國立宜蘭大學生物技術研究所 碩士論文
Graduate Institute of Biotechnology National Ilan University
Master Thesis
影響蛹蟲草固態發酵生產蟲草素之研究
Investigating effects on production of cordycepin by Cordyceps
militaris solid-state fermentation
指導教授:鄭永祥 博士 陳淑德 博士
Advisor : Yeong-Hsiang Cheng Ph. D Su-Der Chen Ph. D
研究生:林冠廷 Guan-Ting Lin
中華民國九十九年六月
中文摘要
蛹蟲草(Cordyceps militaris)發酵能產生具有免疫調節、抗氧化、降血糖 等生理活性成分。本研究採用小麥、薏仁和糙米三種穀類作為蛹蟲草固態 發酵基質;以豬腸膜蛋白粉、血漿蛋白粉、魚粉和酵母粉作為氮源,探討 影響蛹蟲草固態發酵生產蟲草素之條件;及評估殺菌溫度和時間對蛹蟲草 菌存活和生理活性物質滯留率的影響。
結果顯示,以小麥作為蛹蟲草固態發酵之基質,魚粉作為氮源進行發 酵,於 22℃環境下培養,在第 35 天可獲得最高量的蟲草素為 11309.4 μg/g。
熱殺菌處理液態基質方面,在 60℃熱處理 480 秒、70℃熱處理 180 秒、80℃
熱處理 150 秒及 90℃熱處理 90 秒後,能有效抑制蛹蟲草菌絲的生長;熱殺 菌處理固態基質方面,小麥基質裡的菌體在 70℃熱處理 600 秒、80℃熱處 理 600 秒及 90℃熱處理 270 秒後,能被殺滅而停止生長,而此加熱殺菌條 件對蟲草素、腺苷滯留率並無明顯影響。
關鍵詞:蛹蟲草、固態發酵、蟲草素、熱殺菌
Abstract
Cordyceps militaris fermentation can produce immunomodulatory,
antioxidant, hypoglycemic and other physical active components. In this study, adlay, rice and wheat as three different media for Cordyceps fermentation, and adding dried porcine soluble, spray-dried plasma protein, fish meal and yeast powder as nitrogen source. Therefore, the production of cordycepin by Cordyceps militaris solid-state fermentation can be investigated. Moreover, the
effect of sterilization temperature and time on survival and physiological activity of Cordyceps militaris material retention were studied. The results showed that solid-state fermentation of wheat as a medium, fish meal as a nitrogen source for Cordyceps militaris 35 days fermentation at 22℃ obtained the highest
concentration of cordycepin (11309.4 μg/g). Heating treatment at 60℃, 70℃, 80℃, 90℃ for 480 sec, 180 sec, 150 sec, and 90 sec in liquid medium, the growth of Cordyceps militaris can be inhibited, respectively. However, heating Cordyceps militaris in wheat medium at 70℃, 80℃, 90℃ required 600 secs,
600 sec, and 270 sec to inhibit the growth of Cordyceps militaris, respectively.
The retentions of cordycepin and adenosine did not significantly affect during these heating conditions.
Key word: Cordyceps militaris, solid-state fermentation, cordycepin, heat
致謝
首先誠摯的感謝指導教授鄭永祥博士及陳淑德博士,兩位老師細心的 教導使我得以一窺蛹蟲草發酵領域的深奧,不時的討論並指點我正確的方 向,使我在這些年中獲益匪淺。老師對學問的嚴謹更是我輩學習的典範。
本論文的完成另外亦得感謝馬階護專的張讚昌博士大力協助,及生技 所賴裕順老師的支持。因為有你們的體諒及幫忙,使得本論文能夠更完整 而嚴謹。
三年裡的日子,實驗室裡共同的生活點滴,學術上的討論、言不及義 的閒扯、趕作業的革命情感…,感謝眾位學長姐、同學、學弟妹的共同砥 礪,你(妳)們的陪伴讓我的研究生活變得絢麗多彩。
感謝達煒學長、繞哥學長、阿吉學長、易儒學長、以琁學姊和秀芸學 姊們,不厭其煩的指正我研究中的缺失,且總能在我迷惘時為我解惑,也 感謝首成、堯彰、玉婷、靖涵同學的幫忙,幫助我順利走過這三年。實驗 室的暐摯、蝗蟲、阿彬、小潘學弟、宛儒、小雯、欣穎學妹們,當然也不 能忘記,你(妳)們的幫忙及搞笑我銘感在心。
家人在背後的默默支持更是我前進的動力,沒有你們的體諒、包容,
讓這三年的生活有很不一樣的光景。
最後,謹以此文獻給我摯愛的雙親。
總目錄 書名頁
碩士學位論文授權書
碩士學位論文指導教授推薦函 碩士學位論文口詴委員審定書
中文摘要...I 英文摘要...II 致謝…...III 總目錄...IV 表目錄...VIII 圖目錄...IX 附錄...XI
壹、文獻回顧...1
1、前言...1
2、蛹蟲草(Cordyceps militaris)簡介...2
3、蛹蟲草之生理活性成份...7
3.1、蛹蟲草化學成份...7
3.2、蛹蟲草胺基酸組成...8
3.3、蛹蟲草機能性成份...9
4、蛹蟲草之生理功能...14
5、發酵技術...24
6、液態發酵...29
7、影響液態發酵培養之因素...29
7.1、碳源...29
7.2、氮源...30
7.3、碳氮比...30
7.4、微量元素...31
7.5、pH與溫度...31
7.6、接種量與培養基容量...32
8、固態發酵...32
9、影響固態發酵培養之因素...35
9.1、基質特性...35
9.2、基質水分含量、濕度...35
9.3、溫度... 35
10、熱殺菌與非熱殺菌特性與方法...36
10.1、食品的熱殺菌...36
10.2、食品的非熱殺菌...38
貳、研究目的...41
參、研究架構...42
肆、材料與方法...43
1、實驗菌種...43
2、預活化蛹蟲草菌液...43
3、不同活化天數對菌絲體的影響...43
4、腺苷及蟲草素的分析...44
4.1、HPLC分析條件...44
5、菌絲體(Biomass)含量的測定...44
5.1、純菌絲收集……...45
5.2、麥角固醇之測定...45
5.3、菌絲體標準曲線...45
5.4、固態發酵產物菌絲含量之測定...46
6、蛹蟲草固態發酵...46
6.1、不同穀物發酵培養對腺苷和蟲草素的影響...47
6.2、不同氮源發酵培養對腺苷和蟲草素的影響...47
7、熱殺菌詴驗...47
7.1、升溫、降溫曲線...47
7.2、蛹蟲草液態培養液之殺菌詴驗...48
7.3、蛹蟲草固態基質之殺菌詴驗...48
7.4、蟲草素穩定性詴驗...48
7.5、腺苷熱穩定性詴驗...49
8、統計分析...49
伍、結果與討論...50
1、預活化搖瓶培養時間與菌絲體生長之影響...50
2、菌絲體生質量之變化...50
3、固態發酵期間蟲草素和腺苷含量之變化...50
4、不同穀物對菌絲體、蟲草素及腺苷生成之影響...52
5、不同氮源對菌絲體、蟲草素及腺苷生成之影響...54
6、熱殺菌...57
陸、結論...60
柒、參考文獻...85
表目錄
表 1.1、冬蟲夏草與蛹蟲草在外觀和特性方面的比較...3
表 1.2、冬蟲夏草與蛹蟲草三大營養素的含量比較...8
表 1.3、冬蟲夏草與蛹蟲草生理功能的比較...16
表 1.4、固態發酵與深層液態發酵之比較...34
表 5.1 、 不 同 溫 度 (60~90℃) 加 熱 對 蛹 蟲 草 菌 液 之 菌 絲 存 活 的 影 響...61
表 5.2、不同溫度(60~90℃)加熱對小麥培養基中蛹蟲草之菌絲存活的影 響...62
表 5.3、不同溫度加熱處理蟲草素之滯留情形………….……….…………63
表 5.4、不同溫度加熱處理腺苷之滯留情形………….……….………64
圖目錄
圖 1.1、蟲草屬之分類…...4
圖 1.2、蛹蟲草生活史和蛹蟲草菌絲體由無性和有性循環兩方面進行生 殖...6
圖 1.3、蟲草素之化學結構式...9
圖 1.4、甘露醇(D-mannitol)化學結構...10
圖 1.5、麥角固醇(ergosterol)化學結構...11
圖 1.6、蟲草屬之核苷化學結構式...13
圖 1.7、發酵工業的流程圖...28
圖 3、實驗設計流程圖...42
圖 5.1、PDB 搖瓶培養 0~10 天之蛹蟲草菌絲生長曲線...65
圖 5.2、蛹蟲草固態發酵之菌絲生長曲線...66
圖 5.3、以小麥含 5%魚粉為固態基質進行發酵之蟲草素產量變化…...67
圖 5.4、以小麥含 5%魚粉為固態基質進行發酵之腺苷產量變化...68
圖 5.5、不同穀物基質(薏仁、米、小麥)含 5%魚粉經蛹蟲草固態發酵 35 天 之菌絲體含量...69
圖 5.6、不同穀物基質(薏仁、米、小麥)含 5%魚粉經蛹蟲草固態發酵 35 天 之蟲草素含量...70 圖 5.7、不同穀物基質(薏仁、米、小麥)含 5%魚粉經蛹蟲草固態發酵 35 天
之腺苷含量...71
圖 5.8、不同氮源(SDPP、酵母粉、DPS、魚粉)經蛹蟲草固態發酵 35 天之 菌絲體含量...72
圖 5.9、不同氮源(SDPP、酵母粉、DPS、魚粉)經蛹蟲草固態發酵 35 天之 蟲草素含量...73
圖 5.10、不同氮源(SDPP、酵母粉、DPS、魚粉) 經蛹蟲草固態發酵 35 天 之腺苷含量...74
圖 5.11、不同溫度下 PDB 之升溫曲線………...…....75
圖 5.12、不同溫度下小麥固態基質之升、降溫曲線...76
圖 5.13、60~90℃液態殺菌實驗,培養七天之生長情形...77
圖 5.14、60~90℃固態殺菌實驗,培養七天之生長情形...78
圖 5.15、小麥固態發酵基質於滅菌釜中之升溫情形...79
附錄
附圖 A1、麥角固醇標準曲線...80
附圖 A2、菌絲體標準曲線...81
附圖 A3、蛹蟲草菌絲體和麥角固醇濃度相關曲線...82
附圖 A4、蟲草素標準曲線...83
附圖 A5、腺苷標準曲線...84
壹、文獻回顧 1、前言
蛹蟲草為一種昆蟲寄生真菌,主要分布在中國大陸的高山地區,因採 集不易數量稀少,所以價格較為昂貴。蛹蟲草具有各種生物活性成分,包 括多醣、腺苷、蟲草素和甘露糖醇等(Holliday & Cleaver, 2005; Russell &
Paterson, 2008)。近幾年的研究報告證實冬蟲夏草具有抗腫瘤(孫,2006;
Chen et al., 2006; Lin & Chiang, 2008)、調節免疫力(馬等,1995; Kim et al., 2006; Koh et al., 2002; Kuo et al., 1994; Kuo et al., 2007; Won & Park, 2005;
Wu et al., 2006)、降血脂(Koh et al., 2003)、降血糖(黃,2004; Li et al., 2006b;
Kiho et al., 1993)、抗氧化(呂,2007; 莊等,2003; 楊,2001; Yamaguchi et al., 2000; Yu et al., 2006)、增加睪固酮生成(方,2005; Hsu et al., 2003a)、抗
病毒等活性(Holliday et al., 2004)。此外,人工培養的蟲草經科學的分析與 實驗證明,其生理活性成分與天然的蟲草相似 (Li et al., 2006a),人工北冬 蟲夏草二次代謝產物(secondary metabolite)蟲草素(cordycepin)是野生蟲草 的 35 倍,腺嘌呤是野生蟲草的 40 倍,核苷類總含量是野生冬蟲夏草的 6.25 倍(張等,2003);表示天然和人工培養之蟲草子實體或菌絲體發酵液,皆 具有生理活性及功效。
液態發酵的方式雖然能在短時間內獲得大量的菌絲體,但須考量到設 備之成本及空間、下游階段固液分離、廢液處理、濃縮與乾燥的能源成本
等問題。固態發酵可直接使用低水分含量或低水活性的天然顆粒狀穀類,
如:米、小麥、大麥、玉米、大豆或農業廢棄物作為基質(Couto & Sanroman, 2006)。故固態發酵與液態深層發酵相比較,固態發酵培養具有高產率、低 成本的培養基質、減少能源與成本、技術較為簡單、下游分離成本低等優 點(Smits et al, 1996; Castilho et al., 2000; Durand, 2003; Pandey, 2003;
Holker et al., 2004; Philippoussis et al., 2007; Prakash et al., 2008)。
姚(2005)分別將 14 公克米飯、大薏仁、小薏仁、燕麥和喬麥置於培養 皿(petri dish)中,水分含量為 50%,並接入 5 毫升活化菌液於 25℃下進行 北冬蟲夏草固態發酵,第 15 天時米飯基質有最高菌絲體生成量。楊(2005) 則用 14 公克大薏仁、小薏仁、燕麥和喬麥添並加 14 毫升蒸餾水培養於培 養皿,培養溫度為 20℃、接菌量 10%進行冬蟲夏草固態發酵 14 天,菌絲 體生成量為燕麥>小薏仁>大麥>大薏仁>喬麥,多醣生成量以大麥最 多。故穀類可作為蟲草固態發酵的良好培養基質,本研究將以 200 公克穀 類基質培養於 800 mL 玻璃罐中進行蛹蟲草固態發酵。
2、蛹蟲草簡介
蛹蟲草(Cordyceps militaris)別名北冬蟲夏草,又稱蛹草、北方蟲草或北 蟲草,是子囊菌亞門(Ascomycotina)、麥角菌科(Clavicipitaceae)、蟲草屬真 菌(Cordyceps),為蟲生菌的一種。寄主範圍廣泛,主要為鱗翅目的舟蛾科、
天蛾科、大蠶蛾科、蠶蛾科、刺蛾科、尺蛾科、枯葉蛾科;鞘翅目的金龜
科、花金龜科;雙翅目的大蚊科;適合生活的環境,要求土壤表層腐植質 豐富,土壤含水量在15%左右,呈中性或稍偏酸性的山地黃壤土。生長於土 層厚度約在15~20公分,質地疏鬆,少砂礫石區。林間氣候溫度在20~26 ℃,
相對濕度在85%以上,是蛹蟲草的最適合生長條件。表1.1為冬蟲夏草與蛹 蟲草在外觀和特性方面的比較。
表1.1、冬蟲夏草與蛹蟲草在外觀和特性方面的比較(孫和張,2002)。
Table 1.1. Comparison characterstics of C. sinensis and C. militaris.
名稱
子實體 顏色
子座形狀 寄主情況
對生長環境的要 求
冬蟲夏草 黑褐色 圓柱狀
專一昆蟲的帅 蟲上,形態單一
嚴格,海拔以 3000m 以上
蛹蟲草 橙黃色 橢圓狀
多種昆蟲的蛹 上,形態多樣
不嚴格,中山區或 淺山區
蟲草屬(Cordyceps spp.)為廣泛分布的昆蟲寄生型真菌之一(林,2003),
目 前 已 被 發 表 的 種 類 超 過 400 種 , 如 : 中 國 蟲 草 (C. siensis) 、 蟬 花 (C.
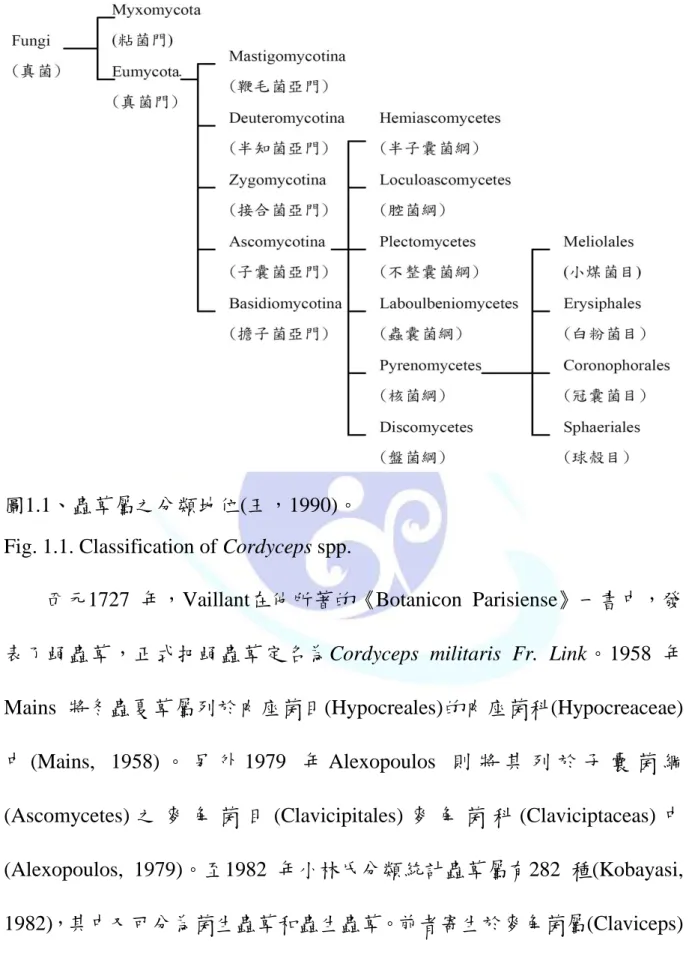
sobolifera)、蛹蟲草(C. militaris)、亞香棒蟲草(C. hawkesii) (王,1995)。圖1.1 為蟲草屬在真菌門之分類地位。
圖1.1、蟲草屬之分類地位(王,1990)。
Fig. 1.1. Classification of Cordyceps spp.
西元1727 年,Vaillant在他所著的《Botanicon Parisiense》一書中,發 表了蛹蟲草,正式把蛹蟲草定名為Cordyceps militaris Fr. Link。1958 年 Mains 將冬蟲夏草屬列於肉座菌目(Hypocreales)的肉座菌科(Hypocreaceae) 中 (Mains, 1958) 。 另 外 1979 年 Alexopoulos 則 將 其 列 於 子 囊 菌 綱 (Ascomycetes) 之 麥 角 菌 目 (Clavicipitales) 麥 角 菌 科 (Claviciptaceas) 中 (Alexopoulos, 1979)。至1982 年小林氏分類統計蟲草屬有282 種(Kobayasi, 1982),其中又可分為菌生蟲草和蟲生蟲草。前者寄生於麥角菌屬(Claviceps) 及大團囊屬(Elaphomyces)的子實體上,後者則寄生於昆蟲及蜘蛛體上。蟲
生 蟲 草 的 寄 主 包 括 : 雙 翅 目 (Diptera) 、 膜 翅 目 (Hymenoptera) 、 鞘 翅 目 (Coleoptera)、鱗翅目(Lepidoptera)、半翅目(Hemiptera)、異翅目(Isoptera)、
直翅目(Orthoptera)、等翅目(Homoptera)等昆蟲綱之帅蟲、蛹與成蟲及蜘蛛 綱(Arachnida)等(Mains, 1958; McEwen, 1963; 徐和曾,1992)。
McEwen(1963)認為蟲草屬的孢子在適當環境,會形成單一或數個發芽 管(germtube),在潮濕陰涼的環境中,其孢子與寄生之體表接觸,會分泌一 些幾丁酵素(chitinase)將寄主體表溶解,因體壁主要是由幾丁質構成,此過 程即是孢子萌芽(germination),而孢子萌芽之前必須打破其休眠期,其機制 目前仍不清楚。一旦孢子在適當環境下萌芽,會形成單一或數個發芽管,
經由寄生之氣孔、節間膜或溶解之體壁直接進入蟲體內而開始菌絲之繁 殖。寄主組織細胞之養分被消耗後,菌體充滿整個寄主體腔。此種內生菌 絲在適當環境下會長出子實體或稱子座(stroma),即所謂的出菇(McEwen, 1963)。其形狀因種類之不同而有圓棍狀、球棒狀、樹枝狀、蛇尾狀或其他 形狀。其顏色有紅、橙、黃、紫、茶、綠、黑、白、褐、棕、灰等多種。
子實體基本上是由緊密成束帄行或交織的菌絲所組成。通常於其頂端、上 端或側邊著生孢子果或稱子囊殼(perithecium),其著生之相對位置為種間分 類之依據。其著生方式也可分為埋生形、半裸形、斜裸形、裸生密佈形、
裸生著衣形。子囊腔之形狀為球形、卵形或橢圓形,其內之子囊(ascus)呈圓 柱狀或長管狀。在子囊中有八條絲狀孢子,即所謂之子囊孢子(ascospore),
彼此帄行排列,有些有隔膜,而將孢子分段成次孢子,也是種間之分類依 據之一(徐和曾,1992)。
菌體生活史如圖1.2 所示,由體腔菌絲(somatic hypha)分別行不同的兩 種生活史世代循環。ㄧ為有性世代循環(sexual cycle),另一為無性世代循環 (asexual cycle)。有性世代循環包括子座(stromata)、子囊(ascus)、子囊孢子 (ascospore)及子囊孢子萌芽(germination)等階段。而無性世代循環包括分生 孢子柄(conidiaphore)、分生孢子(conidia)及分生孢子萌芽(germination)等階 段(黃,2004)。
圖1.2、蛹蟲草生活史和蛹蟲草菌絲體由無性和有性循環兩方面進行生殖 (黃,2004)。
Fig. 1.2. Cordyceps militaris life cycle and there are two aspects generated from somatic hypha as follow: 1) asexual cycle, 2) sexual cycle.(ARS collection of entomopathogenic fungi; ARSEF).
3、蛹蟲草之生理活性成份
蛹蟲草是由蛹蟲草菌感染昆蟲帅蟲或蛹體所形成之藥材,與冬蟲夏草 同屬麥角菌科蟲草屬,也具有相似的生理活性。近年來研究結果顯示,蛹 蟲草為冬蟲夏草重要替代品之一,兩者所含成份十分相似,蛹蟲草的主要 有效成份為:蟲草素、蟲草酸、固醇類、核苷類、蟲草多醣等。藥理學研 究顯示,這些有效成分在對人體醫療、保健等方面發揮著重要作用(柴等,
2003)。
3.1、蛹蟲草化學成份
蛹蟲草藥材一般成份以碳水化合物及粗蛋白為主,總醣、粗脂肪、粗 蛋白質約各佔 14.99%、7.22%、68.29%(表 1.2)。其中粗蛋白中胺基酸的含 量約佔 41%,而灰份約佔 7.36%、甘露醇約佔 4.71%、腺嘌呤約佔 0.054%、
腺苷約佔 0.037%、重金屬含量為 30~40 mg/kg,並有 24 種以上的無機元素,
其中含人體必需微量元素 Fe、Zn、Mn、Cu、Sr、Cr、Ni、Co、S、Sn(孫 和張,2002)。
表 1.2、冬蟲夏草與蛹蟲草三大營養素的含量比較(孫和張,2002)。
Table 1.2. Comparison nutrient contents from C. sinensis and C. militaris.
營養素 冬蟲夏草 蛹蟲草
總醣(%) 24.44 14.99
粗脂肪(%) 10.56 7.22
粗蛋白質(%) 28.45 68.29
3.2、蛹蟲草胺基酸組成
蛹蟲草中九種人體必需胺基酸總量佔41%,冬蟲夏草佔42%(孫和張,
2002),經分析蛹蟲草粗蛋白之水解產物,為麩胺酸(glutamic acid)、苯丙胺 酸(phenyl-alanine)、組胺酸(histidine)、精胺酸(arginine)、纈胺酸(valine)等十 八種胺基酸混合物。在醫藥方面,芳香族胺基酸主要用於營養劑,精胺酸 有促胰島素生成作用,麩胺酸有健腻作用,天門冬胺酸可鎮咳去痰,是蟲 草治虛勞喘咳功效成分之一(貢等,2005)。
3.3、蛹蟲草機能性成份 蟲草素(Cordycepin)
圖1.3、蟲草素之化學結構式(黃,2009)。
Fig 1.3. The chemical structure of cordycepin.
蟲草素為蟲草屬中特有之生物活性物質,而在蟲草屬中,蛹蟲草所含 的蟲草素遠高於其餘蟲草屬的菌種,例如冬蟲夏草、大團囊蟲草、霍克斯 蟲草等。因此蟲草素被認定為蟲草屬中之標定化合物,且是蟲草治病防病 的活性成分(王等,2003)。
1950年Cunningham於蛹蟲草(C.militaris)之發酵製備物中發現具有成分 3‟-deoxyadenosine之核苷抗生素(Cordycepin),將之稱作蟲草素。蟲草素具 有抗病毒活性,是一種重要的核苷抗代謝物,因其能抑制細胞中mRNA後轉 譯作用,是嘌呤再生合成之迴饋抑制劑(Cunningham, 1950)。
蟲草素可作為一種細胞毒性詴劑,和腺苷的結構類似物,能抑制枯草
桿菌(Bacillus spp.)、鳥結核桿菌(Mycobacterium avium)和艾氏腹水癌細胞 (Herlich ascites tumor cells),且蟲草素對RNA和DNA合成有抑制作用(柴 等,2003)。
甘露醇(Mannitol)
圖1.4、甘露醇 (D-mannitol) 化學結構(黃,2004)。
Fig. 1.4. The chemical structure of mannitol.
甘露醇(C6H14O6)的含量在蛹蟲草佔4.71%、在冬蟲夏草中佔6.49%(孫和 張,2002),甘露醇具有抗自由基、抗氧化的作用,是醫藥工業的重要原料,
也是藥品中常用的輔藥,臨床上常用的滲透性利尿藥就含有該成份。滲透 性利尿藥主要用於治療腻水腫及青光眼,是降低顱內壓的安全有效首選藥 品,其在治療腻血栓、腻栓圔、血管痙攣、腻溢血、排除毒素、腎功能衰 竭、促進新陳代謝等方面具有療效。20世紀之後,人們逐漸認識甘露醇的 一些特殊生理功能,如食用後不會引起血糖水帄波動,且不會造成牙齒齲 變及低熱值等特性,因此以甘露醇作為甜味劑和食品添加劑的用量迅速增
加(柴等,2003)。
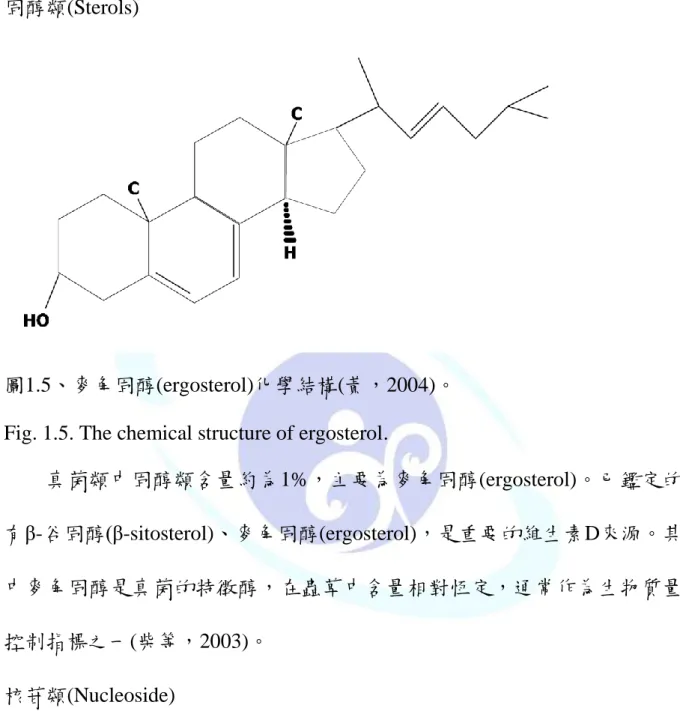
固醇類(Sterols)
圖1.5、麥角固醇(ergosterol)化學結構(黃,2004)。
Fig. 1.5. The chemical structure of ergosterol.
真菌類中固醇類含量約為1%,主要為麥角固醇(ergosterol)。已鑑定的 有β-谷固醇(β-sitosterol)、麥角固醇(ergosterol),是重要的維生素D來源。其 中麥角固醇是真菌的特徵醇,在蟲草中含量相對恆定,通常作為生物質量 控制指標之一(柴等,2003)。
核苷類(Nucleoside)
核 苷 類 是 蟲 草 最 主 要 的 活 性 物 質 之 一 , 包 含 蟲 草 素 在 內 的 腺 苷 (adenosine)、脲嘧啶(uracil)、腺嘌呤(adenine)、次黃嘌呤(hypoxanthine)等物 質(圖1.6)。腺苷為一種抗心律不整藥物,其作用機制為降低心臟傳導速度,
維持細胞膜的完整性並降低冠狀動脈阻力,在心肌缺氧時對心肌有一定的
保護作用,且可有效而選擇性的降低動脈壓力(柴等,2003)。
Adenosine Adenine
Cytidine Guanosine
Hypoxanthine Inosine
Thymidine Uridine
圖1.6、蟲草屬之核苷化學結構式(Thomas & Steen, 1999)。
Fig. 1.6. The chemical structures of nucleosine from Cordyceps spp.
蟲草多醣體(Cordycepic polysaccharide)
多醣類物質是蛹蟲草的重要活性物質之一,近來研究發現多醣對淋巴 系統及巨噬細胞有明顯促進活化作用,能促使淋巴細胞的轉化,是一種非 特選性免疫增強及調節劑,可增強免疫活性細胞,特別是淋巴細胞淋巴因 子,單核巨噬細胞系統和NK細胞等,從而攻擊靶細胞、發揮抗腫瘤作用,
同時具有抗衰老、抗氧化、抗病毒、抗凝血、降血糖、降血脂等作用。蟲 草多醣對體細胞有免疫調節作用,能改善肝功能,用於治療肝炎和恢復肝 損傷(柴等,2003; 貢等,2005)。
超氧化物歧化酶(SOD)
超氧化物歧化酶是一種生物活性蛋白,具有清除人體代謝中產生的超 氧陰離子自由基的能力,在維護有機體活性氧代謝的帄衡中有重要作用。
在預防各種疾病及抗衰老防禦物質中,超氧化物歧化酶是重要的關鍵酵 素,也是目前為止發現唯一以自由基為基質的酵素。近年來,超氧化物歧 化酶得到廣泛的研究,其成果已在醫藥、食品、化妝品、生物製藥等領域 得到廣泛應用(柴等,2003; 貢等,2005)。
4、蛹蟲草之生理功能
西元1756年趙學敏所著的本草綱目拾遺中補述:『出四川江油縣化林 坪、夏為草、冬為蟲,長三寸許,下跌六足,脰以上絕類蠶,羌俗採為上 藥,功與人參同。…夏草冬蟲,乃感陰陽二氣而生,夏至-陰生,故靜而為
草;冬至-陽生,故動而為蟲…』(許,1984)。
近年來的研究結果顯示,蛹蟲草是冬蟲夏草的重要替代品之一(王等,
2003),蛹蟲草最早記載於《新華本草綱要》一書,其功效『味甘、性帄、
有益肺腎、補精髓、止血化痰的功能』,《中華藥海》記載『性味歸經,
入肺腎二經,功能主治健忘不寐、腰膝酸軟、久咳虛喘、勞咳痰血者』,
尚有在降血脂,防治動脈硬化,保護心、腻組織,鎮靜催眠,增強巨噬細 胞活性,抗癌、抗炎、抗菌、抗缺氧等方面具有良好效果(張和張,2001)。
傳統中藥記載蟲草主要功效及典籍,《本草從新》:『保肺、益腎、
補精髓、止血化痰、已勞。』;《藥性考》:『味甘性溫、必精益氣、專 補命門』;《草綱目拾遺》本:『入肺、腎二經、主治肺陰虛、勞咳血;
腎陽虛、遺精早瀉』,『能治諸虛百損』;《柑園小識》:『以酒浸數枚 痰,治腰膝間痛楚,有益腎之功能』;《中藥大辭典》:『冬蟲夏草,性 帄、氣香、無毒、入腎肺二經。有強壯、益肺腎、補精髓、解毒、止血、
化痰之功效』。李時珍在《本草綱目》中指出,蟬花能主治『小兒天吊掠 痢、夜啼、心悸。』蟬花包括蟬草(Cordyceps sobolifera)和蛹蟲草等多種寄 生於蟬體上的蟲草菌(王,1995)。表1.3為冬蟲夏草與蛹蟲草在藥理效用方 面的比較。
表1.3、冬蟲夏草與蛹蟲草生理功能的比較(孫和張,2002)。
Table 1.3. Comparison biofunction of C. sinensis and C. militaris.
名稱 冬蟲夏草 蛹蟲草
免疫作用
對抗免疫系統有雙向調 節作用,腎臟移植中可 抑制免疫排斥作用
具有調節作用
抗腫瘤 有 有
激素作用 無 有雄性激素作用
抗纖維化作用 有 無
抗炎作用 有廣效性的抗菌作用 有抑菌作用
對腎衰的治療作用 能改善腎功能,減輕尿
毒症 無
延緩衰老作用 有 無
抗誘變、抗輻射作用 有 有
造血功能
蟲草結晶物能誘發紅血球細胞系集落形成細胞(colony forming unit-E) 由G0期進入S期(Li et al., 1993),進而刺激這些紅血球前驅細胞的增殖(Xu et al., 1989)。冬蟲夏草萃取物,能在輻射危害初期排除氫氧自由基的產生,與
增強Cbfa(C-Module DNA-Binding Factor)之表現,能刺激造血幹細胞的分 化,減低血球細胞受輻射所造成的缺失(Nakamura et al., 2003; 宋等,2007;
Lin et al., 2007)。
血管擴張
由電磁流量計與恆速灌流之研究發現,蟲草菌絲酒精萃取物有明顯增
加冠狀動脈血流量、降低冠脈、腻及外周血管阻力、具降血壓作用、對血 管帄滑肌之鬆弛作用(馮等,1987)。冬蟲夏草具血管擴張之活性(Chiou et al., 2000)。蟲草中之甘露醇具有使心臟及腻血管擴張的功能,有助於改善動脈 硬化之症狀(柴等,2003; McKenna et al., 2003)。
抗心肌細胞缺氧並給予氧損傷及心臟保護作用
萃取液能降低正常心肌細胞內Ca2+的濃度,減輕缺氧再給氧時細胞內 Ca2+超載造成的損傷現象,並減輕胞內脂質過氧化作用(俞等,1998)。腺苷 為一種抗心律不整藥物,其作用機制為降低心臟傳導速度,維持細胞膜的 完整性並能降低冠狀動脈阻力,在心肌缺血缺氧時對心肌有一定的保護作 用,而且可有效而選擇性的降低動脈壓(Berne, 1980; Toda et al., 1982; Chen, 1995; 柴等,2003; McKenna et al., 2003)。蛹蟲草具有抗心絞痛、保護心臟 的活性(Pelleg & Porter, 1990; Won & Park, 2005; Li et al., 2006a)。蟲草素可 透過調控重組血管壁,抑制COX-2表現以及阻止大鼠主動脈帄滑肌細胞的 增生,對於治療心臟內血管再阻圔(restenosis),為具潛力之探討途徑(Chang et al., 2008)。
降低血壓作用和血脂作用
於家兔靜脈注射蟲草菌乙醇萃取物(0.3 g/kg),有短暫的降血壓作用。
冬蟲夏草亦具降低血壓之活性(Chiou et al., 2000),對以STZ(Streptozotocin) 與菸鹼胺誘導的糖尿病大鼠,冬蟲夏草的子實體具有抗高血壓的活性(Guo
& Zhang, 1995; Kiho et al.,1996; Hsu & Lo, 2002; Zhao et al., 2002; Lo et al., 2004; Li et al., 2004; 鄧等,2005; Li et al., 2006b; Shi et al., 2009)。
蛹蟲草中的多醣galactomannan結構,具有降血脂功能(Kiho et al., 1996;
Li et al., 2006a; Wasser, 2002; Koh et al., 2003; 吳等,2008; Ko et al., 2010)。
降低血糖作用
Kiho等人(1993)指出C.sinensis 菌絲中萃取出的多醣體,能有效降低糖 尿病老鼠血液中血糖。萃取物中主要為半乳甘露醣之結構,具有降低血糖 之 功 能 (Kiho et al.,1996; Ng & Wang, 2005; 宋 等 , 2007) 。 對 以 STZ(Streptozotocin)與菸鹼胺誘導的糖尿病大鼠,冬蟲夏草的子實體能緩解 病症所致的體重下降、劇渴和血糖過高(Lo et al., 2001; Lo et al., 2004; Li et al., 2004; 鄧等,2005; Li et al., 2006ab; Shi et al., 2009)。
抗癌、腫瘤作用
蛹蟲草對人體黑色素瘤B12細胞、人體白血病HL-60細胞、人體白血病 K562細胞具有較好的抗腫瘤效果,而且蛹蟲草的作用優於冬蟲夏草(柴等,
2003; 王等,2004; 陳等,2005a)。由同為蟲草屬的Cordyceps cicadae品種 中抽取之多醣類Galactomannan對於植入老鼠體內之Sarcoma 180腫瘤,具有 抑制其生長的效果(Kai et al.,1983; Shin et al., 2003, 王等,2007a)。冬蟲夏 草可抑制細胞分裂,阻止腫瘤細胞轉移(Ukai et al., 1983; Ohmori et al., 1986;
Xu et al., 1988; Zhu et al., 1998; Holliday et al., 2004; Ng & Wang, 2005)、抗艾 氏腹水癌(Ohmori et al., 1989)、蟲草多醣具抗腫瘤活性(Wasser, 2002)、抑制
肝腫瘤轉移(Nakamura et al., 2003)、抑制H22肝癌活性(孫等,2007;何等,
2008)、透過抑制端粒酶活性,抑制人類肺癌細胞A549(Park et al., 2009),主 要所含成份有蟲草素(Hybell et al., 1985)、大量D-mannitol、多醣體物質及半 乳甘露糖(galactomannan)(Kiho et al., 1996)。
蟲草素是一種重要的核苷代謝物,因其能抑制細胞中mRNA後轉譯作 用,是嘌呤再生合成之迴饋抑制劑,具有抗病毒活性,人類腫瘤細胞之抗 增殖效應與蟲草素之吸收有直接關係(Holliday et al., 2004)。
抗氧化作用
蟲草中含有SOD,而SOD透過對O2起歧化作用合成H2O2,保護生物膜 中不飽和脂肪酸被氧化為過氧化脂質(LOP),降低LOP被溶酶體吞噬後形成 脂褐質在組織中的積累,從而延緩衰老,在此過程中,SOD中的Cu2+可被歧 化反應的產物H2O2還原失去活性,而過氧化氫酶(CAT)可清除產生的H2O2, 對SOD起保護作用;同時O2可使CAT失去活性,SOD又可以對CAT起保護 作用,可明顯提高老年大鼠體內的SOD含量,並增強CAT活性,降低老年
大鼠體內的LOP含量及自由基水帄,從而保護細胞免受O2損害,延緩器官
的衰老(王和韓,2002; 貢等,2005)。蛹蟲草具有抗氧化的活性(呂,2007; 陳 等,2008; Dong & Yao, 2008; 李等,2009; Leung et al., 2009; Yan et al., 2009),且在子實體與毛蟲寄主中具相似之效力(Li et al., 2002)。冬蟲夏草可 抑制活性氧族促使的過氧化氫或Fas-receptor ligation誘導的細胞凋亡,證明
具有抗氧化及抗細胞凋亡之活性(Buenz et al., 2004; Chor et al., 2005; Wang et al., 2005)。蟲草多醣能保護大鼠腎上腺髓質嗜鉻細胞瘤PC 12細胞,免除
過氧化氫造成之損傷;並顯著減少丙二醛,降低變異的穀胱甘肽過氧化酶 與超氧化物歧化酶活性,顯示能使免於自由基誘導之神經細胞毒性,及對 於中樞神經系統之退化性疾病具保護作用(Li et al., 2003; McKenna et al., 2003; Schmidt et al., 2003; Ng & Wang, 2005)。
鎮靜催眠作用
蛹蟲草能明顯減少小鼠自主活動,對抗戊四氮誘發小鼠驚厥和一定程 度協同戊巴比妥鈉誘發小鼠睡眠,顯示蛹蟲草具有一定的鎮靜催眠作用(柴 等,2003)。天然蟲草具有鎮靜的功能,主要成份為谷胺酸、色胺酸及酪胺 酸三種胺基酸,為中樞抑制神經遞質及與動物失眠有密切關係,產生藥理 效應(張等,1991; 張等,1998; 曹等,2005; Won & Park, 2005)。
抗菌作用
蟲草所含的甘露醇能抑制葡萄球菌、鏈球菌的生長。由Cordyceps ophiocordin 二次代謝物分離之ophiocordin 具有抗黴菌效果,是一種典型之
抗生素(Kneifel et al., 1977)。蟲草素還表現出極強的抗真菌(Chu et al., 2006;
Ng & Wang, 2005; Kim et al., 2006; Sugar & McCaffrey, 1998),抗金黃色葡萄 球菌、蠟狀芽孢桿菌、變形桿菌、枯草芽孢桿菌、綠色木霉菌和黃曲霉菌(張 等,1998; 秦等,2006),梭狀芽孢桿菌等桿菌屬(Ahn et al., 2000),抗HIV- I
型病毒和選擇性抑制梭菌屬細菌活性(武等,2008)。
雄性激素作用
蛹蟲草水提取液在劑量5 g/kg 時有促進大鼠糖皮質激素(glucocorticoid) 和性激素分泌的作用,可增加去勢大鼠精囊前列腺重量,具有雄性激素的 作 用 ( 柴 等 , 2003) 。 腺 苷 能 刺 激 腎 上 腺 皮 質 合 成 多 種 皮 質 固 醇 (corticosteroid),其中主要是糖皮質素(glucocorticoid),其他還有雄性素等性 激素(Chen et al., 2008)。冬蟲夏草能促進大鼠的睪固酮之生合成(Guo, 1986;
Zhu et al., 1998; Huang et al., 2001; Hsu et al., 2003ab; Huang et al., 2004)。冬 蟲夏草能透過活化cAMP-protein kinase A訊號路徑,降低P45scc酵素活性,
以保護人類絨毛膜性腺刺激,及萊氏細胞(Leydig cell)之類固醇生成(Bao et al., 1994; Hsu et al., 2003b; Holliday et al., 2004 ; Chen et al., 2005; Ng &
Wang, 2005; 方,2005)。
治療腎臟疾病
鄭等人(1992)在慢性腎功能不全的實驗大鼠研究中發現,蟲草可降低其 血尿素氮及肌酐濃度,降低死亡率,而對慢性腎衰竭患者,其細胞免疫功 能下降、血漿白蛋白及血紅蛋白較正常人低,蟲草可經由提高其細胞免疫 功能來預防續發性感染(鄭等,1992)。冬蟲夏草萃取物被利用在大鼠腎臟的 局部缺血/再灌注,具有對細胞凋亡及發炎基因負向調控之能力(Shahed et al., 2001)。蟲草多醣能夠治療大鼠腎臟部分切除誘發的慢性腎功能衰竭,改善
腎臟功能(陳等,2009b)。冬蟲夏草可藉由抑制癌細胞Bcl-2和Bcl-XL的酪胺 酸磷酸化作用,帄定人類腎間隔細胞的增生,並促進細胞凋亡。對腎臟健 康具保護作用(Guan et al., 1992; Jiang & Gao, 1995; Xu et al., 1995; Yang et al., 2003; 許等,2006; 金和陳,2006; Wojcikowski et al., 2006; 王等,2007b;
尹等,2007ab; Wang et al., 2009)。
免疫調節作用
在植入EL-4淋巴瘤細胞之C57BL/6小鼠,給予蟲草粗抽取物後,可增強 其腹膜巨噬細胞之分化能力,並促進其吞噬能力之回復。蛹蟲草能促使巨 噬細胞增殖、增強吞噬細胞功能、使其內酸性磷酸酶活性增強、促進淋巴 細胞轉化及產生白細胞介素(IL-2)、增強淋巴細胞對IL-2吸收率,對體液性 免疫亦有調節作用(王和晉,1994;馬等,1995; 劉等,2008)。正常人和白 血病患者自然殺手細胞(NK)活性有明顯增強作用,促使急性白血病患者血 中所存在的前NK或未成熟的、功能有缺陷的NK進行合成具細胞毒性之 NK,明顯改善腫瘤患者免疫狀態(劉,1991; Zhang et al., 2004)。冬蟲夏草 能透過免疫調控,調節支氣管肺泡灌洗液細胞的功效(Kuo et al., 2001)。蟲 草素對IL-10具顯著的正向調控作用,明顯提升IL-10 mRNA的表現量,並有 抑制單核白血球增多症的功能(Koc et al.,1996; Zhou et al., 2002)。蟲草萃取 物能增進IL-2產生,提升Con-A誘導的脾臟細胞增生及心肌粒線體ATP產 生,強化免疫功能(Siu et al., 2004)。蟬花萃取物也具免疫調節功能(Weng et
al., 2002)。蟲草素能抑制NF-κB活化和抑制Akt、p38的磷酸化,達到對iNOS
與COX2基因表現的負向調控,進而抑制NO的生成;因此蟲草素在發炎相 關的病症上,也具相當潛力之療效(Kim et al., 2003; McKenna et al., 2003;
Chiang et al., 2005; 王和張,2006; Kim et al., 2006; Kim et al., 2007; Kim et al., 2008; Li et al., 2009; Han et al., 2010)。蛹蟲草具有抗發炎及抑制NO生成
等活性(Won & Park, 2005)。口服蛹蟲草萃取物的小鼠,抽取其腻及肝藏的 RNA發現,能顯著誘導IL-18的轉錄,並且活化IFN-γ增生。蛹蟲草另還具有 調節哮喘的肺氣管發炎之能力(Manfreda et al., 1989; Han, 1995; Qiuo & Ma, 1993; Zheng et al., 1985; Qu et al., 1995; Zheng & Deng, 1995; Halpern, 1999;
Lee et al., 2006; Li et al., 2006a; Hsu et al., 2008)。蟲草多醣能誘導T淋巴細胞 增生,分泌IL-2,IL-6和IL-8,活化胞外訊號調節激酶(ERK),以及促進巨 噬細胞的吞噬能力及激發磷酸酶活性,證實蟲草多醣在觸發免疫上伴演重 要的角色(Schepetkin & Quinn, 2006; 吳和王,2007; 劉等,2007; 陳等,2007;
Masuda et al., 2009; Kawanishi et al., 2009; Jerry et al., 2009)。
保肝作用
蟲草菌絲具有刺激活化肝臟細胞的作用,可藉此改善肝功能、降低GPT (Liu et al., 1986; Yang et al., 1994; 周等,1990; Manabe et al., 1996; Dai et al., 2001; McKenna et al., 2003; 劉等,2004; 方等,2005; 王和張,2006; 張和 沈,2006)。在人類肝星狀細胞中,蟲草能正向調控Fas與Bax,負向調控Bcl-
XL,及Fas配位體和Bcl-2不會有變化;都能顯示蟲草具有抗肝纖維化的潛 力(Chor et al., 2005; 唐等,2006; 金等,2007; 王等,2008; 孫等,2008)。
在四氯化碳誘導肝損傷的SD大鼠詴驗中,蟲草複合物CC-700,能壓抑天門 冬胺酸轉胺酶與丙胺酸轉胺酶之活性,幫助血清膽紅素、血清白蛋白、三 酸甘油脂、總膽固醇、血小板數量和凝血酶原作用時間恢復至正常值,過 氧化氫酵素也會復原到原來水帄,肝膞原組成(纖維化指標)也顯著被抑制下 來,而且在高與低劑量時,都有很好的效果,且都優於控制組silymarin(Ko et al., 2010; 朱等,2009)。而冬蟲夏草也具活化肝臟中巨噬細胞Kupffer cell的
能力(Nakamura et al., 1999)。
抗疲勞
日本擬青霉(Paecilomyces japonia)(為蛹蟲草之無性型)之萃取物,藉由對 脂肪利用的提升,以及延緩血漿乳酸和氨的累積,進而能增強小鼠在強迫 游泳詴驗中的游泳能力(Mizuno, 1999; Zhu & Rippe, 2004; 陳等,2009a;
Holliday et al., 2004; Jung et al., 2004; Ng & Wang, 2005)。有趣的是,在受高 強度的職業運動員與一般常人之比較上,運動員之表現並沒有顯著的提升 (Parcell et al., 2004)。
5、發酵技術
發酵“fermentation”的字是來自“fervere”,即拉丁文的沸騰(boil),此種
沸騰的現象,是存在果汁或麥汁中糖類發生嫌氣代謝所產生的CO2所造成。
但是後來生化學者對“fermentation”則認為是在利用有機化合物時會隨伴產 生能量的意思。而發酵工程學者則認為具有更廣泛的涵義,果汁或麥汁經 由酵母之作用而產生酒精,是以微生物生產之最早發酵工業,因此發酵工 程 學者表 示 fermentation 為 大量培 養微生 物來生 產物 質之 工程 (Hastings, 1971)。
發酵工程的種類
微生物的發酵方法有兩類:深層液態發酵和固態發酵。1945年青黴素 的大規模工業化生產開創了深層液態發酵技術及現代發酵工業,發酵工程 與生化工程也由此應運而生 主要研究對象是純種培養與大規模產業化。
發酵工程在工業上可分成:以菌體為生產目的者、以生產酵素為生產 目的者、以微生物之代謝生產物為生產目的者、使添加之化學物質經由微 生物的化學變化,或修飾後產物的生產目的者,等四種主要的類群(Hastings, 1971)。
微生物之代謝產物
微生物之生長可分成四個階段。當種菌(seed)的菌被添加到培養基中 後,會有一定的期間完全沒有生長,這個期間稱為誘導期(lag phase),這是 菌適應新環境所需要的時間。隨後,菌的生長速度就慢慢加快,很快的就 到達最大生長速度,此時稱為生長期或對數增殖期(log phase),然後菌的生 長就會減緩,為所謂的定常期(stationary phase),再繼續培養時在總菌數中
所佔的生菌數會開始減少,而到達培養上所謂的死滅期(death phase)。在對 數增殖期所生成的生產物則有胺基酸、蛋白質、脂質、碳水化合物等,這 些是菌在生長上所必需的物質。這些物質是代謝的基本物質,把生產這些 一次代謝生產物的時期稱為trophophase(在菌之生長上則相當於對數增殖 期)(Bu'Lock et al., 1965)。
一次代謝生產物的生產量,野生株則只生產供應生產菌本身在生長上 所需之量而已,不會生產過剩,因此在工業上要使生產物更有效的蓄積,
則需利用突變操作來改良菌株或調整培養條件,盡可能的來提高生產物之 產量(Bu'Lock et al., 1965)。
在定常期,一部份之微生物的生產物在trophophase不生成,而在生產與 該菌之本質上的代謝無直接關係之物質,這類物質稱為二次代謝生產物。
生 產 這 些 物 質 的 培 養 時 期 稱 為 idiophase( 在 菌 的 生 長 上 則 相 當 於 定 常 期)(Bu'Lock et al., 1965)。
這些二次代謝生產物之蓄積,對生產菌本身到底具有何種生理意義或 功能則有不同的論說,可能是細胞儲藏物質的一種形式,或微生物在自然 界中生存競爭的一種能力,大多是在微生物生長後期或停止生長後,才開 始合成。很多的二次代謝生產物具有抗菌性、特殊酵素之抑制劑、生長促 進(growth promoter)等機能,顯示各種的藥理活性。因此二次代謝生產物也 成為發酵工業上重要的一部份(Bu'Lock et al., 1965)。
發酵工程之成立
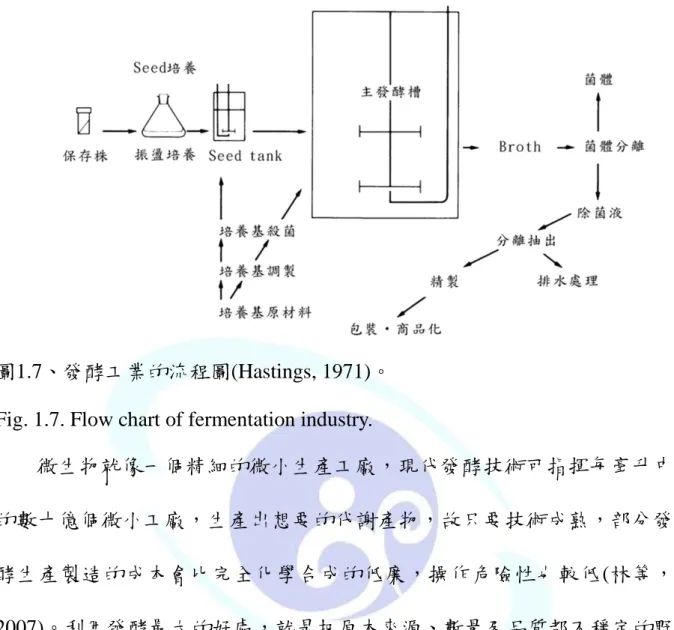
發酵工程是由菌之增殖所用的培養基、製作發酵生產物之培養基等所 有培養基之調製,培養基、發酵槽、其他被用之所有器具的殺菌,給予發 酵足夠量之新鮮種母(seed)的製作,在最適條件下之主發酵槽中之菌的增 殖,生產物之抽出與精製,廢棄物的處理,等六個基本工程所構成(Hastings, 1971)。
這些六個基本的工程之相互關係表示於圖1.7中,但是從研究開發之立 場來看,不僅要注意這些單獨的工程,還要檢討從整體看其流程之效率盡 可能使提高的方法。要確立發酵工程時,首先要分離生產菌,然後加以育 種,使目標生產物,有合乎經濟性的量又能滿足市場要求品質,決定其最 適培養基組成。同時要確立生產物之分離、萃取與純化的流程。另外發酵 工程之研究開發計劃,對使用之微生物的生產能力、生產培養基、萃取純 化工程等之繼續的改善需加以考慮(Hastings, 1971)。
圖1.7、發酵工業的流程圖(Hastings, 1971)。
Fig. 1.7. Flow chart of fermentation industry.
微生物就像一個精細的微小生產工廠,現代發酵技術可指揮每毫升中 的數十億個微小工廠,生產出想要的代謝產物,故只要技術成熟,部分發 酵生產製造的成本會比完全化學合成的低廉,操作危險性也較低(林等,
2007)。利用發酵最大的好處,就是把原本來源、數量及品質都不穩定的野 外採集菇菌類,改成液態或固態發酵,以確保有效成分、縮短時間及放大 產能,也可減少野外採集有助於生態保育(林等,2007)。
大致而言,發酵工程由三部分組成:上游工程包括優良種株的選育,
最適發酵條件(pH、溫度、溶氧和營養組成)的確定,營養物的準備等。發酵 工程主要指在最適發酵條件下,發酵槽中大量培養細胞和生産代謝産物的 技術。在進行任何大規模工業發酵前,必須在實驗室規模的小發酵槽進行 大量的實驗,最後從詴驗資料中再設計出更大生産規模的動力學模型。由
於生物反應的複雜性,在從實驗室到中詴,從中詴到大規模生産過程中會 出現許多流體流動、質傳和熱傳的問題,這就是發酵工程技術放大的課題。
下游工程則主要包含從發酵液中分離和純化産品、乾燥等技術(余,1997)。
6、液態發酵(Submerged fermentation, SmF)
液態發酵培養又稱為深層培養(Submerged fermentation),典型深層液態 培養的水含量高於95%。液態培養的方式能在短時間內獲得大量菌體。許多 學者近年來大多先以少量液態搖瓶發酵的方式,進行最適化的培養條件之 探討,依序放大到中間工廠、最後到工業級量產(田,2001)。液態發酵的優 點包括:(1)菌體生長快速,發酵時間短。(2)可量化生產,不受季節影響。
(3)機能活性成分與野生蟲草相似 (姚,2005;Li et al., 2006a)。(4)較固態發 酵容易散熱及質量傳遞、發酵效率較高。而液態發酵的缺點包括:(1)生產 設備的投資金額比較高。(2)發酵後廢液問題。(3)離心濃縮設備費高。(4)冷 凍乾燥與噴霧乾燥設備費高(林,2008)。
7、影響液態發酵培養之因素 7.1、碳源
碳源為菌體生長主要能量來源之ㄧ,同時也是構成菌絲骨架的主要來 源。Dong & Yao(2005)以單糖之葡萄糖、雙糖之麥芽糖及多醣之糊精作為 碳源,可獲得較佳的菌絲產量。當碳源為糖醇類、蔗糖及麥芽糖可得 4.45~4.26 g/L 菌絲量。蟲草菌於半乳糖培養基發酵培養九天後,殘糖量大
幅驟減,故推估培養基中的碳源已充分被菌體利用轉換,菌絲最高產量為 19.27 g/L (Mao et al., 2005a)。Xiao 等(2006)發現糖蜜與甘油以 2:1 混合做 為碳源時,可得到最高的多醣與菌絲產量。
7.2、氮源
氮源方面,氮是一般為菌體構成胺基酸之重要成分,亦是組成蛋白質 與核酸的主要來源,氮源的種類及濃度會影響菌絲生長形態。氮源是微生 物分解有機物質的重要元素。當分解高碳氮比之基質時,微生物需要額外 供應的有機氮源。充分的氮源供給以致分解過程能達到礦化作用的階段,
這些氮源被利用於微生物體的生長(Girma et al, 2009)。無機氮、胺基酸及 複合有機氮中,冬蟲夏草較能利用複合有機氮中的蛋白腖轉換成菌絲體 (Dong & Yao, 2005)。Park 等(2001)以 1%酵母抽出物為氮源進行發酵,每 公升發酵液可得最高菌絲產量乾重為 17 克;1%玉米粉作為氮源可得胞外 多醣 1.97 g/L。以 6 g/L 花生粕作為碳源可產生最高多醣與菌絲量(Xiao et al., 2006)。
7.3、碳氮比
碳氮比是指總有機碳源與總氮源之比例(C:N),是有機物質提供之指 標。有機碳源對於發酵微生物來說,是主要的能量來源,氮源可以增強利 用這些碳源的效率。基質中含高碳氮比值時,微生物缺乏足夠氮源去分解 碳源,導致發酵過程變緩慢,然而,基質在低碳氮比值時,會加速有機質
的分解,造成氮素的礦化作用,即微生物需求過剩之有機氮源轉化為散佈 空氣中之無機氮源。礦化作用會造成微生物營養生長的停止和起始二次代 謝階段,碳氮源之耗竭,會觸發產生微生物分解纖維素、半纖維素和木質 素等,同樣地,過多的氮源會抑制微生物對木質素之分解(Erwin et al, 1995)。愈高的碳氮比表示基質中愈多的可利用碳源,對菌絲體的生長幫助 也愈大;但碳氮比在超過一臨界值(依菌體和基質有所不同)後,碳源對於 菌體之生長即不再有幫助。這些過量的碳源甚至可能抑制酵素的作用,造 成菌絲生長速率的減慢(Griffin, 1981; Lilly & Barnett, 1951)。碳氮源和其之 間之帄衡關係,對於真菌的生長扮演著重要的角色,因為碳氮比間的交互 作用除影響菌絲生長外,同時也會影響到菌絲體中蛋白質和脂質的含量 (Carlile & Watkinson, 1994)。碳氮比為 12:1 對於冬蟲夏草的菌絲生長有顯 著影響(Dong & Yao, 2005)。Mao 等(2005a)以葡萄糖與蛋白腖作為碳源和 氮源,當碳氮比介於 5:2 可達最高菌絲產量 23.0 g/L。
7.4、微量元素
培養基中所含的微量元素會影響酵素的活性,進而影響菌體生長及代 謝產物分泌的速度。添加 3%酵母抽出物與 3% ZnCl2搖瓶培養 4 天後,均 可大量增加菌絲與多醣產量(Xiao et al., 2006)。
7.5、pH 與溫度
pH 會影響菌體細胞膜上帶電荷的情形,當電荷發生變化,膜的通透性
也會改變,可能影響菌體對於營養物質的吸收與代謝物的分泌速率。在最 適培養溫度範圍,能使菌體生長和代謝加快。在 pH 為 5~6、培養溫度 20℃
冬蟲夏草的菌絲與多醣有最大收量(Park et al., 2001);蛹蟲草最是培養條件 為 pH 5~6、培養溫度 26~28℃(Xiao et al., 2006),故液態培養蟲草菌培養基 在偏酸性 pH 5~6 環境且溫度在 20~28℃環境為較適合。
7.6、接種量與培養基容量
蟲草菌發酵過程是屬於好氧性發酵,發酵過程中的溶氧量對於菌絲生 長有極大影響,溶氧量會隨著菌絲生長而有不足現象,到發酵後期發酵液 會變得較黏稠而影響溶氧量及質傳。接種量 4~6%、裝液量 200 mL 培養基 於 500 mL 錐型瓶可獲得最多菌絲與多醣產量,而較多的培養基與接種量 並不會和菌絲多醣產量有絕對正相關性(Xiao et al., 2006)。
8、固態發酵(Soild-state fermentation, SSF)
固態發酵是指以沒有流動液體的固體作為碳氮源支持物,如傳統的纖 維質廢棄物或是顆粒狀的穀類的原料,再加入適當的水分和營養源後發 酵,發酵期間的酵素來源為:(1)基質本身的酵素,或是(2)來自天然界的微 生物,如:堆肥、高梁酒釀造、乳酪的製造等;以及(3)接種菌種,如菇類 菌種製造、菇類栽培、酵素製造等(Hesseltine, 1972; Aidoo et al., 1982; 王 等,1994; 楊,2002)。固態發酵並沒有大量發酵廢水、通氣、機械攪拌的 高動力耗能,且可節省後段工程中離心濃縮與冷凍乾燥等操作費用。天然
或合成的物質都可作為固態發酵的基質,顆粒狀的穀類如米、小麥、大麥、
玉米、大豆與農業培養的殘餘物,均含有豐富的纖維、澱粉、多醣、蛋白 質及木質素可被菌體作為碳源利用(林,2008; 周,2005)。
固態發酵培養系統無水或接近於無水的發酵過程,與常見的液態發酵 相比具有許多特點。在以固態發酵程序生產二次代謝產物方面,因其培養 特性可產生較高濃度的產物,且培養過程能源消耗少,可應用於高附加價 值之產物生產(黃等,2003; 吳等,2003)。
在農業應用方面,可利用固態發酵進行農業品生產,例如將Agaricus bisporus接種於堆肥和木屑上,栽培洋菇菌種(Wang et al., 1976)。其次為農
業資材之利用,如堆肥製作,可以豬糞和稻草為基質,接種高溫放線菌製 作堆肥(詹等,1999; 楊和陳,2000)。
在工業方面,可進行化學品製造:如酒精之生產或酵素生產,較常見 的有纖維素分解酵素蛋白質水解酵素和澱粉水解酵素等(Bothast & Saha, 1997)。
食品生產方面,固態發酵被廣泛應用於東西方食品之製作,如高梁酒、
醬酒、味噌、乳酪等(Bisping et al., 1993; Wang & Tsao, 1994)。
但固態發酵培養的困難點包括(1)基質床的攪拌非常困難,因此細胞、
養分、溫度、水含量可能會分佈不均勻,使得固態發酵過程控制非常困難。
(2)微生物呼吸或代謝產生熱使得溫度控制非常困難,因為固態基質床的熱
傳導係數非常低,通常強制通氣是控制培養溫度的唯一方法。(3)菌體生長 與其他發酵參數的快速測量非常困難,尚無感測器可供直接測量,通常溫 度是固態發酵控制微生物生長與產物生成的唯一方法,這使得固態發酵連 續操作與自動化非常困難。(4)黴菌或其他菌絲真菌適合生長在低水活性的 基質。(5)由於影響固態發酵高產率因子還是未知數,培養策略完全是靠經 驗與實驗結果(廖,2003;Holker et al., 2004;Sato & Sudo, 1999)。
固態發酵與深層液態發酵之特性比較如表1.4,與深層液態培養相較,
固態發酵具有操作簡單、原料取得容易、產品大多可直接利用等優點,但 基質水分含量較低,不易被所有微生物利用、及培養絛件的控制與偵測不 易等缺點也是急需改進之處(楊,2002)。
表1.4、固態發酵與深層液態發酵之比較 (王等,1994; 楊,2002)。
Table 1.4. Comparison between soild-state fermentation and submerged fermentation.
特性 固態發酵 液態發酵
泡沫 產程控制 投資 廢水 細胞毀壞 產程費時 氧氣供應 熱傳導
微生物與基質 水分
發酵效率
無 較不易 便宜 無
不易發生 長
散怖性 傳導性 靜態 低 低
有 較容易 貴 有
容易發生 短
通氧性 對流性 振盪 高 高
9、影響固態發酵培養之因素 9.1、基質的特性
固態基質顆粒的尺寸大小影響到單位體積反應表面積,進而影響菌絲 的生長、氧氣的供給、二氧化碳的移出與發酵熱的移除。雖然小的基質顆 粒可以明顯提高固態發酵速率,但過小的顆粒則會造成基質的積團,基質 與基質間空隙也變小,對熱傳與質傳也產生不利,故需充分暸解基質的特 性方有助於整個發酵過程(廖,2003)。
9.2、基質水分含量、濕度
水分含量對於固態發酵基質十分重要,典型的基質水分含量約為 30~80%(w/w),當基質水分含量過高會造成固態基質堆聚在一起,使得整個 發酵過程通氣不良,甚至形成缺氧環境,基質最適水分的選擇取決於微生 物的種類與基質本身,低水分固態基質有利於酵母菌與真菌的生長(Yusuf, 1999;廖,2003)。
9.3、溫度
固態發酵過程會產生發酵熱,導致發酵基質溫度升高,發酵熱的移除,
對於熱傳導困難的多孔性固態基質是主要的限制。發酵過程的溫度控制大 多都是藉由蒸發冷卻的方法,使用較乾的空氣、控制空氣溫度提供雖然有 較好的冷卻效果,但會造成基質脫水而乾燥,故常會搭配間歇性的噴灑冷 卻水(Yusuf, 1999; 廖,2003)。
固態發酵的研究包括培養蛹蟲草子實體或菌絲體,如周(2005)利用固態 培養方式以家蠶蛹與白米做為主要培養基,於溫度20℃、相對濕度70%、光 照強度1500~2000 Lux、接菌量10%,固態培養90~120天後,即可獲得蛹蟲 草子實體;郭(2005)利用15 克米置於培養皿中進行蟲草米的製備,於發酵8 天有最高菌絲生成量;且培養溫度以 20℃較25℃和30℃佳,其菌絲在 18℃~22℃下呈較粗壯的白色珊瑚狀,進行冬蟲夏草米固態發酵8天後僅獲 得540 mg/g粗多醣。楊(2005)則利用14 克小薏仁、大薏仁、大麥、燕麥和 喬麥於20℃下發酵14天後,大麥基質方可產生最高的粗多醣含量為321.7 mg/g,大薏仁次之。
10、熱殺菌與非熱殺菌特性與操作
為要保持食品的品質,保證食品的衛生安全,就必須對食品包裝及其 內容物進行殺菌。不同的殺菌方法適用於不同的包裝材料、容器以及不同 的食品生產方式。一般食品殺菌模式可分為熱殺菌和非熱殺菌兩大類(涂 等,2004)。
10.1、食品的熱殺菌
食品的熱殺菌是食品加工與保藏中用於改善食品品質、延長食品貯藏 期的最重要的處理方法之一,其正面作用主要是殺死微生物、鈍化酵素、
改善食品的品質和特性,提高食品中營養成分的可消化性和可利用率;破 壞食品中不符合需要或有害的成分。而熱殺菌的負面作用主要指熱殺菌中
食品的營養和風味,特別是熱敏物質的損失,故對食品的品質和特性有一 定的影響;另外採用熱殺菌的方式消耗能量較大。但對於基質和真菌純種 培養的殺菌條件卻較少出現於論文探討中,而業界為達無菌常過度殺菌,
造成不必要的能源損失。
熱 殺 菌 的 形 式 依 殺 菌 溫 度 的 高 低 , 可 將 熱 殺 菌 分 為 低 溫 殺 菌 (≦100℃)、高溫殺菌(>100℃)、超高溫殺菌(>130℃);按殺菌程度的強弱,
可將熱殺菌分為巴氏殺菌(≦100℃)和商業殺菌(>100℃);按殺菌壓力又可將 熱殺菌分為常壓殺菌(≦100℃)和加壓殺菌(>100℃)。
食品的熱殺菌反應一般遵循一級反應動力學,也就是說熱殺菌反應的 速率與反應物的濃度呈正比關系。這一關係通常被稱為“熱殺菌或熱滅活 的對數狀態”。表示在某一熱處理溫度(足以達到熱滅活或熱殺菌溫度)下,
單位時間內,微生物(食品成分) 被殺滅或被破壞的比例是恆定的。
確定食品熱殺菌條件時,應考慮影響熱殺菌的各種因素。食品的熱殺 菌以殺菌和抑制酵素活性為主要目的,應基於微生物和酶的耐熱性,並根 據實際熱處理時的傳熱情況,確定達到殺菌和抑酶的最佳的熱處理程度。
目前,熱殺菌技術的主要研究動向是熱殺菌條件的最適化和熱殺菌設備。
熱殺菌條件的最適化為協調熱殺菌的溫度-時間(T-t)條件,使熱殺菌的效果 達到期望的作用,而盡量減少不期望的作用。對於熱殺菌設備其最常用的 便是蒸氣殺菌,其次是微波加熱殺菌和電阻加熱殺菌設備(涂等,2004)。
10.2、食品的非熱殺菌
食品的非熱殺菌主要包括物理殺菌和化學殺菌。物理殺菌主要是指: 輻 照殺菌、紫外線殺菌、超高壓殺菌、高壓脈衝電場殺菌、脈衝強光殺菌和 超聲波滅菌等。物理殺菌的主要優點是殺菌效果好,對食品污染小,易於 操作和控制,能更好地保持食品自然風味,但殺菌成本太高。在應用新的 物理殺菌技術時我們應首先考慮三大問題,一是殺菌中是否引起新的污 染;二為是否比傳統方法有明顯的經濟優勢;三為是否能實現規模化生產。
下面將簡要介紹幾種常見的物理殺菌方法:
①超高壓(UHP) 殺菌技術:是指將密封於彈性容器內的食品置於水或 其他液體作為傳壓介質的壓力系統中,經100MPa 以上的壓力處理,使食品 中蛋白質變性、酵素失活,細菌細胞膜受損, 從而達到殺菌、滅酶和改善食 品功能特性等目的。其主要特點是:在常溫或較低溫度下達到殺菌、滅酵 作用,使能耗相對減少,在處理過程中只需在升壓階段以液壓式高壓幫浦 加壓,而恆壓和降壓階段則不需要輸入能量;由於傳壓速度快、均勻,不 存在壓力梯度;超高壓殺菌不受食品的大小和形狀的影響,使得殺菌處理 過程較為簡單。對超高壓處理技術的研究發現,超高壓對食品風味、色澤 有關的小分子以及維生素等沒有太大影響,但可以改變蛋白質、多糖和脂 類等食物(生物)大分子的理化特性,如蛋白質的變性、脂肪結晶和澱粉糊化 等(涂等,2004)。
②高壓脈衝電場(PEF)殺菌技術:它是利用強電場脈波的介電阻斷原理 對食品微生物產生抑制作用。高壓脈波放電產生的強烈衝擊波以及紫外 光、強電流及臭氧等綜合效應較直流電流和低頻交流電壓更能有效殺死微 生物。其特點是能耗低、能量利用率更高、處理時間短、傳遞快速且均勻。
利用此技術殺菌時應考慮場強大小、殺菌時間和食品pH 值對殺菌的影響等 因素,確定殺菌最佳方案。但對高壓脈衝電場殺菌技術的研究迄今仍處於 詴驗階段,規模應用裝置的脈衝電場對食品產生的物理、化學、微生物、
酵素活性以及營養特性等方面的影響有待深入研究(涂等,2004)。
③輻射殺菌技術:它是利用電磁波中的χ射線、γ射線和放射性同位 素(如60Co)射線殺死微生物的方法。其基本作用是破壞菌體的脫氧核糖核酸 (DNA),同時有殺蟲、抑制植物發芽的作用。採用這種技術進行殺菌應遵照 我國輻照食品衛生管理的有關規定,選擇適當的照射劑量及時間,以保證 輻照食品的安全。目前有不少營養口服液及粉狀全營養素等製品均用此法 進行滅菌處理。其他如肉禽類、水產品、蛋品、果蔬類、穀類及其製品、
香料等也採用此技術殺菌。輻射殺菌的特點是射線可以穿透食品包裝和凍 結層,可以在食品包裝後輻照殺菌,避免二次污染;在適當的輻照劑量條 件下,食品營養成分不發生明顯變化,而且保鮮程度很高;利用輻照保藏 可以減少冷藏的壓力,節約能源(涂等,2004)。
④化學殺菌:主要是指在食品中通過添加抑菌劑和防腐劑,如臭氧、二氧 化氯和乳酸鏈球菌素等,從而達到抑菌或殺菌的目的。該方法易於操作、
控制,殺菌效果好,成本較低。但在使用中易受水分、溫度、pH 值和機 體環境等因素的影響,作用效果變化較大。另外食品中殘留的化學詴劑多 次使用可能使菌體產生抗體,同時也會影響食品的自然風味和質構。故在 應用食品殺菌技術時是根據不同食品原料的適用條件和實際工作情況,選 擇適當的殺菌方法,即在保證食品衛生安全的前提下,盡可能減少食品營 養、風味的損失,選擇成本較低的殺菌方法(涂等,2004)。
貳、研究目的
1. 探討蛹蟲草固態發酵條件對生產代謝產物蟲草素的影響。
2. 探討殺菌溫度和時間對蛹蟲草菌存活和生理活性物質滯留率的影響。
參、研究架構
圖 3、實驗設計流程圖。
Fig. 3. Flow chart of experimental design.
蛹蟲草菌(Cordyceps militaris BCRC 32219)
預活化(PDB 培養液,22℃,150 rpm,培養五天)
穀物基質:小麥、糙米、薏仁
氮源:魚粉、酵母粉、噴霧乾燥血漿蛋白粉 (SDPP)、豬腸膜蛋白粉(DPS)
熱殺菌實驗 液態基質(PDB) 固態基質(小麥)
麥角固醇菌絲分 析
腺苷和蟲草素分析
熱水浴(60℃~90℃), 0~10 分鐘
升降溫曲線、
蛹蟲草菌死滅詴驗、
腺苷和蟲草素熱穩定性詴 驗
蛹蟲草菌固態發酵,22℃培養 35 天
肆、材料與方法 1、實驗菌種
蛹蟲草(Cordyceps militaris BCRC 32219)購自食品工業研究所菌種保存 中心(新竹,台灣),以馬鈴薯葡萄糖洋菜培養基PDA(Potato dextrose agar, Difco)帄盤於22℃恆溫培養,每個月於無菌操作台(4HT-24桌上型,海天)內 繼代一次,並以Parafilm(Pechiney, plastic packaging, Chicago) 密封保存。
2、預活化蛹蟲草菌液
以 微 量 吸 管 (Acura 831, Socorex, Swiss)吸 取 血 清 瓶 (Schott, Duran, Germany)中已滅菌之無菌水5 mL,洗下PDA培養皿(Alpha plus scientific Corp., Taoyuan, Taiwan)的蛹蟲草菌絲,接入內含150 mL已滅菌冷卻之 PDB(Potato dextrose broth, Difco)培養基500 mL 搖瓶(Taifong)中,在22℃恆 溫振盪培養箱中(LM-600R, Yihder)以轉速150 rpm震盪培養5天後,即得活化 菌液。
3、不同活化天數對菌絲體的影響
以5 mL無菌水洗下PDA帄板的蛹蟲草菌絲,接入已滅菌冷却之150 mL PDB培養基中,在22℃恆溫振盪培養箱中以轉速150 rpm震盪培養,在第3 天、第5天、第7天、和第10天分別收取樣瓶,以高速冷凍離心機(Kubota, high speed refrigerated centrifuge 6500) 轉速8000 ×g,離心10分鐘後,收集菌絲 體,冷凍乾燥後秤重。
4、腺苷(adenosine)及蟲草素(cordycepin)的分析
以高效能液相層析儀(HPLC)分析腺苷(adenosine)及蟲草素含量(Chang et al., 2005),先製作腺苷及蟲草素標準曲線,將購於Sigma之標準品以15%
甲醇溶液(15 mL 100% methanol, 85 mL H2O)分別稀釋成100、200、300、
400、500及600 μg/mL,再以HPLC分析並繪製標準曲線。將各乾燥發酵樣 品稱取0.5 克裝入10 mL 樣品瓶中,再加入5 mL 15%甲醇溶液,置入100℃
烘箱中,萃取60分鐘。再將萃取液分裝至1.5 mL 微量離心管中,以轉速14000
×g,離心10分鐘,收集上清液,以0.22 μm濾膜進行過濾後,裝入新的微量 離心管中,以利後續HPLC分析。
4.1、HPLC分析條件
移動相(Mobile phase)為0.02 M KH2PO4: MeOH(85:15(v/v)),樣品注入 HPLC體積(Sample volume)為20 μL,流速(Flow rate)控制在1.0 mL/min,UV 偵 測 器 (Detector) 偵 測 波 長 為 254 nm , 分 析 管 柱 為 高 效 逆 相 層 析 柱 LiChrospher® 100 RP-18e (5 μm, Merck, Darmstadt, Germany)。移動相配製方 法為以精密天帄(AND GF-300)稱取 2.7218 克 KH2PO4 溶於850 mL 去離 子水完全溶解後加入100%甲醇150 mL於定量瓶(Taifong)中,以超音波震盪 機(Ultrasonic cleaner)(DELTA® DC 600H) 震盪10分鐘,再以0.22 μm濾膜進 行過濾。
5、菌絲體(Mycelium)的測定
5.1、純菌絲收集
以 5 mL 無菌水洗下 PDA 帄板的蛹蟲草菌絲,接入已滅菌冷却之 PDB 培養基中,在 22℃下轉速 150 rpm 的恆溫振盪培養箱中震盪培養 5 天後離 心,將菌絲體進行冷凍乾燥。
5.2、麥角固醇之測定(Chang et al., 2005)
將麥角固醇標準品(Sigma, USA)配成濃度 50 μg/mL、75 μg/mL、100 μg/mL、200 μg/mL 和 300 μg/mL,以 HPLC 所得面積繪製麥角固醇標準曲 線圖(附圖 A1)。HPLC 包括溶媒輸送幫浦(LC-20AT, Shimadzu, Kyoto, Japan)、紫外光/可見光偵測器(SPD-20A, Shimadzu, Kyoto, Japan),並連接 軟體 SISC 色層分析儀積分數據處理系統(Scientific information service corporation, 訊 華 股 份 有 限 公 司 ) 。 HPLC 條 件 如 下 : 分 離 管 柱 以 LiChrospher® 100 RP-18e(5 μm, Merck, Darmstadt, Germany)注射量為 20 μL,使用 100%甲醇(Labscan Asia Co., Ltd. Thailand)作為移動相,在檢測波 長 282 nm,流速 1.0 mL/min 下進行分析。
5.3、菌絲體標準曲線
取液態發酵後經冷凍乾燥之菌絲體粉末,加入無水酒精(Ethyl alcohol, 島久藥品株式會社,日本) 配製成10 μg/mL、25 μg/mL和50 μg/mL菌絲體標 準溶液,於烘箱(Memmert ULM500-1)中,60℃萃取60分鐘,以轉速8000 ×g 離心10分鐘後,(TERUMO® Syringe with needle (TERUMO®, Philippines))取