 1200
UK-103,320formation(uM)
0 5 10 15
20 5 min
10 min 20 min 40 min
Sildenafil concentration (uM)
0 200 400 600 800 1000 1200
UK-103,320formation(uM)
0 2 4 6 8 10 12 14 16
5 min 10 min 20 min 40 min
Sildenafil concentration (uM)
0 200 400 600 800 1000 1200
UK-103,320formation(uM)
0 2 4 6 8 10 12 14
5 min 10 min 20 min 40 min
Sildenafil concentration (uM)
0 200 400 600 800 1000 1200
UK-103,320formation(uM)
0 1 2 3 4 5
6 5 min
10 min 20 min 40 min
Figure 4
B
D A
C
Table 1. Formation of hydroxyl testosterone (μg/ml) in the metabolism of different concentrations of testosterone with the addition of
different concentrations of sildenafil for different incubation times in rat liver microsomes. (n=3)
Time(min) 5 10 20 40
Tes.(uM)
10 25 100 200 10 25 100 200 10 25 100 200 10 25 100 200
2.5
0.31± 0.17 2.17
± 0.37 7.04
± 0.50 13.4
± 0.21 0.18
± 0.20 1.66
± 0.45 7.64
± 0.37 14.5
± 0.36 0.33
± 0.17 0.92
± 0.60 11.03
± 0.59 14.6
± 0.22 0.34
± 0.18 0.41
± 0.72
9.78 ± 0.47
12.8
± 0.25
5
0.25± 0.17 2.20
± 0.40 9.21
± 0.74 12.4
± 0.18 0.31
± 0.16 1.47
± 0.53 7.51
± 0.30 15.7
± 0.42 0.17
± 0.17 0.61
± 0.64 10.67
± 0.58 13.5
± 0.31 0.34
± 0.15 0.50
± 0.70 10.86
± 0.50 14.5
± 0.42
10
0.31± 0.16 1.93
± 0.47 7.40
± 0.67 14.3
± 0.45 0.33
± 0.19 1.65
± 0.41 8.08
± 0.35 16.3
± 0.22 0.29
± 0.18 0.78
± 0.57
9.86 ± 0.89
15.4
± 0.44 0.32
± 0.16 0.31
± 0.80
9.44 ± 0.50
11.5
± 0.26
50
0.26± 0.20 1.89
± 0.45 6.82
± 0.69 11.8
± 0.46 0.21
± 0.16 1.62
± 0.45 7.02
± 0.49 14.5
± 0.14 0.29
± 0.19 1.53
± 0.34 6.85
± 0.40 15.8
± 0.29 0.35
± 0.11 0.29
± 0.71
7.53 ± 0.69
12.6 ± 0.27
200
0.32± 0.19 2.01
± 0.38 6.32
± 0.29 11.5
± 0.25 0.31
± 0.15 1.58
± 0.41 6.10
± 0.29 14.1
± 0.33 0.29
± 0.19 1.15
± 0.59 7.42
± 0.67 14.3
± 0.39 0.29
± 0.13
0.42 ± 0.65
6.43 ± 1.31
12.0
± 0.31
500
0.32± 0.18 1.94
± 0.42 6.11
± 0.48 14.2
± 0.24 0.21
± 0.18 1.58
± 0.43 6.10
± 0.91 13.8
± 0.30 0.27
± 0.18 1.13
± 0.49 7.44
± 0.29 12.3
± 0.17 0.23
± 0.16 0.45
± 0.66
6.86 ± 0.27
12.5
± 0.12
1000
0.36± 0.14 1.85
± 0.48 6.40
± 0.38 12.9
± 0.36 0.28
± 0.12 2.16
± 0.20 6.16
± 0.28 14.6
± 0.39 0.37
± 0.19 1.08
± 0.30 6.91
± 1.05 15.6
± 0.26 0.16
± 0.18 0.48
± 0.68
5.94 ± 0.61
11.5
± 0.39
Table 2. Formation of 10-1 1 e poxi de c ar bamaz e pi ne ( μg/ ml ) i n t he me t abol i s m of different concentrations of carbamazepine with the addition of different concentrations of carbamazepine for different incubation times in rat liver microsomes. (n=3)
Time(min) 5 10 20 40
Car.(uM)
20 50 150 600 20 50 150 600 20 50 150 600 20 50 150 600
2.5
0 0 0.40± 0.01 0.68
± 0.01 0 0 0.42
± 0.01 0.78
± 0.03 0 0.49
± 0.01 0.64
± 0.01 0.66
± 0.01 0.57
± 0.02 0.71
± 0.01 0.83
± 0.01 1.11
± 0.05
5
0 0 0.41± 0.01 0.68
± 0.01 0 0 0.42
± 0.01 0.70
± 0.02 0 0.49
± 0.01 0.63
± 0.02 0.73
± 0.00 0.57
± 0.01 0.68
± 0.06 0.81
± 0.01 1.02
± 0.04
10
0 0 0.40± 0.01 0.66
± 0.01 0 0 0.43
± 0.01 0.72
± 0.03 0 0.49
± 0.02 0.63
± 0.02 1.02
± 0.04 0.57
± 0.02 0.68
± 0.01 0.79
± 0.01 1.04
± 0.04
50
0 0 0.39± 0.01 0.67
± 0.01 0 0 0.43
± 0.01 0.72
± 0.02 0 0.49
± 0.01 0.63
± 0.02 3.15
± 0.17 0.58
± 0.01 0.72
± 0.02 0.80
± 0.01 0.98
± 0.04
200
0 0 0.40± 0.01 0.68
± 0.02 0 0 0.42
± 0.01 0.74
± 0.04 0 0.48
± 0.01 0.61
± 0.01 5.07
± 0.03 0.58
± 0.03 0.68
± 0.02 0.83
± 0.01 1.03
± 0.02
500
0 0 0.39± 0.02 0.65
± 0.01 0 0 0.43
± 0.01 0.72
± 0.03 0 0.50
± 0.01 0.64
± 0.01 6.20
± 0.06 0.59
± 0.01 0.68
± 0.02 0.81
± 0.01 0.99
± 0.02
1000
0 0 0.39± 0.01 0.65
± 0.02 0 0 0.42
± 0.01 0.71
± 0.03 0 0.51
± 0.02 0.60
± 0.01 6.24
± 0.26 0.60
± 0.01 0.68
± 0.02 0.79
± 0.01 1.03
± 0.04