行政院國家科學委員會專題研究計畫成果報告
靈芝多醣之分析與鑑定
Str uctur e and pr oper ties of the glucans isolated fr om
Ganoderma lucidum
計畫類別:個別型計畫
計畫編號:NSC
91-2316-B-002-011
執行期間:
88 年
8 月
1 日至
91 年
7 月 30 日
計畫主持人 :孫璐西 台灣大學食品科技研究所
共同主持人 :呂廷璋 台灣大學食品科技研究所
王伯徹 食品工業發展研究所
計畫參與人員:張毅偉 台灣大學食品科技研究所
周欣漪 台灣大學食品科技研究所
處理方式: 可立即對外提供參考
(請打¡) ■一年後可對外提供參考
兩年後可對外提供參考
(必要時,本會得展延發表時限)
執行單位:台灣大學食品科技研究所
中華民國
91 年
10 月
31 日
行政院國家科學委員會專題研究計畫成果報告
靈芝多醣之分析與鑑定
Str uctur e and pr oper ties of the glucans isolated fr om Ganoderma
lucidum
計 畫 編 號:NSC-91-2316-B-002-011
執 行 期 限:88 年 8 月 1 日至 91 年 7 月 31 日
主 持 人:孫璐西 台灣大學食品科技研究所
共同主持人:呂廷璋 台灣大學食品科技研究所
王伯徹 食品工業發展研究所
參與研究人員:張毅偉 台灣大學食品科技研究所
周欣漪 台灣大學食品科技研究所
一、中文摘要
本研究建立一個簡單且便宜(1→ 3)-β-D-glucan 含量的螢光檢測法與 一個檢測(1→3)-β-D-glucan 分支度 的酵素-高效陰離子交換層析法。這些 方法讓我們的研究團隊得以檢測靈芝 產品中 glucan 的性質。螢光檢測方法 是 利 用 螢 光 染 劑 aniline blue 中 的”sirofluor”會選擇性與(1→3)-β -D-glucan 錯 合 產 生 螢光 的 特性 而 建 立。(1→3)-β-D-glucan 與 sirofluor 的錯合物會在激發波長 395nm,放射波 長 495nm 有強烈的特徵螢光,而螢光 的強度會因 aniline blue 中 sirofluor 的 含量、(1→3)-β-D-glucan 的含量與 分支度,以及溶液的 pH 與離子強度不 同而有所改變。以數種(1→3)-β -D-glucan:GLG、krestin、laminarin 與 lentinan 進行分析,發現濃度與螢光強 度間具有良好的線性關係,但斜率明顯 不同。倘若標準品不易製備的話,建議 使用商業產品—laminarin 為相對標準 品 來 進 行 檢 測 , 結 果 可 以 laminarin equivalents (LE)來表示。酵素—高效陰 離子交換層析法是利用β-D-glucanase 將(1→6)branched (1→3)-β-D-glucan 水 解 成 葡 萄 糖 與 龍 膽 雙 醣 ( O-6- β -D-glucosyl -D-glucose,gentibiose),再 配合高效陰離子交換層析法分析葡萄 糖與龍膽雙醣的含量即可推算出(1→ 3)-β-D-glucan 的分支度。合併使用 endo-與 exo-(1→3)-β-D-glucanase 可將多醣完全水解。靈芝純化所得的多 醣經過 NMR 確定為(1→6)branched (1→3)-β-D-glucan,分子量約百萬, 分 支 度 0.25 ~ 0.33 。 靈 芝 (1 → 3)-â-D-glucan 的含量、分子量與分支度 會隨品系不同、培養方法不同(子實體 與液態發酵)或培養條件不同而有所差 異,所以在靈芝培養間監控 glucan 的 性質是很重要的。 關鍵字:靈芝、(1→3)-β-D-葡聚糖、 aniline blue、螢光、(1→3) -β-D-glucanaseAbstr act
A simple and inexpensive
fluorometric method for (1→3) –â –D-glucans content determination and enzyme-chromatographic method for determination of degree of branching of (1→3)-â-D-glucans was developed in this studies. The new methods allowed our research group to examine the properties of glucans in Ganoderma products. The fluorometric method was based on the specific interaction of (1→
3)-â-D-glucans with sirofluor, a weakly fluorescent component of aniline blue dye. The interacted complexes of (1→ 3)-â-D-glucans and sirofluor showed a strong fluorescence with an excitation maximum of 395 nm and an emission maximum of 495 nm. The intensity of fluorescence varied with the amount of fluorophore existed in aniline blue, the amount and molecular size of (1→3) -â-D-glucans, and the pH and ionic strength of solution. All tested glucans had shown a good linear correlation between glucan concentration and fluorescence intensity, whereas the linear range and the slope of the linear varied with the different glucan. It is
recommended to use laminarin, a known and commercial available (1→3)-â-D-glucan, as standard to quantify the amount of the glucans in samples, when the glucan from selected material are not available. The results could present as laminarin equivalents (LE), as laminarin had used as standard. The enzyme-chromatographic method was based on the hydrolysis of â-(1→3) glycosidic linkage of D-glucans to yield D-glucose and
O-6-â-D-glucopyranosyl-D-glucopyrano se (gentiobiose) and the quantification of these two sugars. The difficulty of complete hydrolyzing the
macromolecule to its monomer and dimer was overcome by using mix of endo- and exo- (1,3)-â- D-glucanase. The quantification of glucose and gentiobiose was achieved by using high-performance anion-exchange chromatography. The (1→3)-â-D -glucans of Ganoderma lucidium was detected by fluorometric assay and isolated and purified for property analysis. The â-(1→3) glycosidic backbone and the â-(1→6) glycosidic branching linkage was confirmed by NMR analysis. The molecular weight of the D-glucan was larger than million daltons. The degree of branching was
0.25~0.33 elucidated by NMR and enzyme-chromatographic analysis. The content, molecular weight and degree of branching of (1→3)-â-D-glucans of Ganoderma lucidium varied with strains, source (fruiting body or mycelium culture), and cultivation conditions. The results indicated the variation of (1 →3)-â-D-glucans properties was significant, therefore monitoring the properties of the glucans was essential for the quality control of Ganoderma production.
Keywords:Ganoderma lucidum、(1→3) -β-D-glucan、aniline blue、 fluorescence、(1→3)-β -D-glucanase
二、緣由與目的:
靈芝(Ganoderma lucidum)是一 傳統重要中藥,有降血壓、降血脂、降 血糖、抗血拴、抗發炎、抗病毒及抗腫 瘤等生理活性,其中抗腫瘤的活性主要 來自多醣,但並非靈芝中所有多醣都有 效果,具抗腫瘤活性的靈芝多醣被證實 以(1→3)-β- glucan 為主幹,其上具 有多個由一個或兩個葡萄糖以β-(1→ 6)鍵結所構成的分支。分支程度對於 抑制腫瘤的效果有決定性的影響:分支 程度為主幹上每 2.5 到 5 個葡萄糖帶有 一個分支時,其抗腫瘤活性最強,分支 度太多或過少,其活性皆會明顯下降。 靈 芝 原 料 的 生 產 方 式 有 子 實 體 ( fruiting body ) 栽 植 及 菌 絲 體 (mycelium)液態培養。在生產 glucan 的過程中,廠家常以溶劑沈澱或分子量 區分膜配合酚-硫酸法(phenol-sulfuric acid assay)來檢測產品中的水溶性總多 醣量以作為生產多醣的指標,但此法易 受樣品中其他醣類的干擾,且專一性不 足。本研究擬以靈芝子實體與菌絲體培 養液所得活性多醣區分物為基礎,設計 出 一 套 分 析 靈 芝 活 性 多 醣 的 簡 易 方法,再進一步探討靈芝子實體栽植與深 層發酵培養所生成(1→3)-β-D-glucan 結構與含量的差異,以找尋生產具活性 的(1→3)-β-D-glucan 最適液態培養 條件。 Aniline blue 原是用在組織染色上 的 一 種 染 劑 , 後 來 發 現 其 中 所 含 的 sirofluor 對(1→3)-β-D-glucan 具有 專一結合性,擬利用螢光染劑 aniline blue 對(1→3)-β-D-glucan 的專一結 合 性 質 來 定 量 分 析 ( 1 → 3 ) - β -D-glucan。 另外擬利用高度專一性的(1→3) -β-D-glucanase 將(1→3)-β-D-glucan 水解成葡萄糖(主幹部分)與龍膽雙醣 (分支點部分),配合高效陰離子交換 層 析 法 檢 測 水 解 產 物 , 如 此 可 得 β -D-glucan 的含量與分支比例。
三、材料與方法:
(一)多醣的萃取與純化: 以 100℃熱水萃取靈芝子實體粉 末,而後將濾液進行溶劑脫水,乾燥後 即得粗多醣。 將粗多醣溶於水中,加入 10% (w/v)cetylpyridinium chloride 溶液 ( CPC ) 去 除 帶 電 多 醣 , 再 利 用 amyloglucosidase 與量 protease 去除α-(1→4)-D-glucan 與蛋白質,得到純 化的棕白色棉絮狀靈芝多醣 GLPS 共 4.6250 公 克 , 經 酚 — 硫 酸 法(phenol-sulfuric acid assay)檢測知其 總醣含量約佔 80.98%。 (二)多醣的分級沈降: 靈芝多醣 GLPS 溶於水中,逐滴 加入 95%酒精,溶液中酒精濃度約 20 %時得到大分子靈芝葡聚糖 GLG;當 溶液酒精濃度約 60%時得到中分子靈 芝葡聚糖;當溶液酒精濃度約 80%時 得到小分子靈芝葡聚糖。 (三)多醣物化性質的檢測:
利用 phenol-sulfuric acid assay 檢 測多醣中總醣量。利用 Morgan-Elson assay 檢測多醣中 hexosamine 的含量。 利用 m-hydroxydiphenyl method 檢測多 醣中 uronic acid 的量。多醣中蛋白質量 的檢測以 Folin-Lowry method 進行。多 醣 單 醣 組 成 之 鑑 定 依 Miyazaki 與 Nishijima 在 1982 年的水解條件進行。 多醣鍵結形式之測定依 Kim 等在 2000 年發表的方法以核磁共振光譜(nuclear magnetic resonance / NMR)檢測之。 (四)螢光檢測分析: 樣品溶於 3 毫升 0.3N 的 NaOH 中,攪拌 30 分鐘使完全溶解,再用 1N HCl 調整 pH 至 11.5,之後加入 pH 11.5 的 50mM Na2HPO4-NaOH 緩 衝 液 (0.5M NaCl)定容。取 2 毫升入試管, 加入 0.2 毫升濃度為 1 毫克/毫升的 aniline blue 試劑,利用 Vortex 混勻後靜 置 2 個小時,之後進行螢光強度的檢測 (激發波長 395nm,放射波長 495nm)。 (五)酵素—高效陰離子交換層析分 析: 樣品完全溶於 0.3N NaOH 後,添 加 1N HCl 調整 pH 至 5.0,再以 buffer 定容。添加酵素在 37℃下培養 24 小 時,之後 100℃加熱 15 分鐘抑制酵素 活性。利用高效陰離子交換層析法監控 水解產物(葡萄糖與龍膽雙醣)的生成 狀 況 。 , 檢 測器 cell 為 PAD-2 cell (gold),管柱為 Carbopac PA1,Guard column 為 Carbopac PA1 guard,loop 容積為 25μl,以 0.2N NaOH 為移動 相,流速為 1.0ml/min。積分系統為 SISC-32(訊華,台灣)。
(六)GL4 與 GL11 靈芝子實體栽植與 深層發酵培養 1.子實體培養 兆豐興農股份有限公司提供二種 品系赤芝 (Ganoderma lucidum)乾燥 子實體:GL4 及 GL11。 2.靈芝液態培養 財團法人食品工業發展研究所提 供,GL4 及 GL11 均有以大型發酵槽 (20L)發酵及小槽發酵(5L)兩種。
四、結果與討論:
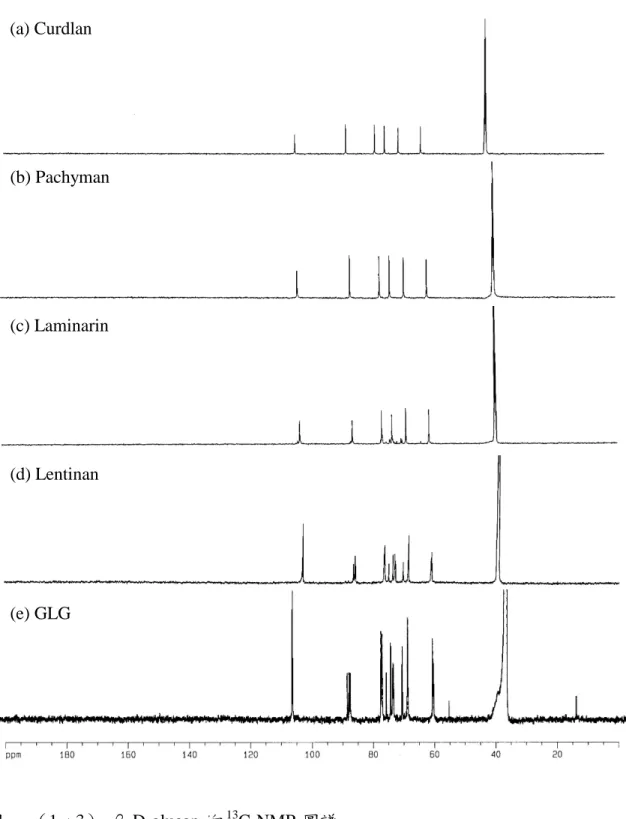
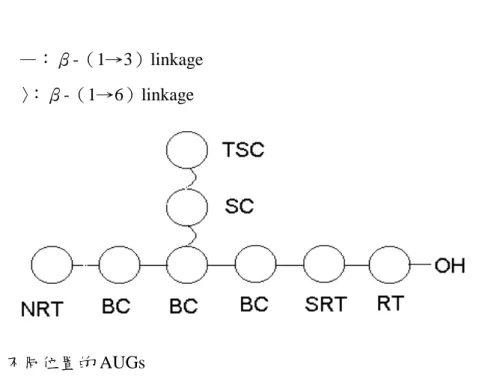
(一)多醣的萃取與純化 用 40.5 公升的熱水萃取 900 公克 靈芝子實體粉末,萃取液經乙醇沈降與 溶劑逐步脫水乾燥後共得 5.12 公克灰 白色棉絮狀的粗多醣,收率(yield)約 0.57%。將所的粗多醣經四級銨鹽去除 酸性多醣、amyloglucosidase 去除(1 →4)-α-D-glucan 及 protease 去除蛋白 質後,共得到純化靈芝多醣 GLPS 共 4.63 公克,收率 0.52%。以酚—硫酸法 檢測其總醣量約 80.98%。 (二)靈芝葡聚糖的製備 取純化靈芝多醣 GLPS 利用不同 濃度的乙醇進行分級沈降,可得到大 (GLG)、中與小分子三個區分,各區 分的總醣量皆提高到 85%以上。 (三)靈芝葡聚糖的分析 為了確認萃取的靈芝多醣 GLPS 中 雜 質 干 擾 的 情 況 , 分 別 檢 測 hexosamine、uronic acid 與 protein 的含 量。Hexosamine 可能來自靈芝細胞壁 的主幹—幾丁質,uronic acid 可能來自 酸性多醣,而蛋白質可能會附著在多醣 上一起被萃取沈降出來,結果在 GLPS 中三者的含量皆少(trace)。各靈芝多 醣區分以三氟醋酸完全水解後,利用 HPLC 檢測各單醣組成與相對含量比 例,發現主由三種單醣所構成:葡萄 糖、半乳糖與甘露糖。大分子區分 GLG 主由葡萄糖所組成(94%)。中與小分 子區分除了葡萄糖外,都含有相當程度 的半乳糖與甘露糖(表一)。依此結果看 來,單純為 glucan 的大分子靈芝葡聚 醣(GLG)結構特徵較接近靈芝活性多 醣(Sone 等人,1985)。 利用 13 C-NMR 來確認靈芝多醣 鍵 結 形 式 。 使 用 標 準 品 — 直 鏈 狀 (curdlan 與 pachyman)與分支狀(1 → 3 ) - β -D-glucan ( laminarin 與 lentinan),發現(1→3)-β-D-glucan 特徵波峰:C-1、C-2、C-3、C-4、C-5 與 C-6 的化學位移(chemical shift)分 別出現在:103.5、73.1、86.1、68.6、 76.4 與 61.1ppm。除主幹上六根波峰 外,還有分支 spliting 造成的其他波 峰:`C-1’:103.0ppm、C-2’:72.9ppm、 C-3’:85.8ppm、C-4’:68.6ppm、C-5’: 75.0ppm 與 C-6’: 68.6ppm ; C-1”: 103.0ppm 、 C-2” : 73.7ppm 、 C-3” : 76.6ppm、C-4”:70.3ppm、C-5”:76.2ppm 與 C-6”:61.0ppm(Saito 等,1977; 1987)。將 GLG 的圖譜比對標準品(圖 一),發現與 lentinan 類似,推測 GLG 為 ( 1 → 6 ) branched ( 1 → 3 ) - β -D-glucan,亦有相當程度的分支。其他 多醣區分並非單純由葡萄糖所組成,其 13 C-NMR 圖譜 broadening 非常厲害, 無法判讀比對。 1 H-NMR 在(1→6)branched (1 →3)-β-D-glucan 結構上的解析直至 2000 年才有重大進展。參照 Kim 等使 用 600HMz 的高頻核磁共振儀檢測靈 芝葡聚糖 GLG,並比對(1→3)-β -D-glucan 標 準 品 laminarin 與 lentinan,發現不同位置構成單元(稱為 anhydrous glucosyl units / AUGs) H-1 出 現 的 位 置 RT : α form 5.02ppm 、 β form 4.43ppm ; SRT : 4.52ppm;BC:4.57ppm;NRT:4.46ppm 與 TSC:4.27ppm(圖二)。利用面積 比 例 可 計 算 聚 合 度 ( degree of polymerization / DP)與分支度(degree of branching / DB),公式如下:
DP=all H-1 peak area / RT(or NRT) H-1 peak area
DB=TSC H-1 peak area / all H-1 peak area in backbone 如圖三(a)所示,lentinan 的分 支度為 0.42(7147 /(7147+10000)), 此值與文獻值 0.40 相近(Sasuki 與 Takasaka,1976)。由於該多醣分子量 巨大,相形之下 RT 與 NRT 的量少, 不見其波峰,故無法計算 DP。由圖三 (b)計算 GLG 的分支度為 0.35(5787 / (5787+10755)),此值與文獻值(0.33) 相近(Sone 等,1985),並座落在(1 →6)branched(1→3)-β-D-glucan 有 效的分支度範圍 0.2~0.5 內(Bohn 和 BeMiller,1995),推測 GLG 即是靈芝 中的(1→3)-β-D-glucan,且同樣不 見 RT 或 NRT 的波峰,無法計算 DP, 推測其分子量很大。 (四)(1→3)-β-D-glucan 螢光檢測 方法之建立:
1.(1→3)-β-D-glucan 與 aniline blue 錯合物的全光譜掃瞄:
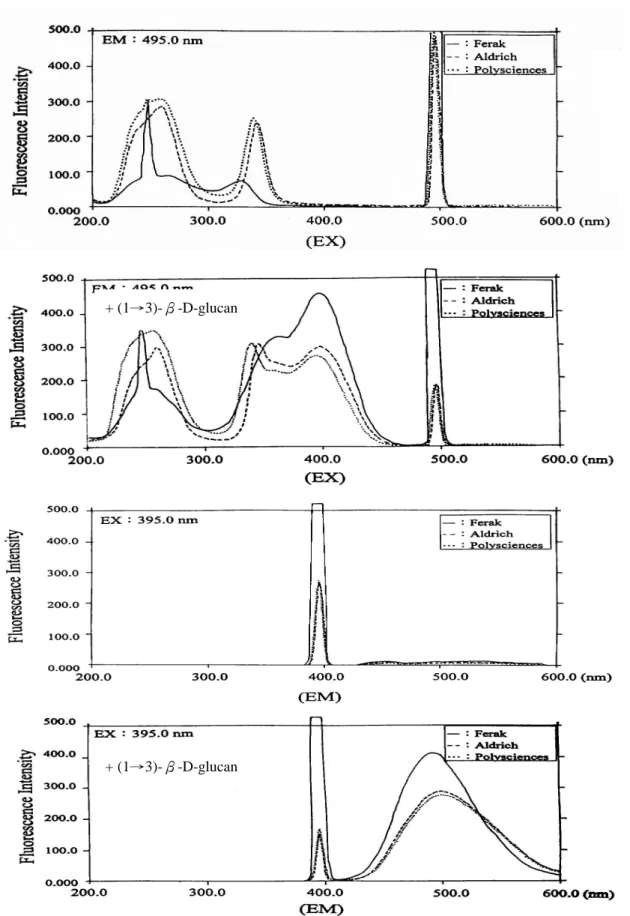
(1→3)-β-D-glucan—curdlan 與 aniline blue 作 用 後 , 在 放 射 波 長 (emission wavelength / EM)固定為 495 nm 的情況下,激發圖譜除了原先 的三個波峰外,在 395 nm 附近出現了 一個很明顯波峰。在激發波長固定為 395 nm 的情況下,放射波長圖譜除了 原先的 396 nm 外,在 495 nm 出現了一 個大的明顯波峰(圖四)。由此得知激 發波長 395 nm 與放射波長 495 nm 是(1 →3)-β-D-glucan 與 aniline blue 錯合 物的特徵螢光光譜波長,將之利用在 aniline blue assay 的檢測。
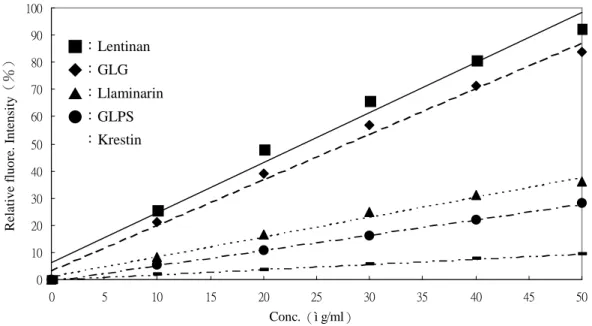
三種廠牌 aniline blue 產生的螢光 強度由強至弱依序為:Ferak >Aldrich > Polysciences , 此 結 果 與 Wood 與 Fulcher(1984)等人認為 Polysciences 產生的螢光強度最強不同,這可能是可 與(1→3)-β-D-glucan 錯合產生螢光 的 sirofluor 含量不同所造成。 由圖四可知 aniline blue 本身也有 螢 光 的 特 性 , 使 用 三 種 不 同 廠 牌 的 aniline blue(Aldrich / Milwaukee、Ferak / Berlin 與 Polysciences / Warrington)依 流程進行 aniline blue assay 後,進行螢 光全光譜的掃瞄,結果如圖四所示。未 添加 curdlan 之前,不論是何種廠牌的 aniline blue , 在 放 射 波 長 ( emission wavelength / EM)固定為 495 nm 的情 況 下 , aniline blue 的 激 發 波 長 (excitation wavelength / EX)圖譜主有 三個波峰:Aldrich—236.8、341.8 與 496.0 nm;Ferak—247.6、327.8 與 495.8 nm;Polysciences—255.0、339.4 與 496.0 nm。在激發波長固定為 395 nm 的情況 下,三個廠牌的放射波長皆只有在 396 nm 出現,因此不論使用何種廠牌的 aniline blue 均可清楚的判別染劑本身 與錯合物產生的螢光。 由 於 ( 1 → 3 ) - β -D-glucan 與 sirofluor 的作用是以單股的分子型態較 強,且多醣於溶液中需完全水和溶解, 故比較 30 與 90 分鐘的溶鹼時間對螢光 呈色強度的影響,結果發現兩者的螢光 強度沒有明顯區別,故樣品溶鹼時間即 訂為 30 分鐘。 2.線性範圍的檢測: 以螢光檢測法測試 curdlan、GLG、
laminarin、lentinan、pachyman 與含有 (1→3)-β-D-glucan 的靈芝多醣 GLPS 與 krestin 均發現有線性相關,且線性 範圍不盡相同。Curdlan 與 pachyman 分子量較大,線性範圍在 0~5μg/ml, 而 GLG、GLPS、krestin、laminarin 與 lentinan 的線性範圍在 0~50μg/ml。 由於(1→3)-β-D-glucan 標準品 的製備步驟繁雜,取得不易,因此建議 可用商業的 laminarin 為相對標準品, 測量值以 laminarin equivalant / LE 表 示。而(1→3)-β-D-glucan 生產廠商 也可用其標準品如 krestin equivalanr / KE 表示之。 3.反應溶液 pH 值對螢光呈色的影響: 依 Young 與 Jacobs 等人(1998) 的發現,在 pH 11.50 時(1→3)-β -D-glucan 與 aniline blue 錯合產生的螢 光最強,但為了嘗試提高 lentinan 的線 性範圍,分別將其最終溶液的 pH 調為 11.0、11.5 與 12.0 以進行 aniline blue assay,結果如圖五所示。pH 11.0 與 pH 11.5 兩組結果並沒有明顯差別,若以線 性關係≧0.97 為一基準點,兩組的線性 範圍皆在 0~30μg/ml 之間。pH 12.0 組 的 螢 光 值 明 顯 比 上 述 兩 組 弱 了 許 多,但線性範圍可延伸至 50μg/ml。由 此實驗得知,線性範圍與螢光強度成反 比,倘若有一 glucan 產生的螢光強度 太強,致使線性範圍太窄,那麼將檢測 環境的 pH 值調成 12.0 以獲取更廣泛的 線性濃度範圍不失為一個可行的方式。 4.Aniline blue 對不同β-D-glucan 的呈
色選擇性:
取(1→3)-β-D-glucan(curdlan、 GLG、laminarin、lentinan、pachyman) 與 ( 1 → 3 ; 1 → 4 ) mixed linked β -D-glucan(barley 與 oat glucan)為測 試標準品,並使用靈芝各區分多醣與 krestin 為測試材料,結果如圖六所示。 curdlan 與 pachyman 的線性範圍很窄(0 ~5μg/ml)。(1→3)-β-D-glucan 標準 品 krestin、laminarin 與 lentinan 進行螢 光檢測,發現各(1→3)-β-D-glucan 皆有明顯螢光,且濃度與螢光值呈線 性,但斜率明顯不同,各線性關係如表 二所示,推測分子量大小與分支度對線 性有影響。若取(1→4)-β-D-glucan 如 : oat- β -D-glucan 與 barley- β -D-glucan 進行試驗,發現其皆無明顯 螢光呈色,換句話說,aniline blue 只對 (1→3)-β-D-glucan 有專一性。 以 各 區 分 靈 芝 多 醣 進 行 aniline blue assay,發現僅有 GLPS 與 GLG 有 明顯螢光值並呈線性關係,且螢光值 GLG 遠比 GLPS 要來的強。至於中、 小分子靈芝葡聚糖則無明顯螢光值,推 測其非(1→3)-β-D-glucan。 5.干擾物質的影響: 螢光檢測法希望也可應用在液態 發 酵 產 品 上 , 故 常 用 培 養 基 組 成 分 (dextrose、glucose、malt extract、potato dextrose broth 、 peptone 與 yeast extract)對此法的影響也進行檢測。在 laminarin 各濃度標準液中添加濃度為 1000μg/ml 與 5000μg/ml 的不同培養 基組成分,檢測其對 laminarin 標準線 的影響。結果如圖七所示。在培養基組 成添加濃度為 1000μg/ml 時,發現各 組成分對標準曲線的影響不明顯,但當 濃度增加為 5000μg/ml 時,相對差異 即顯現,影響程度由大致小分別是: yeast extarct > malt extract > PDB ≒ peptone>glucose。Yeast extract 與 malt extract 皆為粗萃取物,成分複雜,推測 原因是反應液中暗褐色物質對 495nm 波長有吸收而降低螢光呈色值,為避免 暗沈顏色的干擾,需在測試前將顏色漂 白。而 peptone 與 PDB 在高濃度時會 干擾螢光呈色,詳細原因並不清楚,可 能是阻礙了 aniline blue 與 laminarin 的 結合所致。
(五)(1→3)-β-D-glucan 分支之酵 素—高效陰離子交換層析測定
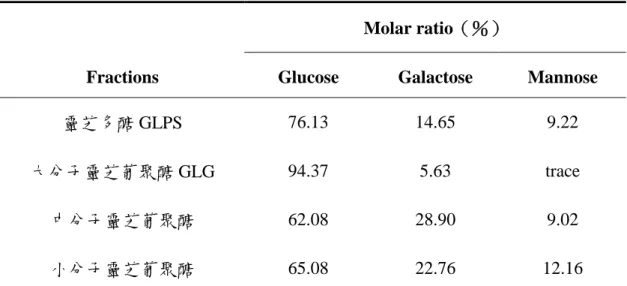
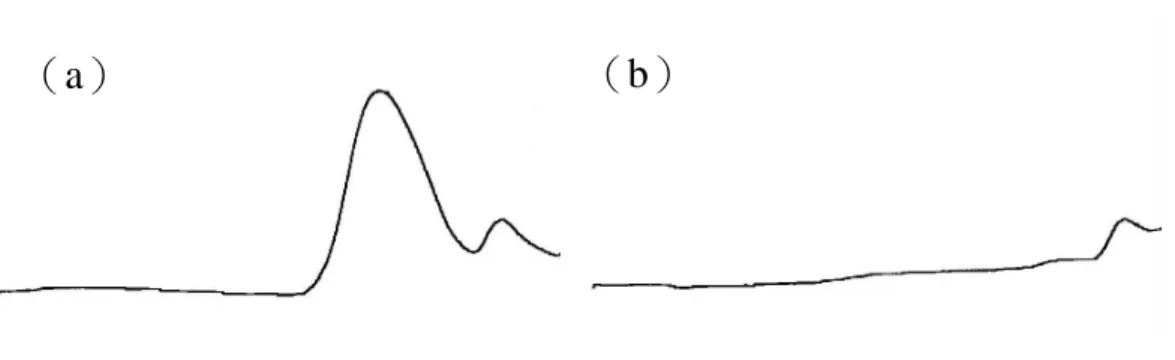
Megazyme exo 與 endo-(1→3)-β-D-glucanase 共用對 lentinan 有良好 的水解效果,在分子篩層析中可見其波 峰明顯消失。將水解液打入 HPAEC 進 行檢測,由葡萄糖與龍膽雙糖的比例計 算其分支度約為 0.34。 利用 0.3N 的 NaOH 使大分子靈芝 葡聚醣 GLG 構形鬆散、完全溶解後, 以 1N HCl 調整 pH 至 6.5,再利用 Megazyme endo + exo- ( 1 → 3 ) - β -D-glucanase 共同水解,水解效果如圖 八所示。在分子篩層析中,GLG 主要 波峰(高分子部分)在酵素水解後明顯 消失,另有較小分子物質波峰,在酵素 處理後仍然存在,推測其非(1→3)-β-D-glucan。由 HPAEC 圖譜(圖九) 知 GLG 酵素水後的產物主為葡萄糖與 龍膽雙糖,經過計算其相對莫耳比例為 龍膽雙糖/葡萄糖=1/3(molar ratio),分 支度約為 0.25,此結果與 NMR 圖譜所 得結果(分支度 0.33)有差距,可能是 靈芝葡聚醣 GLG 被酵素水解成單醣或 雙醣外,還有寡醣(如 laminatriose) 的存在而未納入計算,而在 NMR 上所 有非還原端均會納入計算,因此兩者結 果稍有差異。 靈芝多醣經酵素作用依分子篩層 析波峰面積的消減推算其約含 28%的 (1→3)-β-D-glucan。酵素對中、小 分子靈芝葡聚糖幾無作用,由單醣組成 分析(表一)知兩者主要成分仍為葡萄 糖(約 60%),推測便是葡萄糖相互鍵 結的長度並不長,在幾個葡萄糖鍵結後 即插入半乳糖或甘露糖,造成酵素無法 作用,或其中所含β-(1→3)-glucosidic linkage 含量不高所致。 (六)靈芝子實體栽植與深層發酵培養 所生多醣的差異 子 實 體 與 液 態 發 酵 培 養 靈 芝 (Ganoderma lucidum)菌株 GL4 與 GL11 所獲得之熱水萃取物,以酚—硫 酸法檢測其粗多醣量,再以 aniline blue assay 檢測其中(1→3)-β-D-glucan 的 含 量 , 因 使 用 商 品 化 的 ( 1 → 3 ) -β-D-glucan—laminarin 為標準品,故以 laminarin equivalent(LE)表示(1→3) -β-D-glucan 的檢測量。每公克 GL4 與 GL11 子實體的熱水萃取粗多醣量為 6.07 及 11.32mg , 其 中 ( 1 → 3 ) -β-D-glucan 含量為 0.28 與 0.39meq。 GL4 與 GL11 以發酵方式生成的粗多醣 量在 0.51~0.89meq/ml 間,無論發酵槽 容量為 5 升或 20 升,GL4 生成的(1 →3)-β-D-glucan 量均高於 GL11,GL4 以 5 升及 20 升發酵槽培養所生成的(1 → 3 ) - β -D-glucan 分 別 為 0.21 與 0.24meq/ml , GL11 則 只 有 0.08 及 0.03meq/ml。再利用分子篩層析檢測分 子量的分佈情形,發現多醣及所含(1 →3)-β-D-glucan 的分子量分佈非常 寬,分佈情況亦受培養方式與發酵槽容 量的影響,5 升發酵槽可獲得較多量大 分子量的(1→3)-β-D-glucan。實驗結 果顯示不同菌株、培養方式與條件對(1 →3)-β-D-glucan 的含量與組成有明顯 的影響,為製程中必須監控的重要因 子。
五、計畫成果自評:
本研究自靈芝子實體中萃取純化 出靈芝中性多醣,並確認其中大分子區 分結構為(1→6)branched(1→3) -β-D-glucan,分支度約為 0.33,以其為 標準品,再利用螢光染劑 aniline blue 中 的 ”sirofluor” 會 與 ( 1 → 3 ) - β -D-glucan 產生螢光的特性建立了(1→ 3)-β-D-glucan 螢光檢測定量方法, 此法已可將焦點由樣品中的「總多醣」 縮小至樣品中的(1→3)-β-D-glucan, 實驗也進一步評估其檢測液態培養發酵液中活性多醣的應用性。另外也針對 會影響(1→3)-β-D-glucan 活性的因 子—分支度建立了酵素—高效陰離子 將換層析法,可同時定性與定量。應用 這些方法監控不同培養方法與條件培 養靈芝所生多醣結構與含量的差異,獲 得良好的結果。
六、參考文獻:
王伯徹、陳啟楨、華傑。1998。食藥用 菇類的培養與應用。財團法人食品 工業發展研究所。 王聲遠,許敏玲,李旭生和蕭明熙。 1996。靈芝與雲芝免疫增強作用之 研究 行 政 院 衛 生 署 中 醫 藥 年 報 12(3): 257-275。 黃仁彰。2000。菇類多醣製劑的研發與 應用。CCRC News 42: 1-7。Adachi, Y., Miura, N. N., Ohno, N., Tamura, H., Tanaka, S., and Yadomae, T. 1999. Enzyme immunoassay system for estimating the ultrastructure of (1,6)-branched (1,3)-â-glucans. Carbohy. Polym. 39: 225-229.
Bao, X. F., Wang, X. S., Dong, Q., Fang, J. N., and Li, X. Y. 2002. Structural features of immunologically active polysaccharides from Ganoderma lucidum. Phytochemistry 59:
175-181.
Bitter, T., and Muir, H. M. 1962. A modified uronic acid carbazole reaction. Anal. Biochem. 4: 330-334. Blumenkrantz, N., and Hansen, G. A.
1973. New method for quantitative determination of uronic acids. Anal. Biochem. 54: 484-489.
Cataldi, T. R. I., Campa, C., and Benedetto, G. E. 2000. Carbohydrate
analysis by high-performance anion-exchange chromatography with pulsed amperometric detection: the potential is still growing. Fresenius J. Anal. Chem. 368: 739-758.
Chen, J., Zhou, J., Zhang, L., Nakamura, Y., and Norisuye, T. 1998. Chemical structure of the water-insoluble polysaccharide isolated from the fruiting body of Ganoderma lucidum.
Polym. J. 30(10): 838-842.
Dubois, M., Gilles, K. A., Hamilton, J. K., Rebers, P. A., and Smith, F. 1956. Colorimetric method for determination of sugars and related substances. Anal. Chem. 28(3): 350-356.
Eschrich, W, and Gurrier, H. B. 1964. Identification of callose by its diachrome and fluorochrome reactions. Stain Technol. 39: 303-307. Evans, N. A., and Hoyne, P. A. 1984.
Characteristics and specificity of the interaction of a fluorochrome from aniline blue (siroflour) with polysaccharides Carbohy. Polym. 4: 215-230.
Hong, T. Y., Cheng, C. W., Huang, J. W., and Meng, M. 2002. Isolation and biochemical characterization of an endo-1,3-â-glucanase from Streptomyces sioyaensis containing a
C-terminal family 6
carbohydrate-binding module that binds to 1,3-â-glucan. Microbiology
148: 1151-1159.
Hoogen, B. M., Weeren, P. R., Lopes-Cardozo, M., Golde, L. M. G., Barneveld, A., and Lest, C. H. A. 1998. A microtiter plate assay for the determination of uronic acid. Anal. Biochem. 257: 107-111.
Y., Hayase, S. 1997. Use of a minary mixture of quaternary ammonium salts in fluorometric determination of glycosaminoglycans. Anal. Biochem. 252: 56-61
Kim, H. S., Kacew, S., and Lee, B. M. 1999. In vitro chemopreventive effects of plant polysaccharides (Aloe barbadensis Miller, Lentinus edodes, Ganoderma lucidum and Coriolus versicolor). Carcinogenesis 20(8): 1637-1640.
Kim, Y. T., Kim, E Cheong, C., Williams, D. L., Kim, C. W. and Lim, S. T.
2000 Structural characterization of β-D-(1→3, 1→6)-linked glucans using NMR spectroscopy. Carbohy. Res. 328: 331-341.
Kotak, K., Schulte, A. S., Hay, J., and Sugden, J. K. 1997. Photostability of aniline blue (Cp 42755) and methyl blue (Ci42780). Dye Pigment. 34(2): 159-167.
Kupiec, R. C., Broglie, K. E., Friesem, D., Broglie, R. M., and Chet, I. 1999. Molecular characterization of a novel â-1,3-exoglucanase related to mycoparasitism of Trichoderma harzianum. Gene 226: 147-154.
Lee, Y. C. 1990. High-performance anion-exchange chromatography for carbohydrate analysis. Anal. Biochem. 189: 151-162.
Lownan, D. W., and Williams, D. L. 2001. A proton nuclear magnetic resonace method for the quantitative analysis on a dry weight basis of (1 → 3)- β -D-glucans in a complex, solvent-wet matrix. J. Agric. Food Chem. 49: 4188-4191.
Lowry, H., Rosebrough, N. J., Farr, A. L., and Randall, R. J. 1951. Protein measurement with the folin phenol
reagent. J. Biol. Chem. 1936: 265-275. Maeda, H., and Ishida, N. 1967.
Specificity of binding of hexopyranosyl polysaccharides with fluorescent brightener. J. Biochem. 62(2): 276-278.
Martens, D. A., and Frankenberger, W. T. 1991. Determination of saccharides in biological materials by high-performance anion-exchange chromatography with pulsed amperometric detection. J.
chromatogr. 546: 297-309.
Miyazaki, T., and Nishijima, M. 1981. Studies on fungal polysaccharides. XXVII. Structural examination of a water-soluble, antitumor polysaccharide of Ganoderma lucidum. Chem. Pharm. Bull.
29(12): 3611-3616.
Mizuno, T., Kato, N., Totsuka, A., Takenaka, K., Shinkai, K., and Shimizu, M. 1984. Fraction, structural features and antitumor activity of water-soluble polysaccharide from "Reishi", the fruit body of Ganoderma lucidum. Nippom Nogeikagaku Kaishi 58(9): 871-880.
Mizuno, T., Wang, G., Zhang, J., Kawagishi, H., Nishitoba, T., and Li, J. 1995. Reishi, Ganoderma lucidum and Ganoderma tsugae: bioactive substances and medicinal effects. Food Rev. Int. 11 (1): 151-166.
Morrow, D. L., and Lucas, W. J. 1986. (1 → 3)- β -D-glucan synthase from sugar beet. Plant physiol. 81: 171-176.
Ruiter, J. M., and Burns, J. C. 1987. Characterization of trifluoroacetic acid hydrolyzed subtropical forage grass cell walls. J. Agric. Food Chem.
35: 308-316.
Saito, H., Ohki, T., and Sasaki, T. 1979. A 13C-nuclear magnetic resonance study of polysaccharide gels. Molecular architecture in the gels consisting of fungal, branched (1→ 3)- β -D-glucans (lentinan and schizophyllan) as manifested by conformational changes induced by sodium hydroxide. Carbohy. Res. 74: 227-240.
Saito, H., Ohki, T., Takasuka, N., and
Sasaki, T. 1977. A
13
C-N.M.R.-spectral study of a gel-forming, branched (1 → 3)- β -D-glucan, (lentinan) from Lentinus edodes, and its acid-degraded fractions. Structure, and dependence of conformation on the molecular weight.Carbohy. Res. 58: 293-305. Sengbusch, P. V., Hechler, J., and Muller,
U. 1983. Molecular architecture of fungal cell walls. An approach by use of fluorescent markers. Eur. J. Cell Biol. 30: 305-312.
Shedletzky, E., Unger, C., and Delmer, D. P.1997. A microtiter-based fluorescence assay for (1,3)-â-D-glucan synthases. Anal. Biochem. 249: 88-93.
Smith, M. M., and McCully, M. E. 1978. Enhancing aniline blue fluorescent staining of cell wall structures. Stain technol. 53(2): 79-85.
Smith, M. M., and McCurry, M. E. 1978. A critical evalution of the specificity of aniline blue induced fluorescence. Protoplasma 95: 229-254.
Sone, Y., Okuda, R., Wada, N., Kishida, E., and Misaki, A. 1985. Structures and antitumor activities of the polysaccharides isolated from fruiting body and the growing culture of
mycelium of Ganoderma lucidum. Agric. Biol. Chem. 49 (9): 2641-2653. Stone, B. A., Evans, N. A., Bonig, I., and
Clarke, A. E. 1984. The application of sirofluor, a chemically defined fluorochrome from aniline blue for the histochemical detection of callose. Protoplasma 122: 191-195.
Suzuki, T., Ohno, N., Saito, K., and Yadimae, T. 1992. Activation of the complement system by (1 → 3)- β -D-glucans having different degrees of branching and different ultrastructures. J. Pharmacobio-Dyn.
15: 277-285.
Tanaka, S. 1993. Methods for determining (1 → 3)- β -D-glucan. United states patent 5266461.
Thistlethwaite, P., and Porter, I. 1986. Photophysics of the aniline blue fluorophore: a fluorescent probe showing specificity toward (1→3)-β-D-glucans. J. Phys. Chem.90: 5058-5063.
Wang, G., Zhang, J., Mizuno, T., Zhuang, C., Ito, H., Mayuzumi, H., Okamoto, H., and Li, J. 1993. Antitumor active polysaccharides from the Chinese mushroom Songshan Lingzhi, the fruiting body of Ganoderma tsugae. Biosci. Biotech. Biochem. 57 (6): 894-900.
Wang, Y. Y., Khoo, K. H., Chen, S. T., Lin, C. C., Wong, C. H., and Lin, C. H. 2002. Studies on the immuno-modulating and antitumor activities of Ganoderma lucidum(Reishi)polysaccharides: functional and protemic analyses of a fucose-containing glycoprotein fraction responsible for the activities.
1057-1062.
Wood, P. J. 1984. Specific interaction of aniline blue with (1→3)-β-D-glucan. Carbohy. Polym. 4: 49-72.
Wood, P. J., and Fulcher, R. G. 1980. Specificity in the interaction of direct dyes with polysaccharides Carbohy. Res. 85: 271-287.
Wood, P. J., and Fulcher, R. G. 1978. Interaction of some dyes with cereal â-glucans. Cereal Chem. 55(6): 952-966.
Wood, P. J., And Fulcher, R. G. 1983. Dye interactions: A basis for specific detection and histochemistry of polysaccharides. J. Histochem. Cytochem. 31(6): 823-826.
Yoshioka, Y., Uehara, U., and Saito, H. 1992. Conformatiom-dependent change in antitumor activity of linear and branched (1→3)-β-D-glucans on the basis of conformational elucidation by carbon-13 nuclear magnetic resonance spectroscopy. Chem. Pharm. Bull. 40 (5): 1221-1226.
表一、靈芝多醣各區分的單醣組成
Table 1 Sugar component of different GLPSs
Molar r atio(%)
Fr actions Glucose Galactose Mannose
靈芝多醣 GLPS 76.13 14.65 9.22
大分子靈芝葡聚醣 GLG 94.37 5.63 trace
中分子靈芝葡聚醣 62.08 28.90 9.02
圖一、(1→3)-β-D-glucan 的13
C-NMR 圖譜
Fig. 1. 13C-NMR spectra of various (1→3)-β-D-glucans.
(a)Curdlan.(b)Pachyman.(c)Laminarin.(d)Lentinan.(e)GLG. (a) Curdlan (b) Pachyman (c) Laminarin (d) Lentinan (e) GLG
—:β-(1→3)linkage 〉:β-(1→6)linkage
圖二、不同位置的 AUGs
Fig. 2. AUGs at different locations.
RT:reducing terminal H-1. NRT:nonreducing terminal H-1. SRT:the second AGU next to the reducing terminus H-1. TSC:the branched single or terminal AGU in the side chain. SC:all the AUGs except the terminal AUG in the side chain. BC:all the AUGs except the RT, SRT, and NRT in the backbone.
圖三、(a)Lentinan 與(b)GLG 的 1
H-NMR 圖譜 Fig. 3. 1H-NMR spectra of(a)lentinan and(b)GLG
(a)Lentinan
圖四、不同廠牌 aniline blue 添加(1→3)-β-D-glucan 前後的螢光光譜
Fig. 4. Fluorescence spectra of diffenent aniline blue with and without(1→3)-β -D-glucan.
+ (1→3)-β-D-glucan
Aniline blue assay at differnet pH 0 10 20 30 40 50 60 70 80 90 100 0 10 20 30 40 50 60 70 80 90 100 Conc.(ìg/ml)
Relative fluore. Intensity
(%)
圖五、Lentinan 在不同 pH 環境下的螢光檢測結果
Fig. 5. The effect of pH on the fluorescence intensity of lentinan. ■:at pH 11.0
◆:at pH 11.5 ▲:at pH 12.0
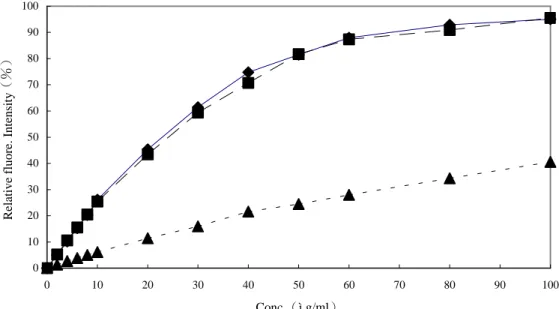
Aniline blue assay of various(1→3)-â-D-glucans 0 10 20 30 40 50 60 70 80 90 100 0 5 10 15 20 25 30 35 40 45 50 Conc.(ìg/ml)
Relative fluore. Intensity
(%)
圖六、不同 glucan 的 aniline blue assay 檢量線
Fig. 6. Standard curves of various(1→3)-β-D-glucans.
表二、各多醣濃度與 aniline blue 呈色反應回歸方程式與回歸係數 Table 2. Functions and R2of various (1→3)-β-D-glucans
Glucan Function R2 Curdlan* Y=37.037X+10.106 0.98 GLPS-F1 Y=0.0167X+0.0344 0.99 Krestin Y=0.0019X+0.0001 0.99 Laminarin Y=0.0073X+0.0112 0.99 Lentinan Y=0.0184X+0.0614 0.98 Pachyman** Y=10.941X+2.85 0.99 *:Curdlan 的回歸範圍為 0~4μg/ml。**:pachyman 的回歸範圍為 0~10μg/ml, 其他多醣回歸範圍為 0~50μg/ml。 ■:Lentinan ◆:GLG ▲:Llaminarin ●:GLPS :Krestin
0 5 10 15 20 25 30 35 40 45 50 0 10 20 30 40 50 Conc.(ì g/ml)
Relative fluore. Intensity
(%) laminarin+glucose laminarin laminarin+PDB laminarin+peptone laminarin+malt extract laminarin+yeast extract (a)1000μg/ml 0 5 10 15 20 25 30 35 40 45 50 0 10 20 30 40 50 Conc.(ì g/ml)
Relative fluore. intensity
(%) laminarin+glucose laminarin laminarin+peptone laminarin+PDB laminarin+malt extract laminarin+yeast extract (b)5000μg/ml
圖七、不同培養基組成分對 aniline blue assay 中 laminarin 標準曲線的影響(a) 組成分濃度為 1000μg/ml.(b)組成分濃度為 5000μg/ml
Fig.7. Fluorescence assay interfered with various media components.(a)at 1000μ g/ml. (b)at 5000μg/ml.
圖八、Megazyme endo-與 exo-(1→3)-β-D-glucanase 共用對 GLG 的作用 Fig. 8. The efficiency of Megazyme exo and endo-(1→3)-β-D-glucanase on
GLG(a)before(b)after.
圖九、GLG 經 Megazyme endo-與 exo-(1→3)-β-D-glucanase 共同作用後之 HPAEC 圖譜
Fig. 9. The HPAEC chromatogram of GLG treated with Megazyme endo- and exo-(1 →3)-β-D-glucanase.