中國醫藥大學營養學系碩士班
碩士論文
樟芝抑制人類乳癌細胞轉移作用及機制探討
Anti-metastatic activity of Antrodia camphorata
in human breast cancer cells
研究生:黃依婷(Yi-Ting Huang)
指導教授:楊新玲 博士 ( Hsin-Ling Yang, Ph.D. )
共同指導教授:許游章 博士(You-Cheng Hseu, Ph.D.)
目錄
謝誌………V 縮寫表 ………VI 中文摘要 ……… VIII Abstract………X 第一章 緒論 ………1 第一節 樟芝介紹 ………2 第二節 乳癌………11 第三節癌細胞轉移過程………13 第四節 基質金屬蛋白酶和轉移的相關性………15 第五節 基質金屬蛋白酶抑制劑和轉移的相關性………20 第六節 纖維蛋白溶解系統和轉移的相關性………20 第七節 血管新生與轉移的相關性………22 第八節 Mitogen-Activated protein Kinase ( MAPK )與轉移的相 關性………23
第九節 Nuclear factor kappa B (NF-kB)與轉移的相關性………26
第十節 研究動機………29 第二章 實驗設計架構圖………30 第三章 實驗器材………32 第四章 實驗方法………36 一、 樟芝發酵液 (AC-10)………37 二、 細胞培養………37 三、 細胞存活率分析(Cell viability)………38
四、 細胞遷移試驗(Wound migration assay)……… 39
六、 MMPs活性分析(Gelatin zymography)………41
七、 uPA 活性分析(Casein zymography)………42
八、 西方墨點(Western blotting)分析法……… 43 九、 抑制劑之處理………48 十、 NF-kB 活性測定(NF-kB reporter assay)………48 十一、 分析統計………49 第五章 實驗結果與圖表………50 第一節樟芝發酵液(AC-10)對 MDA-MB-231 細胞遷移情形與轉 移相關蛋白的表現 ………51 第二節 樟芝發酵液(AC-10)影響 MDA-MB-231 細胞的訊息傳遞 路徑與其可能調控的 MMP-9 表現 ………69 第六章 討論………81 第七章 結論………87 第八章 參考文獻………90
圖表目錄
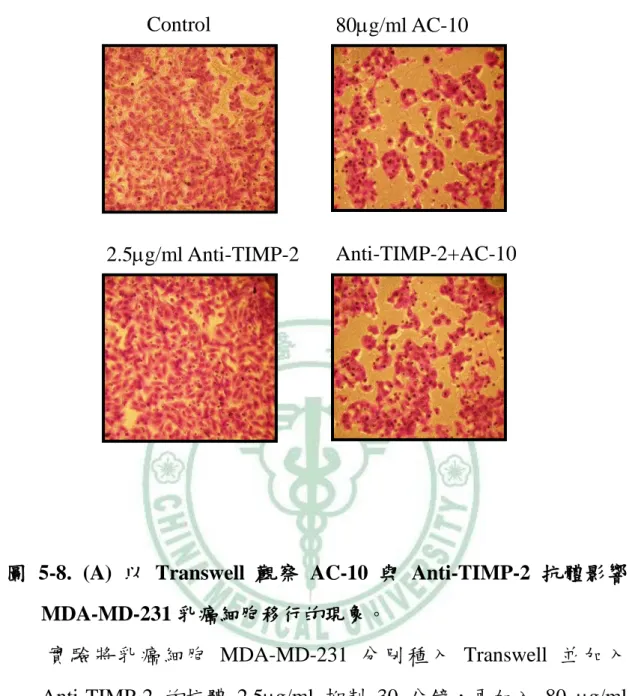
圖 1-1. 樟芝型態………2 圖 1-2. 樟芝子實體與菌絲體發酵液………6 圖 1-3. 乳癌分期 ………12 圖 1-4. 細胞外基質的組成 ………14 圖 1-5. 癌細胞的轉移過程………15 圖 1-6. MMPs的結構 ………16 圖 1-7. 蛋白分解酵素分解細胞外基質相關機制 ………21 圖 1-8. VEGF 促進血管新生與癌細胞轉移機制圖 ………23 圖 1-9. MAPK 與 IkB 影響 NF-kB 調控路徑 ………27 圖 1-10. NF-kB 調控相關基因表現 ………28 圖 4-1. 轉移盤(Transwell) ………41 圖 4-2. 蛋白質轉印 ………47 圖 5-1. 觀察 AC-10 對 MDA-MD-231 乳癌細胞存活率之影響………55 圖 5-2. AC-10 對 MDA-MD-231 乳癌細胞遷移的影響………56 圖 5-3. AC-10 對 MDA-MD-231 乳癌細胞移行的影響………57 圖 5-4. AC-10對MDA-MD-231細胞MMP-9和MMP-2蛋白表現 …58 圖 5-5. AC-10對MDA-MD-231乳癌細胞MMP-9和MMP-2的活性 59 圖 5-6. AC-10對MDA-MD-231乳癌細胞TIMP-1的蛋白表現………60 圖 5-7. AC-10對MDA-MD-231乳癌細胞TIMP-2的蛋白表現………61圖 5-8. 觀察 AC-10 與 Anti-TIMP-2 抗體影響 MDA-MD-231 乳癌 細胞移行的現象 ………62
圖 5-9. AC-10對MDA-MD-231乳癌細胞uPA的蛋白表現 …………64
圖 5-10. AC-10對MDA-MD-231乳癌細胞uPA的活性………65
圖 5-12. AC-10對MDA-MD-231乳癌細胞PAI-1的蛋白表現………67
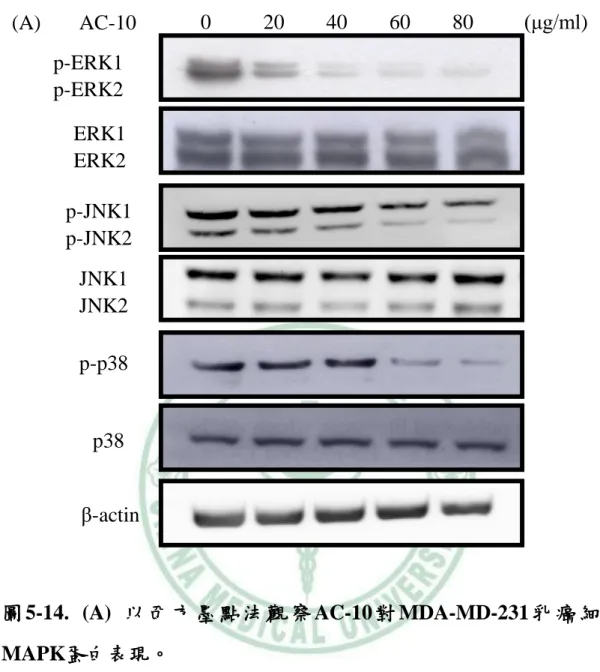
圖 5-13. AC-10 對 MDA-MD-231 乳癌細胞 VEGF 的蛋白表現 …68 圖 5-14. AC-10對MDA-MD-231乳癌細胞MAPK蛋白表現 ………72
圖 5-15. ERK抑制劑U0126與AC-10對MDA-MD-231乳癌細胞 MMP-9的蛋白表現 ………74
圖 5-16. ERK 抑制劑 U0126 與 AC-10 對 MDA-MD-231 乳癌細胞 MMP-9 的活性表現……… 75 圖 5-17. P38 抑制劑 SB203580 與 AC-10 對 MDA-MD-231 乳癌細胞 MMP-9 的蛋白表現……… 76 圖 5-18. P38 抑制劑 SB203580 與 AC-10 對 MDA-MD-231 乳癌細胞 MMP-9 的活性表現……… 77 圖 5-19. JNK 抑制劑 SP600125 與 AC-10 對 MDA-MD-231 乳癌細胞 MMP-9 的蛋白表現 ………78 圖 5-20. JNK抑制劑SP600125與AC-10對MDA-MD-231乳癌細胞 MMP-9的活性表現 ………79 圖 5-21. AC-10 影響 NF-kB 的轉錄活性 ………80 圖 7-1. 結論機制圖………89
表 1-1. Proximate composition in Antrodia camphorata ………3
表 1-2. 三萜類及多醣體有效成份說明……… 5
表 1-3. 乳癌的分期判斷………12
謝誌 忙碌也很充實的兩年研究生活,即將在完成這本論文畫上句點, 感謝指導老師楊新玲教授與共同指導老師許游章副教授這些日子的指 導,在學生實驗上遇到瓶頸時,能適時給予建議以及引領我們不同的思 考方向來解決問題,也特別感謝口試委員呂鋒洲老師、林一郎老師以及 王升陽老師對學生論文寶貴的建議。 另外,這段研究生活裡,謝謝雅婷、美存、依儒、皇琦、永昌與 智聖學長、學姊們,在百忙之中仍不斷教導我實驗的技術,給予我實驗 上莫大的幫助;謝謝我實驗室的同學康妮與彬鈞,一起進入實驗室從不會 到熟練,這段學習與做實驗的生活,感謝有你們的陪伴,遇到實驗不順 與挫折時,容忍我的情緒不穩定,還有幫忙我實驗的學弟妹盈真、書瑋、 雅雯與韻如…等等,讓我實驗能盡快完成,雖然研究的路程很艱辛,壓 力也常常隨著所面臨的問題而增加,但是還好有實驗室的大小夥伴們以 及其他實驗室的同學陪伴,不僅帶給我歡笑,也在我哭泣難過時,給予 安慰與鼓勵,讓我能拾回勇氣繼續向前邁進來完成這本論文。 還有永遠的避風港,感謝父母從小的培育,不在身邊的日子裡,依 然是依婷背後最大的支持與鼓勵,儘管面臨挫折與挫敗的心情,因為有 家人、師長與實驗室的同學與學弟妹在,讓我能秉持不放棄的態度、正 面的思考,順利完成學業。 黃依婷謹致 中國醫藥大學 營養系碩士班 中華民國九十八年七月
縮寫表
AC:Antrodia camphorata DMSO : dimethylsulfoxide
ERK:extracellular responsive kinase ECM:extracellular matrix
ELISA : enzyme-linked immunosorbent assay
EMSA: electrophoretic mobility shift assay
ERα:estrogen receptor alpha
FBS:fetal bovine serum
GFP: green fluorescent protein HRP : horseradish peroxidase IkB: inhibitor of nuclear factor-kB
JNK:c-Jun NH2-terminal kinase/stress-activated protein kinase MMPs:matrix metalloproteinase
MAPKs:mitogen activated protein kinases
MT-MMP: membrane-type matrix metalloproteinase MTT: thiazolyl blue tetrazolium bromide
NF-kB: nuclear factor kappa B
PAI-1:plasminogen activator inhibitor 1 PBS:phosphate-buffered saline
PBST : phosphate buffered saline containing-Tween-20 SDS : sodium dodecyl sulfate
SEAP :secreted alkaline phosphatase tPA:tissue-type plasminogen activator
TEMED: N,N,N,N-tetramethyl-ethylenediamine TIMP:tissue inhibitor of metalloproteinase
uPA:urokinase-type plasminogen activator
uPAR:urokinase-type plasminogen activator receptor VEGF : vascular endothelial growth factor
中文摘要
樟芝(Antrodia camphorata; AC)為台灣特有的藥用真菌,寄生於牛
樟樹上,是常見的傳統中藥,目前對樟芝的研究已顯示具有抗發炎、 抗氧化與抗腫瘤等功效。本實驗室已證實從樟芝發酵液獲得的AC-10 能抑制MDA-MB-231乳癌細胞的生長,誘導乳癌細胞凋亡,但是 AC-10對於抑制MDA-MB-231乳癌細胞轉移的相關機制仍不明確。癌 細胞轉移的過程中通常伴隨著細胞外基質(Extracellular matrix; ECM) 的 分 解 , 而 主 要 扮 演 分 解 ECM 的 角 色 為 基 質 金 屬 蛋 白 酶 (Matrix metalloproteinase;MMPs),以及促進MMPs活化的尿激酶型纖溶酶原 激活物(urokinase-type plasminogen activator ; uPA)。本研究先以MTT 測試細胞存活率,在對於細胞較不具毒殺性且存活率皆大於80%以上 的濃度下進行實驗。首先在細胞遷移試驗 (Wound migration assay)與 細胞移行試驗 (Transwell invasion assay)方法中顯示,隨著0、20、40、 60、80 g/ml的AC-10濃度,可以抑制乳癌細胞的遷移和移行能力。 另外,由西方墨點法結果顯示AC-10能降低MMP-9、MMP-2、uPA、 uPAR與VEGF的蛋白表現,以及MMP-9與uPA的活性。AC-10也增加 基質金屬蛋白酶抑制劑TIMP-1、TIMP-2與血漿纖溶酶原激活物抑制 物PAI-1的表現。進一步探討訊息傳遞,AC-10可以降低ERK1/2、P38 與JNK1/2的磷酸化,結果顯示加了ERK抑制劑UO126與P38抑制劑 SB203580可以降低MMP-9表現,若再加上 AC-10則能更顯著降低 MMP-9的表現,但是在加了JNK抑制劑SP600125與AC-10後,雖然也 有降低MMP-9的活性表現的趨勢,但是較不顯著。AC-10亦能降低 NF-kB的活性表現,因此,本實驗在觀察細胞遷移與轉移相關蛋白的 表現皆顯示AC-10具有降低乳癌細胞轉移的能力,且可能透過抑制 ERK1/2、P38與JNK1/2路徑向下調控核內NF-kB表現,並影響MMP-9
轉移因子的表現進而抑制乳癌細胞的轉移。
關鍵字:樟芝(Antrodia camphorata, AC), 乳癌(Breast cancer), 轉移 (Metastasis), 細胞外基質(Extracellular matrix), NF-kB(nuclear factor kappa B)
Abstract
Antrodia camphorata (AC) is well- known in Tiawan as a traditional
Chinese medicine. It has been shown to exhibit anti-inflammatory, antioxidation and anticancer properties. According to the previous data, AC-10 induced apoptosis in human breast cancer cells (MDA-MB-231). Breast cancer is a common cancer in women. In this study, we investigate anti-metastatic activity of AC-10. Degradation of extracellular matrix is crucial for malignant tumor development and metastasis. Treatment with AC-10 is effective in suppressing cancer cell migration and invasion. These effects were associated with a reduced protein expression of MMP-9, MMP-2, uPA, uPAR and VEGF together with an enhanced expression of TIMP-1, TIMP-2 and PAI-1. AC-10 decreased activities of MMP-9 and uPA, but not effect of MMP-2. Further studies showed that AC-10 regulated MMP-9 production via MAPK signaling pathway, as evidenced by the findings that AC-10 inhibited the phosphorylation of ERK1/2, P38 and JNK1/2. Additionally, pretreatment of MDA-MB-231 cancer cells with 10 and 30 M of U0126, a specific ERK inhibitor, and SB203850, a specific P38 inhibitor and SP600125, a specific JNK inhibitr resulted in a reduced activities of MMP-9. And AC-10 inhibited NF-kB transcriptional activity. These results suggested that AC-10 could reduce the invasion and migration via MAPK and NF-kB signaling pathways in MDA-MB-231 human breast cancer cells, and may use AC-10 as an anti-invasive agent in the prevention and treatment of breast cancer.
Keyworld:Antrodia camphorata, Breast cancer, Metastasis, Extracellular
第一章
第一節樟芝介紹 一、 樟芝簡介
樟芝學名為 Antrodia camphorata,簡稱 AC,亦有人稱為樟菇、
牛樟芝,生長於牛樟樹上,其具有濃厚的樟樹香氣,為台灣特有的 真菌,牛樟樹在台灣地理上的分佈包括桃園角板山、南庄三灣、南 投竹山、高雄六龜,這些地區樟芝的產量較豐富。樟芝的分類上, 是在 1995 年開始的,張東柱依據樟芝子實體型態及真菌的培養基特 性,認為是多孔菌科 Antrodia Karst 屬,因此,食品工業研究所菌種 中心才將樟芝拉丁文命為 Antrodia cinamomea,因此樟芝應屬於無褶 菌目、多孔科、多單生蕈菌類。樟芝型態上具有板狀與鐘狀 (陳等, 2001; 圖 1-1),板狀面呈現橘紅色,因此亦有人稱之為紅樟,表面 含有許多菌孔,面底部有淺黃白色的木栓質,經由木栓質附著在牛 樟樹中空心內壁上而生長,鐘狀形態者,子實層呈橘黃色,充滿菌 孔(4~5 個菌孔/mm),含有苦味的孢子,新鮮時為橘紅色,成熟 後會偏橘褐色,子實體則呈綠褐色。 (陳等; 2001)
二、 樟芝成分分析 (一) 一般成分 樟芝子實體之碳水化合物約含 31.38%,菌絲體部分則含量較 高約有 53.46%,子實體的蛋白質約 7.33%,而菌絲體約含 23.4% (陳 等;2001; 表 1-1),粗脂肪,包括游離脂肪酸、單 (雙或三)甘油酯、 固醇類、磷脂質等,含有 32.23%,此樟芝粗脂肪含量高於一般菇類 (2~8%),因此與樟芝能產生較高量精油有關 (黃; 2000),另外,在 維生素方面,菌絲體比子實體含有較多量的維生素 B6 與菸鹼酸, 但子實體則含有較多的維生素 B12。
表 1-1. Proximate composition in Antrodia camphorata
Assay of contents Fruit body Mycelia
Moisture 7.33% 4.48% Dry matter 92.67% 95.52% Ash 1.73% 9.68% Carbohydrate 36.91% 53.46% Crude protein 6.6% 23.84% Crude fat 32.23% 5.81% Crude fiber 22.53% 7.21% Vitamin B1 5.3ppm 6ppm Vitamin B6 21.3ppm 76.9ppm Vitamin B12 286ng/g 165ng/g Vitamin C 3.8ppm 7.9ppm Niacin 2.6ppm 356.7ppm β-carotene N.D N.D (陳等; 2001)
(二) 有效成分
至今針對菇類成分的研究仍多著重在三萜類與多醣體上 (表 1-2),過去從靈芝及其菌絲體液體培養之菌絲體甲醇萃取物中分離 之三萜類衍生物已有超過八十多種。以HPLC分析發現,靈芝有十八 種不同類型的三萜類。三萜類的研究上證實具有抑制血小板凝集、 抗高血壓、降低血膽固醇及抗HIV病毒活性 (Kim et al; 1999)。文獻 也指出靈芝之三萜類靈芝酸 (ganoderic acid)對肝癌細胞 (hepatoma cells)具有細胞毒性之抗癌活性。靈芝子實體所分離之三萜類及固醇 類 具 有 誘 導 Hep3B 肝 癌 細 胞 計 畫 性 死 亡 (apoptosis) 之 細 胞 毒 性 (Gan et al; 1998)。此外,靈芝三萜類也有活化巨噬細胞及T淋巴球細 胞釋放細胞激素 (cytokine)之抗腫瘤能力,和抑制大鼠mast cell釋放 histamine之抗發炎能力。雖然三萜類之藥理作用仍不明確,但在化 學鑑定上,己成為靈芝在藥理學上品質及分類學上的一種指標物質 (Chen et al; 1999)。 有關樟芝有效成分的研究不多,多由樟芝甲醇或氯仿萃取物中 進 行 層 析 分 離 , 鑑 定 出 的 成 分 多 為 類 三 萜 類 結 構 , 包 括 17 種 4-α-methylergostane type,6種lanostane type及4種l,3-benzodioxloe化 合物。有文獻指出樟芝子實體中所含有之多氧化型之三萜類及固醇 類,可能是造成樟芝具有強烈苦味之主要成分 (吳; 1995)。樟芝子 實體甲醇萃取物可抑制金黃色葡萄球菌和鬚瘡小芽癬菌之生長及抗 膽鹼、腸弛緩運動及血小板凝集作用等,經純化鑑定具生物活性之 化合物有三萜類zhankuic acid A、B 及C,其中zhankuic acid A和C 對P38淋巴癌細胞株有細胞毒殺作用,而zhankuic acid A和B具有
anticholinergic及antiserotonergic的效果 (簡等; 1997)。目前已知多醣 體之研究多集中在抗腫瘤 (Wasser et al; 1999)、調節免疫 (Wang et
al; 1997)、抗泡疹、抗病毒 (Eo et al; 1999)等功能上。現已有研究發
現一些由菇類純化的多醣體具有抗腫瘤作用,均具有α-1,3 glucan (葡 聚糖)的結構。以X-射線繞射分析得知,這種以α-l,3 鍵結的D- glucose骨架呈現3股右旋之螺旋型結構,而有文獻指出此種螺旋結構 可能是抗腫瘤作用的重要結構 (Mizuno ; 1999)。 表1-2. 三萜類及多醣體有效成份說明 名稱 三萜類 (Triterpenoids) 多醣體 (Polysaccharide) 生化結構 1.樟芝苦味的來源。 2.由30個碳組成六角型或五角型的 有機化合物總稱。 3.發現總類200 種以上。 1.由數十萬到數百萬單醣 類組合而成。 2.屬於雜多醣。 3.有效成份為β-D葡聚醣。 生理活性 1.誘導癌細胞凋亡、抑制轉移。 2.抗發炎。 3.免疫調節功能。 4.降低肝功能GOT、GPT 指數。 5.降血壓。 1.刺激免疫系統巨噬細 胞、T 細胞、B 細胞及 自然殺手細胞活性,強化 免疫機能。 功能 1.抗癌。 2.消炎解熱鎮痛。 3.保護肝臟機能治療肝炎。 4.消除疲勞增強體力。 5.調節血壓預防中風。 1.抗癌。 2.預防感冒。 3.促進新陳代謝。
三、 樟芝深層培養 牛樟為台灣稀有珍貴的樹種,若樟樹上要長出樟芝的機會又 更加困難,因此樟芝在台灣屬於昂貴的中藥材,目前樟芝子實體 仍無法使用人工方式來栽培,但是現今利用固態與液態培養(深層 培養,submerged culture)樟芝菌絲體的技術已非常完善且普遍。 深層培養是指藉由固體組成的液態培養基在控制適當的 pH 值、溫度、空氣和震盪速度的條件下,進行微生物的發酵培養, 以製造生質量 (biomass)或是其他的代謝產物 (metabolites)。蕈類 的深層培養液最大的特色在於發酵過程中,所產生的菌絲體是以 圓形球狀物 (pellets)的形式懸浮於液體中,但不產生子實體的部 分。所以利用樟芝深層培養技術進行菌絲體液體培養,可以獲取 最大的菌絲體生成量 (王等; 1998),因此可以供應於市場對樟芝 的需求,降低珍貴牛樟樹被砍伐以及減少成本的消耗。 由於此培養技術上並無子實體的形成,與一般野生所採食用 之樟芝子實體有別,故為了解深層發酵法所產生之菌絲體是否仍 具有其功能性存在,多年來許多研究者利用樟芝子實體萃取物或 樟芝菌絲體發酵液的部分進行試驗 (圖 1-2),探討樟芝對生理的 功效。 樟芝子實體 菌絲體發酵液 (資料來源:葡萄王生技) 圖 1-2. 樟芝子實體與菌絲體發酵液
四、 樟芝功能性研究 (一) 樟芝安全性與毒性研究 (1) 以每日餵食3 g/kg樟芝菌絲體發酵液給予小鼠連續15天,對小 鼠並不具基因毒性,且對化學致癌物dacarbazine所誘發的 DNA損傷有明顯的拮抗作用,即有抗基因毒性的功能 (江等; 2002)。 (2) 連續經口投予樟芝發酵液28天,發現其安全劑量在2 g/kg以 下,血液及尿液生化學的指標均在安全的範圍內 (郭等; 2002)。 (3) 以乾燥菌絲體進行SD大鼠90天活體試驗,發現對肝臟、腎臟 及脾臟不具毒性 (林等; 2000)。 (4) 據急性毒性試驗,大鼠口服樟芝發酵液半數致死劑量 (LD50) 高於15 g/kg。在致畸試驗中,餵食SD雌鼠500 mg/kg/day乾燥 樟芝發酵液,其子宮重量、生育力指數、受精卵著床前流失 率、著床後死亡率及胎鼠平均體重均與控制組無顯著性差 異,由胎鼠之外觀、內臟及骨骼檢查結果顯示,乾燥樟芝發 酵液並不會造成任何畸胎現象 (江等; 2001)。 (5) 對於樟芝發酵液在高濃度下具有毒性的報告,以不同體積發 酵液與大白鼠初代肝細胞培養,發現在20 μl/ml之系統濃度 下,樟芝發酵濾液並不會造成毒性效應,但是以40或60 μl/ml 處 理 8 小 時 後 , 肝 細 胞 乳 酸 去 氫 酶 滲 漏 率 (lactate dehydrogenase leakage; LDH leakage)顯著高於控制組 (p < 0.05),當以60 μl/ml處理24小時,肝細胞會發生形狀變形 (shape deformation) 與 單 層 細 胞 崩 潰 (destruction of cell monolayer)的現象 (沈; 2000)。
(二) 樟芝抗氧化相關研究 (1) 在活體外系統中,樟芝子實體及菌絲體甲醇萃取物均具有清 除自由基及螯合金屬離子能力,且新鮮子實體之抗氧化力優 於乾燥子實體 (Huang et al., 1999)。 (2) 以紅血球以及人類臍帶血管內皮細胞探討樟芝菌絲體水萃取 液是否具有抗氧化及清除自由基的能力,結果顯示其可減少 紅血球溶血的現象,減少細胞死亡率,此外,發現其具有毒 殺血癌細胞 (HL-60cells)的能力,但對正常內皮細胞卻不具細 胞毒性,因此推論樟芝菌絲體可能具有保護性的抗氧化作用 以及抗腫瘤能力 (Hseu et al., 2002)。 (3) 發酵濾液及菌絲體中之多醣體均可藉由誘導胞內抗氧化防禦 能力,以清除胞內活性氧,因此可抑制脂質過氧化作用,減 少DNA的氧化傷害。 (4) 樟芝發酵液與菌絲體均可顯著提升血清總抗氧化狀態,可延 長AAPH誘導紅血球溶血的時間,增加LDL之氧化遲滯期,顯 著提升肝臟中glutathione peroxidase及SOD 酵素活性,因此推 測其具有保護高油飲食所誘發的氧化壓力功能 (劉; 2002)。 (三) 樟芝保肝相關研究 (1) 樟芝菌絲體發酵濾液具有提升SD大鼠初代肝細胞之抗氧化 酵素及抑制脂質過氧化能力。 (2) 相對地,在活體系統中,也有研究證實樟芝子實體與菌絲體 均有降低酒精所誘發之急性肝損傷的功能,而其保護機制應 與其所具有的抗氧化與清除自由基能力有關。以腹腔注射及 口服四氯化碳誘導大鼠慢性肝損傷後,樟芝菌絲體分別呈現
無或有保護肝臟的能力 (戴; 2001; 李等; 2002)。
(3) 在12.4及24.8 ml/kg B.W/day之劑量下,於四氯化碳投予前一 週,每日連續八週餵食樟芝菌絲體發酵濾液,直到實驗終了。 結果顯示樟芝菌絲體發酵濾液對四氯化碳誘發大鼠慢性肝損 傷有保護的能力, 其保護機制應與其提升抗氧化與解毒酵素 (glutathione peroxidase 、 glutathione reductase 及 glutathione S-transferase)活 性與抑制 肝臟與血漿脂質過 氧化能力有關 (陳等; 2001)。 (4) 以四氯化碳連續經口投予八週誘發慢性肝炎,由第四週開始 口服投予樟芝發酵液至實驗終了,由血清生化值及肝臟膠原 蛋白含量顯示,樟芝發酵液 (1 g/kg)具有改善四氯化碳誘發慢 性 肝 炎 的 效 果 。 除 此 外 , 在 2 g/kg 之 劑 量 下 , 能 抑 制 dimethylnitrosamine 所引起的肝臟纖維化。 (四) 樟芝抗癌相關研究 (1) 曾有報告指出樟芝菌絲體之甲醇萃取物在低劑量下,對肝腫 瘤細胞株HepG2即有很強的細胞毒殺作用,其次為水萃取部 份,而發酵液則幾乎不具毒殺能力;但在高劑量 (2000 μg/ml) 下,均對正常人類白血球細胞無毒殺現象。 (2) 以不同濃度之樟芝菌絲體甲醇萃取物測試對不同肝癌細胞株 (Hep3B、J5、J2、G2及SK-Hep-1)的抑制生長能力,以MTT 方法分析結果發現,對Hep G2的影響最大;研究結果指出, 樟芝菌絲體之甲醇萃取物並不會造成Hep G2細胞壞死的現 象,進行細胞週期分析發現,在低濃度 (62.5 ppm)時會使細 胞走向G1 arrest,而高濃度 (250 ppm)時會使細胞走向G2/M
arrest (張等; 2001)。
(3) 無 論 是 經 由 腹 腔 注 射 或 餵 食 樟 芝 菌 絲 體 多 醣 , 均 對 植 入 Sarcoma 180的小鼠腫瘤具有劑量性地抑制能力,並證實此能 力與提升免疫細胞的活化有關 (呂; 2002)。
(4) 樟 芝 發 酵 液 可 抑 制 經 LPS (lipopolysaccharide) 刺 激 之 RAW 264.7巨噬細胞所產生的iNOS (inducible nitric oxide synthase) 以及COX-2 (cyclooxygenase-2)蛋白質之表現,並減少其下游 產物NO (nitric oxide)與PGE2 (prostaglandin E2)之生成,因此 證實樟芝發酵液具有抗發炎之作用 (Hseu et al., 2002)。 (5) 樟芝發酵液處理之HL-60血癌細胞,會造成HL-60細胞之生長 抑制,具有劑量性與時間性的抑制能力,並且能造成HL-60 血癌細胞之細胞凋亡 (Hseu et al., 2004)。 (6) 樟芝發酵液處理之MCF-7乳癌細胞,會造成MCF-7乳癌細胞 之生長抑制,具有劑量性與時間性的抑制能力,並且能造成 MCF-7乳癌細胞之細胞凋亡 (Yang et al., 2006)。 (7) 樟芝子實體萃取物能經由降低人類肝癌細胞株PLC/PRF/5核 內NF-kB的表現而抑制癌細胞的轉移能力 (Hsu et al., 2007)。 (8) 樟芝粗萃取物 (Antrodia camphorata crude extract; ACCE)能
降低膀胱癌細胞株(RT4, TSGH-8301, T24)的生長,並且可以 顯著抑制具有高轉移性的 T24細胞遷移能力 (Peng et al., 2007)。
第二節乳癌 一、 癌症的發生 癌症的發生會隨著個人的性別、年齡、飲食、生活習慣、遺傳、 種族以及環境等所影響,根據衛生署統計資料顯示,民國 97 年國人 的十大死亡原因之首,仍為惡性腫瘤,如何有效預防與治療癌症依舊 是現今重要的課題之一。 腫瘤 (tumor)是由身體內某些細胞或是組織發生不正常或是自發 性且無法控制的增殖,因而形成了細胞族群或組織團塊。腫瘤可依照 臨床行性,將腫瘤簡單分為兩種: (1)良性腫瘤 (benign tumor): 腫瘤細 胞大多分化良好,與來源細胞較為相似,大致上較不易引起病患死亡 的腫瘤; (2)惡性腫瘤 (malignant tumor): 惡性的腫瘤統稱為癌症 (cancer),其腫瘤細胞生長較良性腫瘤快速,可能分化良好或分化較 差,且與來源細胞的差異較大,惡性腫瘤與正常細胞的差異包括:細 胞 與 細 胞 核 的 多 形 性 (pleomorphism) 、 細 胞 核 濃 染 (hyperchromatism)、細胞核與細胞質的比例 (核質比)增加、有絲分裂 增加以及不正常的絲分裂 (abnormal mitosis)等。惡性腫瘤的預後較差 是由於腫瘤的邊界不清楚且不規則,周圍沒有纖維性囊被 (capsule) 包圍著,因此容易隨著腫瘤的生長而對周圍的組織產生侵入或侵襲 (invasion)的現象,造成癌細胞的轉移。 二、 乳癌的發生 乳癌好發於中老年婦女,在衛生署 97 年的統計資料顯示癌症主 要死亡原因,女性乳癌排第四位,相關因素包含:遺傳 (腫瘤抑制基因 BRCA1 異常)、內分泌 (如:動情素產生過多)、環境 (飲食習慣及歐 美生活型態等)、乳房病變等。
腫瘤的轉移方式可分為四種:直接蔓延、血行轉移、淋巴轉移和 播種轉移。而乳癌最容易轉移至同側的腋下淋巴結,以及內乳淋巴結 和鎖骨上淋巴結,另外,也可能侵犯乳房表面皮膚或鄰近的軟組織、 肌肉,甚至經由血液循環轉移到其他的器官。 三、 乳癌的分期與治療 依據美國癌症分期聯合委員會分法,乳癌分第 0 期到第四期 (表 1-3、圖 1-3): 表 1-3. 乳癌的分期判斷 第 0 期 原位癌 第一期 侵襲癌<2 公分 (無淋巴結及遠處轉移) 第二期 侵襲癌<5 公分 (有腋下淋巴結轉移,但無遠處轉移) 侵襲癌>5 公分 (無淋巴結及遠處轉移) 第三期 侵襲癌>5 公分 (有淋巴結轉移) 第四期 任何有遠處轉移的乳癌 (資料來源: http://www.novartis.com.tw) 圖 1-3. 乳癌分期 在 西 方 國 家 臨 床 上 表 示 將 乳 癌 細 胞 上 (estrogen receptor alpha)ERα的有無,作為乳癌患者預後的重要指標 (Brinkman et al.,
會阻礙內分泌治療方式以及有較差的預後,因此許多研究者進行介 入試驗期望能找出讓ERα (-)的乳癌細胞進行修補產生ERα,以利於 治療。依臨床診斷所採取治療的方式包括:手術直接切除、淋巴腺摘 除、荷爾蒙治療、放射線治療與化學治療。 第三節癌細胞轉移過程 癌細胞的轉移伴隨著細胞外基質的降解,以下先介紹細胞外基 質的組成。 一、細胞外基質之組成與功能
細胞外基質 (Extracellular matrix; ECM)主要成分可分為三類 (圖 1-4): ( 一 ) 纖 維 結 構 蛋 白 : 包 括 膠 原 蛋 白 (collagen) 、 彈 性 蛋 白 (elastin)、基膜粘連蛋白 (laminin),和纖維醣連蛋白 (fibronectin)。其 中膠原蛋白為最重要的成分。這些蛋白為巨長的大分子,形成堅韌的 三股螺旋結構纖維。(二)多醣: 透明質酸 (hyaluronan)。(三)蛋白多醣: 含有糖胺多糖的糖蛋白與硫酸肝素的糖蛋白。其功能包括提供細胞的 固定與支持作用,以及調節細胞間的溝通。而細胞與細胞外基質結合 處含有許多黏附因子如粘合素 (integrin)可以傳遞細胞與細胞間各項 訊息。 在胚胎發育和組織修復等過程中,細胞會釋放一些蛋白分解酵 素 , 分 為 四 類 : ( 一 ) 基 質 金 屬 蛋 白 酶 (matrix metalloproteinase, MMP ),(二)Serine 蛋白酶,如:纖溶酶原激活物 (plasminogen activator , PA)、plasmin 等,(三) cysteien 蛋白酶,cathepsin B、cathepsin L,(四) aspartyl 蛋白酶,cathepsin D,皆能將 ECM 分解,因此適量的蛋白分 解酵素可以維持正常的生理功能,但是,當蛋白分解酵素的分泌過量 時,會導致疾病的發生,例如發炎反應、血管新生 (angiogenesis)、
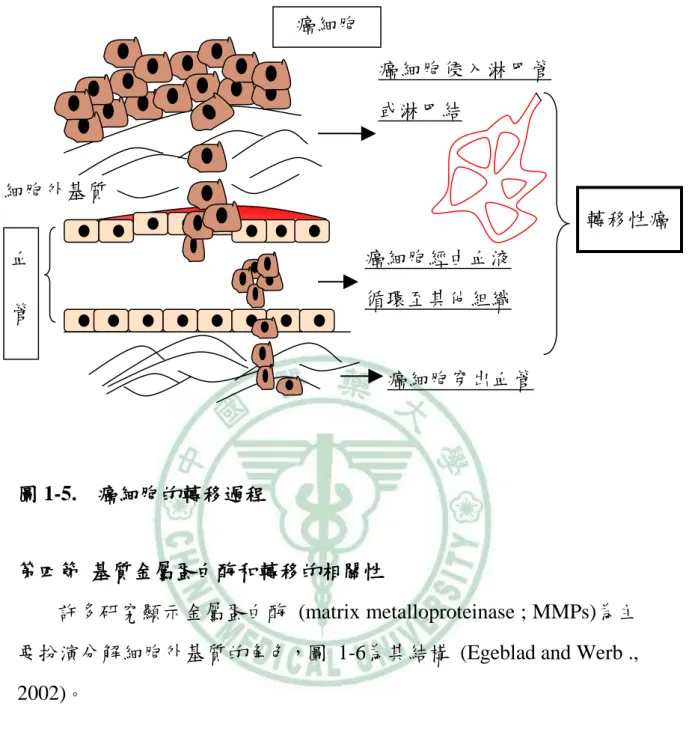
心血管疾病、動脈粥狀硬化的過程 (Galis & Khatri ., 2002)、癌細胞 的生長、轉移和入侵等,都和蛋白分解酵素過度表現有關。 資料來源: (http://www.bbioo.com) 圖 1-4. 細胞外基質的組成 二、癌細胞轉移 造成乳癌患者的死亡原因大都是癌細胞轉移至其他組織和器 官所引起,癌細胞的轉移過程是相當複雜的,包括細胞的黏附 (adhesion)、侵入 ( invasion)、遷移 (migration) (Liotta et al., 1986), 癌細胞轉移過程中需要蛋白分解酶分解細胞外基質 (extracellular matrix ; ECM) (Stetler-Stevenson et al., 1993),癌細胞通過 ECM 後 經由淋巴系統或血液循環轉移至其他組織 (圖 1-5)。
圖 1-5. 癌細胞的轉移過程
第四節 基質金屬蛋白酶和轉移的相關性
許多研究顯示金屬蛋白酶 (matrix metalloproteinase ; MMPs)為主 要扮演分解細胞外基質的角色,圖 1-6為其結構 (Egeblad and Werb ., 2002)。 癌細胞侵入淋巴管 或淋巴結 癌細胞經由血液 循環至其他組織 穿出血管 轉移性癌 血 管 癌細胞穿出血管 細胞外基質 癌細胞
(Egeblad and Werb., 2002) 圖1-6. MMPs的結構
依據在ECM成分中MMPs可被分成不同的種類 (Mook et al., 2004),另外有其他未被分類的蛋白分解酵素 (Creemers et al., 2001; 表1-4)。
(1) 膠原蛋白酶 (Collagenases)
這 類 分 解 酶 主 要 降 解 纖 維 型 的 膠 原 蛋 白 , 包 括 interstitial collagenase (MMP-1) 、 neutrophil collagenase, collagenase-2 (MMP-8)主要降解type I collagenase,collagenase-3 (MMP-13)為降 解 type I I collagenase,另外還有collagenase-4 (MMP-18)。
(2) 明膠分解酶 (Gelatinases)
這類的分解酶主要降解type IV collagen,其富含於基底膜中, 譬如gelatin A (MMP-2)與gelatin B (MMP-9)。Gelatinases的催化位 domain含有三種胜肽序列,其結構與fibronectin一致,當gelatinases 活化時,而能結合fibronectin並將其進行分解。
(3) 基質溶解素 (Stromelysins)
包 含 stromelysin-1 (MMP-3) 、 stromelysin-2 (MMP-10) 、 stromelysin-3 (MMP-11) , 可 以 降 解 許 多 不 同 的 ECM 的 基 質 (Chambers et al;1997; Duffy et al; 1998),如proteoglycans (core protein)以及非膠原蛋白的laminin與fibronectin等。 (4) 細胞膜類基質金屬蛋白酶 (Membrane-type MMPs) 這 一 類 型 的 基 質 金 屬 蛋 白 酶 會 鍵 結 在 細 胞 膜 上 , 包 含 MT1-MMP (MMP-14) 、 MT2-MMP (MMP-15) 、 MT3-MMP (MMP-16)和MT4-MMP (MMP-17)等,能分解膠原蛋白,並且幫 助某些MMP的活化,例如:MT1-MMP被發現可活化MMP-2和 MMP-13 (Sato et al., 1994;Knauper et al., 1996)。
(5) Matrilysin
Matrilysin-1 (MMP-7) 、matrilysin-2 (MMP-26)。Matrilysin由 於缺乏haemopexin-like domain與一般MMPs應具有的區塊,所以 這些酵素能水解多種的基質,像是fibronectin、laminin、casein與 gelatin (Beaudeux et al., 2004)。
(6)其他 有 部 分 MMPs 未 被 歸 類 , 包 括 Metalloelastase (MMP-12) 、 MMP-18、MMP-19、MMP-20、MMP-27、MMP-28等。 所有的MMP都有專一的形式且含有不同的domain,MMP被合成 後是以非活化態的酶原 (zymogen)形式分泌,此時分子內蛋白的 cystiene和catalytic domain上的鋅結合,與其他的蛋白酶作用後, cystiene和鋅的鍵結被打斷後,使鋅完全外露,而成為活化態 (Nelson
et al; 2000)。活化的過程中會將MMP結構中N端的propeptide domain
表1-4. MMPs 的種類
第五節 基質金屬蛋白酶抑制劑(TIMPs)和轉移的相關性 基質金屬蛋白酶抑制劑 (TIMPs)能抑制 MMPs 的活性 (Beaudeux et al., 2004; 圖 1-7),研究指出 TIMPs 至少有兩種生理活性,除了具 有生長因子的功能,也能抑制 MMPs 活性 (Denhardt et al., 1993)。 TIMP 為內生性的 MMP 抑制劑,可以 1:1 與 MMP 成非共價鍵結複合 體,抑制 MMP 的活性,目前已發現有四種形式,如:TIMP-1 (28.5kD)、 TIMP-2 (21kD)、TIMP-3 (21kD)和 TIMP-4 (22kD)。其中 TIMP-1 最早 被發現且具有抑制 MMP-9 活性功能,而 TIMP-2 可以抑制 MMP-2 的活性表現 (Gomez et al., 1997),因此 TIMP-1 與 TIMP-2 在抑制腫 瘤細胞入侵與轉移上扮演重要的角色。TIMP-3 的功能與 TIMP-1、 TIMP-2 較不相同,與調控細胞分化和週期有關 (Stricklin et al., 1983),目前對 TIMP-4 的研究則較少,主要存於人類心臟組織中 (Greene et al., 1996)。
第六節 纖維蛋白溶解系統和轉移的相關性
纖 溶 酶 原 激 活 物 (plasminogen activator; PA) 是 一 群 serine proteinase,依其來源可分為尿激酶型纖溶酶原激活物 (urokinase-type plasminogen activator; uPA) 和 組 織 型 纖 溶 酶 原 激 活 劑 (tissue-type plasminogen activator; tPA),活化後會將纖溶酶原 (plasminogen)轉成 纖維蛋白溶解酵素 (plasmin)進行裂解纖維蛋白 (Collen and Lijnen., 1991)而抑制血栓的形成,降低心血管疾病的發生,另外也具有活化 MMPs 的功能 (Vassalli et al., 1991),分解 ECM 而增加癌細胞的遷移。
uPA 為 53kD 的 serine protein,不活化態時為單股的多胜肽 (Andreasen. et al., 1997; Dano et al., 1985),當 lysine-isoleucine 鍵 (K158-I159)被切斷時,則形成了活化態,uPA 由尿中所分離,主要來
自腎臟,頇與細胞表面接受器 (urokinase-type plasminogen activator receptor; uPAR)結合才具有較強的蛋白分解能力,研究指出抑制 MDA-MB-231 乳癌細胞中的 NF-kB 表現可以降低 uPA 分泌,以及癌 細胞的遷移情形 (Sliva et al., 2002)。 tPA 分子量約為 68kD,由 530 個氨基酸組成,基因位在人類第 八條染色體上,主要由內皮細胞分泌,與 uPA 相同皆以不活化態的 單股形式被分泌出去,在 Arg-275-Ile-276 位置被切斷時,形成雙股, 但是此時並不表示具有酵素活性 (Das et al., 2003),只有在和纖維蛋 白 結 合 後 tPA 才 具 有 活 性 。 uPA 與 tPA 會 受 到 內 生 性 抑 制 劑 (plasminogen activator inhibitor type 1、2; PAI-1、PAI-2)的影響,調節 血栓平衡與癌細胞轉移能力。
(Beaudeux et al., 2004) 圖 1-7. 蛋白分解酵素分解細胞外基質相關機制
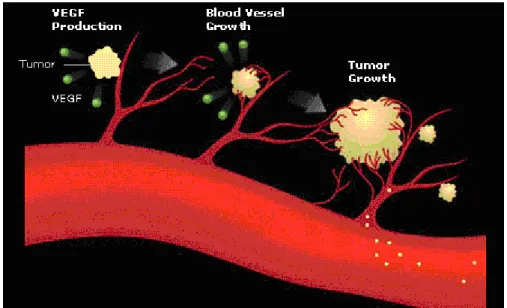
第七節 血管新生與轉移的相關性 原有的血管生成新的血管過程就稱為血管新生作用,在正常的生 理環境,包括胚胎發育、傷口癒合等等,都需要新的血管生成,而當 血管新生失去平衡時,如缺乏的情況會抑制傷口癒合以及冠狀動脈局 部缺血,反之,過度的血管新生與糖尿病視網膜病變息息相關,並且 促進腫瘤生長,腫瘤的血管新生過程是非常複雜的,由於腫瘤細胞會 不斷地釋放血管生長因子,例如:vascular endothelial growth factor (VEGF)、basic fibroblast growth factor (bFGF)等,使腫瘤週遭的血管 內皮細胞受到刺激而增加附近的血管新生,增加腫瘤養分的來源,促 進腫瘤生長(圖 1-8),此時除了癌細胞本身外,血管內皮細胞也會分 泌蛋白分解酵素 (MMP-1、MMP-2、MMP-3 與 MT-MMP),造成血 管基底膜與細胞外基質分解,癌細胞則進入血液循環,侵入其他組織 或器官,增加癌細胞的轉移能力 (Fidler et al., 1994; Folkman et al., 1995)。
許多研究顯示 MMP-2 與 MT-MMP 在血管新生上扮演重要的角色 (Nagase et al; 1999)。另外,增加 MMPs 的表現除了促進癌細胞的轉 移,也會影響 VEGF 的蛋白表現,文獻指出增加 VEGF 會促進人類 肝癌細胞的轉移能力 (Huang et al., 2005; Turlin et al., 2002)。
資料來源:
(http://www.gene.com/gene/research/focusareas/oncolo gy/angiogenesis.html)
圖 1-8. VEGF 促進血管新生與癌細胞轉移機制圖
第十節 Mitogen-Activated protein Kinases (MAPKs)與轉移的相關 性
活化絲裂原活化蛋白激酶家族 (MAPKs),是一種serine/threonine kinases的激酶家族,廣泛存在於各種細胞中的傳導途徑,主要參與細 胞 外 各 種刺 激物的 訊 息 傳遞 反應與 細 胞 凋亡 的信號 轉 導 過程 。 MAPKs 級聯反應包括3 個順序的活化過程:MAPK kinase kinase (MAPKKK)、 MAPK kinase (MAPKK)及MAP kinase。每一種激酶又 由不同成分組成,MAPKKK 由C—RAF,B—RAF,MEKK等組成, MAPKK 由MEK,SEK,MKK3,MKK6等組成,MAPK包括ERK 1/2 (extracellar signal-regluatedkinase) , JNK/SAPK (c-Jun NH2-terminal
kinase)以及p38。MAPK家族有多個成員及多條反應途徑,利用一連 串的蛋白質磷酸化反應來進行訊息的傳導。依照細胞種類、cellular
stmuli、細胞的環境等因素之不同會使各種MAPK在細胞的生理功能 上 產 生 不 同 影 響 , 包 括 發 炎 反 應 (inflammination) 、 細 胞 凋 亡 (apoptosis)、致癌基因轉形、腫瘤細胞轉移 (Chakraborti et al., 2003)。 過去在不同細胞研究顯示經由ERK1/2、P38與JNK路徑會調控MMPs 與uPA的表現 (Chen et al; 2005; Westermarck et al; 1999),因此抑制 MAPKs路徑可能與抑制血管新生、細胞增生與癌細胞轉移有關,以 下將介紹ERK1/2、P38與JNK路徑:
(1)Extracellular signal-regulated kinase (ERK1/2)
最早被發現的分子成員為 MAP-2 kinase (microtubule-associated proteins),在受到胰島素的刺激便會大量活化的蛋白質激酶,進而造 成細胞骨架蛋白質的磷酸化 (Sturgill et al., 1986)。而在之後的研究也 發現 MAP-2kinase 之異構酶 ERK1 (Boulton et al., 1991)。許多物質, 包含生長因子、細胞激素,甚至是致癌物質皆可以活化 ERK1/2 之途 徑。當 ERK1/2 的途徑被啟動時,能促進細胞的增殖,抑制細胞的凋 亡 (Johnson et al., 2002)。然而,在轉移機制方面,有文獻指出抑制 了 p-ERK1/2 可以減少 MMP-2 及 uPA 表現,且抑制腫瘤細胞移行 (Westermarck et al; 1999; Lev et al., 2004),Protein kinase C (PKC)已被 證實可以活化 ERK1/2 的途徑,而近年來發現當 PKCδ、PKCε、PKCδ 過度表現與活化 MMP-1 的 promoter 有關(Rutter et al., 1995)。 MDA-MB-231 乳癌細胞可以經由 TNF-α 誘導 MMP-9 與 AP-1 的表 現,AP-1 能被 ERK1/2 與 P38 調控,當加入 ERK 抑制劑 U0126 與 P38 抑制劑 SB203580 後,能降低 TNF-α 所誘導的 MMP-9 活性表現 (Kim et al., 2008)。
(2) p38 mitogen-activated protein kinase (p38 MAP kinase)
目前發現 p38 家族至少存在有四種異構酶: p38α (MAPK14、 SAPK2a)、p38β (MAPK11、SAPK2b)、p38γ (MAPK12、SAPK3) 及 p38δ (MAPK13、SAPK3)(Kyriakis et al., 2001)。p38 也被證實在細胞 凋亡與存活上扮演雙重角色。在乳癌細胞受 v-Src 所活化的 p38 會藉 由調控 cyclin D 的作用而增加細胞增生 (Zarubin et al., 2005);數種 cellular stress 如 anticancer agent、UV raddiation、oxidative stresses、 growth factor 和 ceramides 等皆透過 p38 而抑制細胞週期的進行或啟 動細胞凋亡 (Olson et al., 2004)。p38 調控轉移機制方面,BT549 人類 乳癌細胞中 p38α 與 p38β 異構物做比較,p38α 為主要調控 uPA 的表 現 (Huang et al., 2000)。另外,phorbol myristate acetate (PMA)為一種 化學致癌劑,能使神經瘤細胞 TIMP-2 表現降低,若抑制了 p38 的磷 酸化,則能增加 TIMP-2 的表現量,降低癌細胞的轉移能力 (Park et
al.,2002)。
(3) c-Jun N-terminal kinase/stress activated protein kinase (JNK/SAPK)
當細胞受到外界的壓力或是一些cytokine的刺激時,便會活化 JNK。JNK主要有3種isoform:JNK1、JNK2及JNK3,前兩者表現在 大部分的組織,而JNK3主要表現在腻部及睪丸 (Barr et al., 2001)。活 化之JNKs可將tumor suppressor gene p53磷酸化,因此抑制p53與 Mdm2的結合而延緩p53之代謝並延長p53啟動細胞凋亡或細胞週期 停滯之活性 (Liu et al., 2005; Cheng et al., 2003)。在JNK影響轉移部 分,抗氧化物質α-Mangostin可以經由抑制前列腺癌細胞 (PC-3)中 JNK磷酸化與降低轉錄因子 NF-kB與AP-1的表現而抑制MMP-2、 MMP-9與uPA的酵素活性 (Hung et al., 2009)。乳癌細胞中含有大量的
leptin,會增加癌細胞的轉移能力,研究顯示leptin刺激乳癌細胞 (MCF-7)的移行能力,並且增加MMP-2的表現,以及活化JNK,而當 抑制JNK磷酸化後,則能降低leptin所誘導的MMP-2表現 (McMurtry
et al., 2009),因此在MAPK路徑中,JNK在抑制癌細胞轉移上也占非
常重要的角色。
第九節 Nuclear factor kappa B (NF-kB)與轉移的相關性
NF-kB 為 細 胞 核 轉 錄 調 節 因 子 , 首 先 被 發 現 於 成 熟 的 B lymphocytes細胞核內,其與免疫球蛋白kappa light chain enhancer region 上 的 B 位 置 特 定 序 列 5’-GGGACTTTCC-3’ 結 合 (Sen and Baltimore., 1986),調控kappa light chain的轉錄,因而稱之細胞核轉錄 因子kB。研究指出NF-kB廣泛存在於各種真核細胞中,其含有五種次 單元能組合成不同的雙聚合體,包括Rel A (p65)、Rel B (p68)、c-Rel (p75)、p50/p105 (NF-kB1)與p52/p100 (NF-kB2),最早被分離出來的 NF-kB為p65/p50所構成的異源雙聚體 (heterodimer),而此種雙聚體 也是最常見的NF-kB。 NF-kB的活性受到IkB蛋白的調控 (圖 1-9),在細胞未受刺激 時,NF-kB與IkB結合形成三聚合體複合物並以無活性狀態存於細胞 質中,IkB家族有三種形式包括IkB-α、IkB-β與IkB-ε,研究指出當細 胞受到刺激時,IkB-α能快速被IkB kinases磷酸化 (Karin., 1999),IkB-α 經由泛素與蛋白酶體系統 (ubiqutination-proteasome system)而降解, 因而NF-kB (p65/p50)露出與DNA結合的位置,具有活性的NF-kB進入 核內則能與DNA上的kB基因序列結合進行調控各種基因的表現 (Patrick et al., 1996)。
白、免疫調節因子 (immune regulation molecule)、生長因子、細胞黏 附因子 (cell adhesion molecule)、細胞凋亡、腫瘤發展、血管新生與 細胞遷移等等 (Sliva et al., 2004; 圖 1-10)。
(Modified from Chakraborti et al., 2003; Lee et al., 2007)
圖 1-9. MAPK 與 IkB 影響 NF-kB 調控路徑
過去研究指出 LPS (lipopolysaccharide)誘發巨噬細胞 RAW 264.7 的 iNOS 與 COX-2 蛋白表現,經由給予 AC-10 後能阻擋 NF-kB 活性
Growth factors Oxidative stress
Environmental stress Cytokines Ras Raf P38 JNK/SAPK MKK5,6 MKK3,6 ERK5,6 MKKK1,2,3,4 ERK1,2 MKK1,2 MKK4,7 p50 p65 Nucleus IkB degradation p50 p65 IkB p50 p65 P TGF-β NF-kB responsive gene P P P P P Cell membrane
表現而被抑制,降低發炎的情形 (Hseu et al., 2005)。在抑制癌細胞轉 移能力部份,許多癌細胞中 NF-kB 的表現量高,包括乳癌 (Nakshatri
et al., 1997)、前列腺癌 (Mukhopadhyay et al., 2001)、頭頸部癌
(Ondrey et al., 1999)與胰癌細胞等等。另外,已證實 NF-kB 能調控 MMP-9 與 uPA 的基因表現,具有影響癌細胞黏附、遷移與移行的能 力 (Lengyel et al., 1996; Banerjee et al., 2002)。
MAPK 也能調控 NF-kB 的活性,P38 與 ERK 的活化可以調控依 賴 NF-kB 的相關基因表現 (Carter et al., 1999),研究顯示一種具抗氧 化物質的 capillarisin 能抑制致癌劑 PMA 所誘導 MCF-7 乳癌細胞中 MMP-9 的表現,並且是透過 P38 與 JNK1/2 調控 NF-KB 轉錄因子而 降低轉移相關因子表現 (Lee et al., 2008),因此 MAPK 與所調控的 NF-kB 路徑被活化後,可能進一步向下調控轉移相關蛋白的表現,影 響癌細胞的轉移能力。
第十節 研究動機 根據 97 年衛生署資料統計,乳癌在女性癌症死亡排行仍為第四 位,造成乳癌患者死亡的主要原因多為癌細胞的轉移,引起其他組織 或器官的病症,因此在預防癌症發生的同時,學者也很迫切想找出能 抑制罹癌患者腫瘤細胞轉移的方法。而在本實驗室之前已證實樟芝發 酵液 AC-10 能促進乳癌細胞 (MCF-7 與 MDA-MB-231)的細胞凋亡 (Hseu et al., 2006; Hseu et al., 2007),而對於抑制乳癌細胞轉移的機制 仍未知,因此本實驗將探討 AC-10 是否也具有抑制 MDA-MB-231 乳 癌細胞轉移的能力以及可能調控的訊息傳遞路徑。
第二章
樟芝發酵液(AC-10)
細胞存活率、 遷移與移行
MTT assay
Wound migration assay Transwell invasion assay
乳癌細胞株 (MDA-MB-231) NF-kB活 性表現 轉移相關蛋白 與活性表現 Western Blotting MMP-2 MMP-9 uPA uPAR TIMP-1 TIMP-2 PAI-1 VEGF MAPK 蛋白表現 Gelatin、Casein zymography MMP-2 MMP-9 uPA NF-kB reporter assay Western blotting ERK P38 JNK MAPK 抑制劑 Western blotting Gelatin zymography MMP-9
第三章
一、 實驗材料
1. 購自 ATCC (American Type Culture Collection)
人類乳癌細胞株(MDA-MB-231)
2. 購置食品工業研所 樟芝發酵液(AC-10) 3. 購自 BIO-RAD
Protein assay dye reagent 、 30% Acrylamide/Bis solution (29:1) 、 Recombinant protein molecular weight marker (All blue)、Ammonium persulfate、N,N,N,N-Tetramethyl ethylenediamide (TEMED)、Sodium dodecyl sulfate (SDS)、Tris-HCl、Glycine
4. 購自 GIBCO
Penicillin- Streptommycine (PS)、L-Glutamine
5. 購自 Hyclone
Trypsin-EDTA
6. 購自 Merck
Methanol
7. 購自 Sigma
Thiazolyl blue tetrazolium bromide (MTT)、Sodium chloride (NaCl)、 Potassium chloride (KCl)、Glucose、Glycerol、Xylene、Triton-X100、 Sodium Bicarbonate (NaHCO3) 、 Sodium phosphate (NaHPO4) 、
Potassium phosphate monobasic (KH2PO4) 、 Tris-base 、
Ethylenediamide-tetraacetic acid (EDTA) 、 Trypan blue 、 Pyruvate sodium、Ascorbic acid 、bovine serum albumin (BSA)、Tris-HCl、β-actin mouse monoclonal antibody、Casein sodium salt from bovine milk、 (N-[2-Hydroxyethyl]piperazine-N’-[2-ethanesulfonic acid]) (HEPES)、 U0126、SB203580、SP600125
8. 購自美國 Santa Cruz
一級抗體:
Phospho-ERK mouse monoclonal antibody 、 ERK rabbit polyclonal antibody、MMP-2 goat polyclonal antibody、MMP-9 goat polyclonal antibody、TIMP-1 rabbit polyclonal antibody、TIMP-2 rabbit polyclonal antibody 、 PAI-1 rabbit polyclonal antibody 、 uPA rabbit polyclonal antibody、uPAR rabbit polyclonal antibody、VEGF mouse monoclonal antibody
二級抗體:
Anti-mouse IgG、Anti-rabbit IgG、Anti- goat IgG
9. 購自 Cell Signaling
一級抗體:
Phospho-SAPK/JNK rabbit polyclonal antibody、P38 MAP Kinase rabbit polyclonal antibody 、 SAPK/JNK rabbit polyclonal antibody 、 Phospho-P38 MAP Kinase rabbit polyclonal antibody
10. 購自 BD BioCoat
BD MatrigelTM Invasion Chamber 24-Well Plate 8.0 Micron
11. 購自安佳
Non-fat Milk
12. 購自Thermo Scentific
Supersignal® West Pico Chemiluminescent Substrate 、 Dulbecco’s Modified Eagle Media (DMEM)
13. 購自Biological Industries
胎牛血清 (Fetal bovine serum;FBS)
二、儀器
1. 天秤:Sartorius
2. 直之式電泳槽:Hoefer 3. 電泳轉印槽:BIO-RAD
4. 電源供應器:BIO-RAD computer power supply Model 3000 Xi 5. 迷你離心機:KUBOTA 2010
6. 分光光度計:Spectrophotometer U-2000 (Hitachi) 7. 細胞培養箱:NUAIRETM US AUTO flow
8. 幫浦:HETO SUE 30Q
9. 無菌操作台:NUAIRETM class Ⅱ TYPE A/B3 10. 倒立式顯微鏡:Nikon Diaphot 300 11. 超音波震盪器:Bransonic PC 620 12. 高壓殺菌釜:TOMIN TM32 13. 數位影像分析儀:Alphar Imager 2000 14. 水浴槽:FIRSTERTM SIENTIFIC 15. 水平式搖晃器:Oribital shaker OS 701 16. 感光夾:HypercassetteTM rpn111649 17. 純水製造機: Millipore milli-Q Plus 18. 震盪器:KS ORBITAL Shaker
19. X 光感光厎片:Kodak Scientfic Imaging Film 20. ELISA Reader:Dynatech MR7000&Dynex MRX 21. ultracentrifuge:Beckman
22. pH meter:Denver Basic
第四章
一、 樟芝發酵液(AC-10)
本實驗採用的樟芝發酵液是以 0.22 Μ 孔徑之濾膜 (filter)過濾後, 分裝於 1.5ml 微量離心管,貯存於-80 ℃。
二、 細胞培養 1. 培養液配製
(1) Dulbecco’s Modified Eagle Media ( DMEM )
取一包 DMEM 粉末溶於 900 ml 的二次水中,加入 3.7 g NaHCO3、
5.9 g HEPES、 110 mg Sodium pyruvate。溶解後調 pH 值至 7.2~7.4, 並將體積定量至 1000 ml。以 0.22μm 血清瓶專用無菌過濾膜 (bottle top filter) 過 濾 , 再 加 入 1% 青 黴 素 (penicillin)/ 鏈 黴 菌 ( streptomycin ),混和後保存於 4℃。 (2) 1X 磷酸緩衝液 ( phosphate-buffer saline;PBS ) 8 g NaCl、0.2 g KH2PO4、0.2 g KCl、2.9 g NaHPO4・12H2O 溶在 900 ml 二次水中,以 1 N NaOH 調整到 pH 值至 7.2~7.4,並將體積定量 至 1000 ml,再經由 121℃、30 分鐘高壓滅菌後使用並存放於室溫備 用。 2. 細胞培養條件 將人類乳癌細胞株 (MDA-MB-231)培養於 DMEM 培養液中,並含 10% 加 熱 去 活 化 之 胎 牛 血 清 (heat-inactivated foetal bovineserum ; FBS),置於 37℃、5% CO2 培養箱中培養。
細胞培養在 T75 培養皿 ( flask )中生長至八、九分滿時,將 DMEM 去除,PBS 清洗 2 次,再用 Trypsin-EDTA 將細胞分離自 T75 培養皿, 再加入新鮮之 DMEM +10 % FBS 並將細胞液收集至 50ml 離心管中, 在室溫下 1000 rpm 離心 5 分鐘,去除上清液,加入 DMEM+10 % FBS 混 勻細胞。取 100μl 細胞懸浮液與等量之 0.4% Trypan blue 溶液混合均勻, 用細胞計數器 (hemocytometer )計數細胞數,依實驗需要之細胞密度均 勻的分至培養盤 ( culture plates ) 。 三、 細胞存活率(viability)之測定 1. 原理 MTT為水溶性的tetrazolium salt,以PBS溶解後會形成淡黃色,在經 由細胞內粒線體的脫氫酶還原,而MTT的環狀結構tetrazolium ring會被 切斷而轉換成不溶水的藍紫色formazan結晶,再利用10% SDS溶解細胞 膜與藍紫色結晶物,在波長570 nm測得吸光值。此反應頇要在活細胞中 進行,因此當藍紫色愈多表示細胞存活愈多。 2. 試劑 (1) 0.5mg/ml MTT溶液: 在避光環境中,取0.5mg MTT粉末,加入1ml PBS為溶劑,溶解後再以0.22M濾膜過濾使用。 (2) 10% SDS : 取10 g SDS粉末,以二次水定量至100ml,使用前需以 0.22μm filter過濾。 3. 方法
將乳癌細胞 MDA-MB-231(2.5×105 cells/well)種於 24well,隔天細
胞貼壁後,以 PBS 清洗,加入含 1%胎牛血清 (FBS)的 DMEM 培養液, 讓細胞同步化後,隔天加入不同濃度的 AC-10,置於 37℃、5% CO2培
時後再加入 400μl 的 10 % SDS,反應 12 小時,抽出 200μl 的上清液於 96 孔微量盤中,以 570nm 波長測吸光值 (Parada-Turska et al., 2007)。 四、細胞遷移之分析 (Wound migration assay)
1. 原理 細胞間會受到彼此分泌生長因子而互相吸引與生長,當細胞間受到 損傷時,即細胞間支持與提供訊號傳遞的 ECM 受到破壞時,細胞會啟 動修復機制,在癌細胞轉移部份,若細胞為高轉移細胞株,當細胞間受 到破壞,加上細胞本身釋放大量的蛋白分解酵素,促進細胞外基質降 解,因此增加癌細胞的遷移情形,造成癌細胞的轉移。 2. 試劑 (1) 75% 乙醇。 (2) Giemsa Stain: 將染劑加入二次水以 1:20 比例稀釋。 3.方法
將 MDA-MB-231 cell (3×105)種於 12 well,隔天以 PBS 清洗 2 次,換 成含 1% FBS 細胞培養液,隔天以 200 l Tip 在 well 底部刮一條空間後, 移除細胞培養液並以 PBS 洗 2 次,加入含 1% FBS 的細胞培養液與不同 濃度的 AC-10,在 0 小時與 24 小時拍照觀察細胞遷移的情形,反應結 束後,以 75% 冰酒精固定 30 分鐘,待酒精完全揮發後,以 Giemsa Stain 染色,於顯微鏡底下 (200X)隨機選三個視野計數遷移的細胞量 (Lin et al., 2008)。
五、細胞移行試驗 (Transwell invasion assay) 1. 原理
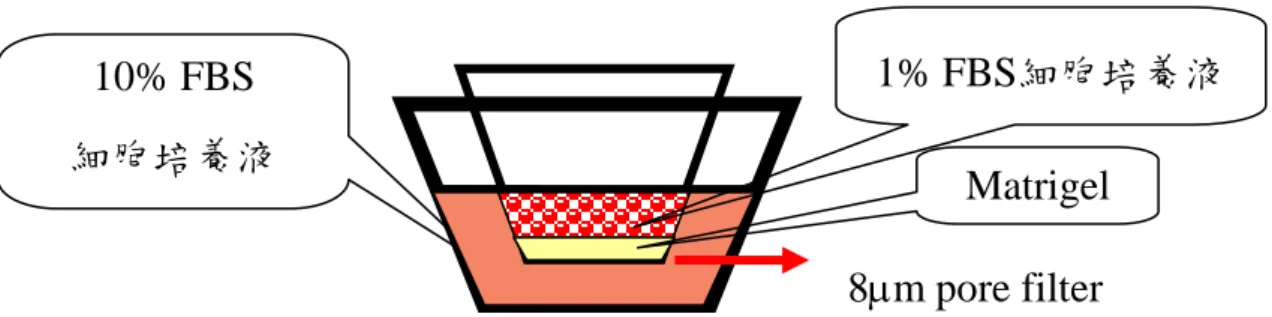
Matrigel 為細胞外基質的成分,可以模擬細胞間環境,此實驗利用 BD MatrigelTM Invasion Chamber , 含 有 Matrigel matrix , 其 來 源 為
Engelbreth-Holm-Swarm 小鼠腫瘤,分離出來的 Matrigel matrix 含有 laminin、collagen type IV、heparan sulfate proteoglycan 以及生長因子 TGF-beta 與 basic-FGF。因此當細胞受到一些生長因子刺激,並且細胞 分泌一些基質分解酵素,將 Matrigel 分解,會吸引細胞的移行,若細胞 移性能力強,染色後可以在濾膜上觀察到細胞。 2. 試劑 (1) 75%乙醇。 (2) Giemsa Stain: 將染劑加入二次水以 1:4 比例稀釋。 3. 方法 下層盤 (24well)中加入 750 l 含有 10% FBS 的 DMEM 當趨化劑, 將下層盤或稱轉移盤 (transwell)放入下層盤,並放入培養箱 2 小時,讓 matrigel 復水 (圖 4-1)。再將 500 l 的 MDA-MB-231 乳癌細胞液(1×105 cells)種入上層盤,含 1% FBS 的 DMEM 培養液以及不同濃度的 AC-10; 並將轉移盤放入下層盤後,移入 37℃、5% CO2的培養箱培養 24 小時, 之後將轉移盤拿出,使轉移盤倒扣在紙巾上,並用棉花棒將轉移盤內的 細胞、培養基移除,以 750 l 的冰乙醇 (75%)固定 15 分鐘,於室溫下 自然乾燥。以 Giemsa Stain 染色後,再用清水洗去多餘的染劑。以光學 顯微鏡在 200Ⅹ視野下觀察,隨機選取位於轉移盤膜下表面中的 9 個視 野,並計算每一視野中的總細胞數;9 個視野細胞數的平均值為每個轉 移盤之乳癌細胞趨化性移動的指標,每次實驗中之各組皆以 3 個重覆之 轉移盤的平均值為代表(Rose et al., 2005)。
圖 4-1. 轉移盤(Transwell) 六、 MMPs 活性分析 (Gelatin zymography) 1. 原理 MMP-2 和 MMP-9 都 是 屬 於 gelatinases , 所 以 可 以 利 用 gelatin zymography來分析MMP-2和MMP-9的酵素活性,當細胞上清液中含有 MMP-2 和 MMP-9 時 , 會 將 膠 體 中 的 gelatin 分 解 , 而 在 分 子 量 72kD (MMP-2)和92kD (MMP-9)位置呈現透明的條紋。 2. 試劑
(1) 6× protein dye:1.5M Tris HCl (pH 6.8)、 30% SDS、60% glycerol、
0.3% bromophenol blue。
(2) 10× Running buffer:取2.9 g Tris base、144 g Glycine、10 g SDS、
以二次水定量到1000ml。
(3) 10×Renaturing buffer:25% TritonX-100 (以二次水配製)。
(4) 10× Developing buffer:取12.1 g Tris base、60 g HCl、117 g NaCl、
7.4 g CaCl2、Brij 0.2%,以二次水定量到1000ml。
(5) 0.5% Coomassie Blue:0.5% Coomassie Blue R-250、50% methanol、
10% acetic acid。 3. 方法
(1) 將乳癌細胞 MDA-MB-231(3×105 cells/well)種於 12well,隔天細 8m pore filter
10% FBS 細胞培養液
Matrigel
胞貼壁後,以 PBS 清洗 2 次,加入含 1% 胎牛血清 (FBS)的 DMEM 培養液,培養至隔天。加入不同濃度的 AC-10,與 1% FBS 的 DMEM 培養液,培養 24 小時後,將 cell culture medium 抽起來放進微量離心 管中,離心 1500 rpm,10 min,取上清液並保存在-80℃。
(2) 取 40l 含有 20 g 蛋白的上清液混合物,加入 6× protein dye,充 分混合,靜置 10 分鐘。將電泳膠片置於電泳槽上,並加入 Running buffer,以 8% SDS-PAGE (含 gelatin 1mg/ml)、50 ~60V 進行電泳,將 48l 樣本混合物及 5 l 的蛋白質標記 (protein marker)注入到各個電 泳膠片槽溝中。直到蛋白質標記 (protein marker)的 50 kD 移至膠底介 面,或是 92 kD 約在膠的中間時即停止電泳。以 Renaturing buffer (2.5% Triton X-100 )和 gel 室溫反應 30 分鐘,以去除 SDS,然後倒掉 Renaturing buffer(1X)並且加入 Developing buffer (1X)室溫下先反應 30 分鐘後,再加入新的 Developing buffer (1X),在 37℃、35rpm 下 作用 20~24 小時。以 0.5% Coomassie Blue 在室溫下,置於 shaker 上 染色約半小時,將 gel 以二次水浸泡至隔天,然後將膠置入數位影像 分析儀 (Alphar Imager 2000)受測物板上照相 (Huang et al., 2004)。 七、 uPA 活性分析 (Casein zymography)
1. 原理 uPA會將Plasminogen分解成plasmin,利用casein zymography (含有 plasminogen)來分析uPA的酵素活性,當細胞上清液中含有uPA時,會將 膠體中的plasminogen分解,而在分子量45kD (uPA)位置呈現透明的條 紋。 2. 試劑
0.3% bromophenol blue。
(2) 10× Running buffer:取2.9 g Tris base、144 g Glycine、10 g SDS、
以二次水定量到1000ml。
(3) 10×Renaturing buffer:25% TritonX-100 (以二次水配製)。
(4) 10× Developing buffer:取12.1 g Tris base、60 g HCl、117 g NaCl、
7.4 g CaCl2、Brij 0.2%,以二次水定量到1000ml。
(5) 0.5% Coomassie Blue : 0.5% Coomassie Blue R-250 、 50%
methanol、10% acetic acid。 3.方法
上清液收集方式gelatin zymography相同,接著取40 l含有20 g蛋白 的 上清液混 合物, 加入 6 x dye充 分 混合,靜 置 10分 鐘後,以 10% SDS-PAGE (含plasminogen 15 g/ml和Casein 1 mg/ml)、50V進行電泳, 當蛋白質標記 (protein marker)45kD約到膠的中間時即停止電泳。之後將 gel放在renaturing buffer (1X)中並以35rpm於室溫作用30分鐘,以去除 SDS,再用清水清洗二次後,以developing buffer (1X)於室溫下先作用30 分鐘後,再換成新的developing buffer (1X),於37℃環境下繼續作用20~24 小時後移除,以清水洗一次,並利用coomassie blue R-250染色30分鐘, 再以二次水浸泡至隔天,然後將膠置入數位影像分析儀 (Alphar Imager 2000) 受 測 物 板 上 照 相 分 析 。 若 樣 本 含 有 urokinase type plasminogen activator (uPA),則在膠體分子量45 kD位置會因plasminogen被酵素分解 而呈現的透明band (Chu et al., 2004)。
八、 西方墨點(Western blotting)分析法 1. 原理
白分開後,為了偵測特定的蛋白質含量時,可利用抗體和抗原專一性結 合原理,將特定的蛋白質找出,分析其含量的相對多寡。先加入能和特 定蛋白質結合的專一性抗體(一級抗體, primary antibody),充分反應 後,再加入能和一級抗體結合的另一專一性抗體(二級抗體, secondary antibody),兩抗體結合後,二級抗體上結合的酵素,可以分解特定之基 質(substrate)而產生冷光,經底片感光後,可用於評估特定蛋白質之 表現。 2. 試劑
(1) Lysis Buffer : 10 mM Tris HCl(pH=8)、0.32 M sucrose、5 mM
EDTA、1mM PMSF、2 mM DTT、1% Triton X-100。
(2) 1.5 M Tris (pH=8.8):取 91g Tris base 溶在 300 ml 二次水,以 1N
HCl 調至 pH=8.8,用二次水量定量至 500ml 以 0.45μm filter 過濾, 4℃儲存 1 個月。
(3) 1 M Tris (pH=6.8):取 12.1g Tris base 溶在 40 ml 二次水,以 1N
HCl 調至 pH=6.8,最後用二次水定量到 100ml,用 0.45μm filter 過 濾,4℃儲存 1 個月。
(4) 30% Acrylamide/Bis solution (29:1)
(5) 10% SDS:取 10g SDS,最後用二次水定量到 100 ml。
(6) Ammonium persulfate:取 0.1g (NH4)S2O8 溶於 1 ml 二次水。
(7) 10% running gel: 8 ml H2O、5 ml 1.5M Tris (pH=8.8)、6.6 ml30%
Acrylamide Mix (acrylamide:bisacrylamide 29:1)、0.2 ml 10% SDS、 0.2 mL 10% ammonium persulfate 及 0.01 mL TEMED。
(8) 12% running gel: 6.6 ml H2O、5 mL 1.5M Tris (pH=8.8) 、8.0 ml
30% Acrylamide Mix (acrylamide:bisacrylamide 29:1)、0.2 ml 10% SDS、0.2 ml10% ammonium persulfate 及 0.01 ml TEMED。
(9) 5% Stacking gel: 3.5 ml H2O、0.625 ml 1.5 M Tris (pH=6.8)、0.825
ml 30% Acrylamide Mix、0.05 ml 10% SDS、0.05 ml、10% ammonium persulfate 及 0.05 ml TEMED。
(10) 6X Protein loading dye: 350 mM Tris-HCl (pH=6.8)、12%
SDS 、35% glycerol、0.02% bromopherol blue、30% β-meanptoethanol
(11) 5X electrode buffer (pH=8.4): 取 54.5 g Tris base、24.8 g Boric
acid、4.7 g EDTA2Na、5g SDS,用二次水定量至 1000ml。
(12) Transfer buffer: 取 18.2 g Tris base、86.5 g Glycine、1200 ml
methol,用二次水定量至 3 升 。
(13) 1X PBS: 取水 800 ml 二次水,加入 8 g NaCl、0.2 g KCl、0.2 g
KH2PO4、2.9 g Na2HPO4.12H2O,調 pH 7.4,並定量 1000 ml。 (14) Washer buffer: 500 ml PBS 再加入 500μl Tween-20。
(15) Block buffer: 取 5% 脫脂奶粉溶於 PBS。
(16) 顯影劑: 取 103 ml developer 加入 370 ml 二次水,稀釋成 4.59
倍,放入避光的瓶子。
(17) 定影劑: 取 103 ml fixter 加入 370 ml 二次水,稀釋成 4.59 倍,
放入避光的瓶子。
(18) SuperSignal substrate solution: 將 SuperSignal I:SuperSignal II
=1:1 混合,加入的量依 PVDF 膜大小調整。 3. 方法
(1) 細胞蛋白質萃取 ( cytosolic protein extraction )
將細胞分盤培養在含有10% FBS的DMEM之6 cm dish中,每個 dish細胞數為1× 106 個細胞。細胞貼壁後,更換成含1% FBS的細胞 培養液,待隔天後,加入不同濃度樟芝 (0-80μg/ml),在培養箱反應 24小時,取出培養盤用刮刀將細胞刮下,收集於50ml離心管中,以
1500 rpm、4℃離心5分鐘,去除上清液將細胞移置1.5ml eppendorf tube,加入1000 μl PBS將細胞pellet完全打散後以1500 rpm、4℃ 離 心10分鐘,去除上清液加入適量lysis buffer將細胞pellet均勻打散, 靜置於冰上反應30分鐘,將溶解後之細胞液以12,000 rpm、4 ℃離心 30分鐘,上清液即為細胞蛋白萃取液 (Hseu et al., 2008)。
(2) 蛋白質濃度定量分析 (Protein concentration assay)
配置不同濃度 (0、2、4、6、8、10 μg /ml)之牛血清白蛋白 (Bovine serum albumin; BSA) 標準品,並將欲測定之蛋白質溶液稀釋成適當 的濃度。取800 μl之二次水加入不同濃度牛血清白蛋白標準品及蛋白 質溶液,加入200 μl Protein assay dye,充分混合後靜置5分鐘後測波 長595 nm之吸光值,並利用牛血清白蛋白標準品所求出的標準曲線 計算出欲測定之蛋白質溶液之濃度。
(3) 聚丙烯醯胺膠體電泳法 (Sodium dodecyl sulfate polyacrylamide gel electrophoresis,SDS-PAGE)
10%或 12% running gel,注入電泳玻片間,待凝膠完全後,加入 5% stacking gel 位於 running gel 之上並插入電泳梳,於室溫下聚合 反應 30 分鐘,完全聚合後去除電泳梳,裝置在電泳槽上,並浸泡於 1X running buffer 中 (stacking gel 之 槽 溝 要 補 滿 1X running buffer) 。取 50 μg~70 μg 之蛋白質,加入相同體積的 6×Protein loading buffer (為樣品 1/6 倍的體積),以 PBS buffer 調整使每一個樣 品之體積相同,並以 97℃加熱 6 分鐘 (使蛋白質變性),立即放回冰 上靜置 5 分鐘,再將蛋白質樣品及 marker 注入各個電泳膠片槽溝中 進行電泳分析,先以 80V 電壓跑 5% Stacking gel,30 分鐘,再改變 電壓為 100V,待 marker 完全展開後即停止電泳,再進行蛋白質的 轉印。
(4) 蛋白質轉印 (transfer protein to PVDF membrane)
取適當大小之 PVDF (polyvinylidene difluoride memberane ),以 甲醇浸潤,連同二張 3M paper 放入 transfer buffer 中浸潤。操作時重 疊順序由下而上依序為海綿、3M paper、gel、PVDF、3M paper、海 棉 (圖 4-2),並使用玻棒去除氣泡,放置於濕式 transfer machine 中, 以 20 V 的電流轉印 14~16 小時,將蛋白質轉印到 PVDF 上。轉印後, 將 gel 取出觀察轉印之效果;PVDF 則進行西方墨點法。 圖 4-2. 蛋白質轉印 (5) 免疫點墨法 (Immunoblotting) 轉印好蛋白質之 PVDF 膜用 Blocking solution 振盪 30 分鐘,以 Blocking solution 作為溶劑分別將欲測之一次抗體稀釋至適的濃 度,加入 PVDF 膜使其均勻的覆蓋於膜上,並置於水平式旋轉器上 室溫以 35rpm 反應 2 小時後,置入 4℃冰箱到隔天,先取出回復室 溫後,用 PBST 洗三次,每次 5 分鐘轉速為 100 rpm。依不同的一次 抗體,加入特異作用的二次抗體,稀釋倍數為 5000 倍,置於水平式 旋轉器上室溫振盪 2 小時,用 PBST 洗三次,每次 5 分鐘轉速為 100 rpm,以清洗未接合二次抗體,同時也可以降低背景值。 PVDF膜 海綿 3M紙 膠 電極+ 電極- 3M紙 海綿
將 PVDF 膜 加 入 SuperSignal® WestPico Chemiluminesent Substrate 溶液,振盪 3 分鐘,將 PVDF 膜用透明夾鍊袋完整包裹, 使用 Kodak BioMax light film 壓片 (依訊息的強弱調整壓片時間的 長短),將底片依序放入顯影劑、水、定影劑,最後再至入水中清洗 並停止顯色。以此方法檢視細胞內各個預測蛋白的變化量。使用冷 光影像儀器部分,則將 PVDF 膜,以塑膠夾子置入黑色金屬盤上, 蛋白面朝上,以 pippet 加入以 1:1 混合的 chemiluminesent Substrate 於膜上,在依螢光強弱決定曝光時間。
九、抑制劑之處理
細胞(1× 106 cells)種於 6 cm dish 中,隔天細胞貼壁後,以 PBS 清洗,加入含 1%胎牛血清 (FBS)的 DMEM 培養液,待細胞同步化後, 隔天先以 vehicle control (0.1% DMSO) 、ERK 抑制劑 U0126 (10、 30μM)、p38 抑制劑 SB203580 (10 、30μM)及 JNK 抑制劑 SP600125 (10、 30μM)培養 1 小時後,低劑量抑制劑結合 AC-10 的組別,則加入 20 g/ml 的 AC-10 培養 24 小時 (Shih et al., 2009)。
十、NF-kB 活性測定 (NF-kB reporter assay) 1.原理:
將有NF-kB啟動子 (promoter)及SEAP (secreted alkaline phosphatase) 作為報告基因的質體 (NF-kB-SEAP)送入細胞中,將質體送入細胞株的 過程稱為轉染作用 (transfection),本實驗以脂質體 (lipofectamine)方式 進行轉染,利用Lipofectamine Transfection Reagent (Invitrogen, U.S.A.)。 當NF-kB有表現時,NF-kB-SEAP被大量分泌到細胞培養液中,藉由抽取 一部份的上清液來偵測SEAP含量多寡,表示NF-kB的活性。