嘉南藥理科技大學專題研究計畫成果報告
檳榔成份誘發細胞自體吞噬訊息傳導路徑之探討
計畫類別:個別型計畫
計畫編號:CN9623
執行期間:96 年 1 月 1 日至 96 年 12 月 31 日
計畫主持人:林美惠
共同主持人:劉永超
計畫參與人員:施雅云
執行單位:生物科技系碩士班
中華民國97 年 3 月 27 日
Arecoline and the 30-100 kDa fraction of areca nut extract
differentially regulate mTOR and respectively induce apoptosis and autophagy—A pilot study
Chin-Hai Lee a, Mei-Huei Lin b,c, Shyun-Yeu Liu d-f, Yu-Rung Hsu b, Ya-Yun Shih b, Wei-Fan Chiang g, Da-Shiung Chou a, Young-Chau Liu c,h,*
a Department of Dentistry, Chi Mei Medical Center, Tainan,
b Department of Biotechnology, Chia Nan University of Pharmacy and Science,
Tainan,
c Department of Medical Research, Chi Mei Medical Center, Tainan,
d Oral and Maxillofacial Surgery Section, Chi Mei Medical Center, Tainan,
e Department of Dentistry, National Defense Medical Center, Taipei,
f Department of Dentistry, Taipei Medical University, Taipei,
gOral and Maxillofacial Surgery Section, Chi Mei Hospital, Liouying,
h College of Liberal Education, Shu-Te University, Kaohsiung County, Taiwan,
ROC
Keywords: Arecoline; Areca nut; Apoptosis; Autophagy; mTOR
* Corresponding author:
Dr. Young-Chau Liu
Division of Natural Science, College of Liberal Education, Shu-Te University, No. 59, Hun Shan Road, Yen Chau 824, Kaohsiung County, Taiwan, ROC.
Tel: 886-7-6158000 ext.4210, Fax: 886-6-2909635,
E-mail: [email protected]
Note: The first four authors contributed equally to this work.
Abstract
Areca nut (AN) is recognized as a human carcinogen; however, few studies of the cytotoxic effects of AN ingredients on cells have been reported.
In Taiwan, AN, lime and inflorescence of Piper betleare the common
components of betel quid. We recently noticed that extract of AN (ANE), but not those of lime and inflorescence of Piper betle, induces rounding cell
morphology and nuclear shrinkage in different types of carcinoma cells. In this study, the rounding cell activity was first traced to the partially purified ≥ 10 kDa fraction (ANE ≥ 10K) and subsequently to the 30-100 kDa fraction (ANE 30-100K). ANE and ANE ≥ 10K stimulated nuclear shrinkage (P < 0.001 in both cases) and the clearance of the cytoplasm. ANE, ANE ≥ 10K, and ANE 30-100K induced the cleavage of LC3-I (P < 0.05, 0.01, and 0.05, respectively) and the emergence of autophagic vacuoles and acidic vesicles. On the other hand, arecoline (Are, the major alkaloid of AN) triggered caspase-3 activation, peri-nuclear chromatin condensation, and micronucleation. Meanwhile, ANE 30-100K, but not Are, inhibited the phosphorylation of the mammalian target of rapamycin (mTOR)-Ser2448. In conclusion, this study demonstrates that different AN ingredients exerting differential impact on mTOR-Ser2448 phosphorylation are capable of triggering apoptosis and autophagy.
1. Introduction
Betel quid (BQ) chewing may represent one of the most popular oral habits with about 600 million users worldwide, and is thought to cause oral leukoplakia, oral submucous fibrosis, and oral cancer (Jeng et al., 2001). In Taiwan, BQ is usually composed of areca nut (AN, Areca catechu L.) and lime, with or without a piece of inflorescence of Piper betle (IPB) or Piper betle leaf.
Around 10% of the Taiwanese population (2-2.8 million people) has this oral habit. Clinical observations support that habitual BQ chewing raises the risk of oral (Jeng et al., 2001), pharynx (Lee et al., 2005a; Tominaga, 1999),
esophagus (Wu et al., 2003; Lee et al, 2005b), liver (Hsiao et al., 2007; Tsai et al., 2004), and stomach (Wu et al., 2004) cancers. Therefore, mechanisms implicated in the cytotoxic impact of AN ingredients on cells are worthy of study.
Some ingredients of AN and IPB, such as arecoline (Are) and
hydroxychavicol, have been shown to induce growth arrest, necrosis, and apoptosis (Chang et al., 2001; Lee et al., 2006; Chang et al., 2002). To our knowledge, there is no evidence so far demonstrating the existence of an autophagy-inducing activity in AN constituents. Autophagy is a regulated self-cannibalism, classified as type II programmed cell death, and is preceded
by the inhibition of the mammalian target of rapamycin (mTOR) (Klionsky, 2004). Numerous hallmarks used for demonstrating autophagy have been developed: for example, the cleavage of the precursor form of
microtubule-associated protein 1 light chain 3 (LC3-I) (18 kDa) to the active form LC3-II (16 kDa) and the emergence of autophagic vacuoles (AVs) and acidic vesicles (Klionsky et al., 2007).
In our previous study, we found that some matrix metalloproteinases are upregulated by arecoline (Are) and/or extracts of lime, AN, and IPB (Liu et al., 2007). Surprisingly, we have recently noticed a unique death pattern induced by the AN extract (ANE) in several types of carcinoma cells from mouth (OECM-1 and SCC-9), bladder (UB-09 and T24), and esophageal
(CE81T/VGH) origins. These dying cells exhibit rounding morphology with a shrunken nucleus and empty cytoplasm. Using OECM-1 cells as the
experimental model, we partially isolated these activities. The rounding
cell-inducing activity in ANE was first traced to the ≥ 10 kDa fraction (ANE ≥ 10K) and subsequently the 30-100 kDa fraction (ANE 30-100K). Only the cell shrinkage activity without rounded morphology was induced by cisplatin, Are, and extracts of lime and IPB. Thus, Are may be a potential apoptosis inducer whereas the ANE, ANE ≥ 10K, and ANE ≥ 30-100K fractions may induce an
unknown death pathway in OECM-1 cells (data are presented in this study).
We have further tried to verify the different death pathways mediated by these reagents by analyzing morphological change, activation status of caspase-3, cleavage of LC3-I, and emergence of AVs and acidic vesicles in OECM-1 cells. Finally, regulation of the phosphorylation status of
mTOR-Ser2448 by Are and ANE 30-100K was also examined.
2. Materials and methods
2.1. Preparation of ANE, ANE ≥ 10K, lime, and IPB Extract
The preparation of ANE was different from other researchers and was similar to that of BQ extract in our previous study (Liu et al., 2007). Briefly, ANE was prepared by grinding tender AN (3.6~4.0 g/nut) at RT, and the squeezed juice was centrifuged at 12,000 × g for 10 min. The supernatant was then regarded as the ANE. ANE was further centrifuged at 2,900 × g for 30 min at 4˚C with 10, 30, or 100 kDa-pored membranous concentration tubes. Thus, ANE and fractions of ANE ≤ 10K, ANE ≥ 10K, and ANE 30-100K used in this study were obtained. They were lyophilized, weighed and re-dissolved in H2O.
The lime and IPB were separated from a commercial BQ and ground in 1 ml H2O. Both extracts were then prepared by the same method as ANE.
2.2. Culture and treatment of cells
Cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM) (Gibco-BRL, Rockville, MD, USA) and 1 × 104 cells were seeded into each
well of a 96-well plate for the observation of morphological change, or into each well of a slide chamber for the nuclear staining, immunofluorescent
analysis, and acridine orange staining, each observed in triplicate. Alternatively, 6 × 106 cells were seeded onto a 10-cm plate for caspase-3 activity, Western blot, TEM, and flow cytometrical analyses and subjected to a 24-h treatment with each reagent under serum-free conditions.
To determine the 50% inhibitory concentration (IC50) of each reagent, cells treated with various concentrations of each reagent were analyzed by XTT reagents (Roche Molecular Biochemicals, Basel, Switzerland) as instructed by the manufacturer.
2.3. Measurement of nuclear diameter
Treated samples on chamber slides were fixed with 3% paraformaldehyde for 30 min, and treated with 0.1% Triton X-100 for 15 min. The slides were coated with 3% BSA for 1 h and stained with 0.01% Hoechst 33258 for 10 min.
The stained cells were photographed under a fluorescent microscope (OLYMPUS DP70), and the diameters of 15 randomly chosen nuclei were measured using a micrometer.
2.4. Measurement of caspase-3 activation status
Treated OECM-1 cells were incubated with 100-fold diluted anti-active caspase-3 antibody (Sigma-Aldrich, USA) for 1.5 h, followed by further incubation with goat anti-rabbit-IgG antibody coupled with fluorescein isothiocyanate (Sigma-Aldrich) for 40 min. Cells were then incubated with 0.1% propidium iodide for 10 min, and photographed under a fluorescent microscope (OLYMPUS DP70).
For the enzyme activity assay, treated OECM-1 cells (as in the
immunofluorescent assay) were lysed in lysis buffer (0.5% NP-40, 50 mM Tris-HCl, pH 7.4). Lysate proteins (100 µg) from each treatment were subjected to caspase-3 activity assay with the Caspase-3 Cellular Assay Kit (Biomol International, PA, USA) as instructed by the manufacturer.
2.5. Observation of morphological change, AVs, and acidic vesicles
Treated samples were observed and photographed using light or electron microscopy. For TEM analysis, samples were fixed in 2.5% glutaraldehyde at
4°C for 1 h and soaked in 1 % OsO4 for 20 min. After dehydration in an ethanol gradient (50% [10 min], 75% [10 min], 85% [10 min], and 95% alcohol [2 × 10 min]), samples were treated with propylene oxide (2 × 10 min), impregnated witha 1:1 mixture of propylene oxide/EA (Epon 812:Araldite
502:DDSA:DMP-30 =1.34:0.93:2.60:0.14, Electron Microscopy Sciences, PA, USA), and embedded in 100% EA. Ultrathin sections werestained with uranyl acetate and lead citrate. Sections wereexamined by TEM(JEOL JEM-1200EX) at 80 kV. Photographs were made with electron microscopyfilm 4489 Estar Thick Base (Kodak, NY, USA).
For the observation of acidic vesicles, treated samples were stained with acridine orange (1 µg/ml) for 10 min and photographed under a fluorescent microscope, or flow cytometrically analyzed by 525 nm (FL1) and 575 nm (FL2) with FACSCalibur (BD Biosciences, NJ, USA).
2.6. Western blot analysis
Lysate proteins (60 µg) from treated OECM-1 cells were subjected to Western blot analysis following the protocols previously described (Liu et al., 2007). Briefly, the first antibodies used against LC3 (MBL International
Corporation, MA, USA), mTOR, and phosphor-mTOR-Ser2448 (Cell Signaling Technology, Inc., MA, USA), as well as β-actin proteins (Sigma-Aldrich) were 1,000- and 10,000-fold diluted, respectively. The secondary antibody was goat anti-mouse-IgG or goat anti-rabbit-IgG coupled with horseradish peroxidase (Sigma-Aldrich) and 10,000-fold diluted.
2.7. Statistical analysis
Two groups of data presented as mean ± SD were analyzed by Student’s t test. A value of P < 0.05 was regarded as statistically significant.
3. Results
3.1. Characterization of the unique cytotoxic effect of ANE
Extracts of the three BQ components (lime, AN, and IPB), Are, and cisplatin exhibited cytotoxic effects on OECM-1 cells. The average IC50 of these reagents is listed in Table 1. OECM-1 cells treated with the IC50 of these reagents showed a shrunken phenotype, except that the ANE-treated cells exhibited rounding membrane and condensed nucleus-like spheres (Fig. 1A).
The rounding cell-inducing activity of ANE was also observed in other
carcinoma cells, including SCC-9, UB-09, T24, and CE81T/VGH (Fig. 1B). To address the molecular weight of this activity, the ANE was separated into two fractions by passing it through a centrifugal concentration tube with 10 kDa pores. The results showed that this activity was in the ANE ≥ 10K fraction (1120 µg/ml), whereas fractions of ANE ≤ 10K (1120 µg/ml), lime ≥ 10K (400 µg/ml), lime ≤ 10K (400 µg/ml), IPB ≥ 10K (220 µg/ml) and IPB ≤ 10K (220 µg/ml) only induced cell shrinkage (Fig. 1C).
After treatment with the IC100 of ANE and ANE ≥ 10K, most of the OECM-1 cells exhibited rounding morphology, and each of them contained a
shrunk nucleus-like sphere (data not shown). We further subjected these cells to Hoechst 33258 staining, and found that the suspected nucleus-like spheres were stainable by this dye (Fig. 1D). Compared to the non-treated control cells, the nuclei were significantly smaller in ANE- and ANE ≥ 10K-treated cells (Fig.
1E; P < 0.001 in both cases, n = 15).
3.2. Induction of caspase-3 activation by ANE and Are
Caspase-3 is known to be one of the down-stream effector caspases of apoptosis (Porter and Janicke, 1999). Although Are has been shown to induce apoptosis in KB cells (Lee et al., 2006), it remains uncertain whether it induces apoptosis in OECM-1 cells. Furthermore, it is not known whether this protease plays a role in ANE- and ANE ≥ 10K-mediated signals. Thus, the induction of caspase-3 activation by Are, ANE, ANE ≥ 10K, and cisplatin (used as the positive control) was analyzed. First, immunofluorescent analysis with
anti-active caspase-3 antibody revealed that cells treated with the IC50 of ANE, Are, and cisplatin exhibited stronger staining than those of control and ANE ≥ 10K (IC50)-treated cells (Fig. 2A). Secondly, protease activity assessed with a commercial kit also demonstrated a significant elevation of caspase-3 activity
in the lysates of ANE-, Are-, and cisplatin-treated cells compared to that of ANE ≥ 10K-treated cells (Fig. 2B; P < 0.05, 0.01, and 0.05, respectively).
3.3. Induction of cellular autophagy by ANE, ANE ≥ 10K, and ANE 30-100K To verify whether ANE ≥ 10K induces autophagy, the ratio of
LC3-II/LC3-I, a specific index of autophagy, was analyzed by Western blot.
The results showed that the IC50 of ANE and ANE ≥ 10K treatment
significantly enhanced LC3-I cleavage in OECM-1 cells (Fig. 3A, Fig. 4C and 4D; P < 0.05 and 0.01, respectively). Moreover, ANE ≥ 10K was further shown to dose-dependently increase the LC3-II/LC3-I ratio (Fig. 3A).
Further, OECM-1 cells were subjected to TEM analysis after the IC50 of ANE and ANE ≥ 10K treatments. The results showed that numerous AVs appeared in ANE [Fig. 3B, ANE (1)]- and ANE ≥ 10K [Fig. 3B, ≥ 10K (1)]-treated cells, whereas the Are (IC50)-treated cells exhibited peri-nuclear chromatin condensation [Fig. 3B, Are (1)] and micronucleation [Fig. 3B, Are (2)]. Interestingly, in some ANE- and ANE ≥ 10K-treated cells, the cytoplasmic substances, including nucleus, were massively degraded before the
disintegration of the cell membrane [Fig. 3B, ANE (2) and ≥ 10K (2)].
The IC50 of ANE ≥ 10K is about 30-fold greater than ANE (Table 1),
meaning that the autophagy-inducing component was not enriched in the ANE
≥10 kDa fraction. We further used concentration tubes with 30 and 100 kDa pores to separate ANE, and found that the rounding cell-inducing activity was localized in the 30-100 kDa fraction (ANE 30-100K, Fig. 4A). The IC50 of ANE 30-100K against OECM-1 cells was 15 ± 5 µg/ml (Table 1), about 2.5-fold lower than that of ANE. Thus, the effectiveness of ANE ≥ 10K in autophagy induction may be interfered with by 10-30 kDa molecules in ANE.
ANE 30-100K (IC50) also induced numerous AVs in OECM-1 cells (Fig. 4B), and significant LC3-I cleavage (Fig. 4C and 4D; P < 0.05).
3.4. Induction of acidic vesicles by ANE, ANE ≥ 10K, and ANE 30-100K Acidic vesicles are also one the autophagy hallmarks. We found that cells treated with the IC50 of ANE, ANE ≥ 10K, and ANE 30-100K, as well as those cultured in non-glucose medium (used as the positive control), exhibited numerous acidic vesicles after acridine orange staining. In contrast, the acidic vesicles were barely detected in non-treated control and Are (IC50)-treated cells (Fig. 5A). The percentages of acidic vesicle-containing cells were 1.0%, 88.0%, 90.2%, 88.9%, and 16.3% in control, glucose-starved, ANE-, ANE 30-100K-, and Are-treated cells as analyzed by flow cytometry, respectively (Fig. 5B).
3.5. Differential regulation of the mTOR-Ser2448 phosphorylation by Are and ANE 30-100K
The phosphorylation status of mTOR-Ser2448 was recently shown to be correlated with the extent of autophagy (Morimoto et al., 2007). We were thus interested to assess whether it is differentially regulated by Are and ANE 30-100K. The results showed that the phosphorylation level of mTOR-Ser2448 was significantly inhibited after 24-h treatment with ANE 30-100K (IC50) (Fig.
6A and 6C; P < 0.05). In contrast, Are (IC50) treatment did not attenuate the phosphorylation level on this site within 24 h (Fig. 6B and 6C).
4. Discussion
We have demonstrated that ANE, ANE ≥ 10K, and ANE 30-100K are capable of triggering cellular autophagy. To our knowledge, this study has provided the first evidences of an autophagy-inducing AN ingredient (AIAI). In addition to KB cells (Lee et al., 2006), Are was also shown to induce apoptosis in OECM-1 cells. Thus, AN may contain both apoptosis- and
autophagy-inducing ingredients.
The shrunken nuclei in OECM-1 cells may represent the unique autophagic death pattern induced by AIAI. It might be thought that the
degradation of nuclear contents is strictly "off-limits", given the importance of maintaining the integrity of genes, but it has been reported that partial
degradation of the nucleus may occur in yeast (Mijaljica et al., 2007). In Saccharomyces cerevisiae, a process called "piecemeal microautophagy of the nucleus" (PMN) pinches off and degrades nonessential portions of the nucleus.
PMN is a constitutive process induced to high levels by starvation or
rapamycin and occurs at nucleus-vacuole junctions (Kvam and Goldfarb, 2007).
It might be reasonable to speculate that nuclear contents may be degraded progressively, which reduces nuclear size in the process of autophagic death.
Clarke et al. observed that during the autophagic death of isthmo-optic neurons
in chick embryos, nuclei of the dying neurons lost more than half of their DNA content and became more electron dense and shrank (Clarke and Hornung, 1989). Here, we have illustrated that ANE and ANE ≥ 10K also induced significant nuclear shrinkage (Fig. 1D and 1E). Taken together, it is suggested that the nucleus may be shrunk before its disintegration in different modes of autophagy.
It is speculated that the rounding morphology of SCC-9, UB-09, T24, and CE81T/VGH cells induced by ANE treatment reflects the initiation of an autophagic pathway in these different cancer cells. We have recently observed the same morphological change in a primary culture of oral fibroblasts and three independent and normal human fibroblasts after ANE 30-100K treatment (data not shown). Thus, AIAI may have the potential to transmit autophagic signals to both normal and malignant cells.
Although various modes of autophagy conspire to degrade virtually every compartment of the eukaryotic cell, the plasma membrane and nucleus may be the most resistant compartments during autophagic degradation. Szende et al.
observed that these two compartments were resistant to L-leucine methyl ester-induced autophagic digestion in mouse peritoneal macrophages (Szende et al., 1991). Gómez-Santos et al. also demonstrated that the plasma membrane
and nucleus maintained their integrity in the dopamine-induced autophagy in human neuroblastoma cells (Gómez-Santos et al., 2003). In our study, the cell membrane and nucleus also seemed to be the dominant remnants in the
AIAI-induced autophagy as revealed by light microscopy (Fig. 1A-C and Fig.
4A). Furthermore, the degradation of the nucleus before the rupture of the plasma membrane after ANE and ANE ≥ 10K treatments [Fig. 3B, ANE (2) and ≥ 10K (2)] suggests that in the AIAI-induced autophagy, the cell membrane may be the most stable compartment in some OECM-1 cells.
Increase of caspase-3 activity may not necessarily result in cellular
apoptosis. Sadasivan et al. recently showed that withdrawing amino acids from the culture medium of PC-12 cells activated this protease but induced
autophagic cell death (Sadasivan et al., 2006). Similarly, we have also
demonstrated that although caspase-3 was evidently activated by ANE (Fig. 2), meaning that the apoptotic signals might have been turned on by the
apoptosis-inducing ingredients, the cells underwent autophagy. The AIAI (most likely in the ANE 30-100K fraction) may thus trigger the dominant autophagic phenotype in the presence of apoptosis-inducing molecules, like
hydroxychavicol and Are.
Artificial inhibition of mTOR by rapamycin, for example, often results in
autophagy (Cao et al., 2006). Therefore, inhibition of mTOR-Ser2448
phosphorylation by ANE 30-100K suggests that the induction of autophagy by AIAI may be due to the inhibition of mTOR activity. On the other hand,
although Are does not affect the phosphorylation level of mTOR-Ser2448, whether it interferes with other regulators along the mTOR pathway remains unknown. In conclusion, the identification of apoptosis and autophagy induced by Are and ANE ≥ 10K (and ANE 30-100K) respectively may serve as an ideal model to further delineate the difference and interaction between these two death pathways in our future studies.
Acknowledgement
This work was supported in part by grants from the National Science
Council (96-2314-B-041-001) and Chi Mei Medical Center (CMFHR 9715 and CLFHR 9703).
References
Cao C., Subhawong T., Albert J.M., Kim K.W., Geng L., Sekhar K.R., Gi Y.J., Lu B., 2006. Inhibition of mammalian target of rapamycin or apoptotic pathway induces autophagy and radiosensitizes PTEN null prostate cancer cells. Cancer Research 66 (20), 10040-10047.
Chang M.C., Ho Y.S., Lee P.H., Chan C.P., Lee J.J., Hahn .LJ., Wang Y.J., Jeng J.H., 2001. Areca nut extract and arecoline induced the cell cycle arrest but not apoptosis of cultured oral KB epithelial cells: association of glutathione, reactive oxygen species and mitochondrial membrane potential.
Carcinogenesis 22 (9), 1527-1535.
Chang M.C., Uang B.J., Wu H.L., Lee J.J., Hahn L.J., Jeng J.H., 2002.
Inducing the cell cycle arrest and apoptosis of oral KB carcinoma cells by hydroxychavicol: roles of glutathione and reactive oxygen species. British Journal of Pharmacology 135 (3), 619-630.
Clarke P.G., Hornung J.P., 1989. Changes in the nuclei of dying neurons as studied with thymidine autoradiography. The Journal of Comparative Neurology 283 (3), 438-449.
Hsiao T.J., Liao H.W., Hsieh P.S., Wong R.H., 2007. Risk of betel quid
chewing on the development of liver cirrhosis: a community-based case-control study. Annals of Epidemiology 17 (6), 479-485.
Jeng J.H., Chang M.C., Hahn L.J., 2001. Role of areca nut in betel
quid-associated chemical carcinogenesis: current awareness and future perspectives. Oral Oncology 37 (6), 477-492.
Klionsky D.J., 2004. Cell biology: regulated self-cannibalism. Nature 431 (7004), 31-32.
Klionsky D.J., Cuervo A.M., Seglen P.O., 2007. Methods for monitoring autophagy from yeast to human. Autophagy 3 (3), 181-206.
Kvam E., Goldfarb D.S., Nucleus-vacuole junctions and piecemeal
microautophagy of the nucleus in S. cerevisiae. Autophagy 3 (2), 85-92.
Lee K.W., Kuo W.R., Tsai S.M., Wu D.C., Wang W.M., Fang F.M., Chiang F.Y., Ho K.Y., Wang L.F., Tai C.F., Kao E.L., Chou S.H., Lee C.H., Chai C.Y., Ko Y.C., 2005a. Different impact from betel quid, alcohol and cigarette: risk factors for pharyngeal and laryngeal cancer. International Journal of Cancer 117 (5), 831-836.
Lee C.H., Lee J.M., Wu D.C., Hsu H.K., Kao E.L., Huang H.L., Wang T.N., Huang M.C., Wu M.T., 2005b. Independent and combined effects of alcohol intake, tobacco smoking and betel quid chewing on the risk of esophageal
cancer in Taiwan. International Journal of Cancer 113 (3), 475-482.
Lee PH, Chang MC, Chang WH, Wang TM, Wang YJ, Hahn LJ, Ho Y.S., Lin C.Y., Jeng J.H., 2006. Prolonged exposure to arecoline arrested human KB epithelial cell growth: regulatory mechanisms of cell cycle and apoptosis.
Toxicology 220 (2-3), 81-89.
Liu S.Y., Liu Y.C., Huang W.T., Huang G.C., Chen T.C., Lin M.H., 2007.
Up-regulation of matrix metalloproteinase-8 by betel quid extract and arecoline and its role in 2D motility. Oral Oncology 43 (10), 1026-1033.
Mijaljica D., Prescott M., Devenish R.J., 2007. Nibbling within the nucleus:
turnover of nuclear contents. Cellular and Molecular Life Sciences 64 (5), 581-588.
Morimoto N., Nagai M., Ohta Y., Miyazaki K., Kurata T., Morimoto M., Murakami T., Takehisa Y., Ikeda Y., Kamiya T., Abe K., 2007. Increased autophagy in transgenic mice with a G93A mutant SOD1 gene. Brain Research 1167, 112-117.
Porter A.G., Janicke R.U., 1999. Emerging roles of caspase-3 in apoptosis. Cell Death and Differentiation 6 (2), 99-104.
Sadasivan S., Waghray A., Larner S.F., Jr. Dunn W.A., Hayes R.L., Wang K.K., 2006. Amino acid starvation induced autophagic cell death in PC-12
cells: evidence for activation of caspase-3 but not calpain-1. Apoptosis 11 (9), 1573-1582.
Tominaga S., 1999. Major avoidable risk factors of cancer. Cancer Letter 143 (Suppl 1), S19-S23.
Tsai J.F., Jeng J.E., Chuang L.Y., Ho M.S., Ko Y.C., Lin Z.Y., Hsieh M.Y., Chen S.C., Chuang W.L., Wang L.Y., Yu M.L., Dai C.Y., 2004. Habitual betel quid chewing and risk for hepatocellular carcinoma complicating cirrhosis. Medicine 83 (3), 176-187.
Wu M.T., Wu D.C., Hsu H.K., Kao E.L., Lee J.M., 2003. Relationship between site of oesophageal cancer and areca chewing and smoking in Taiwan.
British Journal of Cancer 89 (7), 1202-1204.
Wu M.T., Chen M.C., Wu D.C., 2004. Influences of lifestyle habits and p53 codon 72 and p21 codon 31 polymorphisms on gastric cancer risk in Taiwan.
Cancer Letter 205 (1), 61-68.
Szende B., Lapis K., Timar J., Antoni F., Csuka I., 1991. Morphological studies on the effect of L-leucine methyl ester on mouse peritoneal macrophages in vitro. Experimental Pathology 42 (3), 121-127.
Gómez-Santos C., Ferrer I., Santidrián A.F., Barrachina M., Gil J., Ambrosio S., 2003. Dopamine induces autophagic cell death and alpha-synuclein
increase in human neuroblastoma SH-SY5Y cells. Journal of Neuroscience Research 73 (3), 341-350.
Figure legends
Fig. 1. Unique death pattern induced by ANE. (A) OECM-1 cells treated with the IC50 of lime, IPB, ANE, Are, and cisplatin (Cis) were photographed under light microscope. Ctr: non-treated control cells. (B) SCC-9, UB-09, T24, and CE81T/VGH (CE81T) cells were treated with or without ANE (20 µg/ml) and photographed. (C) OECM-1 cells treated with two different fractions of lime [lime ≤ 10K (400 µg/ml) and lime ≥ 10K (400 µg/ml)], ANE [ANE ≤10K (1120 µg/ml) and ANE ≥ 10K (1120 µg/ml)], and IPB [IPB ≤ 10K (220 µg/ml) and IPB ≥ 10K (220 µg/ml)] were photographed. Bar (A-C) = 10 µm. (D) OECM-1 cells treated with the IC100 of ANE and ANE ≥ 10K were stained by 0.01% Hoechst 33258 and photographed under fluorescent microscope. Bar = 5 µm. (E) Average nuclear diameter of these ANE- and ANE ≥ 10K (≥
10K)-treated OECM-1 cells was presented as mean ± SD. These experiments were repeated at least for three times with similar results. ***P < 0.001. Ctr:
non-treated control cells.
Fig. 2. Activation of caspase-3 by ANE, Are, and cisplatin. (A) OECM-1 cells treated with the IC50 of ANE, ANE≥ 10K, Are, and cisplatin (Cis) were
analyzed by immunofluorescent staining with anti-active caspase-3 antibody and photographed under fluorescent microscope. Bar = 20 µm. (B) Lysate proteins (100 µg) from OECM-1 cells identically treated as in (A) were
analyzed with Caspase-3 Cellular Assay Kit. Average OD405 absorbance of each treatment was presented as mean ± SD. These experiments were repeated at least for three times with similar results. Ctr: non-treated control cells. ≥ 10K:
ANE≥ 10K. *P < 0.05, **P < 0.01.
Fig. 3. Induction of cellular autophagy by ANE, ANE ≥ 10K. (A) LC3 proteins in OECM-1 cells treated with ANE (IC50) and the indicated concentrations of ANE ≥ 10K were analyzed by Western blot. The
LC-3-II/LC3-I was listed under each treatment. (B) OECM-1 cells treated with the IC50 of ANE, ANE ≥ 10K (≥ 10K), and Are were subjected to TEM analysis.
Two representative pictures from each treatment were shown. N: nucleus; mN:
micronucleated nucleus. Bar = 2 µm. These experiments have been repeated at least twice.
Fig. 4. Induction of cellular autophagy by ANE 30-100K. (A) OECM-1 cells treated with or without ANE 30-100K (IC50) (30-100K) were photographed
under light microscope. Bar = 10 µm. (B) ANE 30-100K-treated cells were further subjected to TEM analysis. Bar = 2 µm. (C) LC3 proteins in OECM-1 cells treated with the IC50 of Are, ANE, ANE 30-100K (30-100K), and ANE ≥ 10K (≥ 10K) were analyzed as in Fig. 3A. Ctr: non-treated control cells. (D) Average LC3-II/LC3-I ratios from three independent experiments were repeated as mean ± SD. *P < 0.05, **P < 0.01.
Fig. 5. Induction of acidic vesicles by ANE, ANE ≥ 10K, and ANE 30-100K.
(A) Non-treated control (Ctr), glucose-starved (Non-glucose), ANE (IC50)-, ANE ≥ 10K (IC50)-, ANE 30-100K (IC50)-, and Are (IC50)-treated cells were stained by acridine orange and photographed under fluorescent microscope.
Bar = 5 µm. (B) The Ctr, Non-glucose-, ANE-, ANE 30-100K (30-100K)-, and Are-treated cells were further subjected to flow cytometric analysis.
Fig. 6. Differential inhibition of mTOR-Ser2448 phosphorylation by Are and ANE 30-100K. Phosphorylated mTOR-Ser2448 (pmTOR-Ser2448), total mTOR (mTOR-Total), and β-actin proteins in OECM-1 cells treated with the IC50 of ANE-30-100K (A) and Are (B) for the indicated periods were analyzed by Western blot. (C) Average pmTOR-Ser2448/mTOR-Total ratios of ANE 30-100K
(IC50, 24 h) - and Are (IC50, 24 h)-treated cells from three independent
experiments were compared to that of non-treated control cells and presented as mean ± SD. *P < 0.05.
Figure 1
Figure 1
Figure 2
Figure 3
Figure 4
Figure 5
Figure 6
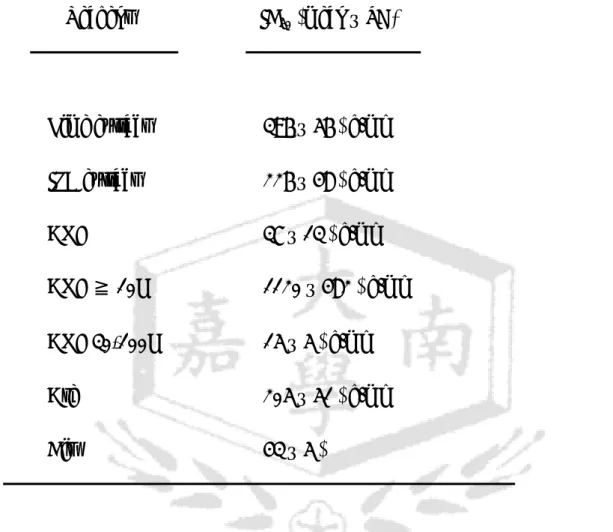
Table 1. The average IC50 of each reagent against OECM-1.
Reagent IC50 (mean ± SD)
Lime extract 396 ± 56 µg/ml IPB extract 226 ± 48 µg/ml
ANE 37 ± 13 µg/ml
ANE ≧ 10K 1120 ± 482 µg/ml ANE 30-100K 15 ± 5 µg/ml
Are 205 ± 51 µg/ml
Cis 43 ± 5 µM
IC50: 50% inhibitory concentration; ANE: areca nut extract;
Are: arecoline; Cis: cisplatin.