國立臺灣大學醫學院藥理所
國立臺灣大學醫學院藥理學研究所 碩士論文
Graduate Institute of Pharmacology
碩士論文
O r e x i n s 對小鼠海馬迴神經突觸可塑性之研究
李家旭 撰
College of Medicine
National Taiwan University Master Thesis
Orexins 對小鼠海馬迴神經突觸可塑性之研究
Effect of orexins on synaptic plasiticity of mouse hippocampal slices
李家旭
Lee Chia-Hsu
指導教授:邱麗珠 博士
Advisor: Chiou Lih-Chu, Ph.D.
中華民國 97 年 7 月 July, 2008
97 7
目 錄
頁次
一、 口試委員會審定書 ……… i
二、 誌謝 ……… ii
三、 縮寫表……… iii
四、 中文摘要……… iv
五、 英文摘要 ……… vii
六、 緒論……… 1
七、 實驗材料及方法……… 14
八、 實驗結果……… 19
九、 討論……… 26
十、 結論……… 32
十一、 圖表與附圖 ……… 33
十二、 參考文獻 ……… 53
誌謝
時間飛快地流逝,在這不長不短的兩年裡,我從帶著獸醫背景進
入藥理的世界,實驗室的大小事在我眼中總是新鮮又陌生,我也很高
興能進入這樣的實驗環境,首先要感謝 邱麗珠老師,總是為了學生著
想,不論是學業或生活態度,總是耐心地討論,讓我知道我哪裡要改
進,雖然時常有不盡理想的表現,老師還是以無比的耐心包容。這兩
年的收穫是很豐富的,有歡笑有汗水,從跌倒的地方明瞭自己的缺失,
從讚美之中明白有努力付出才有收穫。
實驗室成員們大家感情都很融洽,時常互助合作。短短兩年我想
最辛苦的就是彥昱學長,大小事情都一把罩,平日常給學長添的麻煩,
由衷感謝學長的照顧。另外一位要感謝的重要成員就是欣蓉學姐,電
腦軟硬體教學多職,不但協助各項雜務,教導許多實用的軟體,精神
上也常常給予我許多支持。此外還有許多要感謝的人,像是要謝謝廖
蕙萍學姊帶我做實驗;謝謝范碧娟醫師在 seminar 給予指導;謝謝李
建興學長和李君耀學長在 seminar 給予指正;謝謝實驗室的夥伴,俐
君、昱征、心慈和宏烈平日的互助鼓勵;謝謝珮筠一直都在我身邊鼓
勵我幫助我;謝謝建誠、士函、建邦和于凡等許多好友們的關心鼓勵,
我們做到了;感謝我的父母與妹妹多年來無怨無悔的支持我,最後我
想將此份快樂獻給在天上的爺爺,希望您會開心。
縮寫表( Abbreviation )
ACSF artificial cerebral spinal fluid
AMPAR alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor CA1 cornu ammonis 1
cAMP cyclic adenosine monophosphate
DG dentate gyrus
fEPSP field excitatory post-synaptic potential GABA gamma-aminobutyric acid
GIRK G-protein coupled inwardly rectifying potassium channel GPCR G-protein coupled receptor
HFS high frequency stimulation LFS low frequency stimulation LTP long term potentiation LTD long term depression NMDA N-methyl-D-aspartate
NMDAR N-methyl-D-aspartate receptor OXR oreixn receptor
OX1R orexin 1 recepor OX2R orexin 2 receptor
PKA cAMP-dependent protein kinase PKC protein kinase C
PLC phospholipase C PTX pertusis toxin
PPF pair pulse facilitation PPR pair pulse ratio
PTP post tetanic potentiation REM rapid eye movement
SB 334867 N-(2-Methyl-6-benzoxazolyl)-N``-1,5-naphthyridin-4- yl urea TBS theta brust stimulation
VTA ventral tegmental area
摘要
分佈於下視丘為主的orexin neurons 會經由腦部廣泛投射的軸突釋出 orexins,
可分為orexnin A (hypocretin 1)和 orexin B (hypocretin 2),會與 orexin receptors 結 合。這個受體是在1998 年被去孤兒化的 G 蛋白偶合受體,又分做兩種亞型,orexin 1 receptor (OX1R)和 orexin 2 receptor (OX2R)。OX1R 對於 orexin A 的親和力比 orexin B 高,但是 OX2R 對於兩者的親和力則相似。這兩種 orexin receptors 的在腦 中分佈極廣,並與自下視丘orexin neurons 投射出的軸突所釋放的 orexins 接合,被 証實與許多生理病理的機轉有關。hippocampus 中也有 orexin neurons 投射的軸突 和orexin receptor 的分布,且在許多以大鼠實驗對象的 in vivo 實驗中被證實會影響 hippocampus 的 突 觸 可 塑 性 以 及 學 習 和 記 憶 的 表 現 。 而 以 大 鼠 和 小 鼠 的 hippocampus 腦切片為實驗材料的 in vivo 實驗中,亦發現 orexins 有影響神經基礎
突觸傳遞和突觸可塑性的現象。但 orexin 的角色仍眾說紛紜。因此,我們利用小
鼠hippocampus 切片,觀察 Schaffer Collateral-CA1 區域中,外給 orexins 對基礎突 觸傳遞和神經可塑性,包括長期增益現象和去增益現象,有何種影響。藉由在 Schaffer Collateral 至 CA1 的傳遞路徑上給予電刺激,CA1 區域中的 stratum radiatum 進行胞外紀錄,所得到的場興奮性突觸後電位(fEPSPs)進行分析。長期增益現象 (long term potentiation, LTP)是由高頻所引發刺激,我們所用的高頻刺激是由三個相 隔15 秒的 theta 頻率構成之刺激組成,每個 theta 頻率的刺激是由 4 段 100 HZ 的 高頻刺激構成,每段相隔200 秒。去增益現象(depotentiation)則是在高頻刺激後四 十分鐘,由連續的低頻(1 Hz)刺激 15 分鐘所引發。
灌流oirexin A (30, 100, 300 nM) 10 分鐘後,所測得 fEPSPs 的斜率,並沒有影 響基礎傳遞突觸。但orexin A 會有意義的抑制 LTP 且具濃度相依性。高頻刺激後 4 分鐘內的fEPSPs 的斜率變化,即後強直增強效應(post-tetanic potetanic, PTP),亦 有輕微濃度相依的抑制情形,但不具統計上的差異。而各個濃度對去增益現象的 影響,亦無統計上顯著差異。
灌流[Ala11, D-Leu15]-orexin B (3, 10, 30, 100, 300, 1000 nM) 10 分鐘後,並沒有
發現影響基礎傳遞突觸,在低濃度即可有意義的抑制 LTP,但隨濃度的升高並無
法增加抑制 LTP 的程度。觀察 PTP,各組間不具統計上的差異。而各濃度對
depotentiaion,亦無統計上的顯著影響。
將3 μM 專一性 OX1 拮抗劑 SB 334867 與 100 nM orexin A 同時灌流,發現可 部分反轉orexin A 對 LTP 所造成的抑制。對 PTP 的輕微抑制亦有些許的反轉,但 皆未達統計上顯著的差異。而各個濃度對 depotentiation 的影響,亦無統計上顯著 差異。單獨投予SB 334867 對基礎突觸傳遞、LTP、PTP 和 depotentiation 並無影響。
再將3 μM 專一性 OX1 拮抗劑 SB 334867 與 300 nM [Ala11, D-Leu15]-orexin B 同時灌流,發現可部分反轉[Ala11, D-Leu15]-orexin B 對 LTP 所造成的抑制。對 PTP 並無影響。各組間對depotentiaion 的影響,則無統計上顯著差異。
將30 μM 專一性 OX2 拮抗劑 Compound 29 與 100 nM orexin A 同時灌流,發 現可部分反轉 orexin A 對 LTP 所造成的抑制,對 PTP 並無影響。各濃度對 depotentiation 的影響,則無統計上顯著差異。單獨投予 Compound 29 對基礎突觸 傳遞、LTP、PTP 和 depotentiation 並無影響。
再 將 30 μM 專 一 性 OX2 拮 抗 劑 Compound 29 與 300 nM [Ala11, D-Leu15]-orexin B 同時灌流,發現可部分反轉[Ala11, D-Leu15]-orexin B 對 LTP 所造 成的抑制。對PTP 並無影響。各組間對 depotentiation 的影響,則無統計上顯著差 異。
在hippocampus CA1 突觸會因極短的成對刺激(50 ms)後,鈣離子於突觸前堆 積,而出現paired pulse facilitation 的特性。計算 paired pulse ratio (PPR),由第二個 fEPSP 斜率除與第一個 fEPSP 斜率,來評估突觸前神經傳遞物釋放的變化。我們 發現在給予高頻刺激後40 分鐘會有 PPR 下降(1.62±0.04 vs. 1.51±0.03, n=4),即突
觸前神經傳遞物釋放增加。一般認為LTP 是以突觸後 NMDAR 開啟來為主,而我
們認為LTP 應同時也有增加突觸前神經傳遞物釋放。orexin A (100 nM)並不影響基 礎刺激下的PPR,但有輕微減少 PPR 下降程度的現象。
根據我們目前的實驗結果,在小鼠hippocampus CA1 區域,發現 orexin A 和 [Ala11, D-Leu15]-orexin B 皆會部分抑制 LTP,但並不影響 depotentiation。對 LTP 的 抑制效果,可能同時經由OX1R 和 OX2R,但 OX1R 可能佔有較大的比重。再者,
突觸前的抑制效果可能很輕微,推測可能是在突觸後,藉由活化 OX1R 和 OX2R
活化PLC/PKC 的途徑,造成抑制 NMDA 所媒介的電流為主。
關鍵字:海馬迴,Orexin,神經突觸可塑性,長期增益現象,去增益現象。
Abstract
Orexins are a novel family of two hypothalamic peptides, consisting of orexin A and orexin B, which were also named hypocretin 1 and hypocretin 2, respectively. They were identified to be the endogenous agonists of an orphan GPCR in 1998, which was, therefore, named orexin receptor and consists of OX1 and OX2 receptors. OX1 receptors display higher affinity for orexin A than orexin B, while OX2 receptors have similar affinity for both orexin A and orexin B. Orexin-containing neurons are mostly localized in the lateral hypothalamus and send projections widely through the brain, including the hippocampus. Orexin A had been reported to affect learning and memory tasks and hippocampal synaptic plasticity in vivo, mostly in the rat, as well as affected synaptic plasticity of hippocampal slices of rats and mice in vitro. However, controversial results were reported. We, therefore, investigated the effects of orexins on the synaptic plasticity, long-term potentiation (LTP) and depotentiation, at Schaffer Collateral-CA1 synapses of mouse hippocampal slices. Field excitatory postsynaptic potentials (fEPSPs) were evoked by stimulating the Schaffer collateral pathway and recorded in the stratum radiatum of the CA1 region of hippocampal slices. LTP was induced by high frequency stimulation (HFS) which consisted of 3 trains of theta burst stimulation (TBS), separated by 15s. Each TBS consisted of 4 pulses at 100 Hz, separated by 200 ms. Depotentiation was induced by low frequency stimulation (LFS, 1Hz, 15 min) 40 min after HFS.
Orexin A (30-300 nM) had no effect on the baseline fEPSPs after 10 min treatment.
Orexin A treated 10 min before and during HFS, decreased HFS-induced LTP dose-dependently. The magnitude of LTP (% of the baseline fEPSPs) 40 min after HFS was reduced by pretreatment with orexin A 100 nM and 300 nM. The magnitude of PTP (% of the baseline fEPSPs) 40 min after HFS was mild dose-dependently reduced by
pretreatment with orexin A 30 nM, 100 nM and 300 nM. Depotentiation induced by LFS (expressed by the % restoration of the magnitude of LTP) 40 min after LFS was not different with orexin A.
[Ala11, D-Leu15]-orexin B (3~1000nM) also had no effect on the baseline fEPSPs after 10 min treatment. [Ala11, D-Leu15]-orexin B treated 10 min before and during HFS, decreased HFS-induced LTP dose-dependently. The magnitude of LTP (% of the baseline fEPSPs) 40 min after HFS was reduced by pretreatment with [Ala11, D-Leu15]-orexin B 30 nM, 300 nM and 1000 nM. The magnitude of PTP (% of the baseline fEPSPs) 40 min after HFS had no different by pretreatment with [Ala11, D-Leu15]-orexin B. Depotentiation induced by LFS (expressed by the % restoration of the magnitude of LTP) 40 min after LFS was not different with [Ala11, D-Leu15]-orexin B.
SB 334867 (3 μM), a selective OX1 receptor antagonist, partially antagonized the effect of orexin A. The magnitude of LTP (% of the baseline fEPSPs) in slices treated with orexin A+ SB-334867 was higher in slices with 100 nM orexin A, which was significantly lower than the control slices. The magnitude of PTP (% of the baseline fEPSPs) in slices treated with 100 nM orexin A had no significantly difference in slices with orexin A+ SB-334867, which was not significantly different with the control slices.
SB-334867 alone had no effect on basal transmission, PTP, LTP and depotentiation.
SB 334867 (3 μM) slightly antagonized the effect of [Ala11, D-Leu15]-orexin B.
The magnitude of LTP (% of the baseline fEPSPs) in slices treated with 300 nM [Ala11, D-Leu15]-orexin B + SB 334867 was higher in slices with [Ala11, D-Leu15]-orexin B, which was significantly lower than the control slices. The magnitude of PTP (% of the baseline fEPSPs) in slices treated with 300 nM [Ala11, D-Leu15]-orexin B had no
statistically significant difference in slices with [Ala11, D-Leu15]-orexin B + SB 334867, which was not significantly different with the control slices. SB 334867 alone had no effect on basal transmission, PTP, LTP and depotentiation.
Compound 29 (30 μM), a selective OX2 receptor antagonist, partially antagonized the effect of 100 nM orexin A. The magnitude of LTP (% of the baseline fEPSPs) in slices treated with 100 nM orexin A + Compound 29 was higher in slices with 100 nM orexin A, which was significantly lower than the control slices. The magnitude of PTP (% of the baseline fEPSPs) in slices treated with 100 nM orexin A was no different in slices with orexin A + Compound 29, which was no significantly difference with the control slices. Compound 29 alone had no effect on basal transmission, PTP, LTP and depotentiation.
Compound 29 (30 μM) slightly antagonized the effect of [Ala11, D-Leu15]-orexin B. The magnitude of LTP (% of the baseline fEPSPs) in slices treated with [Ala11, D-Leu15]-orexin B + Compound 29 (133±7.1%, n=5) was higher in slices with 300 nM [Ala11, D-Leu15]-orexin B (120.8±7.2%, n=5), which was significantly lower than the control slices (141.9±3.5%, n=16). The magnitude of PTP (% of the baseline fEPSPs) in slices treated with 300 nM [Ala11, D-Leu15]-orexin B (146.8±8.7%, n=5) was not significantly different in slices with [Ala11, D-Leu15]-orexin B + Compound 29 (186.5±28%, n=5), which had no significantly difference with the control slices (181.5±10.4 %, n=16). Compound 29 alone had no effect on basal transmission, PTP, LTP and depotentiation.
Paired-pulse facilitation (PPF) of fEPSPs, a presynaptic phenomenon resulting from Ca2+ accumulation in the presynaptic terminals in response to stimulation by a pair pulses with short interval is induced by paired-pulse (50 ms interval) stimulation. The
PPR during LTP decreased, as compared to basal PPF ratio (PPR), suggesting that increased presynaptic transmission plays a role in the LTP evoked by TBS in CA1 region of mouse hippocampal slices although this LTP is mostly mediated by NMDA receptors. Orexin A (100 nM) did not affect basal PPR but slightly reversed the PPR decreased during LTP.
It is concluded that orexin A and [Ala11, D-Leu15]-orexin B decreased LTP but not affect depotentiation in CA1 region of mouse hippocampal slices. The impairment of LTP is mediated by both OX1 and OX2 receptors. The efficacy mediated by OX1 receptors is higher than that by OX2 receptors. It is suggested that presynaptic inhibition plays a little role in orexin-induced impairment; instead, postsynaptic NMDA receptor inhibition might contribute to this impairment through PLC/PKC pathway activated by OX1 and OX2 receptor activation.
Key word: hippocampus, orexin, neural plastisity, long term potentiaion, depotentiation
緒論
1. Hypocretins/ Orexins 的發現
1998 年de Lecea 等學者發現大鼠下視丘( hypothalamus )有兩種結構近似於腸 道激素,secretin,的胜肽,由於只分佈於下視丘,因此被命名為hypocretin。這兩 種hypocretins 是由同一種前驅蛋白, preprohypocretin,經過水解後所得( Fig. 1 )。
其中含有38 個胺基酸的胜肽稱為hypocretin 1,而29 個胺基酸的胜肽則稱為 hypocretin 2。同時,他們也利用北方點墨法( Northern blot ) 及原位雜交法( in situ hybridization ) , 發 現 在 小 鼠 的 第 11 對 染 色 體 可 在 出 生 後 三 週 開 始 轉 錄 出 preprohypocretins mRNA,且只分布在下視丘背側與外側區( dorsal and lateral hypothalamuic area)的神經細胞體,由於下視丘背側與外側區與進食( feeding )、血 壓與中樞對免疫系統等的調控有關,因此起初推測hypocretin 在這些生理功能中扮 演重要角色(de Lecea et al., 1998)
同年,Sakurai 等學者(Sakurai et al., 1998b)使用大鼠全腦萃取物在表現各種G 蛋白偶合孤兒受體(orphan G protein-couple receptor, orphan GPCR )的細胞株上篩選 這些G 蛋白偶合孤兒受體的內生性受質時,發現大鼠全腦萃取物中有兩種新的神 經胜肽,對某orphan GPCR 有高親和力,並且有使得細胞內的鈣離子顯著上升的 特性。在動物實驗的結果顯示,此兩種胜肽都會增加老鼠攝食行為,故被命名為 orexins,其中orexin A 具有兩個分子內雙硫鍵且含有33 個胺基酸、分子量為3562 Da。orexin B 則含有28 個胺基酸、分子量為2973 Da 。這兩種神經胜肽皆從同一 種前驅蛋白,prepro-orexins,而來。同時,這個會與oreixns 結合的orphan GPCR 被 稱為orexin receptors(Sakurai et al., 1998a)。Sakurai等學者表示,他們所分離而得的 orexins 與de Lecea 等學者所發現的hypocretin 是相同的胜肽,意指orexin A 即為 hypocretin 1,而orexin B 為hypocretin 2 ,兩者的差異僅在於hypocretin 1較orexin A 在C端多出5個無功能的胺基酸,而hypocretin 2 在N 端則多出一個Glycine (Fig.
1)(Sakurai et al., 1998b)。這兩個實驗團隊所發現的orexins與先前其他已知的胜肽完 全不同,是為一個全新種類的胜肽。因此,後續許多作者發表相關結果時,都會
使用orexins/ hypocretins 或hypocretins/ orexins 通稱之(Kilduff and Peyron, 2000)。
2. Orexins 的介紹
會有orexin( hypocretin ) neurions的神經細胞,主要聚集於下視丘,包括於下視 丘外側( lateral hypothalamus, LHA ) 和下視丘背側( posterior hypothalamus, PHA ) 和穹窿狀周邊區( perifornical area, PFA )(de Lecea et al., 1998;Sakurai et al., 1998a),但是其投射到非常廣泛的腦區,除了小腦以外,皆屬其投射的範圍,包括:
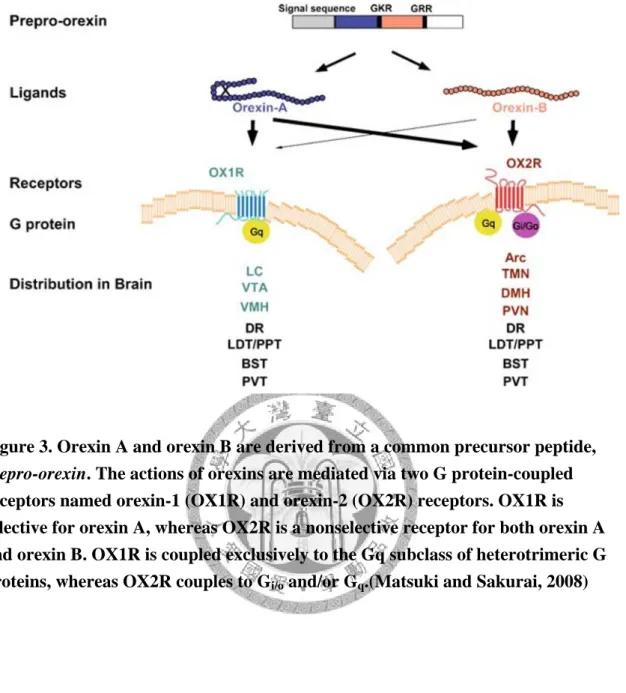
central medial nucleus (CM) 和paraventricular nucleus of the thalamus (PVT);下視丘 的arcuate nucleus (Arc),ventromedial nucleus (VMH),dorsomedial nucleus (DMH) 和 tuberomammillary nucleus (TMN);locus coeruleus (LC) 和 ventral tegmental area (VTA) 等區(Fig. 2)(Mintz et al., 2001; Nambu et al., 1999a; Peyron et al., 1998)。
由orexin neurons 投 射 纖 維 所 分 泌 出 的 orexins 乃 是 屬 於 神 經 多 胜 類 (neuropeptide), 如前段所提,orexins 又分為A 及B 兩種型態(即 hypocretin 1 及 hypocretin2 ),在哺乳類中,兩者皆來自於共同一段130~131氨基酸的前驅物,
prepro-orexin,經由裁切後可形成orexin A 或orexin B,分別含33 和28 個氨基酸。
比較其結構,orexin A有兩個雙硫鍵,而orxin B則是線狀的胜肽。Orexin A在人、
豬、狗、大鼠和小鼠有相似的結構,但orexin B在種別間則差異較大(Kukkonen et al., 2002)。此外,與orexin生成有關的基因密碼已證實位於人類染色體17q21 這個位置 上(de Lecea et al., 1998; Sakurai et al., 1998a),這個位置與人類神經退化性疾病有 關,例如:巴金森氏症(parkinsonism) 以及失智症(dementia)(Wijker et al., 1996;
Wilhelmsen et al., 1994)。同樣的,在動物行為實驗中,orexins 也已經證實可以影 響記憶的獲取(acquisition)、固化(consolidation)和提取(retrieval)記憶的能力。(Akbari et al., 2008; Akbari et al., 2006; Akbari et al., 2007; Aou et al., 2003; Selbach et al., 2004; Wayner et al., 2004)。
3. Orexin receptors
當 人 類 基 因 被 解 碼 之 後 , 近 三 百 種 的G 蛋 白 偶 合 受 體 (G protein-couple receptors, GPCR)被養殖出來,這些受體有著穿膜七次的特質。其中有三分之二被 認為是體內神經傳導物質的受體,另外一百多種被養殖出來的G 蛋白偶合受體尚 未發現其內生性受質,因而被稱為G 蛋白偶合孤兒受體(orphan GPCR)(Chiou et al., 2007)。這些orphan GPCR被認為是許多人類束手無策疾病的可能標的,而廣為科 學家或製藥界重視,被視為在藥物發展上良好的作用標的(Stadel et al., 1997)。這些 orphan GPCR若找到其受質,其將被去孤兒化( de-orphanized ),OX受體即是第二 個被去孤兒化的GPCR。由1998 年Sakurai 等學者如前述方式,藉由大鼠全腦萃取 物在含有orphan GPCR細胞株篩選,找出兩種新的神經胜肽orexins,並將可以與 orphan GPCR結合的受體命名為OX 受體。這類受體兩種亞型,便分別命名為OX1 受體(OX1R) 與OX2 受體(OX2R),而OX2R 有64﹪的氨基酸序列與OX1R 相同,
從此這兩種G 蛋白偶合受體便被去孤兒化。此外Sakurai 等學者發現orexins 對於 OX1 受體(OX1R)與OX2 受體(OX2R)有不同的親和力,OX1R 對orexin A 的親和 力比orexin B 高,orexin A (EC50=30 nM) 大於orexin B (EC50=2500 nM),但OX2R 對orexin A 及orexin B 親和力則相同(Sakurai et al., 1998a)。2003 年Zhu 等學者使 用BIM 細 胞 株 ( 由 human neuroblastoma cells 及 thymidine auxotrophs of rat nerve-like cells 雜交而來 )(Ishii et al., 1990) 表現OX1R 及OX2R, 發現與OX1R 偶合的G-protein 是PTX( pertussis toxin )-insensitive G-protein,目前認為是Gq/11和 Gs為主,其可藉由活化磷脂酶C ( phospholipase C, PLC )和磷脂酶A ( phospholipase A, PLA )進而活化其下游機制。而OX2R 則可能分別與PTX-sensitive ( Gi/o ) 及 -insensitive G-protein ( Gq/11 ) 偶合(Fig. 3)(Matsuki and Sakurai, 2008; Spinazzi et al., 2005; Zhu et al., 2003)。為了能夠詳盡地研究OX1R 與OX2R 的特性,於是科學家 研發對這兩種受體有選擇性胜肽類與非胜肽類的致效劑與拮抗劑( Tab. 1)(Cheng et al., 2005)。在本實驗中,分別使用SB-334867、[Ala11, D-Leu15] orexin-B和compound 29,三個合成的藥物來研究兩類OXR所媒介的反應。SB-334867是為對OX1R 具有
選擇性的拮抗劑(Smart et al., 2001); 而[Ala11, D-Leu15] orexin-B ,是由orexin B 將 其中的胺基酸取代並修飾過後而得,對於OX2R 是高度選擇性的致效劑(Asahi et al., 2003),其OX1R:OX2R 的選擇性為1:400。此外,本實驗中亦利用OX2R 之 選擇性拮抗劑N-Acyl 6,7-dimethoxy-1,2,3,4-tetrahydroisoquinoline,簡稱為compound 29,對OX2R的選擇性為OX1R的250倍(Hirose et al., 2003)。利用上述幾類種藥物則 可研究orexins 的作用是經由OX1R 或OX2R 所引發。
4. Orexins的細胞作用機制
由於認為兩個OXR 的G protein 有所不同,OX1R 是會經由Gq/11和Gs,而 OX2R 則Gq/11 ,Gi/o 和Gs。藉由上述的G 蛋白,orexins 對於神經細胞內離子和通 道的作用,總括而言,可簡單分為4 類:(1)增加細胞內鈣離子濃度(2)抑制鉀離子 通道和GIRK 通道(3)開啟非選擇性的陽離子通道( non-selective cation channel, NSCC )(4) 活 化 Na+/Ca2+ exchanger(Kukkonen and Åkerman, 2005) 。 用 Chinese hamster ovary (CHO) cell或human embryonic kidny 293( HEK 293) cell等細胞株表現 OXR 的體外實驗( in vitro)之中,發現orexin A 和orexin B 均可透過OX1R 會增加 細胞內鈣離子濃度,而orexin B 在OX1R 作用能力比orexin A差,orexin A 以及 orexin B 透過OX2R 增加細胞內鈣離子濃度的能力相似(Sakurai et al., 1998a)。
Kukkonen以及Åkerman ( 2001)在相同實驗模式下,發現orexin A 增加細胞內鈣離 子是經由orexin- receptor- operated Ca2+ influx 以及store- operated Ca2+ influx 造成 (Kukkonen and Akerman, 2001)。利用同時表現OX1R和OX2R以及GIRK通道的 HEK-293細胞株,發現orexin A可瞬間興奮後長期地抑制GIRK通道。由於興奮的作 用可受到PTX的抑制,但抑制的作用則否。因此推論活化GIRK是經由Gi/o的Gβγ, 而抑制的作用則是經由Gq所媒介的PLC途徑所引發(Hoang et al., 2003)。在nucleus tractus solitarius,除了GIRK通道外的鉀離子通道亦會受到orexin A的經由活化 PLC/PKC的抑制現象(Yang et al., 2003)。有趣的是,許多去極化的研究都顯示,
Voltage-dependent calcium channel (VDCC) 都有參與其中,因此認為這是因為
orexins 藉由減少鉀離子通道電導而開啟VDCC(Kukkonen and Åkerman, 2005)。此 外,Uramura等學者在ventral tegmental area 的dopaminergic 神經元,發現orexin A 增加細胞內鈣離子的濃度是透過phosphatidyl choline- specific PLC- PKC- L/ N- type 鈣離子通道的訊息傳遞,並發現有增加NMDA所媒介電流的現象(Borgland et al., 2006; Uramura et al., 2001)。在非神經細胞實驗中,在人類與大鼠腎臟髓質上發現,
給予orexin A 經由OX1R 造成cortisol 分泌的現象,會因給予adenylate cyclase 或 是PKA 抑制劑而使分泌量下降,因此推測OX1R 可能偶合於Gs 蛋白 (Spinazzi et al., 2006)。
此外,orexins也會影響NMDA所媒介的電流,在ventral tegmental area可以使 NMDA所媒介的電流上升(Borgland et al., 2006)。但在hippocampus CA1區域,分別 利用大鼠和小鼠的腦薄片,利用胞外紀錄的方式,觀察NMDA電流的變化,卻發 現無論是投與30nM orexin A 30 min 或100 nM 5 min 皆有減少NMDA電流 20~30%的現象(Aou et al., 2003; Selbach et al., 2004),似乎暗示著在hippocampus CA1區域的NMDAR會受到orexin A的影響,而有功能或是數量上削減的跡象。
5. Hypocretins/ Orexins 以及OXR 的分佈
科學家以原位雜交法( in situ hybridization )(Bingham et al., 2001; Date et al., 2000; Hervieu et al., 2001; Peyron et al., 1998; van den Pol, 1999)及免疫組織染色法 ( immunohistochemistry )(Cutler et al., 1999; de Lecea and Sutcliffe, 1999; Kilduff and Peyron, 2000; Sakurai et al., 1998a; van den Pol, 1999) 得知orexins 和OXR 在CNS 的分佈區域,發現orexins 的蛋白質與OXR 之mRNA 或蛋白質可在腦及脊髓中發 現(Fig.2,3)。
6. Orexins 的生理功能與病理角色
自orexins 被發現以來,許多研究結果發現orexins 在清醒/睡眠、攝食行為、
報償機制、疼痛調節和記憶學習等機制,扮演著重要的調控角色,以下將逐一簡 單介紹。
6.1 清醒/睡眠(sleep-wake)機制及猝睡症(narcolepsy)
清醒/睡眠機制及猝睡症的病理機制中,orexins 的參與其中是近來亟為令人振 奮的突破。若於老鼠的腦室內給予orexin A,利用腦波圖(electroencephalogram, ECG) 監測睡眠覺醒週期,發現使得動物增加警覺期(wakefulness),延緩睡眠的開始(onset time)且不易入睡,入睡後明顯縮短快速動眼期( rapid eye movement, REM ) 及非快 速動眼期( non-REM )(Beuckmann and Yanagisawa, 2002; Mieda and Yanagisawa, 2002),但orexin B 卻沒有顯著差異(Hagan et al., 1999)。因為orexins 可以活化許多 與動物保持清醒有關的核區,包括locus coeruleus 的noradrenergic 神經元 (Hagan et al., 1999),ventral tegmental area 的dopaminergic 神經元(Nakamura et al., 2000),
dorsal raphe 的serotonergic神經元 (Brown et al., 2002; Liu et al., 2002),和TMN 的 histaminergic 神經元(Yamanaka et al., 2002)。較有進展的部分是對histamine的研 究,對已於側腦室投與orexin A的大鼠給予H1 接受器阻斷劑,pyrilamine,則會發 現有大量減少其清醒時間的現象(Yamanaka et al., 2002)。若利用H1 接受器缺失的 小鼠,可以觀察到其清醒時間並不會受到orexin A 投予而有增加的現象(Huang et al., 2001)。因此推論orexin使動清醒的機制可能與分佈許多histaminergic 神經元之 TMN核區有著極大的關係。直接觀察preproorexin 基因剔除(knockout)的老鼠,會 有猝睡症的現象,以及破壞狗的OX2R 的基因,也會導致猝睡症(Chemelli et al., 1999; Lin et al., 1999),因而推測orexins在猝睡症的病理機制中扮演重要的角色。若 測量病人腦脊髓液( cerebrospinal fluid, CSF )中orexins 濃度,發現在猝睡症病人的 CSF 與控制組比較,猝睡症病人的CSF 測不到或測到濃度很低的orexins ,因此 orexins 的濃度可視為嗜睡症的臨床診斷指標之一(Dalal et al., 2001; Nishino et al., 2000)。
6.2 攝食行為 (feeding behavior)
Sakurai等學者( 1998a)使用免疫染色法發現orexin neurons主要存在下視丘,其 中LHA這個區域與進食行為( feeding behavior) (Anand et al. 1951)而其他的下視丘 區域,例如 :Arc (arcuate nucleus), VMN (ventomedial nucleus), DMH (dorsomedial hypothalamic nucleus), PVN (paraventricular hypothalamic nucleus )則與能量代謝 ( energy hemostasis)有關(Elmquist et al., 1999)。若將orexins 直接注入老鼠的側腦室 後,會顯著促進動物的進食與飲水量,且orexin A 的效用比orexin B 顯著;若將 動物禁食48 小時,或外給胰島素造成體內血糖下降(hypoglycemia),則促使內生性 orexin 的mRNA 代償性增加(up-regulation),進而影響 hypothalamic- pitutary- adrenal axis 的調節(Kunii et al., 1999; Sakurai et al., 1998a)。在電生理in vitro 的實 驗上發現,hypothalamic orexin neurons會受到胞外葡萄糖濃度的影響,而改變其靜 止 膜 電 位 和 自 發 性 動 作 電 位 的 頻 率 。 當 胞 外 葡 萄 糖 濃 度 上 升 , 會 有 過 極 化 (hyperpolarization)和減低自發性動作電位的頻率的現象(Yamanaka et al., 2003)。相 反的,胞外葡萄糖濃度下降,會有去極化(depolarization)和增加自發性動作電位的 頻率的現象(Burdakov et al., 2005; Yamanaka et al., 2003)。在臨床觀察上,測量肥胖 小孩血清的orexin A含量發現較正常組為低(Lou et al., 2006)。而若他們藉由減少飲 食攝取和多運動,發現隨著體重的減輕,orexin A會有回升的趨勢(Bronsky et al., 2007)。在成人中,亦發現肥胖病人的血清oreixn A濃度較低(Adam et al., 2002;
Baranowska et al., 2005),同樣的,在血清orexin A濃度較低的肥胖病人進行胃縮小手 術 , 術 後 病 人 體 內 orexin A 出 現 上 升 的 情 形 (Heinonen et al., 2005) 。CSF 中 prepro-orexin 濃 度 , 會 與 小 鼠 體 內 的 血 中 葡 萄 糖 濃 度 和 食 物 攝 取 量 呈 反 比 (Yamanaka et al., 2003)。綜觀上述結果,orexins傾向在身體處於缺乏飢餓時釋放,
而在身體能量充足時減少。
但觀察猝睡症病人在臨床上的表現後發現,典型猝睡症的病人因為orexins系 統的缺失,因而有減少食物攝取量的傾向,但卻因基礎代謝率的下降,使得身體 質量指數增加(body mass index, BMI)的現象(Chabas et al., 2007; Lammers et al.,
1996; Schuld et al., 2000),同樣的,在OXR 基因剔除小鼠亦有減少食量,但卻有 後期過胖(late-onset obesity) 的現象(Hara et al., 2001; Hara et al., 2005)。以上證據顯 示orexin 屬於除了會促進食慾外,其對體內基底代謝率的維持也有相當的影響 (Honda et al., 1986; Schuld et al., 2000)。
6.3 壓力(stress)
與壓力相關的研究中,已知orexin neurons會因為寒冷和束縛( immobilized )而 活化(Sakamoto et al., 2004)。將orexins 注入動物的側腦室中,會促進血液中腎皮質 固醇( corticosterone )的濃度(Hagan et al., 1999),可能是經由刺激腦下垂體釋放促腎 上腺素皮質醇( Adrenocorticotropic hormone;ACTH )(Engelmann et al., 2004),此項 結果暗示著orexin 可能參與壓力(stress)有關的生理功能(Dun et al., 2000; Ida et al., 2000)。
6.4 疼痛( pain)
若 將orexin A 以 腦 室 、 腹 腔 及 靜 脈 投 予 後 觀 測 老 鼠 在 疼 痛 發 炎 模 式 ( carrageenan-induced thermal hyperalgesia)及熱板(hot plate test) 測試下的痛覺反 應,發現orexin A 以腦室和靜脈投予時,在疼痛發炎模式中具有止痛的效果,並 可用OX1R 拮抗劑對抗orexin A 的止痛效果。當脊髓注射orexin A 後再給予SB 334867,則看見止痛效果被抑制,單獨腹腔給予SB 334867,則使疼痛加劇,表示 此種發炎模式有內生性orexin A釋出。其對發炎反應所產生的止痛效果,可能與腦 部和脊髓部位的OX1R 有關。但在熱板測試下的急性疼痛(acute pain) 模式,並不 會受到SB 334867的影響,因此推論在急性疼痛模式下,沒有內生性orexin A 的釋 放(Bingham et al., 2001)。術後疼痛模式上,單獨給予SB 334867 會看見老鼠有更 痛的情況產生,因此推測術後疼痛模式可能有內生性的orexins 參與;以上可推知 orexin A 對疼痛反應所產生的止痛效果,可能大多與OX1R 有關。由於naloxone 不能對抗orexin A的止痛效果,所以其作用與鴉片系統無關(Cheng et al., 2005)。
6.5 自主神經系統(autonomic nervous system)
在自主神經系統有關的研究中,發現orexins 注射於側腦室時,可增加心跳速 率(heart rate)及平均動脈壓,但如先投予α或β受體拮抗劑時,這個效果就會消失 (Shirasaka et al., 1999)。同時也可以可觀察到在缺乏orexin的小鼠血壓會低於一般的 小鼠(Kayaba et al., 2003; Zhang et al., 2006) 上述的結果似乎都支持orexin會增加交 感神經(sympathetic tone)的興奮性。因此,缺乏orexin的小鼠便減低了能量的耗損。
也就是說,orexins雖然增加了攝食行為,但同時orexins 也增加了代謝的速率 (Lubkin and Stricker-Krongrad, 1998) 。 這 也 解 釋 了 動 物 在 攝 食 時 所 需 的 警 覺 (aware),活力( active) 以及與攝食時所需的回饋機制的參予。
6.6 內分泌系統(endocrine system)
在 內 分 泌 系 統 有 關 的 研 究 中 , 由 腦 室 內 給 予orexins 會增加黃體生成素 (luteinizing hormoin)的濃度(Pu et al., 1998)以及腎皮質類固醇(corticosterone)而抑制 了泌乳素(prolactin)及生長激素(growth hormone)(Hagan et al., 1999)。除了在中樞影 響內分泌外,進一步的研究發現,腎上腺髓質亦有其受體的存在(Lopez et al., 1999)。在病理研究上,在正常的腎上腺髓質中,OX1R 的mRNA 表現量比OX2R 的mRNA 表現量大,但形成腎上腺髓質瘤( pheochromocytomas)後,便轉變成OX2R 的mRNA 表現量比OX1R 的mRNA 表現量大(Mazzocchi et al., 2001)。
6.7 消化道系統(digestive system)
中樞投與orexin 後,會促進動物胃酸的分泌,一旦將動物施以迷走神經切除 (vagotomy)或投與副交感神經阻斷劑阿托平(atropine),則會使上述orexin 的作用喪 失;若經由周邊給予orexin,則對動物胃酸分泌無影響,因此,暗示orexin 會經由 中樞的迷走神經系統來刺激胃酸分泌(Takahashi et al., 1999)。另外研究發現,在腸 胃道中有orexins 及其受體的存在,且orexins 會顯著刺激腸胃道神經元的活性,及 促進腸胃道的運動(Kirchgessner and Liu, 1999)。
6.8 運動行為(motor activity)
Orexin A 和 orexin B 都會促進動物自發性活動力( spontaneous physical activity)
以及增加一些行為表現上的改變。在動物行為上發現,由腦室給予 orexin 會增加
動物的活動行為(locomotor activity)、跳躍次數及增加警覺(Hagan et al., 1999; Kotz, 2006),此外還發現有 freezing behavior、毛髮整理(grooming)以及咀嚼等動作的增 加(Hagan et al., 1999; Ida et al., 2000; Jones et al., 2001)。若直接將 orexins 微量注入 LC,會促進同側或雙側肢體的肌肉張力(Kiyashchenko et al., 2001)。
6.9 報償系統(reward system)與成癮機制(addiction)
在 回 饋 系 統 與 成 癮 機 制 中 ,orexins 亦被發現有其十分重要的角色。從 hypothalamus來的orexin neurons有大量的神經纖維投射在ventral tegmental area釋 出orexin(Fadel and Deutch, 2002; Peyron et al., 1998),直接活化在ventral tegmental area 上的dopaminergic 神經元(Korotkova et al., 2003; Nakamura et al., 2000)。如本 文6.8,由腦室給予orexin 會有增加動物的活動行為(locomotor activity)、跳躍次數 及增加警覺(Hagan et al., 1999; Kotz, 2006)的效果。但若投予dopamin 受體拮抗 劑 ,haloperidol , 可 以 有 效 抑 制 側 腦 室 注 射 orexins 所 造 成 活 動 行 為 的 增 加 (hyperlocomotion)以及刻板行為(stereotypy) (Nakamura et al., 2000)。如果給予 cocaine 或morphine 引起成癮行為後,給予orexin 可以看到成癮行為或攝食行為 被加強(Boutrel et al., 2005; Harris et al., 2005)。若是給予OX1R 拮抗劑SB 334867,
會使成癮表現降低,推測內生性orexin A 可能藉由活化ventral tegmental area 產生 成癮行為(Borgland et al., 2006; Narita et al., 2006);此外,Borgland 等學者(Borgland et al., 2006)進一步利用電生理記錄的方式,發現給予orexin A 會經由OX1R 活化 PLC/PKC,在大鼠ventral tegmental area 的dopamine neuron 上引發NMDAR 所媒 介的電流外,藉由此一方式,更強化了AMPAR所媒介的電流。推論在ventral tegmental area 這個區域,orexins 可能強化回饋系統和成癮作用。
6.10 學習(learning)與記憶(memory)
在動物行為的實驗中,Telegdy 等學者,首先利用 orexin A 注入大鼠側腦室 (i.c.v.),利用被動逃避試驗(passive avoidance test),發現給予 orexin A 可以促進學 習(learning)和記憶的固化(consolidation),在 560 pmol orexin A 可以促進學習,而 280 pmol orexin A 即具有強化記憶固化的效果,這個增進的效果可能藉由許多受體 活 化 而 達 到 , 利 用 專 一 性 的 受 體 拮 抗 劑 發 現 , 包 括:dopamin 2 receptor 、 α -adrenoreceptor、β- adrenoreceptor、muscarinic cholinergic receptor、GABAA receptor 等 都有參與(Telegdy and Adamik, 2002b)。Jager 等學者同樣利用 orexin A 注入 CD-1 和SMAP 小鼠側腦室後,進行主動和被動逃避試驗,發現 orexin A(3 nmol)可促進 CD-1 主動和被動逃避記憶的固化,並增強 4 個月大 SMAP 小鼠的被動逃避記憶固
化,更可幫助有記憶衰退現象的12 個月大 SMAP 小鼠的被動逃避記憶固化。然而,
在水迷宮試驗(Morris water maze task) 中,於 Aou 等學者於側腦室投予 1~10 nmol orexin A,會使大鼠有記憶衰退的情形。Akbari 等學者,同時利用水迷宮以及被動 逃避試驗,並於CA1 和 DG 區域投予 OX1R 專一性的抑制劑 SB-334867(6 mg/0.5 ml)後,發現在水迷宮試驗中,若阻斷 CA1 的 OX1R 會同時減低大鼠獲取 (acquisition)、固化(consolidation)和提取(retrieval)記憶的能力。但在 DG 卻只會造
成獲取和固化記憶的能力下降。在被動逃避學習試驗中,於CA1 抑制了獲取和固
化記憶的能力,DG 投與的結果僅抑制了提取記憶的能力,暗示內生性的 oresin A 在不同記憶模式中,皆扮演了重要的角色。(Akbari et al., 2008; Akbari et al., 2006;
Akbari et al., 2007)。Wayner 等學者於大鼠 DG 局部投與 orexin A (900 pmol),利用 in vivo 的胞外電生理紀錄模式,觀察到 LTP 有被增強的現象,且此一現象可以被
腹腔投予之OX1R 專一性拮抗劑,SB-334867(10mg/kg) 所完全抑制,因此推論為 OX1R 所媒介的反應(Wayner et al., 2004)。
利用in vitro 胞外紀錄的電生理實驗中,Aou 等學者觀察 orexin A(1~30 nM)對 大鼠 CA1 區域長期增益現象(longterm potentiation, LTP)的影響,發現 orexin A(30nM)對 LTP 有抑制的情形,而不影響長期抑制效應和基礎突觸傳遞。此外,
利用低鎂和AMPAR 拮抗劑觀察投予 orexin A 時 NMDA 電流的變化,發現有下降 的情形,因此認為LTP 的抑制是由於 NMDA 電流的降低而來(Aou et al., 2003)。但 Selbach 等 學 者 給 予 orexin A (100nM) 於 具 有 完 整 CA3-CA1 迴 路 的 小 鼠 hippocampus 腦切片中,卻看到基礎突觸傳遞有增強的趨勢,而且此一趨勢並不會 因為將orexin A 洗去而消失。在將 CA3-CA1 迴路切斷後,這個增強的現象就會消 失,而投予 orexin B (100nM)不會這個現象。他們並認為這個基礎突觸傳遞增強的
趨勢可能是由於CA3 區域的細胞群自發性去極化程度上升所造成,而此一現象可
能是 orexin A 藉由活化局部區域 glutamatergic,noradrenergic,cholinergic 和 gabaergic 細胞所導致。同一篇實驗中,發現用低鎂、AMPA 拮抗劑和 GABAA拮 抗劑存在下,刺激引起的 NMDA 電流,發現 orexin A(100 nM)對 NMDA 電流有抑 制的情形(Selbach et al., 2004)。這個結果與 Aou 等學者發現的相同(Aou et al., 2003)。
然而,我們知道 LHA 的神經元可以將 orexin 投射到 hippocampus 中,且 hippocampus 亦有 orexin 的受體。也有報導指出於活體實驗中,刺激 LHA 會抑制 DG 長期增益現象(LTP)(Wayner et al., 1997a; Wayner et al., 1997b)。因此推論 orexins 有機會抑制 hippocampus 長期增益現象(LTP)的形成,上述實驗的結果,有許多的 不一致。由於目前的解剖證據指出,雖然大鼠和小鼠的hippocampus 皆可發現 orexin 的受體(Cluderay et al., 2002; Marcus et al., 2001; Selbach et al., 2004; Trivedi et al., 1998),但兩個種別間,受體的兩個亞型卻有分佈區域上的不同。在大鼠無論是分 析ORX mRNA 或是蛋白質,都發現 CA1 和 CA2 區域是以 OX1R 為主,OX2R 非 常的少。此外,在CA3 則是以 OX2R 為主,而 DG 則是兩種亞型都有,以 OXR1 較多(Cluderay et al., 2002; Marcus et al., 2001; Trivedi et al., 1998)。但在 C57/BL6 小 鼠 hippocampus,無論是 CA1、CA3 或 DG 上,卻可以同時擁有 OX1R 和 OX2R 的mRNA 和蛋白質的表現(Selbach et al., 2004)。因此,本實驗便選用 C57BL/6J 小 鼠,於 hippocampus CA1 區域進行胞外電生理紀錄,研究 orxcin A 和[Ala11, D-Leu15]-orexin B 對基礎突觸傳遞和神經可塑性的影響,並利用 OX1R 之選擇性拮
抗劑 SB 334867 或 OX2R 之選擇性拮抗劑 compound 29,與 orxcin A 或[Ala11, D-Leu15]-orexin B 同時給藥,希望可以進一步釐清在小鼠和大鼠間的 orexin 受體分 佈差異在基礎突觸傳遞和神經可塑性上,OX1R 和 OX2R 是否有參與且扮演何種 的角色。
實驗材料及方法
1.實驗動物
實驗使用的動物為成年雄性小鼠(C57BL/6J mice),週齡為 8~12 週。小鼠購自
國家實驗動物中心,在台大醫學院實驗動物中心飼養於室溫下,光照週期為12h:12h
光照:黑暗,鼠籠配置食物及飲水,至少飼養一週後開始使用,。
2.海馬回(hippocampus)活體腦切片製備
將小鼠斷頭犧牲後,快速取出腦,置於冰溫且充氧(5% CO2 and 95% O2)的解 剖用人工腦脊髓溶液(dissection artificial cerebral spinal fluid , ACSF),成分組成如下 (in mM): NaCl 87, KCl 2.5, MgCl2 7, CaCl2 0.5, Na2HPO4 1.2, NaHCO3 25, sucrose 75 (pH 7.4)。取出的腦先沿著大腦縱隔的溝壑切成左右兩個腦半球,將腦半球的小腦 (cerebellum)、延髓(medula)、嗅球(olfactory bulb)與前腦(prefrontal cortex)迅速切除,
再將得到體積大約0.5×0.4×0.4 cm3 包含海馬回(hippocampus)的腦區塊,再用快乾 膠 把 腦 塊 的 小 腦 切 除 面 黏 在 切 片 機 標 本 台 上(DTK-1000, DOSAKA, Kyoto, Japan),以 Coronal dissection 方式 ,切出厚度為 400 μm 的海馬回(Hippocampus) 腦切片,切下的海馬回腦切片放置在人工腦脊髓溶液(artificial cerebral spinal fluid, ACSF) 中,室溫下平衡至少一小時。生理 ACSF 成分如下: (in mM) NaCl 117, KCl 4.5, MgCl2 1.2, CaCl2 2.5, Na2HPO4 1.2, NaHCO3 25, Glucose 11 (pH 7.4)。
3.電生理學記錄
3.1 電生理訊號記錄方式
在腦切片的 hippocampus CA1 區域使用胞外紀錄(extracellular recording)的方 式,記錄場興奮性突觸後電位(field excitatory post-synaptic potential,fEPSP)。本實 驗利用多頻道電極記錄系統(Panasonic MED-64 system)(圖一)。記錄槽底面佈有 64 個可選作刺激或記錄的電極與四個參考電極(圖二),這四個參考電極有助於增加訊 號/雜訊比值,以這四個參考電極的平均值對應可擷取到的場興奮性突觸後電位 (fEPSP),可獲取更加清晰且雜訊較小的 fEPSP。所獲得的訊號再經由電腦軟體 (Conductor 3.1)數位化後擷取與分析。64 個可刺激或記錄的電極排列成 8×8 矩陣,
64 個電極覆蓋面積約為 1 mm2 ,每個電極面積約為50 ×50 μm2,每個電極間距 150 μm,在 50Hz 測量下電阻值約為≦50kΩ。含多頻道電極記錄槽在第一次使用 前,浸泡在室溫下PEI 溶液( 1 ml 0.1% polyethylene imine 溶在 25 mM, Borate buffer ) 至少 12 小時,增加記錄槽的電極面的親水性而加強其腦切片之貼附性,
之後用蒸餾水溫和地沖洗至少3 次以上。
依前述方法分離之含腦切片,在室溫下至少平衡一小時後,待海馬體的神經 細胞恢復活性,平衡後,將 hippocampus 腦切片移到紀錄槽,透過連接到電腦的 顯微鏡(OLYMPUS MIC-D)將 hippocampus 區域定位到電極上,用鑲有尼龍網子的 馬蹄形白金棒將hippocampus 薄片固定住。定位後照相存檔再將含有腦切片的記錄 槽安置在接有接上放大器(SU-MED640, Panasonic)的連接器(Connector),透過電腦 軟體(Conductor 3.1)操作實驗,並定位記錄槽,接上灌流裝置後,以 95% O2 / 5% CO2
充氧之ACSF 持續灌流。記錄槽內灌流液體積約為 0.5~0.6 ml,灌流速率維持約在 0.8~1 ml/min,溫度保持在室溫(約 27 ℃),酸鹼度維持在 pH 7.4。因為此套系統刺 激電流傳導,腦片刺激效果與記錄訊號的大小與腦片和記錄電極的貼合度息息相 關,故需待腦切片與記錄槽上電極達穩定貼合(約一小時)後,開始進行電生理紀 錄。依 hippocampus 標本與對應位置選擇在 Schaffer-collateral 刺激 CA1 的 apicak dendric zone, stratum radiatum 紀錄。基礎刺激條件是頻率0.03Hz、0.1 ms,基礎刺激
強度調整在可以產生最大fEPSP 幅度的 40%-50%。fEPSP 的丈量方式為量取 fEPSP 的起始斜率的大小為比較指標。
(圖一) (圖二)
3.1.1 長期增益效應(Long-Term Potentiation,LTP)
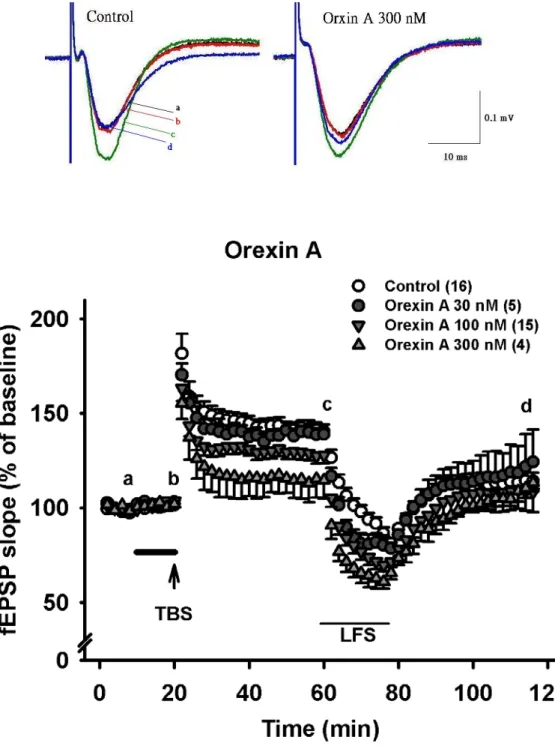
長期增益效應是經由高頻刺激(High Frequency Stimulation, HFS)引起,如前述 在基礎刺激條件,0.03Hz,0.1ms 刺激下得 到穩定的 fEPSP 20 分鐘以上之後,紀錄 十分鐘的fEPSP 之平均值當作 baseline,再接下來加藥十分鐘後,改變刺激條件為 高頻刺激。,高頻刺激的條件為Theta burst stimulation (TBS): 三次相隔 15 秒的 theta 波刺激 ,每次 theta 波刺激由十個相隔 200ms 的 100Hz 波串(train)組成,每個 train 由4 個相隔 10 ms 的脈波組成(圖三)。HFS 後再轉成 0.05Hz 的基礎刺激頻率持續 觀察40 分鐘。長期增益效應(LTP)定義為高頻刺激(TBS)過後轉成基礎刺激頻率所 引發的fEPSP 比未高頻刺激前的 fEPSP 大且可以持續至少 20 分鐘以上。LTP 強度 突觸塑性改變程度計算,是 HFS 後 40 分鐘每一腦切片的 fEPSP 斜率增加程度。
本篇中n 值為使用之腦片數量,每隻小鼠約使用 1~2 片。
(圖三)
3.1.2 去增益效應(Depotentiation)
在 TBS 四十分鐘後,去增益現象是由給予 1Hz 的低頻刺激(Low frequency Stimulation, LFS)15 分鐘所引發。在 LFS 之後,再轉成前述的 0.33Hz 基礎刺激頻 率下觀察40 分鐘。LFS 後原本因 TBS 產生 LTP 可以被 LFS 削弱稱為去增益效應。
削弱的程度以LFS 後 40 分鐘腦切片的 fEPSP 斜率與 LTP 時 fEPSP 斜率相減後之 值,再以LTP 的增大程度為 100%,計算而得。
3.1.3 強直性後增效應 Post-Tetanic Potentiation (PTP)
緊接在高頻刺激 (High-Frequency Stimulation, HFS) 後的幾分鐘內,突觸後神 經元對原本的基礎刺激(0.03Hz)產生較強直刺激前大很多的 fEPSP ,這樣的現象就 是強直性後增效應(Post-Tetanic Potentiation, PTP)這是因為突觸前末稍聚積鈣離 子(Korogod et al., 2005)。PTP 與 LTP 的差異是,PTP 評估 HFS 後短期內的神經傳
遞加強現象,大量鈣離子在HFS 後蓄積神經末梢內,造成神經傳遞增加,可以用
來評估藥物對突觸前影響神經傳遞可塑性的作用。測量PTP 的方法是觀察 HFS 後
四分鐘內的fEPSP 比未 PTP 前增加的程度。
3.1.4 Paired-pluse facilitation (PPF)
為了進一步評估藥物對突觸前訊息傳遞物釋放之影響,本篇利用一對兩個快 速的刺激,第一個刺激和第二個刺激之間隔時間為50 ms(Aou et al., 2003; Lauri et al., 2007)。將第二個刺激所得之 fEPSP 斜率與第一個所得之斜率相除,而得一比值,
稱之為 Paired pluse ratio (PPR)。一對電刺激給予的時間點分別是在加藥前,加藥10 分鐘後,高頻刺激後40 分鐘以及低頻刺激後 40 分鐘後,並計算其 PPR。
4. 藥品來源
本實驗所用的藥品為orexin A、[Ala11,D-Leu15] orexin-B、 SB-334867 皆購自 Tocris ( Bristol, UK )。Compound 29 則感謝美國國家藥物濫用研究院化學生物研究 組(Chemical biology research branch, National insitute on Drug Abuse)的 Dr. Kenner C.
Rice 慷慨捐贈。藥品除了 SB-334867 溶於 DMSO 之外,其餘藥品都是使用二次 水溶解,泡製成實驗使用濃度1000 倍的 stock solution,每 30 ml 成一管,儲存於
-20 ℃,實驗時均取 stock solution 30 μl 加入 30ml 的 ACSF 中稀釋為實驗所需之 濃度。
5. 資料分析
在ACSF 持續灌流下由前述基礎刺激條件引發的 fEPSP slope 達 steady state 後 取加藥前十分鐘的fEPSP 斜率視為 baseline fEPSP slope,其平均值則視作 100%。
評估加藥物對basal synaptic transmission 影響,是觀察加藥後達 steady state 後 10 分鐘內的fEPSP slope 與未加藥的 baseline fEPSP slope 比較改變程度。評估 LTP 與 depotentiation 程度是分別觀察高頻刺激和低頻刺激後四十分鐘 fEPSP 斜率 ,與未 加藥的baseline fEPSP slope 比較。評估 PTP 則是在測量 HFS 後四分鐘內與 baseline fEPSP slop 相比的 fEPSP slope。
Time course 圖形表示每點是由每兩分鐘的 fEPSP slope 平均一次,即由四個 fEPSP 斜率平均之,標準化後用百分比加上 standard error bar 表示,並標註 N 值,
指的是腦切片數目,統計方法使用paired Student t-test 於同一片腦片間的比較,而 unpaired Student t-test 則於不同濃度的組別間比較,P 值小於 0.05 視為有統計上差 異,標以* ,若 P 值小於 0.01,則標以** 做區分。
實驗結果
本 實 驗 之 目 的 擬 觀 察 orexins 是 否 會 對 小 鼠 hippocampus Schaffer collateral/commissural-CA1 間突觸之長期增益現象(LTP)以及高頻刺激後去增益現 象(depotentiaion)造成影響。在腦片穩定記錄到 0.03 Hz 基礎刺激誘發的 fEPSP 後,
灌流orexin A(30, 100, 300nM)或 [Ala11, D-Leu15]-orexin B (3, 10, 30, 100, 300, 1000 nM)於小鼠 hippocampus 腦薄切片 10 分鐘,再換成 ACSF,並給予高頻率刺激(TBS) 後,刺激頻率轉回0.03Hz,觀察 fEPSP 變化來評估 40 分鐘後的 LTP 程度,再給 予低頻率刺激(LFS),之後再轉回 0.03Hz,觀察 fEPSP 變化 40 分鐘,來評估 depotentiation 的程度。藉由上述方式,我們希望可以觀察 CA1 區域之神經突觸可 塑性是否受到 orexins 的影響。此外,本實驗亦利用 OX1R 之選擇性拮抗劑 SB 334867 或 OX2R 之選擇性拮抗劑 Compound 29 分別與 orexin A 和 [Ala11, D-Leu15]-orexin B 一併灌流十分鐘,希望進一步釐清 orexins 在 hippocampus CA1 區域的作用所媒介的OXR 亞型。
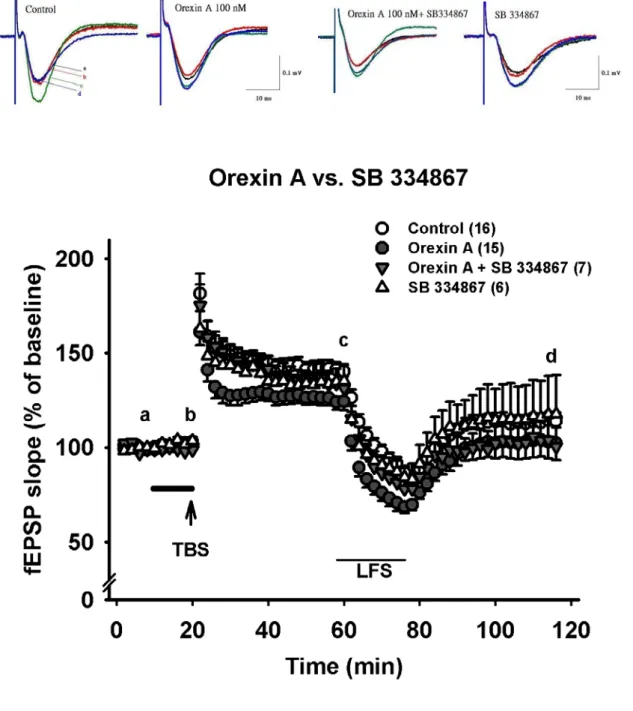
Orexin A 有意義的抑制長期增益現象(LTP)的程度
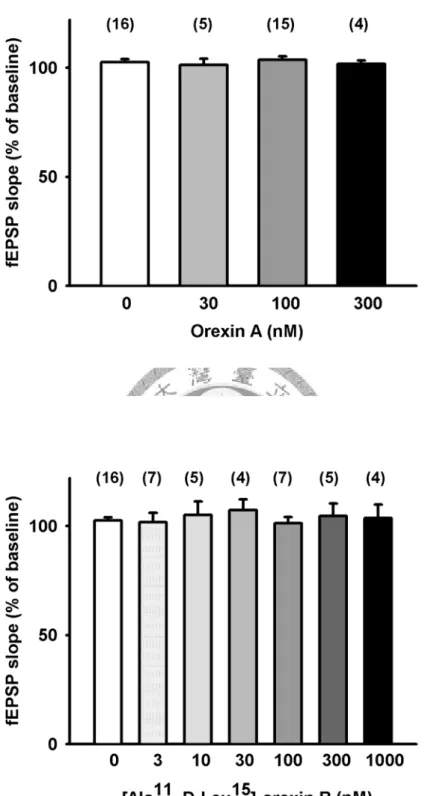
在腦片穩定記錄10 分鐘後,灌流 orexin A (30, 100,300 nM) 10 分鐘,發現基 礎突觸傳導並無顯著的變化(orexin A 30, 100, 300 nM baseline 分別為: 101.9±0.9 %, n=5; 99±0.7 %, n=15; 100.3±0.3 %, n=4,drug application 後分別為: 101.3±2.7 %, n=5;
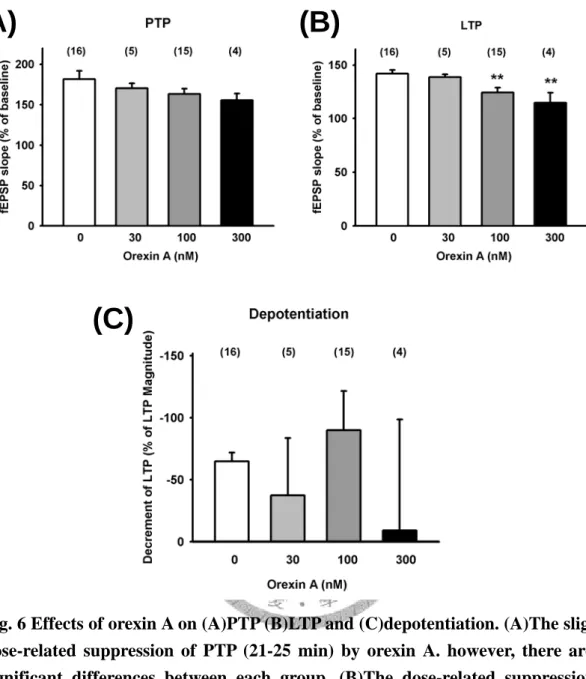
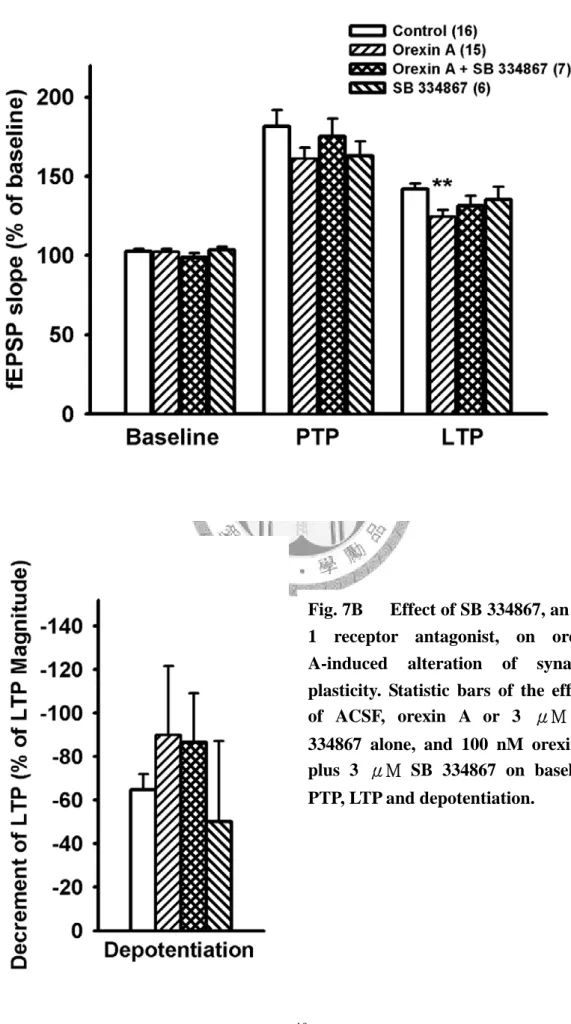
102.5±1.7 %, n=15; 101.8±1.6 %, n=4 )(Fig. 4A)。在高頻刺激後的後強直增強效應 (PTP),有觀察到隨著 orexin A 濃度上升而使得 PTP 有減弱的趨勢,雖無顯著的統 計差異(control: 181.5±10.4 %, n=16; orexin A 30, 100, 300 nM 分別為: 170.3±5.9%, n=5; 161.1±7 %, n=15; 155.3±8.5 %, n=4)(Fig. 5, Fig. 6A)。繼續觀察高頻刺激(TBS) 後40 分鐘,我們發現 orexin A 濃度相依的抑制長期增益(LTP),在 100 nM 和 300 nM 達到有意義的抑制效果(control: 141.9±3.5%, n=16; orexin A 30, 100, 300 nM 分 別為: 138.8±2.5%, n=5; 124.4±4.5%, n=15, p<0.01; 114.6±9.5%, n=4, p<0.01) (Fig 5, Fig 6B)。在去增益現象(depotentiaiton)上,各組間並無統計上顯著差異 (control:
-64.8±7 %, n=16; orexin A 30, 100, 300 nM 分別為: -37.3±46.1%, n=5; -90±31.6%, n=15; -9±31.6 %, n=4) (Fig. 5, Fig. 6C)。
Orexin A 對突觸前神經傳遞物釋放有輕微抑制的傾向
由於PTP 可能與突觸前末稍聚積鈣離子有關(Delaney and Tank, 1994)。因此我 們更進一步的利用paired pluse faciliation 的現象來分析突觸前神經傳導物釋放的變 化程度,並以paired pluse ratio (PPR)來表示之,希望能更進一步釐清我們在 PTP 所觀察到的抑制趨勢。我們在orexin A (100 nM 和 300 nM) 灌流前,灌流 10 分鐘 後,高頻刺激後40 分鐘以及低頻刺激後 40 分鐘分別給予 paired pluse stimulation,
計算其PPR。在基礎刺激頻率(0.03 Hz)下灌流 orexin A 100 或 300 nM 十分鐘後,
並無明顯影響basal stimulation 下的 PPR。在 orexin A 300 nM 灌流標本,在 LTP 時 的PPR 似乎有比 baseline 時的 PPR 降低的趨勢,不過統計上並無意義(1.49±0.04 vs.
1.57±0.07, n=4 )(Tab. 2)。若比較 PPR 在 LTP 與 baseline 時變化的比值,在未加藥 的腦薄片,LTP 的比值為 0.938,暗示此條件刺激在小鼠 hippocampus CA1 區域所 引起的LTP 時有突觸前訊息傳遞物釋放會增加的可能。在高頻刺激(TBS)後 40 分 鐘,orexin A 100 nM 和 300 nM 皆可反轉控制組之 PPR 的下降程度(control:
0.93±0.02, n=8; orexin A 100 nM: 0.98±0.03, n=5; orexin A 300 nM: 0.94±0.02, n=4)。
在低頻刺激(LFS)後 40 分鐘,orexin A 100nM 和 300 nM 亦可輕微反轉控制組 PPR 減少的程度(control: 0.94±0.02, n=8; orexin A 100 nM: 1.01±0.04, n=5; orexin A 300 nM: 0.99±0.05, n=4)。但這些變化並無達到統計上顯著的意義,亦無與 orexin A 濃 度上升有相關性(Tab.2)。
OrexinR 拮抗劑對基礎突觸傳導和突觸可塑性之影響
根據上述之結果,我們可以肯定的是orexin A 會抑制長期增益現象(LTP)的程 度。為了瞭解orexin A 是藉由何種受體作用而影響 LTP。因此我們分別灌流 OX1R 之選擇性拮抗劑SB 334867 和 OX2R 之選擇性拮抗劑 Compound 29 來觀察他們對
orexin A 所造成的抑制是否有拮抗的效果。我們先分別單獨灌流此兩種拮抗劑 10 分鐘後,觀察他們對基礎突觸傳導,強直後增效應(PTP),長期增益現象(LTP)和去 增益現象(depotentiaion)是否有造成影響。
OX1R 拮抗劑 SB 334867 對基礎突觸傳導和突觸可塑性並無影響
在基礎刺激頻率(0.03 Hz)下灌流 3 μM SB 334867 十分鐘後,並沒有發現顯著 影響基礎突觸傳導的現象,fEPSP slpoe 加藥後為加藥前的 103.5±2.1 % (n=8) (Fig.
7A△ , Fig. 7B)。在高頻刺激後的強直後增效應(PTP)上,SB 334867 3 μM 亦無顯 著影響(control: 181.5±10.4 %, n=16; 3 μM SB 334867: 162.9±9.2 %, n=8) )(Fig. 7)。
在高頻刺激(TBS)後 40 分鐘,3 μM SB 334867 不影響長期增益現象(LTP) (control:
141.9±3.5%, n=16; 3 μM SB 334867: 135.3±8.2%, n=8) ) (Fig. 7A, △ Fig.
7B )。在低頻刺激(LFS)後 40 分鐘, 3 μM SB 334867 也沒有影響去增益現象 (depotentiaion) 的情形(control: -64.8±7 %, n=16; 3 μM SB 334867 : -50.2±36.9 %, n=8) (Fig. 7A, △ Fig. 7B )。
OX1R 拮抗劑 SB 334867 對 orexin A 造成之抑制長期增益現象有輕微拮抗的效果 若同時灌流orexin A 100 nM 和 3μM SB 334867 十分鐘後,基礎突觸傳導並無 顯著的變化,fEPSP slpoe 加藥後為加藥前的 98.7±2.9%(n=6) (Fig. 7A▼, Fig.
7B )。Orexin A 對高頻刺激後的強直後增效應(PTP)的輕微抑制傾向,在這裡也 會受到SB 334867 些許的反轉,但未達統計上的差異(control: 181.5±10.4 %, n=16;
orexin A 100 nM: 161.1±6.5 %, n=15; orexin A 100nM + 3μM SB 334867:
170.6±11.9%, n=7 ) (Fig. 7A▼, Fig. 7B )。然而,與 orexin A 100 nM 抑制 LTP 的程度比較,同時灌流orexin A 100 nM 和 3μM SB 334867 可以得到輕微的拮抗效 果(control: 141.9±3.5%, n=16; orexin A 100 nM: 124.4±4.5%, n=15; orexin A 100nM + 3μM SB 334867: 133.86±6.6%, n=7) (Fig. 7A▼, Fig. 7B ),但未達統計意義。在 低頻刺激(LFS)後 40 分鐘,orexin A 100 nM 和 3μM SB 334867 不影響高頻刺激後
去增益現象(depotentiaion) 的表現的程度(control: -64.8±0.07 %, n=16; orexin A 100nM: -90±31.6 %, n=15; orexin A 100nM + 3μM SB 334867: -86.6±22.4%, n=6) (Fig. 7A▼, Fig. 7B )。
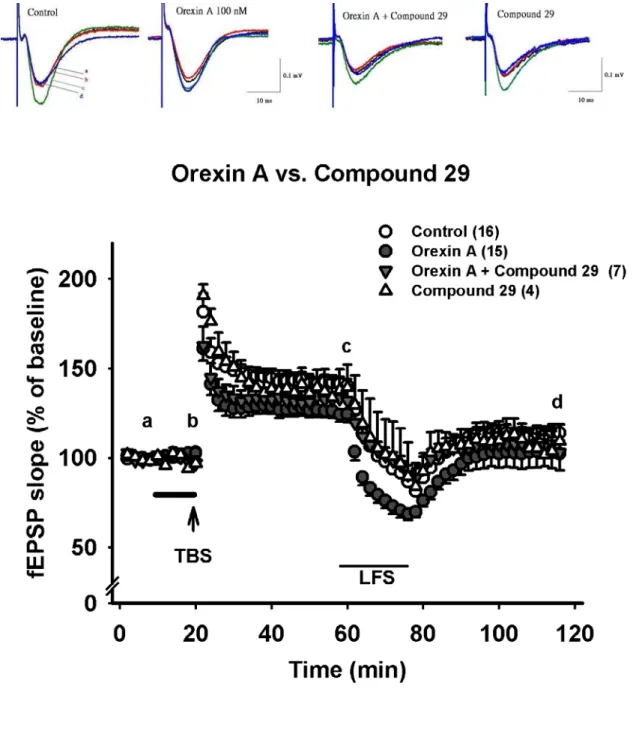
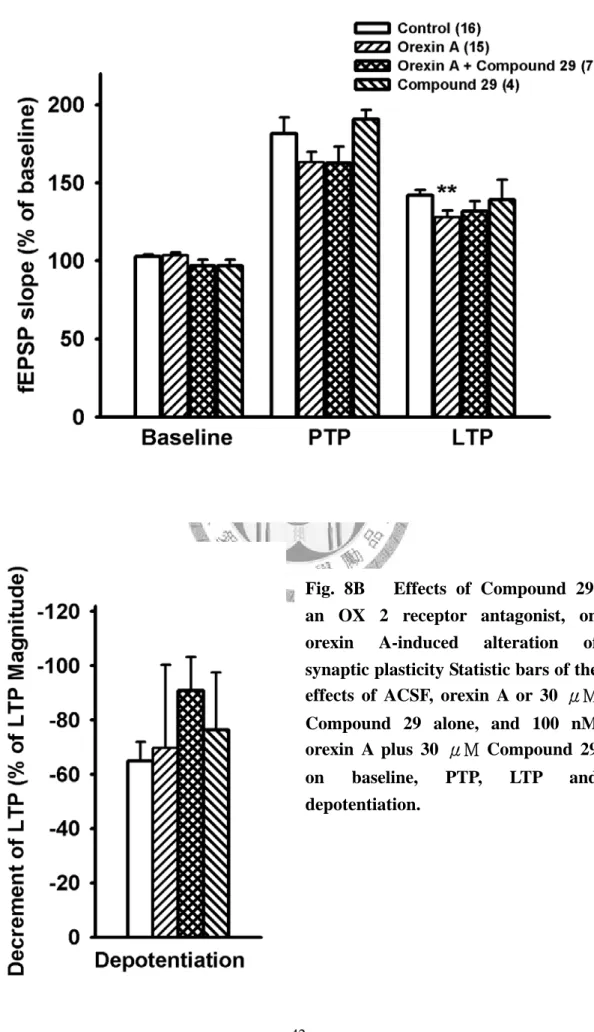
OX2R 拮抗劑 Compound 29 無顯著抑制基礎突觸傳導的現象
在基礎刺激頻率(0.03 Hz)下灌流 Compound 29 30 μM 十分鐘後,發現 30 μM Compound 29 有輕微抑制影響基礎突觸傳導的現象,但不具統計上的差異,fEPSP slpoe 加藥後為加藥前的 96.7±3.8 % (n=4 )(Fig. 8A △, Fig. 8B )。在高頻刺 激 後 的 強 直 後 增 效 應(PTP)上,30 μM Compound 29 並無顯著影響(control:
181.5±10.4 %, n=16; 30 μM Compound 29: 190.7±6.1 %, n=4) (Fig. 8A △, Fig.
8B )。在高頻刺激(TBS)後 40 分鐘,30 μM Compound 29 亦沒有增強長期增 益現象(LTP)的效果(control: 141.9±3.5%, n=16; Compound 29 30 μM: 139.1±12.9 %, n=4) (Fig. 8A △, Fig. 8B )。在低頻刺激(LFS)後 40 分鐘,於高頻刺激時灌流 的 30 μM Compound 29 沒有改變去增益現象(depotentiaion) 的程度(control:
-64.8±0.02 %, n=16; 30 μM Compound 29: -76.3±21.1 %, n=4) (Fig. 8A △, Fig.
8B )。
OX2R 拮抗劑 Compound 29 對 orexin A 造成之抑制長期增益現象亦有輕微拮抗的 效果
同時灌流100 nM orexin A 和 30 μM Compound 29 十分鐘後,基礎突觸傳導有 些許下降的現象,且程度與單獨灌流30 μM Compound 29 相近,fEPSP slope 加藥 後為加藥前的96.8±3.9%( n=7 ) (Fig. 8A▼, Fig. 8B ),但未達統計差異。高頻刺 激後的強直後增效應(PTP)灌流 100 nM orexin A 和 30 μM Compound 29 並不顯著影 響其程度(control: 181.5±10.4 %, n=16; orexin A 100 nM: 161.8±7 %, n=15; orexin A 100nM + Compound 29 30 μM: 162.6±10.6%, n=7 ) (Fig. 8A▼, Fig. 8B )。當與 orexin A 100 nM 對 LTP 的抑制程度比較時,同時灌流 orexin A 100 nM 和 30 μM
Compound 29 可以得到輕微的拮抗效果(control: 141.9±3.5%, n=16; orexin A 100 nM:
124.4±4.5%, n=15; orexin A 100nM + 30 μM Compound 29: 131.7±6.4%, n=7 ) (Fig.
8A▼, Fig. 8B ),但統計上並沒有差異。在低頻刺激(LFS)後 40 分鐘,於高頻 刺激時灌流的 orexin A 100 nM 和 30 μM Compound 29 亦無影響去增益現象 (depotentiaion) 的 表 現 的 程 度 (control: -64.8±0.02%, n=16; orexin A 100nM:
-90±31.6%, n=15; orexin A 100nM + 30 μM Compound 29: -90.8±12.4%, n=8) (Fig.
8A▼, Fig. 8B )。
[Ala11, D-Leu15]-orexin B 有意義的抑制長期增益現象
在基礎刺激頻率(0.03Hz)穩定記錄十分鐘後,分別灌流 3, 10, 30, 100, 300, 1000 nM [Ala11, D-Leu15]-orexin B 十分鐘後,基礎突觸傳導並沒有顯著的變化,與 baseline 相比,[Ala11, D-Leu15]-orexin B 3, 10, 30, 100, 300, 1000 nM 分別為:
101.8±4.2%, n=7; 105.1±6.1%, n=5; 107.3±4.8%, n=4; 101.3±2.8%, n=7; 300 nM:104.6±5.5 %, n=5; 103.6±6 %, n=4) (Fig.4B, Fig. 9)。在高頻刺激後的強直後增效 應(PTP)中,並無顯著統計差異(control: 181.5±10.4 %, n=16; [Ala11, D-Leu15]-orexin B 3, 10, 30, 100, 300, 1000 nM 分別為:: 148.7±5.3%, n=7; 146.3±9.7%, n=5;
159.7±13.4 %, n=4; 167.8±8.3 %, n=7; 146.8±8.7 %, n=5; 161.4±14.9 %, n=4) (Fig. 9, Fig 10A)。再繼續觀察高頻刺激(TBS)後 40 分鐘,長期增益現象(LTP)的程度,發 現 [Ala11, D-Leu15]-orexin B 30, 300 和 1000 nM 可以有意義的抑制長期增益(LTP) 的現象,但其抑制的程度 不完全與[Ala11, D-Leu15]-orexin B 濃度的上升為增加,表 示在 30 nM 或 300 nM 抑制已達飽和,且很快就到達最大的抑制效果(control:
141.9±3.5%, n=16; [Ala11, D-Leu15]-orexin B 3, 10, 30, 100, 300, 1000 nM 分別為:
130.3±7.9%, n=7; 130.4±6%, n=5; 123.3±3.8%, n=4, p<0.05; 131.2±2.9%, n=7;
120.8±7.2%, n=5, p<0.05; 123.4±4.6%, n=4, p<0.05) (Fig. 9, Fig 10B)。在去增益現象 (depotentiaiton)上,各組間沒有統計上顯著的差異存在。(control: -64.8±7%, n=16;
[Ala11, D-Leu15]-orexin B 3, 10, 30, 100, 300, 1000 nM 分別為: -60±29.6%, n=7;
-40.2±16.1%, n=5; -48.8±30.9%, n=4; -36.2±14.4%, n=7; -104.9±44.8%, n=5;
-88.9±50.5 %, n=4) (Fig. 9, Fig 10C)。
OX1R 拮抗劑 SB 334867 對[Ala11, D-Leu15]-orexin B 造成之抑制長期增益現象可觀 察到輕微拮抗的現象
同時灌流[Ala11, D-Leu15]-orexin B 300 nM 和 3μM SB 334867 十分鐘後,基礎 突觸傳導無顯著的變化,fEPSP slope 加藥後為加藥前的 99±4.7% (n=7) (Fig. 11A▼, Fig. 11B ) 。 高 頻 刺 激 後 的 強 直 後 增 效 應 (PTP) 亦 無 明 顯 的 變 化 (control:
181.5±10.4%, n=16; [Ala11, D-Leu15]-orexin B 300 nM:146.8±8.7%, n=5; [Ala11, D-Leu15]-orexin B 300 nM + 3μM SB 334867: 156.5±10.3%, n=7 ) (Fig. 11A▼, Fig.
11B )。然而,與[Ala11, D-Leu15]-orexin B 300 nM 抑制 LTP 的程度相較,若一 起灌流[Ala11, D-Leu15]-orexin B 300 nM 和 3μM SB 334867 可以得到輕微的拮抗效 果(control: 141.9±3.5%, n=16; [Ala11, D-Leu15]-orexin B 300 nM:120.8±7.2%, n=5;
[Ala11, D-Leu15]-orexin B 300 nM + 3μM SB 334867: 130.2±9.5%, n=7) (Fig. 11A▼, Fig. 11B ),但未達統計差異。在低頻刺激(LFS)後 40 分鐘,[Ala11, D-Leu15]-orexin B 300 nM 和 3μM SB 334867 亦不影響去增益現象(depotentiaion) 表現的程度 (control: -64.8±2.9%, n=16; [Ala11, D-Leu15]-orexin B 300 nM: -104.9±44.8%, n=5;
[Ala11, D-Leu15]-orexin B 300 nM + SB 334867 3 μM: -49.9±23.1%, n=7) (Fig. 11A▼, Fig. 11B )。
OX2R 拮抗劑 Compound 29 對[Ala11, D-Leu15]-orexin B 造成之抑制長期增益現象 亦可發現有輕微拮抗的能力
[Ala11, D-Leu15]-orexin B 300 nM 和 30 μM Compound 29 同時灌流十分鐘,基 礎突觸傳導沒有顯著的差異,fEPSP slope 加藥後為加藥前的 99±4.7%(n=5) (Fig.
12A▼, Fig. 12B )。高頻刺激後的強直後增效應(PTP)也沒有明顯的變化(control:
181.5±10.4%, n=16; [Ala11, D-Leu15]-orexin B 300 nM:146.8±8.7%, n=5; [Ala11,
D-Leu15]-orexin B 300 nM + 30 μM Compound 29: 186.5±28%, n=5 ) (Fig. 12A▼, Fig.
12B )。與[Ala11, D-Leu15]-orexin B 300 nM 抑制 LTP 的程度比較,灌流[Ala11, D-Leu15]-orexin B 300 nM 和 30 μM Compound 29 可以得到輕微的拮抗效果(control:
141.9±3.5%, n=16; [Ala11, D-Leu15]-orexin B 300 nM:120.8±7.2%, n=5; [Ala11, D-Leu15]-orexin 300 nM + 30 μM Compound 29: 132.9±7.2%, n=5) (Fig. 12A▼, Fig.
12B ),但未達到統計上的差異。於低頻刺激(LFS)後 40 分鐘, [Ala11, D-Leu15]-orexin B 300 nM 和 30 μM Compound 29 並 不 影 響 去 增 益 現 象 (depotentiaion) 表現的程度(control: -64.8±2.9%, n=16; [Ala11, D-Leu15]-orexin B 300 nM: -104.9±44.8%, n=5; [Ala11, D-Leu15]-orexin B 300 nM + 30 μM Compound 29:
-97.8±33.8%, n=5) (Fig. 12A▼, Fig. 12B )。
討論
在本實驗中,利用 in vitro 胞外紀錄的方式,探討 orexins 對小鼠 hippocampal Schaffer collateral-CA1 的突觸傳導以及突觸塑性之影響。實驗結果顯示(1) orexin A 會抑制長期增益現象(LTP)。(2) [Ala11, D-Leu15] orexin-B 亦會抑制長期增益現象 (LTP)。(3) Orexin A 和[Ala11, D-Leu15] orexin-B 所造成的抑制程度會輕微因 OX1R 拮抗劑SB 334867 和 OX2R 拮抗劑 compound 29 的投予而減輕。(4) [Ala11, D-Leu15] orexin-B 的抑制效果有限,而 orexin A 可達較大的抑制效價(efficiency)。(5)無論是 orexin A 或[Ala11, D-Leu15] orexin-B 都不會影響 depotentiaion 的程度。
許多解剖上的證據顯示,在LHA 中生成 orexin 的神經元,會將含其有 orexin 的末稍深入hippocampus(Chen et al., 1999; Moriguchi et al., 1999; Nambu et al., 1999b),而目前的證據亦指出無論是大鼠或是小鼠的 hippocampus 皆可發現 orexin 的受體(Cluderay et al., 2002; Marcus et al., 2001; Selbach et al., 2004; Trivedi et al., 1998)。但兩個種別間,受體的兩個亞型卻有分佈區域上的不同。在大鼠無論是分 析ORX mRNA 或是蛋白質,都發現 CA1 和 CA2 區域是以 OX1R 為主,OX2R 非 常的少。此外,在CA3 則是以 OX2R 為主,而 DG 則是兩種亞型都有,以 OXR1 較多(Cluderay et al., 2002; Marcus et al., 2001; Trivedi et al., 1998)。但在 C57/BL6 小 鼠 hippocampus,無論是 CA1、CA3 或 DG 上,卻可以同時擁有 OX1R 和 OX2R 的 mRNA 和蛋白質的表現(Selbach et al., 2004)。因此 orexins 在大鼠和小鼠 hippocampus CA1 對神經突觸可塑性的影響可能與 OXR 亞型扮演的角色可能有所 不同。以下將分成幾點討論本實驗中利用orexins 在小鼠 hippocampus CA1 區域基 礎突觸傳遞,長期增益現象(LTP)所造成的影響。
1. Orexins 對基礎突觸傳導的影響
在 我 們 的 實 驗 中 , 並 沒 有 發 現 orexin A(30~100 nM) 和 [Ala11, D-Leu15] orexin-B(3~1000 nM) 可以有意義的增加 CA1 區域基礎突觸傳導(Fig. 4)。而之前進 行orexins 對基礎突觸傳導影響進行研究的報導亦指出,orexin A (30nM)並不會顯 著的改變大鼠CA1 區域基礎突觸傳導(Aou et al., 2003)。但在小鼠的腦片標本上,
orexin A(100 nM) 被 發 現 可 藉 由 興 奮 CA3 上 游 的 局 部 區 域 glutamatergic , noradrenergic,cholinergic 和 gabaergic 細胞,進而使得 CA1 基礎突觸傳導有增加 的趨勢,但orexin B 則否(Selbach et al., 2004)。推測實驗結果差異的可能是由於不 同切面,迴路完整性不同所致。因為本篇所使用獲得的腦薄片的方法為利用 corornal dissection 所得,而在發現有會增加基礎突觸傳導的報導中則是使用 horizotal dissection 所取得(Selbach et al., 2004)。根據同一篇的報導(Selbach et al., 2004)指出,若將 CA3-CA1 之間的連接切斷,這種 orexin A 增進基礎突觸傳導就會 消失。為此我們便回溯本實驗所使用的腦薄片後發現,我們所採用的腦薄片皆偏 尾部區域Ammon’s horn 較大的腦薄片。由於海馬體結構上的限制,使得 CA3-CA1 兩個區域的連結在製備過程受到傷害而不甚完整,因此在這裡我們無法觀察到相 同的反應。
2. Orexins 抑制長期增益現象(LTP)
在本篇利用 C57BL/6j 的小鼠 hippocampus CA1 區域發現,灌流 orexin A (100 nM)發現有抑制 LTP 的現象(Fig.6)。與此實驗結果相仿的,Aou 等學者利用大 鼠於hippocampus CA1 區域紀錄長期增益現象(LTP),在給予 orexin A (30~100 nM) 後,亦發現有抑制的情形(Aou et al., 2003)。此外,我們更進一步的發現 orexin A (100 nM)的抑制效果可被 OX1R 專一性拮抗劑 SB 334867(3 μM)和 OX2R 專一性拮抗劑 compound 29 (30 μM)所減輕,因此認為此抑制的效果可能同時經由 OX1R 和 OX2R 而達成。這個結果與C57/BL6 小鼠 hippocampus,在 CA1 上同時擁有 OX1R 和 OX2R 的mRNA 和蛋白質的表現(Selbach et al., 2004)相呼應。