行政院國家科學委員會專題研究計畫 期中進度報告
猴病毒四十型 T/t-common 基因用於治療 HER-2/neu 大量表
現的癌症的評估(2/3)
計畫類別: 個別型計畫 計畫編號: NSC92-3112-B-002-008- 執行期間: 92 年 05 月 01 日至 93 年 04 月 30 日 執行單位: 國立臺灣大學醫學院免疫學研究所 計畫主持人: 王萬波 報告類型: 完整報告 報告附件: 出席國際會議研究心得報告及發表論文 處理方式: 本計畫可公開查詢中 華 民 國 93 年 4 月 30 日
Abstract
In the previous report, we have demonstrated that T/t-common could specifically induce apoptosis in HER-2/neu-overexpressing cancer cells but not in HER-2/neu low-expressing cancer cells and normal cells. To investigate whether this is the general property of T/t-common, for the past year, we included more
HER-2/neu-overexpressing and HER-2/neu low-expressing cancer cells and normal cells in our study. The results confirm our previous conclusion and indicate that T/t-common may be developed as a selective therapeutic agent against
HER-2/neu-overexpressing cancers. We also demonstrated that T/t-common induced apoptosis of HER-2/neu-overexpressing cancer cells by activating the caspase activity and that the ability of T/t-common to induce apoptosis could be inhibited by Bcl-2. As regard to the mechanism underlying T/t-common induction of apoptosis, we found that T/t-common induced apoptosis in HER-2/neu-overexpressing cancer cells primarily by repressing HER-2/neu expression. Finally we demonstrated that T/t-common could sensitize HER-2/neu-overexpressing cancer cells to anti-cancer drug cisplatin. We also showed that T/t-common could inhibit angiogenesis induced HER-2/neu-overexpressing cancer cells. Taken together, these data suggest that T/t-common may be developed as an efficient anti-cancer gene therapy agent. Keyword: HER-2/neu, SV40, T/t-common, Apoptosis, Angiogenesis
中文摘要
我們先前的報告證明了猴病毒 40 型大 T 抗原和小 t 抗原胺基端共同片段 ( T/t-commom polypeptide )會特異性地誘導大量表現 HER-2/neu 基因的癌細胞凋 亡, 但不會誘導其他癌細胞及正常細胞凋亡。過去一年我們用更多的細胞株證明 這個論點。此外我門證實 T/t-common 可經由活化 caspase 來誘導大量表現
HER-2/neu 基因的癌細胞凋亡,而且 T/t-common 誘導細胞凋亡的能力會被 Bcl-2 抑制。有關 T/t-common 誘導細胞凋亡的機轉,我門證實 T/t-common 可經由抑制 HER-2/neu 來誘導大量表現 HER-2/neu 基因的癌細胞凋亡,最後我們證實
T/t-common 可讓大量表現 HER-2/neu 基因的癌細胞更容易被化療藥物 cisplatin 殺死,且 T/t-common 可抑制大量表現 HER-2/neu 基因的癌細胞去誘發新生血管 生成。綜合以上結果,我門認為 T/t-common 有發展成抗癌基因試劑的潛力。
關鍵詞:HER-2/neu, 猴病毒 40 型, T/t-common, 細胞凋亡, 新生血管生成
Introduction
HER-2/neu is an epidermal growth factor receptor-related transmembrane protein
Yamamoto et al., 1986). The HER-2/neu receptor contains an extracellular domain, a transmembrane domain, and an intracellular tyrosine kinase domain.
Identification of the oncogenic HER-2/neu oncogene and the oncogenic property of the HER-2/neu were originally demonstrated in the rat neu oncogene that was isolated from rat neuroblastoma (Bargamann et al. 1986; Hung et al. 1986). The activated rat neu oncogene contains a point mutation in the transmembrane domain of its protein, which results in a constitutively active tyrosine kinase (Bargmann et al. 1986; Hung et al. 1989). This increase in HER-2/neu tyrosine kinase activity can also be found in many human cancers that overexpress HER-2/neu. In human, the normal HER-2/neu gene is frequently overexpressed or amplified without mutation in many types of cancer, including cancers of the breast, ovary, lung, kidney, colon, bladder, esophagus, stomach, mouth and salivary gland (reviewed by Hynes and Stern, 1994). Enhanced expression of the HER-2/neu gene is correlated with chemotherapeutic drug resistance in some cancer cells (Tsai et al., 1993; Tsai et al. 1994; Ueno et al. 1997; Yu et al. 1998; Hynes and Stern, 1994), enhanced metastatic potential in human breast, ovarian, and non-small-cell lung carcinoma cells (Slamon et al., 1987; Tan et al. 1997; Yu et al. 1993; Hynes and Stern, 1994)
and shortened survival in breast, ovarian, and prostate cancer patients (Slamon et al., 1987; Slamon et al., 1989; Berchuck et al., 1990; Sadasivan et al., 1993; Hynes and Stern, 1994). Transgenic mice carrying mutation-activated HER-2/neu or
overexpressing normal HER-2/neu can induce mammary adenocarcinoma with a
high frequency of metastasis (Muller et al., 1988; Guy et al., 1992; Hynes and Stern, 1994). Enhanced expression of the HER-2/neu gene can also increase experimental metastasis in human lung cancer cells (Yu et al., 1994). Taken together, these results indicate that HER-2/neu is a potent oncogene, and its overexpression plays a crucial role in the development of human malignancies. These results also suggest that suppression of HER-2/neu expression may lead to the treatment of the HER-2/neu-overexpressing human cancers. In fact, inhibition of HER-2/neu activity by various means, such as by anti-HER-2/neu antibodies, have been
demonstrated to be an effective way to treat HER-2/neu-overexpressing cancers (Yu and Hung, 2000).
Two viral oncoproteins, adenovirus 5 E1A and simian virus 40 (SV40) large T, have been reported to repress transcription from the HER-2/neu promoter and consequently repress the transforming activity of HER-2/neu (Yu et al., 1990; Yu et al., 1991; Matin and Hung, 1993; Hung et al., 1995; Chen et al., 1997). When E1A is stably transfected into cells, it can repress HER-2/neu overexpression and
suppress the tumorigenic and metastatic potential of activated
cancer cells that overexpress HER-2/neu (Yu et al., 1991; Yu et al., 1992; Yu et al., 1993; Yu et al., 1995). Moreover, in mice bearing HER-2/neu-overexpressing ovarian, lung, or breast cancer cells, E1A delivered either by adenovirus or liposome significantly inhibits tumor growth and prolongs mouse survival (Hung et
al., 1995; Yu et al., 1995; Zhang et al., 1995; Chang et al., 1996; Chang et al., 1997). These results strongly suggest that by repressing HER-2/neu expression, it is
possible to achieve therapeutic effects on cancers that overexpress HER-2/neu. In fact, a clinical trial of E1A gene therapy in HER-2/neu-overexpressing breast and ovarian cancer has been conducted and promising results indicating the feasibility of this approach have been obtained (Hung et al., 2000; Hung, 2000).
Previously we have shown that SV40 small t and its derivative, T/t-common (which contains the N-terminal common domain of large T and small t), possess a strong transcriptional repression activity (Wang et al., 1994). Subsequently, we showed that small t and T/t-common can repress the HER-2/neu promoter, with T/t-common has the strongest repressing activity between the two proteins (Lin et al., 2000). We then demonstrated that T/t-common can suppress the tumorigenic potential of the
HER-2/neu-overexpressing ovarian carcinoma SK-OV-3 cells both in vitro and in vivo through repressing HER-2/neu expression (Lin et al., 2000). These results lead to a conclusion that T/t-common may be used as a therapeutic agent against the
HER-2/neu-overexpressing cancers just like adenovirus E1A does. In this report, we demonstrate that SV40 T/t-common could specifically induce apoptosis in
HER-2/neu-overexpressing cancer cells but not in HER-2/neu-low-expressing tumor cells and non-transformed cells. We also demonstrate that T/t-common could sensitize HER-2/neu-overexpressing cancer cells to anti-cancer drug cisplatin and that
T/t-common could inhibit angiogenesis induced by HER-2/neu-overexpressing cancer cells. Together, these data suggest that T/t-common has the potential to be developed into an anti-cancer gene therapy agent.
Materials and Methods Cell Lines and Culture
.The SK-Br-3, MDA-MB-453, SK-OV-3, and SK-OV-3 ip1 cells were grown in DMEM/F12 nutrient mixture (Gibco, Grand Island, NY) supplemented with 10% FBS in 5% CO2. The SK-OV-3 derivative cells (SK-T/t-1, SK-T/t-2, SK-T/t-3, SK-T/t-4, SK-NEO-1, and SK-NEO-2) were grown under the same conditions, except that G418 (200 µg/ml) was added to the culture media. The AU565, MCF-7, NPC-TW04, HEp2, HeLa, MRC-5, CV-1, and NIH3T3 cells were cultured in DMEM (Gibco, Grand Island, NY) supplemented with 10% FBS in 5% CO2.
Plasmids and Viruses.
Plasmids pRSV.3-BglII, pRSV-T/t-common, pCMVHER2, and pcDNA3 were described previously (Lin et al., 2000). The pRc/CMV plasmid, which contains the neomycin resistance gene, was purchased from Invitrogen. The recombinant adenovirus rAdT/t which contains the T/t-common gene, and the vector adenovirus rAdV were constructed according to the protocol provided by B. Vogelstein (He et al., 1998).
Transfection and G418-resistant Colony Formation Assay.
DNA transfection was performed as described previously (Wang et al., 1994). Cells were seeded in 35-mm-diameter plates 24 h before transfection. Cells were transfected at 70% confluency by the calcium phosphate coprecipitation technique (Sambrook et al., 1989); 1 µg of pEGFP-C1, a plasmid which contains the CMV promoter-driven GFP gene, was also cotransfected as an internal control for
transfection efficiency. The cells were exposed to the transfection precipitate for 8 h, washed twice with fresh medium, and fed with fresh DMEM containing 0.5% FBS and appropriate concentration of G418. After 5 days, the media were removed and cells were refed with DMEM containing 10% FBS and appropriate concentration of G418. The G418-resistant colonies were counted two weeks latter.
Flow Cytometry Analysis.
For flow cytometry analysis of apoptosis by propidium iodide staining, floating and attached cells were collected, combined, and washed three times in PBS, and fixed in 80% ethanol at -20oC. Fixed cells were washed twice in PBS/1% BSA/0.1% Tween, and stained in 1ml of PBS containing 50 µg of propidium iodide and 10 µg of boiled RNase, at 4 oC for 1 h or longer. Stained cells were then analyzed on a flow cytometer (Becton Dickinson). Results were obtained as the percentage of subdiploid nuclei, which represent the apoptotic cells.
TUNEL Assay.
Cells were fixed in 4% paraformaldehyde for 45 min at room temperature. After rinsing with PBS, cells were permeabilized in a solution of Triton X-100 (0.1%) in sodium citrate (0.1%) for 2 min on ice. Samples, washed with PBS, were then
incubated in the TUNEL reaction mix (Roche) for 1 h at 37 oC in the dark. The stained samples were washed three times in PBS containing 0.2% BSA and 0.1% Triton X-100, and then visualized by a fluorescence microscope.
Results and Discussion
I. T/t-common specifically induces apoptosis in HER-2/neu-overexpressing cancer cells.
In the previous report, we have demonstrated that T/t-common could specifically induce apoptosis in HER-2/neu-overexpressing cancer cells but not in HER-2/neu low-expressing cancer cells and normal cells. To investigate whether this is the general property of T/t-common, for the past year, we included more
HER-2/neu-overexpressing and HER-2/neu low-expressing cancer cells and normal cells in our study. The results confirm our previous conclusion and indicate that T/t-common may be developed as a selective therapeutic agent against
HER-2/neu-overexpressing cancers. We also demonstrated that T/t-common induced apoptosis of HER-2/neu-overexpressing cancer cells by activating caspase activity. Finally we demonstrated that T/t-common’s effect could be blocked by re-expression of HER-2/neu and by overexpression of Bcl-2. The latter result suggests that
T/t-common induces apoptosis of HER-2/neu-overexpressing cancer cells through inhibiting HER-2/neu. Below are summaries of these studies.
1) T/t-common could specifically induce cell death in HER-2/neu-overexpressing cancer cells but not in normal cells and HER-2/neu low-expressing cancer cells.
Previously we generated a recombinant adenovirus, rAdNT/t, carrying the T/t-common gene. We found that rAdNT/t infection could permanently suppress the soft-agarose colony-forming ability of the HER-2/neu-overexpressing cancer cells SK-OV-3 and AU565. Since the expression of T/t-common is transient in
rAdNT/t-infected cells, the above finding implies that T/t-common may induce cell death in rAdNT/t-infected cells. To test whether T/t-common could indeed induce cell death in HER-2/neu-overexpressing cancer cells, we cotansfected either a
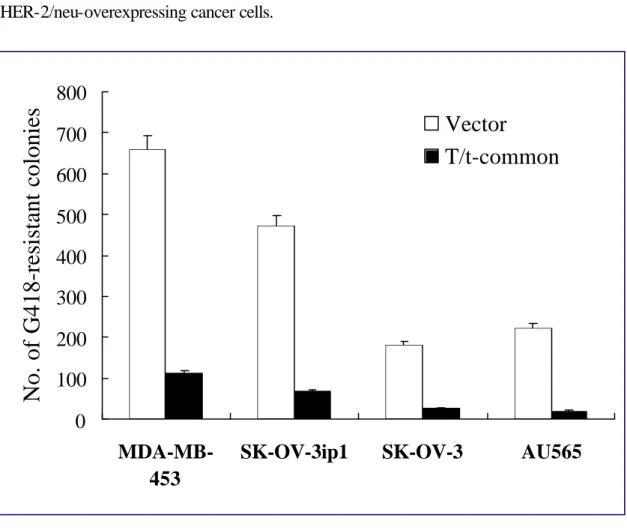
T/t-common-expressing plasmid or its vector control plasmid together with a neomycin resistance gene-encoding plasmid into HER-2/neu-overexpressing cancer cell lines SK-OV-3 (ovarian cancer cell line), SK-OV-3 ip1(ovarian cancer cell line), MDA-MB-453 (breast cancer cell line) and AU565 (breast cancer cell line), or HER-2/neu low-expressing cancer cell lines MCF-7 (breast cancer cell line), NPC-TW04 (nasopharyngeal cancer cell line) , HeLa (cervical cancer cell line) and HEp-2 (laryngeal cancer cell line), or non-transformed cell lines CV-1 and NIH3T3. We found that T/t-common cotransfection led to much less neomycin-resistant colonies than control cotransfection in all of the HER-2/neu-overexpressing cancer cell lines (Fig. 1). In contrast, in non-transformed cell lines and HER-2/neu
low-expressing cancer cell lines, cotransfection with T/t-common or vector plasmid led to similar number of neomycin-resistant colonies (Fig. 2). These data suggest that
T/t-common could specifically induce cell death in HER-2/neu-overexpressing cancer cell lines but not in non-transformed cells and HER-2/neu low-expressing cancer cells.
2) T/t-common-expressing SK-OV-3 cells underwent apoptosis under low serum condition.
To investigate whether T/t-common induced apoptosis in
HER-2/neu-overexpressing cancer cells, we grew previously established
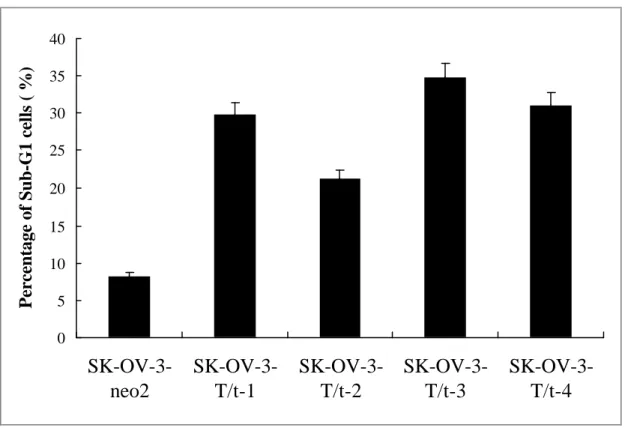
T/t-common-expressing SK-OV-3 clones or vector-transfected SK-OV-3 clone in low-serum media. The cells were harvested 4 days later, fixed, stained with propidium iodide, and subjected to FACS analysis. As shown in Fig. 3, the
T/t-common-expressing clones (SK-OV-3-T/t-1, SK-OV-3-T/t-2, SK-OV-3-T/t-3, and SK-OV-3-T/t-4) contained more cells in sub-G1 phase than did the vector-transfected clone (SK-OV-3-neo2). These data indicate that T/t-common-expressing SK-OV-3 cells underwent apoptosis under low serum condition. To further confirm that these T/t-common-expressing clones underwent apoptosis, TUNEL assays were performed. The T/t-common-expressing SK-OV-3 clones or the vector-transfected clone were incubated in low-serum media. Four days later, the cells were fixed and subjected to TUNEL assay. As shown in Fig. 4, all of the T/t-common-expressing clones
(SK-OV-3-T/t-1, SK-OV-3-T/t-2, SK-OV-3-T/t-3, and SK-OV-3-T/t-4) were TUNEL-positive while the vector-transfected clone (SK-OV-3-neo2) was TUNEL-negative. The nuclei of the cells were also stained with DAPI. While the nuclei of the T/t-common-expressing cells displayed apoptotic feature, that is they were condensed and stained brightly with DAPI, the nuclei of vector-transfected cells were evenly stained and not condensed (data not shown). Together these data strongly indicate that the T/t-common-expressing SK-OV-3 cells underwent apoptosis under low serum condition.
3) T/t-common could induce apoptosis in HER-2/neu-overexpressing cancer cells but not in HER-2/neu low-expressing cancer cells and normal cells.
The above data indicate that T/t-common could induce apoptosis in
HER-2/neu-overexpressing ovarian carcinoma SK-OV-3 cells. To investigate whether T/t-common could induce apoptosis in other cells, recombinant adenovirus carrying T/t-common (rAdT/t) and vector adenovirus (rAdV) were used to infect
HER-2/neu-overexpressing breast cancer cells (SK-Br-3, AU565, and MDA-MB453), HER-2/neu-overexpressing ovarian cancer cells (SK-OV-3), HER-2/neu
low-expressing breast cancer cells (MCF-7), and non-transformed cells (CV-1 and MRC-5). After infection, the cells were incubated in low-serum media. Various times after infection, the cells were harvested, fixed, stained with propidium iodide, and
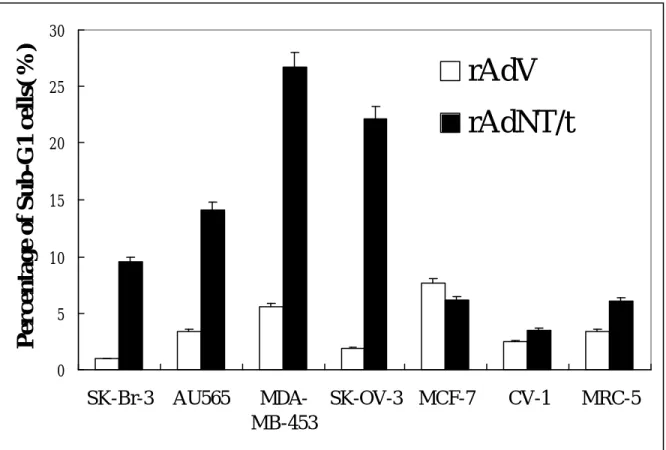
subjected to FACS analysis. As shown in Fig. 5, compared to rAdV infection, rAdT/t infection dramatically increase the number of apoptotic cells (i.e., sub-G1 cells) in HER-2/neu-overexpressing cancer cells (SK-Br-3, AU565, MDA-MB453, and SK-OV-3). In contrast, in HER-2/neu low-expressing breast cancer cells (MCF-7) and non-transformed cells (CV-1 and MRC-5), rAdT/t infection caused similar number of apoptotic cells as rAdV infection. These data indicate that T/t-common could
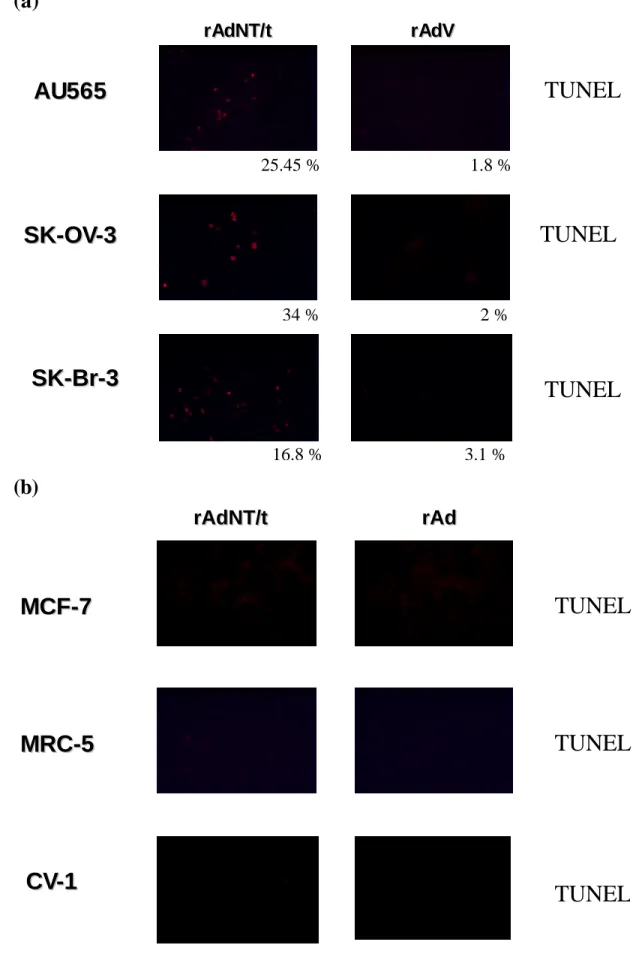
specifically induce apoptosis in HER-2/neu-overexpressing cancer cells but not in HER-2/neu low-expressing cancer cells and non-transformed cells. The TUNEL assays (Fig. 6a and 6b) also confirmed the above conclusion.
4) T/t-common expression in HER-2/neu-overexpressing cancer cells leads to activation of caspase acitivity.
To study whether T/t-common induces apoptosis of HER-2/neu-overexpressing cancer cells through activating caspase activity, we measured caspase activity of rAdNT/t or rAdV infected MDA-MB453 cells by detecting the cleavage product of poly(ADP-ribose)polymerase (PARP). As shown in Fig. 7, T/t-common virus
infection caused significant increase in the cleavage product of PARP than vector virus infection, indicating that T/t-common expression could lead to activation of caspase activity.
5) The ability of T/t-common to induce apoptosis of HER-2/neu-overexpressing cancer cells was derived from its ability to repress HER-2/neu expression.
As described in our previous paper (Lin et al., 2000), T/t-common could suppress the tumorigenic potential of HER-2/neu-overexpressing cancer cells through repressing HER-2/neu expression. Previous studies (Roh et al., 2000; Cuello et al., 2001) showed that repression of HER-2/neu expression in HER-2/neu-overexpressing cancer cells can lead to apoptosis. Taken together, these observations suggest that T/t-common may induce apoptosis through repressing HER-2/neu expression in
HER-2/neu-overexpressing cancer cells. To investigate whether this hypothesis is true, we reintroduced the HER-2/neu gene under the control of CMV IE promoter (which
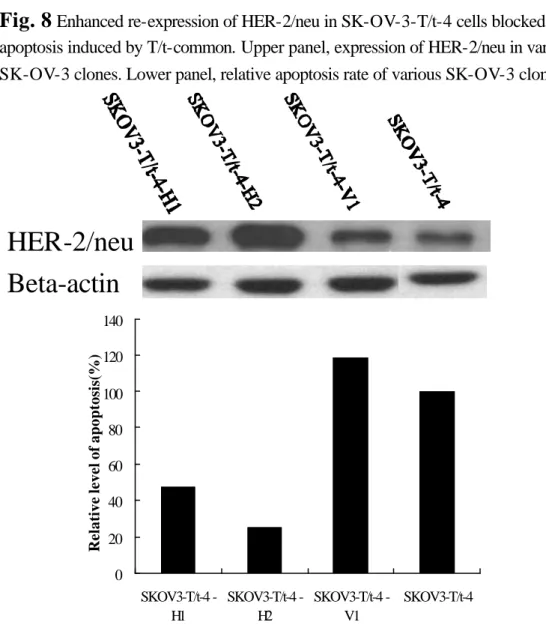
is not repressed by T/t-common) into the T/t-common-expressing SK-OV-3 clone (SK-OV-3-T/t-4). As shown in Fig. 8 upper panel, both clones SK-OV-3-T/t-4-H1 and SK-OV-3-T/t-4-H2 (lanes 1and 2, respectively) produced a large amount of
HER-2/neu, while the vector-transfected clone (SK-OV-3-T/t-4-V1; lane 3) produced similar amount of HER-2/neu as the parental SK-OV-3-T/t-4 clone (lane 4). The
parental SK-OV-3-T/t-4 clone, the vector-transfected clone (SK-OV-3-T/t-4-V1), and incubated in low serum media and then subjected to TUNEL assays. As shown in Fig. 8 lower panel, while the parental cells and the vector-transfected cells contained a significant amount of apoptotic cells, the HER-2/neu-reexpressed cells contained
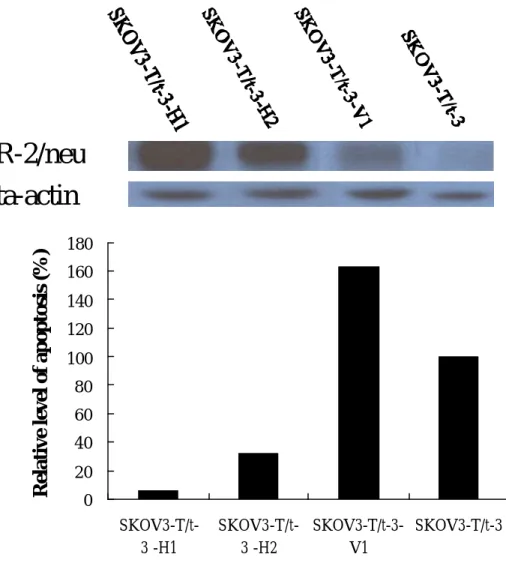
much less apoptotic cells. These data indicate that re-expression of HER-2/neu could block T/t-common’s ability to induce apoptosis in HER-2/neu-overexpressing cancer cells. To further verify this observation, we also reintroduced the HER-2/neu gene under the control of CMV IE promoter into another T/t-common-expressing SK-OV-3 clone (SK-OV-3-T/t-3). As shown in Fig. 9, similar results as those shown in Fig. 8 were obtained. To rule out the possibility that the above observations are due to clonal selection of non-apoptotic clones, we also did transient cotransfection assays. We cotransfected β−galactosidase-encoding plasmid with either T/t-common-encoding plasmid or T/t-common-encoding plasmid plus CMV promoter-driven HER-2/neu plasmid into AU565 breast carcinoma cells. As shown in Fig. 10, T/t-common cotransfection led to reduced number of blue cells as compared to vector
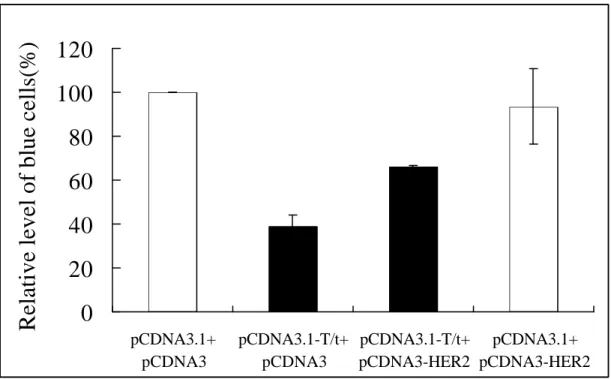
cotransfection (compare bar 3 with bar 4), indicating that T/t-common caused the death of the transfected cells. Interestingly when HER-2/neu was cotransfected with T/t-common, the number of blue cells increased significantly as compared to
T/t-common transfection alone (compare bar 1 with bar 3). This data indicates that cotransfection of HER-2/neu could block T/t-common’s killing effect. Taken this data together with above ones, we conclude that re-expression of HER-2/neu could block T/t-common’s ability to induce apoptosis in HER-2/neu-overexpressing cancer cells. Since T/t-common is known to inhibit HER-2/neu, these data also suggest that
T/t-common’s ability to induce apoptosis of HER-2/neu-overexpressing cancer cells is derived from its ability to inhibit HER-2/neu.
6) Bcl-2 could block T/t-common’s ability to kill HER-2/neu-overexpressing cancer cells.
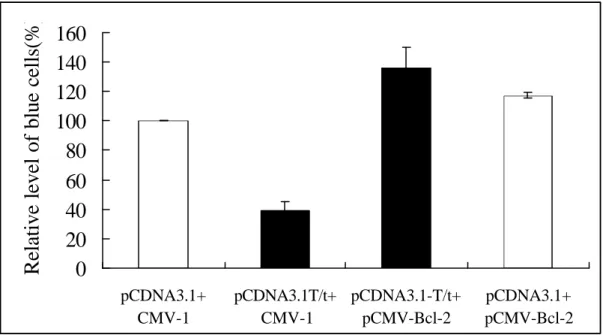
Bcl-2 is known to inhibit apoptosis induced by various agents. To investigate whether Bcl-2 could block T/t-common’s ability to kill HER-2/neu-overexpressing cancer cells, we did transient cotransfection assay as described above. We
cotransfected β−galactosidase-encoding plasmid with either T/t-common plasmid or T/t-common plasmid plus Bcl-2 plasmid into AU565 breast carcinoma cells. As shown in Fig. 11, T/t-common cotransfection led to reduced number of blue cells as compared to vector cotransfection (compare bar 3 with bar 4), indicating that T/t-common killed the transfected cells. When Bcl-2 was cotransfected with T/t-common, the number of blue cells increased significantly as compared to T/t-common transfection alone (compare bar 1 with bar 3). This data indicates that Bcl-2 could block T/t-common’s ability to kill HER-2/neu-overexpressing cancer cells.
II. T/t-common sensitized HER-2/neu-overexpressing cancer cells to anti-cancer drug cisplatin.
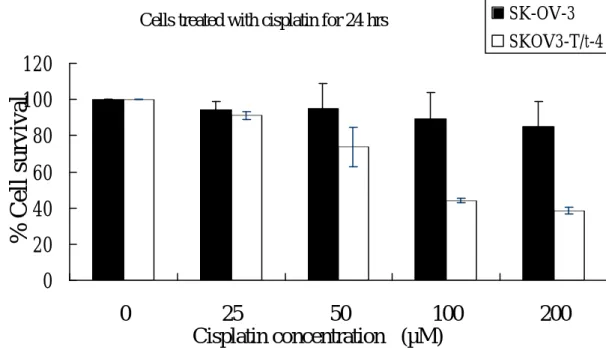
T/t-common can inhibit HER-2/neu, we reasoned that T/t-common might sensitize HER-2/neu-overexpressing cancer cells to ant-cancer drugs. To test this, we incubated T/t-common-expressing SK-OV-3 cells and parental SK-OV-3 cells in various amounts of cisplatin for 24 or 48 h. The survival of these cells was estimated by MTT assays. As shown in Fig. 12, after cisplatin treatment, the survival rate of
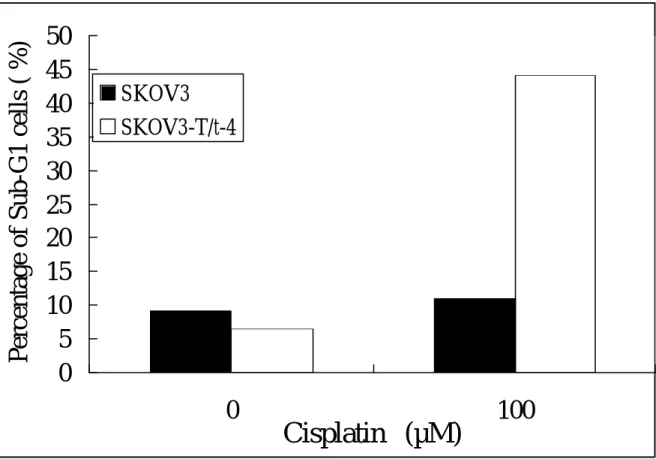
T/t-common-expressing SK-OV-3 cells was much lower than the parental cells, indicating that T/t-common sensitized these HER-2/neu-overexpressing cancer cells to cisplatin. To further investigate this phenomenon, we analyzed the appearance of apoptotic cells after cisplatin treatment by sub-G1 analysis. We found that more T/t-common-expressing SK-OV-3 cells underwent apoptosis than parental SK-OV-3 cells after cisplatin treatment (Fig. 13). Together these data clearly indicate that
T/t-common can enhance apoptosis induced by cisplatin in HER-2/neu-overexpressing cancer cells.
III. T/t-common inhibited angiogenesis induced by HER-2/neu-overexpressing cancer cells.
HER-2/neu overexpression is known to stimulate the synthesis of angiogenic factors. The fact that T/t-common can inhibit HER-2/neu suggests T/t-common may inhibit angiogenesis induced by HER-2/neu-overexpressing cancer cells. We therefore tested the angiogenesis-inducing activities of T/t-common-expressing SK-OV-3 cells and Vector-transfeced SK-OV-3 cells in nude mice. We mixed these cancer cells with matrigel and inoculated them subcutaneously into nude mice. One week later, the mice were sacrificed and the matrigel were exposed and photographed. As Shown in Fig. 14, the matrigel containing vector-transfected SK-OV-3 cells appeared dark red, indicating that angiogenesis occurred. In contrast, the matrigels containing
T/t-common-expressing SK-OV-3 cells appeared pale white, indicating that angiogenesis did not occur. These data suggest that T/t-common can inhibit angiogenesis induced by HER-2/neu-overexpressing cancer cells.
References
Bargmann CI, Hung M-C, and Weinberg RA. (1986) Cell, 45, 649-657.
Berchuck A, Kamel A, Whitaker R, Kerns B, Olt G, Kinney R, Soper JT, Dodge R, Clarke-Pearson DL, Marks P, McKenzie S, Yin S and Bast JRC. (1990). Cancer Res., 50, 4087-4091
Chang JY, Xia W, Shao R and Hung M-C. (1996). Oncogene, 13, 1405-1412. Chang JY, Xia W, Shao R, Sorgi F, Hortobagyi GN, Huang L and Hung M-C. (1997). Oncogene, 14, 561-568.
Chen H, Yu D, Chinnadurai G, Karunagaran D and Hung M-C. (1997). Oncogene, 14, 1965-1971.
Chen H and Hung M-C. (1997). J. Biol. Chem., 272, 6101-6104.
Coussens L, Yang-Feng TL, Liao Y-C, Chen E, Gray A, McGrath J, Seeburg PH, Libermann TA, Schlessinger J, Francke U, Levinson A and Ullrich A. (1985). Science, 230, 1132-1139.
Cuello et al., Cancer Res. 61:4892-4900, 2001
Gavrieli I, Sherman Y, Ben-Sasson SA. (1992). J. Cell Biol., 119, 493-501.
Guy CT, Webster MA, Schaller M, Parsons TJ, Cardiff RD and Muller WJ. (1992). Proc. Natl. Acad. Sci. USA, 89, 10578-10582.
Hassan W, Sanford MA, Woo SLC, Chen S-H, and Hall SJ. (2000). World J. Urol., 18, 130-135.
He T-C, Zhou S, DA Costa LT, Yu J, Kinzler KW and Vogelstein B. (1998). Proc. Natl. Acad. Sci. USA, 95, 2509-2514.
Hung K, Hayashi R, Lafond-Walker A, Lowenstein C, Pardoll D and Levitsky H. (1998b). J. Exp. Med., 188, 2357-2368.
Hung M-C. (2000). Abstract of International Symposium on DNA Vaccine and Gene Therapy Technology, Academia Sinica, Taipei, p.15.
Hung M-C, Hortobagyi GN, and Ueno NT. (2000). Adv. Exp. Med. Biol., 465, 171-180.
Hung M-C and Lau Y-K. (1999). Semin. Oncol., 26 (suppl. 12), 51-59.
Hung M-C, Chang JYJ and Xing XM. (1998). Adv. Drug Delivery Rev., 30, 219-227. Hung M-C, Matin A, Zhang Y, Xing X, Sorgi F, Huang L and Yu D. (1995). Gene, 159, 65-71.
Hung M-C, Schechter AL, Chevray PY, Stern DF and Weinberg RA. (1986). Proc. Natl. Acad. Sci. USA, 83, 261-264.
Hynes NE and Stern DF. (1994). Biochim. Biophys. Acta, 1198, 165-184.
Kao M-C, Liu G-Y, Chuang T-C, Lin Y-S, Wuu J-A and Law S-L. (1998). Oncogene, 16, 547-554.
Lin Y-C, Peng J-M and Wang W-B. (2000). Oncogene, 19, 2704-2713. Matin A and Hung M-C. (1993). Cell Growth Diff., 4, 1051-1056.
Roh et al., Cancer Res. 60:560-565, 2000
Sadasivan R, Morgan R, Jennings S, Austenfeld M, van Veldhuizen P, Stephens R and Noble M. (1993). J. Urol., 150, 126-131.
Shtrichman R and Kleinberger T. (1998). J. Virol., 72, 2975-2982.
Simons JW, Mikhak B, Chang JF et al. (1999). Cancer Res., 59, 5160-5168. Slamon DJ, Clark GM, Wong SG, Levin WJ, Ullrich A and McGuire WL. (1987).
Science, 235, 177-182.
Slamon DJ, Godolphin W, Jones LA, Holt JA, Wong SG, Keith DE, Levin WJ, Stuart SG, Udove J, Ullrich A and Press MF. (1989). Science, 244, 707-712.
Tan CM, Yao J, and Yu D. (1997). Cancer Res. 57, 1199-1205.
Tsai C-M, Chang K-T, Perng R-P, Mitsudomi T, Chen M-H, Kadoyama C and Gazdar AF. (1993). J. Natl. Cancer Inst., 85, 897-901.
Tsai C-M, Perng R-P, Chen M-H, Jan Y-H, Hung M-C, Ku T-Y, and Chang K-T.
(1994). J. Natl. Cancer Inst., 86, 1018-1020.
Ueno NT, Yu D, and Hung M-C. (1997). Oncogene, 15, 953-960.
Wang W-B, Bikel I, Marsilio E, Newsome D and Livingston DM. (1994). J. Virol., 68,
6180-6187.
Xing X, Martin A, Yu D et al. (1996). Cancer Gene Ther., 3, 168-174.
Yamamoto T, Ikawa S, Akiyama T, Semba K, Nomura N, Miyajima N, Saito T and Toyoshima K. (1986). Nature, 319, 230-234.
Yu D and Hung MC. (2000). Oncogene, 19, 6115-6121.
Yu D, Jing T, Liu B, Yao J, Tan M, McDonnell TJ, and Hung M-C. (1998). Mol. Cell, 2, 581-591.
Yu D, Hamada J-I, Zhang H, Nicolson GL and Hung M-C. (1992). Oncogene, 7, 2263-2270.
Yu D, Matin A, Xia W, Sorgi F, Huang L and Hung M-C. (1995). Oncogene, 11, 1383-1388.
Yu D, Scorsone K and Hung M-C. (1991). Mol. Cell. Biol., 11, 1745-1750.
Yu D, Suen T-C, Yan D-H, Chang L-S and Hung M-C. (1990). Proc. Natl. Acad. Sci. USA, 87, 4499-4503.
Yu D, Wang S-S, Dulski KM, Tsai C-M, Nicolson, GL and Hung M-C. (1994). Cancer Res., 54, 3260-3266.
Yu D, Wolf JK, Scanlon M, Price JE and Hung M-C. (1993). Cancer Res., 53, 891-898.
Zhang Y, Yu D, Xia, W and Hung M-C. (1995). Oncogene, 10, 1947-1954.
Evaluation of Accomplishment
Overall this study has provided evidences that demonstrate that T/t-common has the potential to be developed into an anti-cancer gene therapy agent.
Fig.1
Inhibition of G418-resistant colony formation by T/t-common in HER-2/neu-overexpressing cancer cells.Fig. 2
T/t-common did not inhibit G418-resistant colony formation in HER-2/neu low-expressing cancer cells and non-transformed cells.0 20 40 60 80 100 120 140 160
NPC-TW04 Hep2 HeLa CV-1 NIH3T3 MCF-7
NO. of G418-resistant colonies
Vector
T/t-common
0
100
200
300
400
500
600
700
800
MDA-MB-453
SK-OV-3ip1
SK-OV-3
AU565
No. of G418-resistant colonies
Vector
T/t-common
Fig. 3
T/t-common-expressing SK-OV-3 clones underwent apoptosis under low serum condition. 0 5 10 15 20 25 30 35 40SK-OV-3-neo2
SK-OV-3-T/t-1
SK-OV-3-T/t-2
SK-OV-3-T/t-3
SK-OV-3-T/t-4
Percentage of Sub-G1 cells ( %)
Fig. 4
TUNEL assay showed that T/t-common-expressing SK-OV-3 clones underwent apoptosis under low serum condition.SKOV3-T/t-1
SKOV3-T/t-2
SKOV3-T/t-3
SKOV3-T/t-4
Fig. 5
T/t-common-containing adenovirus infection induced apoptosis inHER-2/neu-overexpressing cancer cells but not in HER-2/neu low-expressing cancer cells and non-transformed cells.
0 5 10 15 20 25 30 SK-Br-3 AU565 MDA-MB-453 SK-OV-3 MCF-7 CV-1 MRC-5
Percentage of Sub-G1 cells( %)
rAdV
rAdNT/t
Fig. 6
TUNEL assay showed that T/t-common-containing adenovirus infection induced apoptosis in HER-2/neu-overexpressing cancer cells (a) but not in HER-2/neu low-expressing cancer cells and non-transformed cells (b)(a)
(b)
r
r
A
A
d
d
N
N
T
T
/
/
t
t
r
r
A
A
d
d
M
M
C
C
F
F
-
-
7
7
TUNEL
M
M
R
R
C
C
-
-
5
5
TUNEL
C
C
V
V
-
-
1
1
TUNEL
r rAAddNNTT//tt rrAAddV VA
A
U
U
5
5
6
6
5
5
TUNEL
S
S
K
K
-
-
O
O
V
V
-
-
3
3
TUNEL
TUNEL
S
S
K
K
-
-
B
B
r
r
-
-
3
3
34 % 2 % 25.45 % 1.8 % 16.8 % 3.1 %Fig. 7
Activation of caspase activity by T/t-common in HER-2/neu-overexpressing MDA-MB-453 cells. Cells were infected with T/t-common-carrying adenovirus (rAdN/T/t) or vector virus (rAdV) and then incubated in low serum media. 72 h later, cell lysates were analyzed for caspase-cleaved product of PARP by western blot.Fig. 8
Enhanced re-expression of HER-2/neu in SK-OV-3-T/t-4 cells blocked apoptosis induced by T/t-common. Upper panel, expression of HER-2/neu in various SK-OV-3 clones. Lower panel, relative apoptosis rate of various SK-OV-3 clones.Cleaved PARP
GAPDH
mock
rAdV
rAd
N
T/
t
HER-2/neu
Beta-actin
0 20 40 60 80 100 120 140 SKOV3T/t4 -H1 SKOV3T/t4 -H2 SKOV3T/t4 -V1 SKOV3-T/t-4Fig. 9
Enhanced re-expression of HER-2/neu in SK-OV-3-T/t-3 cells blocked apoptosis induced by T/t-common. Upper panel, expression of HER-2/neu in various SK-OV-3 clones. Lower panel, relative apoptosis rate of various SK-OV-3 clones0 20 40 60 80 100 120 140 160 180 SKOV3-T/t-3 -H1 SKOV3-T/t-3 -H2 SKOV3-T/t-3-V1 SKOV3-T/t-3
Relative level of apoptosis (%)
HER-2/neu
Beta-actin
Fig. 10
Transient cotransfection assays showing that cotransfection of HER-2/neu can block T/t-common’s ability to kill HER-2/neu-overexpressing cancer cells.PCMV-βgal were cotransfected into AU565 cells with pCDNA3.1-T/t-common plus pCDNA3-HER2 (bar 3), pCDNA3.1 plus pCDNA3-HER2 (bar 4),
pCDNA3.1-T/t-common plus pCDNA3 (bar 2), or pCDNA3.1 plus pCDNA3(bar 1). After transfection, cells were incubated in low serum media, and stained for
β-galactosidase-expressing blue cells.
0
20
40
60
80
100
120
pCDNA3.1+ pCDNA3 pCDNA3.1-T/t+ pCDNA3 pCDNA3.1-T/t+ pCDNA3-HER2 pCDNA3.1+ pCDNA3-HER2Fig. 11
Transient cotransfection assays showing that cotransfection of Bcl-2 can block T/t-common’s ability to kill HER-2/neu-overexpressing cancer cells.PCMV-βgal were cotransfected into AU565 cells with pCDNA3.1-T/t-common plus pCMV-Bcl2 (bar 4), pCDNA3.1 plus pCMV-Bcl2 (bar 3), pCDNA3.1-T/t-common plus pCMV-1 (bar 2), or pCDNA3.1 plus pCMV-1(bar1) After transfection, cells were incubated in low serum media, and stained for β-galactosidase-expressing blue cells.
0
20
40
60
80
100
120
140
160
pCDNA3.1+ CMV-1 pCDNA3.1T/t+ CMV-1 pCDNA3.1-T/t+ pCMV-Bcl-2 pCDNA3.1+ pCMV-Bcl-2Fig. 12
T/t-common-expressing SK-OV-3 cells are more sensitive to anti-cancer drug cisplatin than parental SK-OV-3 cells. Cells were treated with variousconcentration of cisplatin, 24 h (upper panel) or 48 h (lower panel) later, viable cells were counted by MTS assays.
Cells treated with cisplatin for 24 hrs
0
20
40
60
80
100
120
0
25
50
100
200
Cisplatin concentration (µM)
% Cell survival
SK-OV-3 SKOV3-T/t-4Cells treated with cisplatin for 48hrs
0 20 40 60 80 100 120 0 25 50 100 200