台北醫學大學 保健營養學系碩士論文
Master Thesis
School of Nutrition and Health Sciences Taipei Medical University
兒茶素和菊苣糖對於非酒精性脂肪變性肝炎大鼠的影響 Effects of Green Tea-Derived Catechins and Inulin
on Non-alcoholic steatohepatitis Rats
研究生 : 蘇倩瑢 撰 (Chien-Jung Su) 指導教授 : 陳俊榮 博士 (Jiun-Rong Chen, Ph.D.) 中華民國九十六年七月 July 2007
摘要
本研究目的是探討兒茶素與菊苣糖對於非酒精性脂肪變性肝炎 (Non-alcoholic steatohepatitis, NASH) 的影響。餵食雄性 Wistar 大白鼠 十二週高油高膽固醇飲食,誘發 NASH 後,隨機分為四組: N 組 (NASH 誘發飲食)、 C 組 (NASH 誘發飲食 + 兒茶素) 、 F 組 (NASH 誘發飲食 + 菊苣糖)、 FC 組 (NASH 誘發飲食 + 兒茶素 + 菊苣糖)。結果發現:飲食中額外添加兒茶素時,可降低血漿三酸甘油 酯,並且可以改善肝臟膽固醇和三酸甘油酯堆積的情形;且可藉由減 少肝臟中脂質過氧化物的堆積,進而降低肝臟抗氧化酵素的活性。此 外,也可降低 CYP2E1 蛋白質的表現量及 TNF-α 濃度。飲食中菊苣 糖的添加, 也可降低血漿三酸甘油酯,以及藉由減少游離脂肪酸濃 度,降低肝臟脂質過氧化物進而降低肝臟抗氧化酵素的活性,並且降 低 CYP2E1 蛋白質的表現量及 TNF-α 濃度。由研究結果推論,飲食 中兒茶素與菊苣糖的添加,雖然不具有加成作用,可能會藉由其特殊 的生理活性:調整血脂濃度、減少肝臟脂質堆積、降低體內高濃度游 離脂肪酸、降低體內氧化壓力、並減輕肝臟發炎反應,延緩肝臟細胞 走向纖維化,進而延緩 NASH 疾病進程 。 關鍵詞:非酒精性脂肪變性肝炎、兒茶素、菊苣糖、氧化壓力
Abstract
The aim of this study was to investigate the effects of catechin and inulin on hepatic steatosis and oxidative stress in non-alcoholic steatohepatitis (NASH). Sixty male Wistar rats were fed a high-fat diet for 12 weeks to induce NASH and then were divided into four groups: the N group (NASH diet), C group (NASH diet + catechin ), F group (NASH diet + inulin), and FC group (NASH diet + catechin + inulin). After the 6-week experimental period, the results showed that catechin and inulin significantly improved dyslipidemia induced by NASH diet and it lowered plasma triglyceride concentrations. Additionally, catechin intake also reduced the hepatic lipid depots of triglycerides and cholesterol, and decreased the concentrations of lipid peroxides. In an analysis of the antioxidant status, rats fed the catechin and inulin diet showed improved antioxidant potential due to decrease liver TBARS and a decrease in the protein expression of CYP2E1. In conclusion, catechin and inulin may improve the liver function in patients with NASH by lowering lipid levels in the blood and liver, increasing the antioxidant capacity.
致謝 在北醫的兩年, 經歷了很多事情, 難忘的事情很多 ,學到的事情也 很多。兩年的碩士生活,並不是一個人就可以完成的,是靠很多人的幫 忙,才可以把事情完成,因此要感謝的人有很多。 首先最要感謝的人就是又高又帥的指導老師 - 陳俊榮,老師提供了最 充足的資源,讓我可以沒有後顧之憂,完成實驗,並且給予寶貴的意見, 協助我完成碩士論文,不僅如此,老師也常常帶大家去吃 <好料的>, 老師對我的大方讓同學都羨慕萬分,在實驗室的生活可以說是<吃好的 用好的>,從進來的那一刻開始,就一直讓老師照顧 ,感謝老師的耐心 與關心,老師的好,是說也說不盡的,心裡面對於老師只有感激 – < 老師謝謝你>。 在我完成碩士的論文過程中,是靠許多人的幫忙才得以完成,首先最要 感謝的是欣怡學姐,不管是實驗還是生活上的問題,只要有困難,學姐 絕對不會吝於伸出援手,總是陪著我慢慢的把問題解決,學姐在我心中 是無所不能,所有問題只要碰到學姐就會迎刃而解,在這兩年裡給學姊 添了無數的麻煩,對於學姊除了感謝還有抱歉;也要感謝淑子姐,在每 一次報告前,總是不厭其煩給予建議,要跟學姐說一聲謝謝,讓我可以 在台上報告時,減少錯誤的發生;另外還有惠婷學姐,妳是一個很棒的 學姐,謝謝妳平常的照顧與關心,並且總是適時的給予鼓勵與打氣,真
的要感謝妳的陪伴,有妳真好;另外還有湘琦學姊,平時常常煩妳、吵 妳,可是有問題找妳時,學姐還是會伸出援手,解救我;再來要感謝的 人是 – 最棒的雅惠學姊,妳是我的救命恩人,是我生命的泉源,是我 的陽光,沒有妳我會枯萎,只有死亡。不論是抽血、犧牲以及分析,因 為妳的幫助才可以順利並且迅速完成,還要特別感謝妳就算畢業了,妳 還是常回來<照顧>我,二話不說幫我解決問題,真的很感激妳,我愛妳; 還有波姐,剛開始進入實驗室,要不是沒有妳一步步的教我做分析,我 根本沒辦法完成我的實驗,感謝妳花時間,慢慢的教我並且犧牲假日教 我分析,甚至幫我做實驗做到三更半夜,妳的大恩大德我會一直放在心 裡;我最正學妹 – 詩涵,對於妳,我只有滿滿的感激,要不是妳照顧 我,我可能沒辦法好好的完成實驗,不管是實驗或是生活,妳把我照顧 的無微不至,還要常常哄我,陪我哭陪我笑<我快哭了>,沒有妳我都不 知道該怎麼辦,我只能說認識妳的人都是幸福的,所以妳知道嗎?因為 妳 – 我是幸福的,我愛妳;Shu <昕嬬>,人雖然身在遠方,但還是掛 念著我,謝謝妳的鼓勵,妳還沒去美國就忙著幫我做實驗,到美國之後, 還要擔心我,妳真是辛苦了;最後還要謝謝我最可愛的學妹 – 宜靜、 筱珊、麗雯、宛儒、于瑩、玟暄,謝謝妳們的大力支持,妳們真是辛苦 了,除了幫忙實驗之外還要逗我開心,看我發瘋,我要謝謝妳們。 當然還有我的同學們 – 沂錚、柏方、宇葳、明樺、紓慧、佳妤、苯丸、 俐岑、萾芬、芷琪、歆倩、小毛,感謝這兩年來一起走過的路,充滿歡
笑淚水,都是我美好的回憶。還要謝謝小朱學長的照顧、嘉文的幫忙、 給我力量的小珍、給我歡笑的邱及修致。
最後要感謝我的家人<包含小巨人>,一直以來給予我溫暖、鼓勵、支持, 有你們全心全力,不求回報的支持,我才會有信心與勇氣走下去,感謝 你們。
目錄 摘要 ... I Abstract ... II 致謝 ... III 目錄 ... VI 表目錄 ... VIII 圖目錄 ... IX 附表目錄 ... X 前言 ... 1 第一章 研究動機與目的 ... 4 第一節 非酒精性脂肪變性肝炎 ... 4 第二節 兒茶素簡介 ... 7 第三節 兒茶素影響脂質代謝之可能機轉 ... 9 第四節 兒茶素影響脂氧化壓力之可能機轉 ... 11 第五節 兒茶素影響發炎反應之可能機轉 ... 13 第六節 菊苣糖影響脂質之可能機轉 ... 14 第七節 菊苣糖影響脂肪變性之可能機轉 ... 16 第二章 材料與方法 ... 17 一、實驗動物 ... 17 二、實驗設計 ... 17 三、樣品收集 ... 18 四、分析項目 ... 21 五、統計方法 ... 31 第三章 結果 ... 32 一、NASH 疾病動物誘發 ... 32 二、攝食量、體重 ... 32 三、口服葡萄糖耐受試驗 ... 33 四、血液生化分析 ... 33 五、脂肪組織重量 ... 35 六、肝臟生化分析 ... 35 七、肝臟病理分析 ... 38 八、糞便生化分析 ... 38
第四章 討論 ... 39 第五章 總結 ... 48 參考文獻 ... 49
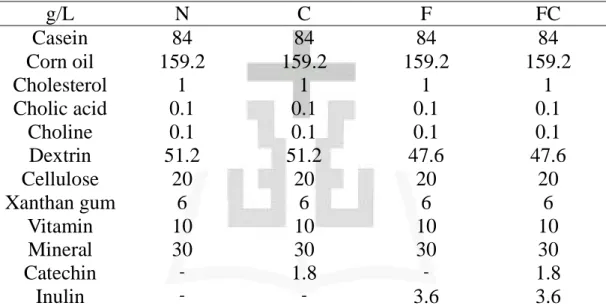
表目錄 表一、各組實驗飲食...57 表二、餵食 NASH 誘發飲食第十二週血液生化值 ...58 表三、各組體重、攝食量、攝食效率 ...59 表四、各組血液生化值...60 表五、各組肝臟重量、肝臟與體重比、脂肪組織、肝臟脂質濃度...61 表六、各組糞便總膽固醇和膽酸濃度 ...62 表七、各組肝臟脂質過氧化物濃度及抗氧化酵素活性 ...63 表八、各組肝臟 TNF-α、Hydroxyproline 濃度和骨髓過氧化酶活性64
圖目錄
圖一、餵食 NASH 誘發飲食第十二週肝臟病理切片- H&E stain ...65
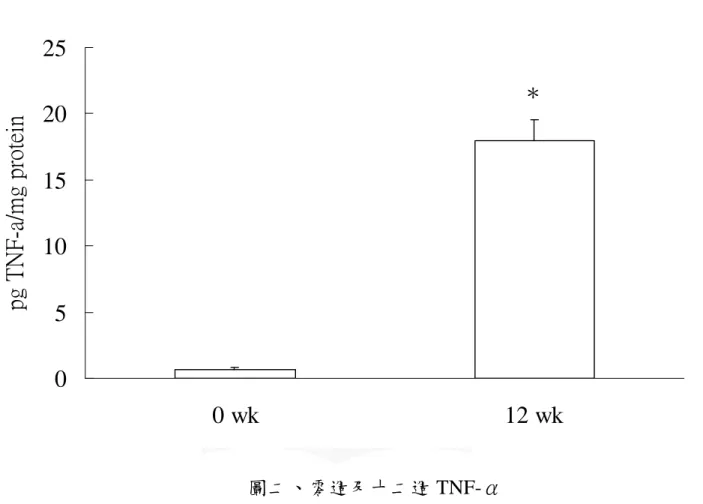
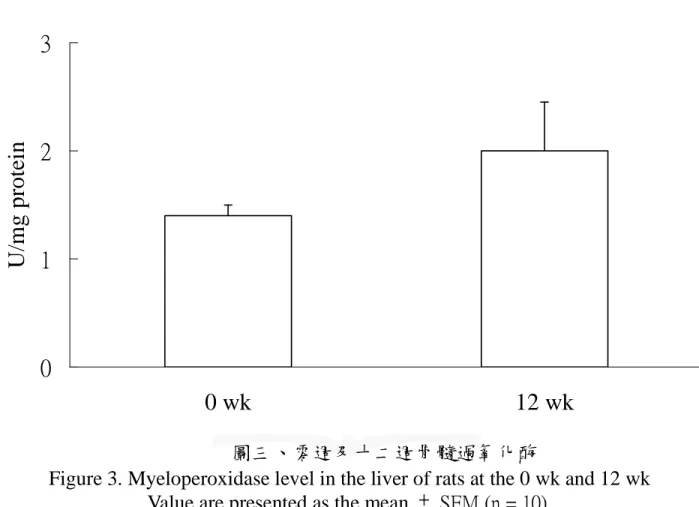
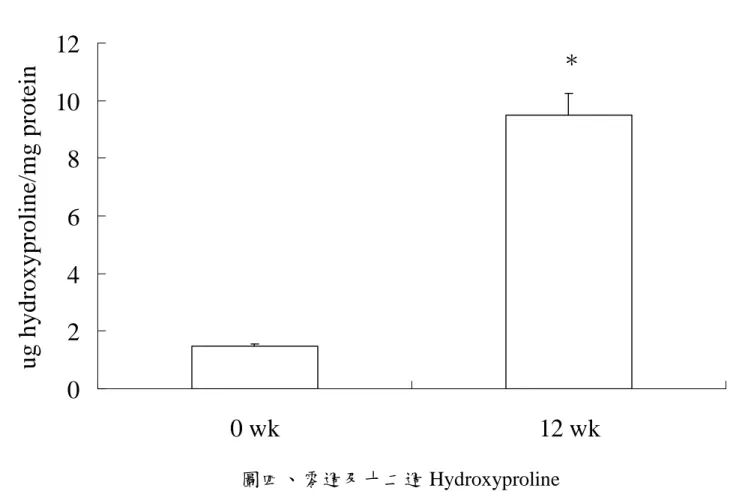
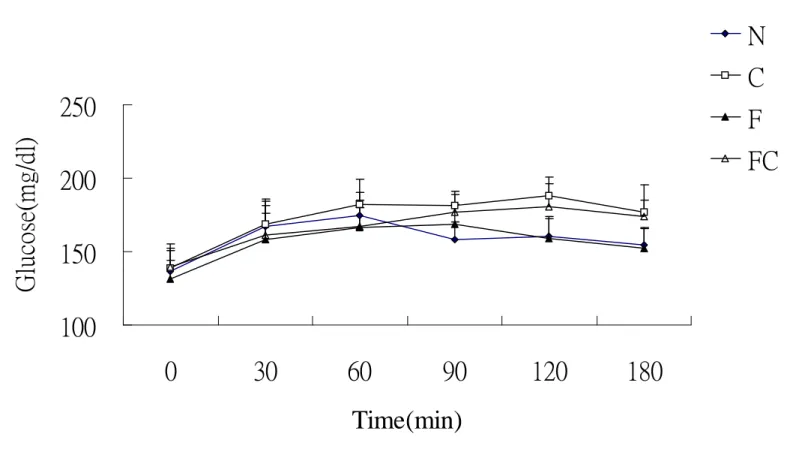
圖二、零週及十二週 TNF-α...66 圖三、零週及十二週骨髓過氧化酶 ...67 圖四、零週及十二週 Hydroxyproline...68 圖五、口服葡萄糖耐受性試驗-血漿葡萄糖濃度變化...69 圖六、口服葡萄糖耐受性試驗-血漿胰島素濃度變化...70 圖七、肝臟病理切片- H&E stain...71 圖八、各組血漿游離脂肪酸濃度 ...72 圖九、各組肝臟游離脂肪酸濃度 ...73 圖十、各組肝臟 CYP2E1 蛋白質表現量 ...74 圖十一、肝臟病理切片- Silver stain...75
附表目錄
前言 肝臟是身體內代謝功能的主要器官,在體內負責儲存肝醣、製造 膽汁、消化、排除多種代謝產物,每天攝取的食物經過消化後的產物 會直接運送進入肝臟,經過代謝後,產生維持身體運作的營養素。因 此若是肝臟受損,對健康的影響既深且廣。肝臟疾病一向是國人常見 的疾病,對於國人的健康造成很大的威脅,根據行政院衛生署的統計, 台灣地區 2006 年十大死因中,慢性肝病和肝硬化位居十大死因第七 位。肝臟疾病的類型和發生的原因很多,根據研究顯示:非酒精性脂 肪肝病 (Non-alcohol fatty liver disease, NAFLD) 為肝臟疾病中常見的 一種 (Bacon et al., 1994),發生於沒有飲酒習慣的患者,其肝臟細胞有 脂質堆積的現象;廣義來看,NAFLD 除了單純的脂肪變性之外,還涵 蓋了脂性肝發炎、肝纖維化,甚至於肝硬化等各種不同嚴重程度的肝 臟疾病,其肝組織學變化與酒精性脂肪肝相似,病變主體在肝小葉, 並且以肝細胞脂肪變性和脂肪蓄積為病理特徵的臨床綜合徵狀,其發 生率有逐年增加的趨勢 (Mulhall et al., 2002)。
非酒精性脂肪變性肝炎 (Nonalcoholic steatohepatitis, NASH) 和體 內脂質代謝的異常有關,會產生肝臟脂肪堆積的情形,最後甚至會進
展為不可回復性的肝纖維化或肝硬化,在 NAFLD 進展至硬化的過程 中為主要關鍵。NASH 於 1980 年由 Ludwig 等所提出,主要描述無 飲酒史病患的肝臟切片,在組織學上的變化及臨床表徵皆與酒精性肝 炎相似 (Ludwig et al., 1980)。流行病學調查發現,肥胖、高脂血症、 糖尿病及高血壓等代謝異常的疾病,是造成 NASH 的危險因子。研究 認為可能與這些疾病所造成的胰島素抗 性有關,因胰島素抗性會進而 影響脂質代謝 (Marchesini et al., 1999)。大部分 NASH 病患皆無明顯 的症狀,因此在疾病發生初期很容易被忽略,以往利用抗氧化劑,例 如:維生素E或維生素C來降低其氧化傷害,或者是利用藥物來改善胰 島素抗性及提高肝細胞的存活率 (Harlander et al., 2001)。 綠茶中的多酚類,近年來因具有特殊生理活性而受到矚目,在先 前的動物實驗中指出,綠茶中的多酚類可藉由抑制膽固醇合成及改善 高三酸甘油酯血症,達到調節血脂的作用 (Bursill et al., 2006)。除此之 外,兒茶素也具有抗氧化作用,具有清除過氧化氫及降低體內氧化壓 力等作用 (Kurihara et al., 2004)。茶飲料雖然長久以來廣泛的被飲 用,但是目前並沒有研究直接討論兒茶素與 NASH 之間的關係,推論 上述功效可能與改善 NASH致病機轉相關。因此本研究以兒茶素介入 實驗動物飲食中,觀察兒茶素對於 NASH 的影響。
然而除了高脂,高多元不飽和脂肪酸飲食之外,針對 NASH 病患 的飲食習慣進行調查研究中發現,和一般人比較,NASH 病患在飲食 上攝取較多膽固醇,以及較少的纖維 (Musso et al., 2003)。目前已有研 究證實,飲食中的水溶性膳食纖維具有降低血脂的作用 (Cho et al., 2007),並且還可以促進脂質代謝,而且有研究更進一步證實果寡糖具 有改善肝臟脂肪變性作用 (Daubioul et al., 2005),因此推論水溶性膳食 纖維的攝取或許對於 NASH 病程的改善具有功效。 因此本研究以NASH疾病模式動物,來探討飲食中兒茶素的補充, 是否可以藉由降低氧化壓力、改善脂肪變性及肝臟發炎情形,以延緩 NASH 的疾病進程。另外添加菊苣糖 (inulin) 於飲食中,觀察其對於 NASH 的改善作用是否具有加乘效果。
第一章 研究動機與目的
第一節 非酒精性脂肪變性肝炎
非酒精性脂肪變性肝炎 (Non-alcoholic steatohepatitis, NASH),這 個名詞首先於 1980 年被提出 (Ludwig et al., 1980),主要描述一群無飲 酒 史 的 女 性 糖 尿 病 患 者 , 其 肝 臟 病 理 表 現 卻 有 大 泡 型 脂 肪 (macrovesicular fat) 、 肝 細 胞 壞 死 、 類 竇 旁 纖 維 化 (perisinusoidal fibrosis)、肝細胞發炎、ballooning degeneration 及 mallory hyaline 等類 似於酒精性肝炎的現象。
1998 年有學者提出「Two hit hypothesis」,加以說明 NASH 的疾 病進程,第一步:肝臟細胞脂質大量堆積,第二步:肝臟中氧化壓力 增加 (James et al., 1998)。因此,肝細胞中脂質的堆積是造成肝傷害的 先決條件,根據先前的研究指出:肥胖、糖尿病、高血壓、高血脂等 代謝異常的疾病,皆為造成 NASH 的危險因子,並指出可能與這些疾 病所造成的胰島素抗性有關 (Marchesini et al., 1999)。當胰島素抗性產 生時,會進一步的影響肝臟中脂質的調節作用,引起脂肪變性、氧化 傷害等負面影響。正常情況之下,肝臟中脂肪的合成與代謝會維持一
時,就會影響肝臟脂質的代謝,進而提高脂肪酸的合成,同時造成脂 肪酸的β-oxidation 受阻 (Saito et al., 2007);另一方面,會加強 ApoB 蛋白質水解作用,使得極低密度脂蛋白合成能力降低,而無法順利將 脂肪送出肝外,脂肪酸便會以三酸甘油酯的形式堆積於肝臟中 (Powell et al., 1990),導致大量脂肪蓄積於肝臟中。當肝臟中脂質堆積時,會降 低 PPARs (peroxisome proliferators-activated receptors) 的表現,PPARs 是一群在細胞核內調節脂肪細胞基因表現和分化之主要轉錄因子,在 醣類和脂肪代謝過程中扮演重要角色,當 PPARs 的表現降低時,會降 低胰島素感受性,使得肝臟及組織脂質堆積情形更為顯著 (Gelman et al., 2007)。 然而單純的脂肪變性並不會造成肝臟的損傷,必須有第二種傷害 的存在,才會使脂肪肝進展至脂肪變性壞死及纖維化,會使肝臟中 Collagen 指標-Hydroxyproline 濃度增加 (Sun et al.,2007),根據先前的 研究結果,氧化壓力的增加是造成脂肪變性肝炎的第二種傷害。當肝 臟中存在過量脂肪酸,會使脂肪酸的氧化作用增加,產生大量的超氧 陰離子 (superoxide anions) 及過氧化氫 (hydrogen peroxide)。此外,大 量 的 脂 肪 酸 也 會 誘 發 Cytochrome P450 family 的 表 現 , 特 別 是
有密切的關係。而這些活性氧屬會攻擊細胞膜上的多元不飽和脂肪 酸,並引發脂質過氧化作用進行,生成大量的脂質過氧化物,如 malondialdehyde (MDA) (Robertson et al., 2001),進而活化肝臟星狀細 胞 , 而 星 狀 細 胞 可 藉 由 活 化 NF-κB, 使 前 發 炎 細 胞 激 素 (proinflammatory cytokines) 大量分泌,促進發炎反應 (Letteron et al., 1996; James and Dat, 1998; Browning and Horton, 2004)。
在脂肪變性發炎過程中,氧化壓力及脂質過氧化與細胞激素間的 交互作用,扮演了重要角色。研究指出,NASH 病患體內有較高腫瘤 壞死因子 (tumor necrosis factor-α,TNF-α) 的表現,而 TNF-α 可誘發 其他細胞激素,如 IL-6 (interleukin-6) 及 IL-8 (interleukin-8) 的產生, 加重肝細胞發炎情況,導致肝細胞壞死 (Crespo et al., 2001)。另一方 面,在肥胖小鼠的研究中發現,因脂質堆積所造成的粒線體功能不全, 會提高肝細胞對 TNF-α cytotoxicity 的敏感性,進而加重肝細胞的損害 (Tilg and Diehl, 2000)。
大部分NASH病患皆無明顯的症狀,因此在疾病發生初期很容易被 忽略,以往利用抗氧化劑,如維生素 E 或維生素 C 來降低其氧化傷 害 , 或 者 是 利 用 藥 物 來 改 善 胰 島 素 抗 性 及 提 高 肝 細 胞 的 存 活 率
的飲食介入方法可以延緩 NASH 的進程,是本研究探討的主題。 第二節 兒茶素簡介 茶飲料的飲用歷史悠久,其中不經過發酵過程的綠茶,含有最多 兒茶素 (Jin et al., 2006)。乾燥的綠茶中,多酚類佔約 36% (w/w),綠 茶 中 的 多 酚 類 大 多 為 兒 茶 素 , 而 這 些 兒 茶 素 則 屬 於 黃 酮 類 (flavonids),含有兩個芳香環 (C6-C3-C6),兒茶素主要有四種結構異構 物:(-)-epicatechin (EC)、(-)-epigallocatechin (EGC)、(-)-epicatechin gallate (ECG)、(-)-epigallocatechin gallate (EGCG),佔兒茶素的 90% 以上。 其中的 EGCG,因含有八個氫氧基,最具有生理活性,佔有總兒茶素 的 54~59% (Kobayashi et al., 2005)。亞洲民族飲用綠茶的歷史悠久,近 年來綠茶中的多酚類因具有特殊生理活性而受到矚目,目前許多研究 針對綠茶中兒茶素的抗肥胖及降血脂等功效;研究結果顯示,綠茶中 的多酚類,可以增加人體及動物體內棕色脂肪組織 (Brown adipose tissue) (Dulloo et al., 2000),脂質的氧化作用及攝食產熱效應。研究證 實,綠茶中的兒茶素會藉由抑制 catechin-O-methyltransferase (會降解 正腎上腺素的酵素) ,達到抑制磷酸脂解酶對於 cAMP 的降解,進而
促進了體內脂肪的氧化作用 (Dulloo et al., 1999)。
除此之外,綠茶中的兒茶素也會透過促進脂質代謝的酵素活性, 包 括 : 透 過 PPARs 所 調 控 的 Acyl-CoA oxidase 及 Acyl-CoA dehydrogenase,進而促進脂質的代謝 (Murase et al., 2002),綠茶中的兒 茶素在動物實驗中證實具有降低血漿膽固醇濃度活性 (Raederstorff et al., 2003)。另一方面,許多研究證實綠茶中的多酚類可以增加大鼠糞便 中膽固醇的排出,藉由減少飲食中膽固醇的吸收,減少血漿中膽固醇 濃度;除此之外,綠茶中多酚類還具有抗氧化能力、清除自由基的能 力,而且其抗氧化能力與含有兒茶素的量呈正相關性 (Yamamoto et al., 2006)。另外,兒茶素也具有減少冠狀動脈疾病的發生及抗癌能力,研 究證實兒茶素具有減少動脈膽固醇堆積 (Steinberg et al.,1989),同時動 物實驗也證實具有減少由高油飲食所導致的體內脂質堆積,進而減少 動脈粥狀硬化疾病的發生。雖然,目前尚無研究針對兒茶素與 NASH 之間的相關性,然而以現有的文獻推論,兒茶素的特殊生理活性可能 會影響 NASH 的致病機轉,因此將透過下列章節詳加討論。
第三節 兒茶素影響脂質代謝之可能機轉 近來茶葉已被證實含有許多生理功效成分,由先前臨床實驗指 出,兒茶素可降低高膽固醇血症病患之血脂質濃度,並且具有抑制膽 固醇合成酶的特殊生理活性,進而降低膽固醇的生合成 (Akanji et al., 1989)。除此之外,茶葉中的兒茶素,還可藉由與飲食中膽固醇的結合, 減少脂質的吸收,進而降低血漿中膽固醇濃度。先前研究提出兒茶素 可藉由與飲食中膽固醇的結合,而達到抑制膽固醇的吸收,並且增加 糞便固醇以及膽酸的排出,影響腸肝循環,加速體內膽固醇代謝的作 用,以利降低血膽固醇 (Yang et al., 2005; Muramatsu et al., 1989)。在內 生性脂質合成過程中,需要許多酵素參與反應,而研究發現兒茶素可 降低 SREBP (Sterol-regulatory element binding protein) 的活性形式,藉 以調節脂質代謝酵素活性,例如:Cholesterol synthase,以降低脂質的 合成的速率 (Bursill et al., 2006)。在動物實驗方面,以大白鼠為實驗對 象,兒茶素的攝取可降低血中總膽固醇,並且降低低密度脂蛋白膽固 醇 (Kuo et al., 2005)。 兒茶素除了可以抑制膽固醇合成以及促進膽固醇代謝之外,還具 有降低血漿當中三酸甘油酯濃度的能力。在動物實驗中發現兒茶素會
藉由抑制小腸中胰脂解酶活性、影響腸胃道的乳糜化作用、減少飲食 中脂肪之吸收,進而降低餐後血漿三酸甘油酯濃度 (Yang et al., 2005)。除了減少飲食中脂肪酸的吸收之外,兒茶素也可透過抑制脂肪 酸的相關合成酵素達到降低三酸甘油酯的合成速率,不論是細胞或是 動物實驗皆證實,兒茶素會藉由減少肝臟中脂肪酸的合成作用 (Ikeda et al., 2005; Fiorini et al., 2005)。許多實驗已證實兒茶素會降低脂肪酸合 成酶活性,在 Wang 等 (2005) 的研究中指出,兒茶素是藉由抑制脂 肪酸合成酶中的 β-ketoacyl reductase 而抑制脂肪酸的合成,進而減少 游離脂肪酸濃度;Bursill 等 (2001) 的研究更進一步指出,兒茶素可 以透過調節 SREBP-1 的表現,減少脂肪酸合成酶 mRNA 的表現,進 而達到減少肝臟中脂質的生合成。營養素代謝產生的 acetyl CoA 會與 草醋酸結合進入 TCA cycle,而後會再藉由 ATP-citrate lyase 的作用, 使 acetyl CoA 釋放至細胞質中,形成 malonyl CoA,進而生成脂肪酸。 因此推論兒茶素可藉由抑制脂肪酸合成酶 (fatty acid synthase, FAS) 的活性,降低脂肪酸的合成,並進一步減少體內以及肝臟中三酸甘油 酯合成,減少血漿脂質濃度。因此推論,兒茶素可以藉由其降血脂特 性,改善因 NASH 所造成的血脂異常。
第四節 兒茶素影響脂氧化壓力之可能機轉 許多研究指出兒茶素具有抗氧化活性,具有清除體內自由基能 力。自由基是人體代謝之代謝產物,過多的自由基會對人體會造成傷 害, 例 如: 低密度脂 蛋白之 氧化以及 脂質過 氧化作用 (Steinberg, 1997)。經研究證實,綠茶中的多酚類具有抗氧化特性,特別是具有八 個氫氧基的 EGCG,其存在量與抗氧化能力成正比,EGCG 可作為 自由基清除者,其化學活性具有提供電子的特性;該結構具有位移的 能力,藉此穩定未配對電子;在它的基本化學結構之 B 環上的 3’、4’ 位置,有兩個相鄰的氫氧基 (-OH),這個結構提供了自身氧化電子位 移時,形成較穩定的酚自由基 (Kondo et al., 1999)。由於 EGCG 在結 構上含有多個羥基,具有提供質子的活性,能與過氧化物作用,進而 減少體內的氧化壓力。 除了直接與自由基的作用之外,另一項兒茶素結構式的特色是: 可以透過 B 環將游離的金屬離子螯合成複合物 (Srichariratanakool et al., 2006),以減少金屬離子之促氧化作用。由於兒茶素具有清除自由基、 螯合金屬離子之能力,因此具有降低體內氧化壓力能力。除此之外, 綠茶中的多酚類,也可以藉由清除體內自由基,而增加抗氧化物的活
性狀態。在動物實驗中證實,同時給予大鼠兒茶素及 vitamin E,發現 兒茶素可避免 α-tocopherol 的氧化作用,進而提升 vitamin E 在體 內的抗氧化作用 (Frank et al., 2006)。因此推論,兒茶素可以以其抗氧 化特性,改善因 NASH 所造成的氧化傷害。
第五節 兒茶素影響發炎反應之可能機轉 近年來有研究認為綠茶中的兒茶素 - EGCG,除了具有降血脂、抗 氧化能力之外,還具有抗發炎的功能。Shin 等 (2007) 的研究結果指 出,EGCG 會藉由抑制 TNF-α 基因表現,而減少體內 TNF-α 的濃 度;在 2005 年的動物實驗中發現,綠茶的飲用可以減少發炎細胞激 素的基因表現,減少肝臟損傷時所增加的發炎細胞激素,例如:TNF-α, IL-1β 基因表現,進而降低血漿中細胞激素濃度,如 TNF-α 所誘發的 IL-6 及 IL-8 的產生 ,減輕肝細 胞發炎情 況所導致的肝細胞 壞死 (Crespo et al., 2001)。因此推論兒茶素可以以其抗發炎的作用,改善因 NASH 所造成的肝細胞損害。
第六節 菊苣糖影響脂質之可能機轉 菊苣糖 (Inulin) 是由菊苣根部所萃取,是以果糖為基本單位所構 成的聚醣類,在末端以 -(1-2) 的方式鍵結成為異果寡醣 ,人類並沒 有分解此結構的酵素,所以攝取後在人體內很難被胃酸水解或體內消 化酵素所消化、吸收,大部分的菊苣糖可以進入人體結腸內,進而被 結腸內的細菌加以發酵利用 (Roberfroid et al., 1993)。菊苣糖會被結腸 中的 bifidobacteria 代謝、發酵,並生成大量短鏈脂肪酸 (SCFA),短 鏈脂肪酸經腸道吸收後可能具有調節脂肪代謝的功效 (Agheli et al., 1998)。因此,菊苣糖的生理活性相當於水溶性膳食纖維,可減短食物 在腸胃道滯留時間、增加飽食感、預防便秘、並且具有降低血漿中膽 固醇三酸甘油酯濃度的作用 (Van et al., 1995)。 目前已有實驗證實,飲食中的水溶性膳食纖維具有降低血脂作 用,根據目前研究認為,果寡糖降血脂的作用,可能是透過下列二個 機轉: (一) 促進脂質代謝: 經由增加腸道雙叉桿菌的增生,增加腸道中細菌膽鹽水解酵素的 活性,促進膽汁及膽固醇在糞便中的排出,進而促進脂質的代謝
(Tahri et al.,1996; Roos and Katan, 2000)。 (二) 影響肝臟脂質生合成:
促進腸道益生菌的增生,進而增加短鏈脂肪酸的產生,影響肝臟 膽固醇的合成限制酶 HMG-CoA reductase 以及脂肪酸合成酶活 性 (Hidaka et al., 1991; Marcil et al., 2003)。
寡糖對於血脂質的影響,目前在動物實驗中比較有一致性的結 果,在動物實驗中發現,寡糖的添加可以降低血清總膽固醇,推測可 能是短鏈脂肪酸使實驗動物肝臟膽固醇的合成減少,以及代謝作用增 加的結果 (Beylot et al., 2005)。另外,在 Roberfroid 等 (1993) 的實驗 中,餵食實驗動物高飽和脂肪酸飲食,再添加果寡糖後發現,可降低 血清膽固醇及三酸甘油酯濃度,同時也減少肝臟中三酸甘油酯的堆 積,推測可能是肝臟利用短鏈脂肪酸的代謝反應,減少游離脂肪酸的 合成,而三酸甘油酯則轉而代謝為酮體。另有研究指出,寡糖發酵後 產 生 之 短 鏈 脂 肪 酸 , 可 被 腸 壁 吸 收 進 入 肝 臟 , 降 低 carnitine acyltransferase 之活性,減少 acyl-CoA 進入粒線體,進而減少三酸甘 油酯之合成 (Aoeh et al., 1986; Hata et al., 1983)。因此推論,菊苣糖可 以以其降血脂特性,改善因NASH所造成的血脂異常。
第七節 菊苣糖影響脂肪變性之可能機轉
根據最近的動物研究指出,飲食中添加菊苣糖,可以減少肝臟中 三酸甘油酯堆積 (Sugatani et al., 2006),改善肝臟脂肪變性情形。研究 中指出攝取菊苣糖,可以增加飽食感,避免過量飲食,同時也會減少 飲食中脂質吸收;除此之外,還可以促進脂質代謝,增加糞便中脂質 的排出 (Verlinden et al., 2006; Cani et al., 2004)。因此認為,菊苣糖可以 藉由減少攝取、降低吸收、增加代謝,進而減少肝臟中脂質的堆積, 改善肝臟脂肪變性情形。若能在飲食中添加菊苣糖,可以改善 NASH 疾病脂肪變性情形。除了飲食中的攝取之外,NASH 疾病動物模式的 胰島素抗性,使NASH疾病動物模式脂質代謝異常,導致體內脂質堆 積。研究指出,菊苣糖可以藉由減少葡萄糖吸收,延緩餐後血糖上升, 降低胰島素濃度,進而改善胰島素抗性所造成的脂質代謝異常,減少 肝臟脂質堆積,改善 NASH 疾病脂肪變性情形 (Daubioul et al., 2005; Kaur et al., 2002)。因此認為菊苣糖可以以其生理特性,改善因 NASH 所造成的脂肪變性。
第二章 材料與方法
一、實驗動物
Wistar 品系雄性大白鼠 60 隻,購自國家實驗動物繁殖及研 究中心,置於室溫 23 ± 2°C,溼度控制於 55% ± 10%,12 小時明暗更 替的飼育室內,給予固型飼料 (Rodent Laboratory Chow 5001) 及去離 子水,預養二週後進行為期十八週的實驗。實驗飼料組成主要參考 Lieber 等 (2004) 發表之 NASH 疾病動物誘發飲食為基礎。首先給予 大白鼠十二週液態高油高膽固醇飲食以誘發 NASH,於零週及第十二 週進行動物犧牲(n = 20),之後依體重隨機分為 4 組 (n = 10),分組 如下,N 組 (NASH 誘發飲食)、C 組 (NASH 誘發飲食 + 兒茶素)、 F 組 (NASH 誘發飲食 + Inulin)、FC 組 (NASH 誘發飲食 + 兒茶素 + Inulin),其中 C、F 與 FC 組之兒茶素和菊苣糖是額外添加 0.5% 兒 茶素或 1% 菊苣糖,各組詳細飼料配方如表一所示。
二、實驗設計
每週測量體重。十二週後依體重平均分為四組,分別給予 NASH 誘發 飲食、NASH 誘發飲食 + 兒茶素、NASH 誘發飲食 + 菊苣糖、NASH 飲食 + 兒茶素 + 菊苣糖,攝食量及體重測量方法同上。於實驗零週 和實驗第十二週進行動物抽血,並且分別犧牲十隻,作為基準值以及 確立疾病動物模式的成立,於第十七週進行口服葡萄糖耐受試驗,第 十八週犧牲前收集糞便樣本。實驗末犧牲動物,採集血夜樣本並取出 實驗動物肝臟與體脂肪組織進行分析。 三、樣品收集 血液 第 十 七 週 進 行 口 服 葡 萄 糖 耐 受 試驗 , 經 口 投 予 葡萄 糖 溶 液 (1 g glucose/kg-BW),分別於第 0、30、60、90、120 及 180 分鐘由實驗 動物尾靜脈採血,血液收集於含有 EDTA 的試管中,充分混合均勻 後,以 1,700 ×g、4°C 離心 15 分鐘後,將血漿保存於 -70°C 之冷凍 櫃,以待分析血糖和胰島素濃度。於第十二週以及實驗末則將動物以 巴比妥鈉鹽 (50 mg/kg-BW) 麻醉後犧牲,開腹腔由腹腔動脈採血,血 液收集於含肝素的試管中充份混合後,以上述相同條件離心,將血漿
保存於 -70°C 之冷凍櫃,留待日後分析。 肝臟 肝臟病理分析:動物犧牲後,以 0.9% 生理食鹽水由門靜脈灌流後將 肝臟摘除,再以生理食鹽水清洗,剝除多餘的結締組織後,以濾紙拭 淨、秤重,剪取最大葉肝臟,置於 19% 福馬林中固定處理,經脫水製 成蠟塊,薄切為組織切片,以蘇木紫-伊紅進行染色 (haematoxylin & eosinstain; H & E stain)、Masson trichrome stain、Silver stain,置於光學 顯微鏡下觀察。肝臟標本由病理醫師判讀,依據 Teramoto 等 (1993) 的方法,評估大白鼠肝臟細胞內脂肪顆粒堆積的程度,以及肝臟組織 纖維化程度。剩餘肝臟以鋁箔紙包住,以夾鏈袋密封,儲存於 -70°C, 留待日後分析。 肝臟脂質分析:依照 Folch 等 (1957) 方法,剪取 1 克肝臟組織,加 入萃取溶劑 (chloroform:methanol = 2:1, v/v),在冰浴中以均質機磨 碎後,過濾至有蓋試管,以萃取液定量至 10 mL,加入 2 mL 之 0.05% CaCl2 (w/v) , 離 心 (3500×g, 4°C) 3 分 鐘 後 , 去 除 上 層 液 , 再 以 chloroform:methanol:water = 3:48:47 (v/v/v) 定量至 12 mL,離
心 (3500×g, 4°C) 3分鐘後,去除上層液,先加入甲醇定量至 10 mL, 再以萃取溶劑 (chloroform:methanol = 2:1, v/v) 定量至 25 mL,收集 於樣品瓶中,儲存於 -20°C。總膽固醇和三酸甘油酯以市售試劑組 (Randox laboratories kit) 進行分析。
肝臟抗氧化分析:剪取 0.5公克肝臟組織,與4°C之緩衝液 (0.25 M sucrose, 1 mM EDTA, 10 mM Tris-HCl) 混合,經均質化後,離心 10 分 鐘 (4500×g, 4°C),分離肝臟細胞質部分,儲存於 -80°C 下待分析。 MDA、catalase 及 SOD 分析方法同前。 體脂肪組織 動物犧牲後,取其副睪兩側與腎臟周圍的體脂肪,以生理食鹽水清洗, 再以濾紙拭淨後秤重。 糞便 於第十七週收集動物糞便,將收集之糞便冷凍乾燥後,以研缽研磨, 置入夾鏈袋密封,儲存於 -20°C,留待日後分析。
四、分析項目
血液
(1) 血漿總膽固醇濃度之測定
以全自動分析儀 (HITACHI 7170 Autoanalyzer, Tokyo, Japan) 分 析,分析原理如下:以 cholesterolesterase 將血漿中 cholesterolester 分解成膽固醇和脂肪酸,此膽固醇和原本存在的膽固醇一起受 cholesteroxidase 作用,產生 H2O2 。此 H2O2 在 peroxidase 存在
下和 4-aminoantipyrin 及 DAOS (3.5- dimethoxy- N- ether- N- (2-hydroxy-3-Sulfoproyl)-aniline) 作用,氧化縮合產生青紫色色素,測 其吸光度變化換算成膽固醇濃度。
(2) 血漿三酸甘油酯濃度之測定
以全自動分析儀 (HITACHI 7170 Autoanalyzer) 分析,分析原理如 下 : 採 用 Free Glycerol Blanking Method , 即 三 酸 甘 油 酯 受 lipoprotein lipase 作 用,產 生 glycerol 後加 入 glycerokinase 與 glycerol-3-phosphateoxidase,產生 H2O2。測 H2O2 與 peroxidase、
吸光度變化換算成三酸甘油酯濃度。 (3) 血漿高密度脂蛋白膽固醇濃度之測定
以全自動分析儀 (HITACHI 7170 Autoanalyzer) 分析,分析原理如 下:使用 Homogeneous assay,以 dextran sulfate 與鎂離子將高密 度脂蛋白以外的脂蛋白沉澱後,以上述分析膽固醇的方法分析上層 懸浮液中的膽固醇含量,即高密度脂蛋白膽固醇濃度。 (4)血漿低密度脂蛋白膽固醇濃度之測定 以全自動分析儀 (HITACHI 7170 Autoanalyzer) 分析,分析原理如 下:利用試劑破壞所有非低密度脂蛋白後,再無色反應下移除釋出 的膽固醇。然後再以試劑與低密度膽固醇作用使其釋出膽固醇,此 膽固醇以上述的呈色反應定量。 (5)血漿游離脂肪酸濃度之測定
以市售試劑組 (Randox FA115, Antrim, UK) 進行分析。取血漿或標 準液 50 μL 與反應劑 1000 μL 混勻,37°C 下靜置 10 分鐘,再加入 酵素反應劑1000 μL 混勻,同樣在 37°C 下靜置 10 分鐘,以分光光 度計在 550 nm 波長下測量吸光值,與標準液吸光值對照而得非酯 化脂肪酸濃度,即為血漿游離脂肪酸濃度。
以全自動分析儀 (HITACHI 7170 Autoanalyzer) 分析,分析原理如 下:在反應劑中加入適量緩衝液混勻備用,之後於 100 μL 血漿中 加入 1 mL 的稀釋反應劑,混合均勻,讀取第 1、2、3、4 分鐘的 吸光值 (340 nm, 37°C)。計算公式: U/I= 1746×吸光值變化量。 (7)血漿葡萄糖濃度之測定
以市售試劑組 (Randox GL1611, Antrim, UK) 進行分析。取血漿或 標準液 10 μL 與反應劑 1000 μL 混勻,37°C 下靜置 10 分鐘, 以分光光度計在 500 nm 波長下測量吸光值,與標準液吸光值對照 而得血漿葡萄糖濃度。
(8)血漿胰島素濃度之測定
使用市售試劑組 (Rat Insulin Mercodia 10-1124-01, Uppsala, Sweden) 以酵素免疫分析法 (ELISA),在 450 nm 波長下,與標準溶液之 吸光度對照,而得胰島素濃度。
肝臟
(1) 肝臟膽固醇濃度之測定
取肝臟萃取液 10 μL,先以減壓濃縮機抽乾後,以市售試劑組 (Randox CH201, Antrim, UK) 進行分析。加入反應劑 1000 μL 混
勻,37°C 下水浴 5 分鐘,以分光光度計在 500 nm 波長下測量吸光 值,與標準液吸光值對照而得肝臟膽固醇濃度。
(2) 肝臟三酸甘油酯濃度之測定
取肝臟萃取液 10 μL,先以減壓濃縮機抽乾後,以市售試劑組 (Randox TR213, Antrim, UK) 進行分析。加入反應劑 1000 μL 混 勻,37°C 下水浴 5 分鐘,以分光光度計在 500 nm 波長下測量吸光 值,與標準液吸光值對照而得肝臟三酸甘油酯濃度。
(3) 肝臟游離脂肪酸濃度之測定
以市售試劑組 (Randox FA115, Antrim, UK) 進行分析。取肝均質液 或標準液50 μL 與反應劑 1000 μL 混勻,37°C 下靜置 10 分鐘,再 加入酵素反應劑 1000 μL 混勻,同樣在 37°C 下靜置 10 分鐘,以分 光光度計在 550 nm 波長下測量吸光值,與標準液吸光值對照而得 非酯化脂肪酸濃度,即為肝臟游離脂肪酸濃度。 (4) 肝臟抗氧化酵素與過氧化物質 肝臟 Catalase 活性測定:依據 HugoAebi (1947) 的方法測定,以 50 mM phosphate buffer 將肝臟均質液稀釋至適當稀釋倍率後,取 400 μL 的待測肝臟均質稀釋液加入 30 mM H2O2 200 μL,於 240 波長下測一分鐘之吸光值變化,得 H 減少之速率,代入
公式推算活性,並以 Lowery method 來測定肝臟均值稀釋液的蛋白 質含量來進行蛋白質含量的校正。
肝臟 superoxidase dismutase (SOD) 活性之測定:以市售試劑組 (Rondox SD125, Antrim, UK) 分析,取 25 μL肝臟均質液加入 850 μL 的混合基質液 (mixed substrate) 及 125 μL 的 xanthine oxidase 溶 液,混合均勻後,在 37°C、505 nm 波長下測定 30 秒後的吸光值 (A1),3 分鐘後,測量吸光值 (A2),利用吸光值變化量及與標準品
比較後求得濃度,並以 Lowery method 測定肝臟均質稀釋液的蛋白 質含量以進行蛋白質含量的校正。
肝臟過氧化物質濃度之測定:使用市售試劑組 (Lipid peroxidation assay kit, CALBIOCHEM®, San Diego, CA) 分 析 , 以 50 mM phosphate buffer 將肝臟均質液稀釋至適當稀釋倍率後,取 200 μL 的待測肝臟均質稀釋液或標準液加入 650 μL 反應劑混勻,再加入 150 μL 之 15.4 M methane sulfonic acid 混勻,45°C 下水浴 40 分 鐘,取出後立即冰浴,以分光光度計在 586 nm 波長下測量吸光 值 , 與 標 準 液 吸 光 值 對 照 而 得 malondialdehyde (MDA) + 4-hydroxyalkenals (4-HNE) 濃度。並以 Lowery method 來測定肝 臟均值稀釋液的蛋白質含量來進行蛋白質含量的校正。
肝臟 Glutathione reductase 活性之測定:使用市售試劑組 (Randox GR 2536, Goldberg, DM) 分析,將肝臟均質液稀釋至適當稀釋倍率 後,取 5 μL 的待測肝臟均質稀釋液加入 125 μL 基質溶液混勻, 加入 25 μL NADPH,混合均勻以分光光度計,於 37°C,340 nm 下 測量五分鐘內吸光值變化。
肝臟 Glutathione reductase 活性之測定:使用市售試劑組(Randox RS 504, Paglia)分析, 將肝臟均質液稀釋至適當稀釋倍率後,取 4 μL 的待測肝臟均質稀釋液加入 200μL 反應劑混勻,以及 Cumene hydroperoxide,混合均勻,於 37°C, 340 nm 下測量三分鐘內吸光 值變化。 (5) 肝臟組織之微粒體 剪取 0.5 g 肝臟組織,加入 2.5 mL 的均質緩衝液 (0.25 M sucrose, 10 mM Tris-HCl, 0.25 mM Phenyl-methyl-sulfonyl fluoride),經均質 化後,離心 20 分鐘 (10000 ×g, 4°C),取出上層液分裝至超高速離 心機專用離心管 (Beckman 362305, 13×56 mm, 3.2 mL),離心 1 小 時後 (105,000×g, 4°C),去除上層液,將下層沉澱物懸浮於 50 mM potassium phosphate buffer 中,即得微粒體懸浮液。
polyacrylamide gelelectro-phoresis):微粒體懸浮液蛋白質定量後, 將蛋白質濃度調整為 10 mg/mL ,取 10 μL 樣 品加入等 體積 Protein dye 混合均勻,於 100°C 條件下加熱 5 分鐘,之後移至 4% acrylamide gels 樣品槽中,以 100 伏特電壓進行電泳分析。 西方點墨法 (Western blotting) 與免疫分析:將所得之電泳膠片, 利用濕式轉印槽,於固定電流 80 mA 下轉印 18 小時,將膠片上 的 蛋 白 質 轉 印 至 轉 印 膜 (polyvinylidene difluoride membrane, PVDF)。將含有蛋白質的 PVDF 放入含 5% 脫脂牛奶的 blocking buffer,4°C 下隔夜。取出 PVDF 以 PBS-T buffer 沖洗三次,每 次 5 分鐘,接著加入以 PBS-T buffer 稀釋 100 倍的 CYP2E1 一 級單株抗體 (monoclonal anti-rat CYP2E1 antibody, Oxford PM32), 置於室溫下作用 2 小時;之後取出 PVDF,同樣以 PBS-T buffer 沖洗三次,每次 5 分鐘,再加入以 PBS-T buffer 稀釋 5,000 倍的 二 級 抗 體 (goat anti-mouse IgG peroxidase conjugated antibody, Chemicon AP124P),置於室溫下作用 2 小時之後取出 PVDF,以 PBS-T buffer 沖洗三次,每次 5 分鐘;最後加入 1 mL 的冷光劑 (chemiluminescence reagent, Western lightingTM, Perkinelmer),作用 1 分 鐘 後 , 用 保 鮮 膜 包 覆 後 固 定 於 壓 片 匣內 ,於 暗 房 中 將 Fuji
medicalx-ray film 放入壓片匣進行壓片,經適當時間反應後將其取 出放入沖片機 (Konica, RX-1300, Japan) 沖洗底片,底片經掃描後 由 Image-ProPlus 軟體分析其蛋白質表現量,並以 internal control 校正。 (6) 肝臟 TNF-α 濃度之測定 將冷凍的肝臟取出解凍,依 Das 等 (2005) 方法,剪取 0.5 g 肝臟 組織,加入 1.5 mL 均質液 (50 mM Tris-base, 150 mM NaCl, 1%Triton-X100) 混合,經均質化後,於 4°C 下震盪 90 分鐘,後 離心 15 分鐘 (3000×g, 4°C),取出上層液,儲存於 -80°C 下待分 析 。 使 用 市 售 試 劑 組 (Rat TNF-α/TNF SF1A, DuoSet®
ELISA Development System, Abingdon, UK) 以酵素免疫分析法 (ELISA), 在 450 nm 波長下,與標準溶液之吸光度對照,而得 TNF-α 濃 度。 (7) 肝臟中骨髓過氧化酶 (Meyloperoxidase) 剪取 0.2 g 肝臟組織,與 4°C 之緩衝液 (0.5% hexadecyltrimethyl ammonium bromide) 混合,經均質化後,離心 5 分鐘 (12500×g, 4°C),取上清液,儲存於 -80°C 下待分析。取 10μL 肝臟上清均質 液,加入 280 μL working reagent(1.5% H 5.26 mM O-dianisidine
hydrochloride50 mM potassium phosphate buffer),於 460 nm 下測 量 5 分鐘吸光稙變化,觀察 5 分鐘內酵素活性變化。 (8) 肝臟中 Hydroxyproline 含量 剪取 5 g 肝臟組織,將肝臟組織冷凍乾燥,再稱取 0.25 g 冷凍乾 燥之肝臟以 1 mL 蒸餾水均質,儲存於 -80°C 下待分析。取 25 μL肝均質液或標準品容液 (2-100 μg) 加 25 μL 4 N NaOH (終濃 度 2N),於 120°C 下水解 20 分鐘,加入 450 μL chloramine T 混合均勻,於室溫下氧化 25 分鐘,再加入 550 μL Ehrichs ragent 混和均勻,於 65°C 下水浴 20 分鐘以呈色 (紅色),於 550 nm 下 測吸光值,與標準溶液之吸光度對照,而得 Hydroxyproline 濃度。 糞便 (1)中性固醇 依據 Folch 等 (1957) 方法,精秤 0.5 g 糞便粉末,加入 15 mL 萃 取溶劑 (chloroform : methanol = 2:1, v/v),振盪後以濾紙過濾,於濾 液中加入 0.05% CaCl2 混勻,脫氣 15 分鐘,離心 (3500 xg, 4°C) 5 分鐘後,去除上清液,再以萃取溶劑定量至 25 mL,收集於褐色塑 膠離心管中,儲存於 -20°C 備用。取糞便中性固醇萃取液 10 μL,
先以減壓濃縮機抽乾後,以市售試劑組 (Randox CH201, Antrim, UK) 進行分析。加入反應劑 1000 μL 混勻,37°C 下水浴 5 分鐘,以分 光光度計在 500 nm 波長下測量吸光值,與標準液吸光值對照換算而 得糞便中性固醇濃度。 (2)膽酸 依據 Grundy 等 (1965) 方法,精秤 0.1 g 糞便粉末至 2 mL 塑膠離 心管中,加入 1 mL 無水酒精混勻,脫氣 15 分鐘後,離心 15 分 鐘 (3500×g, 20°C),取上清液以減壓濃縮機抽乾,重複操作兩次;於 無水酒精已蒸乾之試管內再加入 1 mL 石油醚混勻,脫氣 15 分 鐘,離心 (3500×g, 20°C) 15 分鐘後,去除上清液,重複操作兩次, 將沉澱物以減壓濃縮機抽乾後,以甲醇溶解定量至 2 mL,離心 (3500×g, 20°C) 15 分鐘後,再取上清液收集於褐色塑膠離心管中, 並儲存於 -20°C 備用。取糞便膽酸萃取液 200 μL,以市售試劑組 (Randox BI1605, Antrim, UK)進行分析,利用 3α-hydroxysteroidde hydrogenase 及 diaphorase 與 bile acids 作 用 , 產 生 紫 藍 色 formazan 衍生物,在 540 nm 波長下測量吸光值,對照標準曲線求 得濃度。
五、統計方法
實驗結果以 SAS (the Statistical Analysis System, version 8.2) 統計軟體 進行分析,所有數據均以 Mean ± SEM 表示,不同組間之差異採用變 異數分析 (Analysis of variance, ANOVA) 中之 One-way ANOVA 進 行分析,並以 least significant difference (LSD) 進行組間比較,以 p 值 小於 0.05 表示統計上有意義。
第三章 結果 一、NASH 疾病動物誘發 1) 第十二週血液生化值 給予 NASH 誘發飲食十二週,實驗動物的血液生化值如表一。結果 顯示:肝功能指數 (GOT、GPT) 酵素活性顯著高於零週 (p < 0.05), 同時血漿中膽固醇濃度以及低密度蛋白膽固醇濃度顯著高於零週 (p < 0.05),並且顯著的降低血中高密度脂蛋白膽固醇與低密度脂蛋白 膽固醇比例 (p < 0.05)。 2) 第十二週肝臟切片 給予 NASH 誘發飲食十二週,對於肝臟脂肪空泡堆積與發炎情形的影 響如圖一所示。經由病理醫師判讀結果,給予 NASH 誘發飲食十二週 之肝臟脂肪空泡堆積程度有明顯增加情形。 二、攝食量、體重 1) 每日攝食量
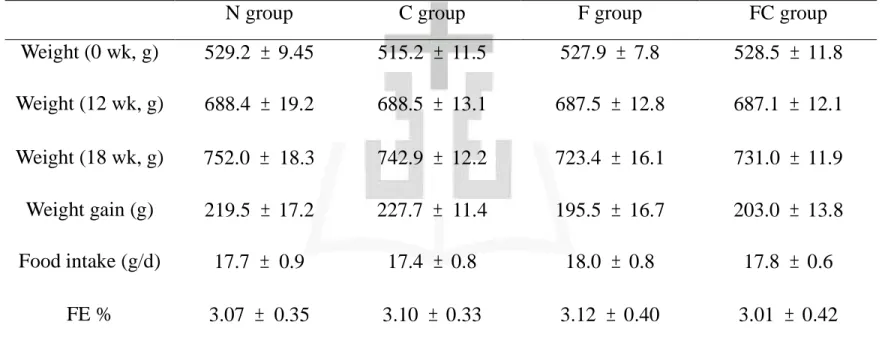
不同飼料對於大白鼠每日攝食量的影響如表三所示。結果顯示:各 組間並無統計上的差異 (p > 0.05)。 2) 體重 不同飼料對於大白鼠體重的影響如表三所示。結果顯示:各組實驗 期間的體重並無統計上的差異 (p > 0.05)。 三、口服葡萄糖耐受試驗 1) 血漿葡萄糖濃度 不同飼料對於血漿葡萄糖濃度變化的影響如圖五中所示。結果顯 示:各組血漿葡萄糖濃度皆於投予葡萄糖溶液30分鐘後達到最大 值。而各組間無顯著差異 (p > 0.05)。 2) 血漿胰島素濃度 不同飼料對於血漿胰島素濃度變化的影響如圖六所示。結果顯示; 各組血漿胰島素濃度皆於投予葡萄糖溶液後 30 分鐘達到最大值。 而各組間無顯著差異 (p > 0.05) 。 四、血液生化分析 1) 血漿三酸甘油酯濃度
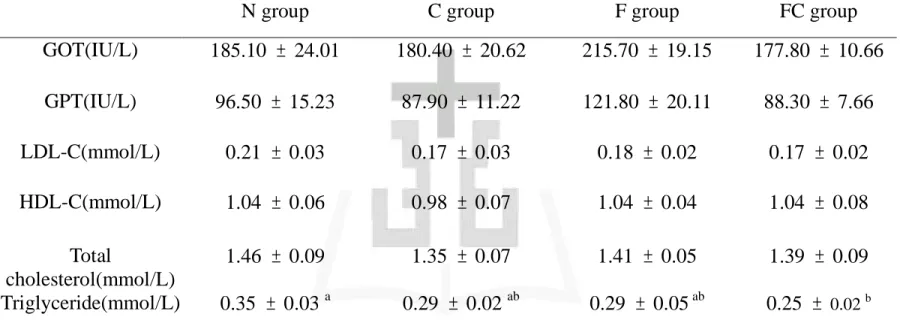
不同飼料對於血漿三酸甘油酯濃度的影響如表四所示。結果顯示:C 組、F 組、FC 組顯著低於 N 組 (p < 0.05)。 2) 血漿總膽固醇濃度 不同飼料對於血漿總膽固醇濃度的影響如表四中所示。結果顯示: 各組間並無顯著差異 (p > 0.05)。 3) 血漿高密度脂蛋白膽固醇濃度 不同飼料對於血漿高密度脂蛋白膽固醇濃度的影響如表四中所示。 結果顯示:各組間皆無顯著差異 (p > 0.05)。 4) 血漿低密度脂蛋白膽固醇濃度 不同飼料對於血漿低密度脂蛋白膽固醇濃度的影響如表四中所示。 結果顯示:各組間皆無顯著差異 (p > 0.05)。 5) 血漿游離脂肪酸濃度 不同飼料對於大白鼠血漿游離脂肪酸濃度的影響如圖八所示。結果 顯示:結果顯示,F 組和 FC 組顯著低於 N 組 (p < 0.05)。 6) 肝功能指數 不同飼料對於大白鼠肝功能指數 (GOT、GPT) 的影響如表四所 示。結果顯示:各組間皆無顯著差異 (p > 0.05)。
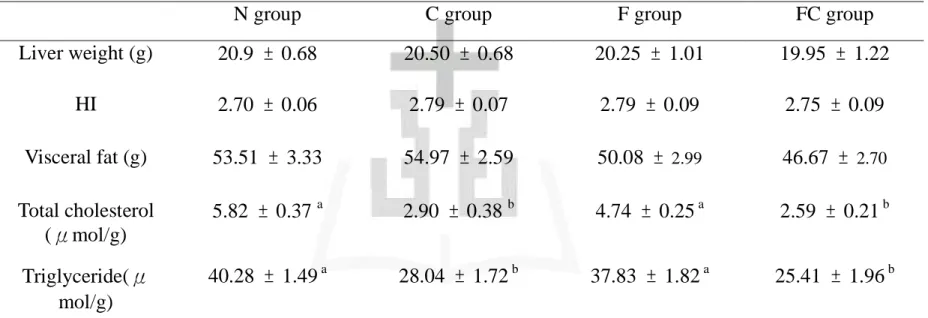
五、脂肪組織重量 1) 重量 不同飼料對於大白鼠脂肪組織重量的影響如表五所示。結果顯示: 各組間並無顯著差異 (p > 0.05)。 六、肝臟生化分析 1) 重量 不同飼料對於大白鼠肝臟重量的影響如表五所示。結果顯示:各組 間並無顯著差異 (p > 0.05)。相對肝重方面各組間並無顯著差異 (p > 0.05)。 2) 肝臟脂質濃度(膽固醇) 不同飼料對於肝臟脂質濃度的影響如表五中所示。結果顯示:C 組 與 FC 組肝臟膽固醇濃度顯著高於 N 組與 F 組 (p < 0.05)。 3) 肝臟脂質濃度(三酸甘油酯) 不同飼料對於肝臟脂質濃度的影響如表五中所示。結果顯示:C 組 與 FC 組肝臟三酸甘油酯濃度顯著高於 N 組與 F 組 (p < 0.05)。 4) 硫代巴比妥酸反應性物質 (TBARS)
不同飼料對於肝臟 TBARS 的影響結果如表七所示。結果顯示:C 組、F 組、FC 組之肝臟硫代巴比妥酸反應性物質顯著低於 N 組 (p < 0.05)。 5) 過氧化氫酶 (Catalase) 活性 不同飼料對於肝臟過氧化氫酶 (Catalase) 活性的影響如表七所示。 結果顯示: C 組、F 組、FC 組的肝臟過氧化氫酶活性顯著高於 N 組 (p < 0.05)。
6) 超過氧化物歧化酶 (Superoxide dismutase, SOD) 活性
不同飼料對於肝臟超過氧化物歧化酶 (SOD) 活性的影響如表七所 示。結果顯示:FC 組肝臟中的超過氧化物歧化酶活性顯著高於 N 組 (p < 0.05)。 7) 麩胱甘肽過氧化酶 (Glutathione peroxidase) 不同飼料對於肝臟中麩胱甘肽過氧化酶 (Glutathione peroxidase) 活 性的影響如表七。結果顯示:各組間並無顯著差異 (p > 0.05)。 8) 麩胱甘肽還原酶 (Glutathione reductase) 不同飼料對於肝臟中麩胱甘肽過氧化酶 (Glutathione peroxidase) 活 性的影響如表七。結果顯示:各組間並無顯著差異 (p > 0.05)。
不同飼料對於肝臟 CYP2E1 蛋白質含量的影響如圖十所示。結果顯 示:N 組肝中 CYP2E1 蛋白質含量顯著高於 C 組、F 組、FC 組 (p < 0.05)。 10) 肝臟 TNF-α 濃度 不同飼料對於肝中 TNF-α 濃度的影響如表八中所示。結果顯示:N 組肝中 TNF-α 濃度顯著高於 C 組、F 組、FC 組,C 組濃度顯著 高於 FC 組 (p < 0.05)。 11) 肝臟髓過氧化酶 (Myeloperoxidase) 不同飼料對於肝臟中 MPO 濃度的影響如表八中所示。結果顯示: 各組間並無顯著差異 (p > 0.05)。 12) 羥基脯胺酸 (hydroproline) 不同飼料對於肝臟中羥基脯胺酸濃度的影響如表八所示。結果顯 示:N 組肝中羥基脯胺酸濃度顯著高於 C 組、F 組、FC 組 (p < 0.05)。
七、肝臟病理分析 不同飼料對於肝臟脂肪空泡堆積與發炎情形的影響如圖七所示。經由 病理醫師判讀結果,N 組之肝臟脂肪空泡堆積程度高於其他三組。另 外,在 Massion stain 染色結果如圖十二,發現 N 組之染色結果顯著 高於 C 組 (p < 0.05);Silver stain 染色結果如圖十一,發現 N 組之染 色結果顯著高於 F 組 (p < 0.05)。 八、糞便生化分析 1) 中性固醇 不同飼料對於糞便中性固醇濃度的影響如表六所示。結果顯示:N 組、C 組、F 組糞便中性固醇濃度顯著低於 FC 組 (p < 0.05)。 2) 膽酸 不同飼料對於糞便膽酸濃度的影響如表六所示。結果顯示:C 組與 F 組糞便膽酸含量顯著低於 N 組與 FC 組 (p < 0.05)。
第四章 討論
由本次實驗結果發現,餵食實驗動物十二週 NASH 誘發飲食,導 致實驗動物,肝功能指數顯著上升、高膽固醇血症、肝臟細胞明顯脂 肪空泡堆積。除此之外,肝臟中 TNF-α 濃度顯著增加,使肝臟細胞發 炎情形增加,以及肝臟中 Hydroxyproline 濃度顯著上升等指標症狀。 本次實驗飲食主要是參考 Lieber 等研究 (Lieber et al.,2004),此研究指 出給予實驗動物攝食高油飲食,會導致實驗動物,血脂異常、肝臟脂 質堆積以及肝臟細胞發炎等症狀,與本次實驗結果相同,因此可以確 立NASH疾病動物模式的誘發。 在實驗飲食對動物生長情形的影響方面,由本實驗的結果可以得 知,不論是添加兒茶素和菊苣糖的組別與 NASH 控制組相比實驗動物 在體重增加、攝食量、攝食效應方面皆無顯著差異,顯示不論是兒茶 素或是菊苣糖的添加皆不會影響實驗動物的生長情形以及營養狀態。 胰島素抗性所引起的脂質代謝異常,會造成實驗動物肝臟中 脂質的大量堆積,導致肝臟細胞產生發炎和壞死 (Marchesini et al., 1999)。由實驗結果得知,在葡萄糖耐受試驗中兒茶素與菊苣糖的給予 並不會影響血漿中葡萄糖和胰島素濃度,所以推論飲食中兒茶素和菊
苣糖的攝取並不能影響 NASH 疾病動物的胰島素抗性。 在本實驗中我們發現飲食中兒茶素的攝取可以顯著降低 NASH 疾病模式動物血漿中三酸甘油酯濃度。Kuo 等 (2005) 在動物實驗中 也發現,在飲水中添加兒茶素,飲用三十週後,具有降低血漿中三酸 甘油酯濃度的功效。另外,在動物實驗 (Kuo et al., 2007) 中也發現兒 茶素會降低飲食中脂質在小腸內的吸收,進而降低血漿中三酸甘油酯 濃度;研究中指出綠茶中的兒茶素,特別是 EGCG 會降低食物的乳糜 化作用、消化、以及微膠粒內脂質溶解,降低了脂質在腸道的吸收作 用。除了降低脂質吸收之外,兒茶素中的 EGCG 會因其特殊的化學結 構而抑制腸道中胰脂解酶的活性,減少脂質吸收 (Nakai et al., 2005; Wang et al., 2006)。由研究推論兒茶素可能藉由減少飲食中脂質的吸 收,降低血漿中三酸甘油酯濃度,改善因 NASH 疾病所導致的血脂異 常。 實驗動物攝取 NASH 誘發飲食除了會導致血脂異常,也會顯著造 成肝臟中脂質的堆積。由本研究結果發現,攝取兒茶素可以降低肝臟 中膽固醇濃度以及因 NASH 飲食所造成的肝臟三酸甘油酯堆積。研究 指出兒茶素可能是透過抑制肝臟脂肪酸合成相關酵素活性以及降低膽 固醇合成,降低肝臟中三酸甘油酯和膽固醇堆積 (Powel et al., 1990)。
Akanji 等 (1989) 指出兒茶素具有抑制膽固醇合成酶的特殊生理活 性,進而降低膽固醇的生合成;除此之外,兒茶素還可藉由與飲食中 膽固醇的結合,而減少飲食中膽固醇吸收,進而降低膽固醇濃度。因 此,兒茶素可能可以藉由減少膽固醇的生成以及吸收來減少肝臟中膽 固醇濃度。另外,不論是細胞或是動物實驗皆證實,兒茶素會降低肝 臟中脂肪酸的合成作用 (Ikeda et al., 2005; Fiorini et al., 2005),調節肝 臟脂質濃度。Tian (2006) 的研究中證實兒茶素會降低脂肪酸合成酶活 性,其實驗結果顯示兒茶素藉由抑制脂肪酸合成酶活性,減少肝臟中 脂肪酸合成,進而減少肝臟中三酸甘油酯濃度。所以兒茶素也可能藉 由抑制脂肪酸合成酶的活性而達到降低脂肪酸的合成的作用,並進一 步減少體內和肝臟中三酸甘油酯的堆積 (Wang et al., 2005)。 除了抑制脂質生合成之外,兒茶素降低血脂和肝臟脂質堆積,也 有可能透過促進糞便中膽酸的排出、加速肝臟中膽固醇再合成膽酸的 速率、增加膽固醇代謝,所以能改善因 NASH 所造成之體內脂質代謝 異常的情況。根據研究指出,兒茶素可與飲食中膽固醇結合,而達到 抑制膽固醇的吸收,並且增加糞便中性/酸性固醇以及膽酸排出,而影 響腸肝循環,加速體內膽固醇代謝的作用,以利降低血膽固醇、促進 腸道中膽汁、膽固醇的排泄 (Yang et al., 2000; Muramatsu et al., 1989)。
由本研究結果發現,兒茶素的攝取,會增加糞便中膽固醇的排出,所 以可能藉此改善因 NASH 所造成之體內脂質異常的情況。
在 NASH 疾病進展至纖維化時,體內高濃度游離脂肪酸,會增加 體內氧化壓力,產生大量的超氧陰離子 (superide anions) 及過氧化氫 (hydrogen peroxide),造成肝臟損害或引起發炎反應,加速 NASH 疾 病進程 (Robertson et al., 2001)。由研究結果發現,兒茶素的介入會降 低肝臟內高濃度的游離脂肪酸,推論可能因此降低體內脂質過氧化作 用,進而降低氧化壓力,延緩 NASH 疾病進程。Bastie 等 (2007) 的 研究發現,若降低肥胖動物血脂及肝臟脂質堆積時,可以降低體內高 濃度的游離脂肪酸。除此之外,不論是體內或體外試驗皆證實,兒茶 素會藉由抑制脂肪酸合成酶活性,減少肝臟中脂肪酸的合成作用,降 低肝臟中游離脂肪酸濃度 (Zhang et al., 2006, Wang et al., 2003)。因此 推論,兒茶素可能藉由降低肝臟中游離脂肪酸濃度,以減少 NASH 所 造成之氧化損傷以及發炎反應。 在菊苣糖對脂質代謝的影響方面,在本研究中我們同樣發現飲食 中菊苣糖的攝取,會降低實驗動物血漿中三酸甘油酯濃度。根據動物 和臨床實驗的證實,菊苣糖可藉由降低脂質吸收,降低血漿中三酸甘 油酯濃度 (Kuo et al., 2007)。另有研究指出菊苣糖也有可能透過,大腸
內細菌的發酵作用 (Roberfroid et al., 1993),生成短鏈脂肪酸 (Guarner, 2005),使肝臟利用短鏈脂肪酸,減少游離脂肪酸的合成。由結果推論, 菊苣糖可藉由減少三酸甘油酯的吸收,以及減少游離脂肪酸的合成, 降低血漿三酸甘油酯濃度 (Roberfroid, 2005)。 除了抑制脂質生合成之外,菊苣糖改善血脂代謝異常,也有可能 透過促進糞便中膽酸的排出,加速肝臟中膽固醇再合成膽酸的速率, 增加膽固醇代謝,改善因 NASH 所造成之體內脂質異常的情況。根據 研究認為,飲食中菊苣糖的攝取,使雙叉桿菌的增生,增加腸道細菌 膽鹽水解酵素的活性,促進膽汁及膽固醇在糞便中的排出 (Tahri et al.,1996; Roos and Katan, 2000)。在本研究結果也發現,菊苣糖可以增 加糞便中膽固醇排出量,促進脂質代謝,可能藉此改善因 NASH 所造 成之體內脂質異常的情況。 除了對於血漿中脂質濃度的影響之外,本研究也發現,菊苣糖可 以降低血漿以及肝臟中游離脂肪酸濃度。根據先前研究認為,當血漿 中脂質降低時,也會減少體內游離脂肪酸濃度 (Bastie et al., 2007),降 低活性氧物質生成,減少脂質過氧化作用以及 CYP2E1 蛋白質的表現 造成氧化傷害以及發炎反應 (Robertson et al., 2001; Marchesini et al.,
力,減少肝臟細胞發炎以及損傷。 在本實驗中我們也發現,飲食中兒茶素的攝取會降低體內高濃度 的游離脂肪酸,推論可能因此降低體內氧化壓力的增加,延緩 NASH 疾病進程;在抗氧化分析結果中顯示,兒茶素確實會降低體內氧化壓 力,減少過氧化物的生成。Ji 等 (2007) 的研究中指出,當體內過氧化 物濃度低時,肝臟內抗氧化酵素的活性也會下降。在本研究中確實發 現,飲食中兒茶素的攝取,減少過氧化物生成,降低了 SOD 以及 Catalase 酵素活性。有關兒茶素的抗氧化力,主要是因為兒茶素中的 EGCG, 其結構上含有多個羥基,具提供質子的活性,能與過氧化物作用,進 而降低體內的氧化壓力。 NASH 疾病所造成的胰島素抗性,造成脂質代謝異常,體內游離 脂肪酸濃度顯著上升,而高濃度的游離脂肪酸會誘發 CYP2E1 蛋白質 的表現,產生大量的過氧化氫等活性氧,而這些活性氧屬會攻擊細胞 膜上的多元不飽和脂肪酸,引發脂質過氧化作用進行 (Robertson et al., 2001)。在本研究結果中發現,兒茶素的攝取可以顯著的降低肝臟中 CYP2E1 蛋白質的表現,因此認為,兒茶素可降低 NASH 所造成的氧 化壓力。Jimenez 等 (2004) 的研究發現,兒茶素對 CYP2E1 蛋白質所 產生的氧化壓力具有保護作用;Morillas 等 (2006)的研究也指出,茶飲
料中的多酚類具有降低氧化壓力能力。因此推論,兒茶素可能是藉由 特殊化學結構降低了氧化壓力,或者是藉由降低游離脂肪酸濃度,間 接地減少了肝臟中 CYP2E1 蛋白質的表現,進而延緩 NASH 疾病進 程。因此由研究結果可以得知,兒茶素可以降低體內氧化壓力,進而 降低肝臟細胞的氧化傷害以及發炎反應。 由脂肪酸氧化作用以及 CYP2E1 蛋白質所產生的活性物質,會引 發脂質過氧化作用,會產生脂質過氧化物,形成脂質自由基,繼續產 生氧化自由基以及氫過氧化物。自由基除了會提高氧化壓力,也會攻 擊細胞,造成細胞死亡以及肝臟發炎。當肝臟細胞發炎時,會活化星 狀 細 胞 , 促 使 前 發 炎 激 素 以 及 膠 原 蛋 白 生 成 , 導 致 細 胞 纖 維 化 (Browning and Horton, 2004);同時 TNF-α 也會大量表現,體內 TNF-α 濃度增加時會刺激相關細胞激素表現,被認為與 NASH 疾病進程,進 展至肝硬化有關。 於本研究結果發現,兒茶素顯著降低肝臟中 TNF- 濃度,嗜中性 白血球中 Myeloperoxidase 活性也有下降趨勢。根據先前的研究認為, 兒茶素降低 TNF- 的可能機制為降低體內發炎反應 (Widlansky et al., 2005),Wang 等 (2005) 更進一步指出,兒茶素是透過抑制 TNF-基 因的表現,進而向下調控 mRNA 表現,降低 TNF- 濃度,以及相關
細胞激素濃度 (Shin et al., 2007)。兒茶素除了對 TNF-的作用之外, 也有可能是透過降低血脂、肝臟中脂質堆積,進而減少游離脂肪酸濃 度,減少氧化壓力,進而改善 NASH 所引起肝細胞傷害,發炎反應, 因此降低 TNF-濃度,降低肝臟細胞發炎現象。 肝臟細胞產生發炎反應時,TNF- 以及其他相關酵素,會加重肝 細胞發炎情形,使肝細胞壞死 (Crespo et al., 2001);而當肝臟細胞發 炎、壞死,可能會導致肝細胞纖維化。本研究結果發現,補充兒茶素 可降低肝臟中 Hydroxyproline 濃度,同時肝臟切片中的膠原纖維以及 網狀纖維呈色結果亦有明顯改善,因此推論兒茶素藉由降低肝臟發炎 反應,延緩 NASH 疾病進程。 Robertson 等 (2001)指出,飲食中菊苣糖的攝取,可以藉由改善脂 質代謝異常,降低血漿以及肝臟中游離脂肪酸濃度,進而降低 CYP2E1 蛋白質的表現。由本研究結果發現,菊苣糖的攝取確實減少 CYP2E1 蛋白質的表現,減少 CYP2E1 蛋白質所產生的活性物質所引發的脂質 過氧化作用,減少脂質過氧化物,降低體內氧化壓力,進而降低 SOD 以及 Catalase 抗氧化酵素活性 (Ji et al., 2007)。菊苣糖降低脂質過氧 化作用,可能是透過降低體內脂質的過度堆積,減少脂質的過氧化作
一步減少肝細胞損害以及發炎反應,延緩 NASH 疾病進程 (Song and Yang, 2003)。 由研究結果發現,菊苣糖的攝取顯著的降低肝臟中 TNF- 濃度, 顯示菊苣糖降低了肝臟發炎反應。菊苣糖可能是透過改善脂質代謝異 常,減少游離脂肪酸濃度,降低氧化壓力,改善 NASH 所引起肝細胞 傷害,發炎反應,因此降低 TNF- 濃度,降低肝臟細胞發炎現象。 本研究結果發現,菊苣糖降低肝臟中 Hydroxyproline 濃度以及肝臟切 片中的膠原纖維以及網狀纖維呈色結果,因此推論菊苣糖可能藉由降 低肝臟發炎反應,延緩 NASH 疾病進程。
第五章 總結 飲食中兒茶素與菊苣糖的介入,兩者合併給予雖然不具有加成作 用,但是兒茶素及菊苣糖仍會藉由其特殊生理活性降低體內高濃度游 離脂肪酸濃度,進而降低體內氧化壓力,而減少肝臟的損害,減少肝 臟細胞發炎以及減少肝臟纖維化。由研究結果推論,飲食中兒茶素與 菊苣糖的介入,可以延緩 NASH 疾病進程。
參考文獻
Agheli N, Kabir M, Berni-Canani, Petitjean E, Boussairi A, Luo J, Borner F, Slama G and Rizkalla SW (1998) Plasma lipids and fatty acid synthase activity are regulated by short-chain fructo-oligosaccharides in sucrose-fed insulin-resistant rats. J Nutr 128:1283-8.
Aoeh S, Suzuki Y, Hirota T, Ahiko K and Togawa Y (1986) The effects of lactic acid bacteria and intestinal microflora on cholesterol metabolism in rats. Abstrs, 6thJapan Bifidus Center Conf. Tokyo
Bursill CA and Roach PD (2006) Modulation of cholesterol metabolism by the green tea polyphenol (-)-epigallocatechin gallate in cultured human liver (HepG2) cells. J Agric Food Chem. 54:1621-6.
Bacon BR, Farahvash MJ, Janney CG, Neusch W and Te tri BA (1994) Non-alcoholic steatohepatitis: an expanded clinical entity. Gastroentrology 107:1103-9.
Bursill C, Roach PD, Bottema CD and Pal S (2001) Green tea upregulates the low-density lipoprotein receptor through the sterol-regulated element binding Protein in HepG2 liver cells. J Agric Food Chem 49:5639-45.
Beylot M (2005) Effects of inulin-type fructans on lipid metabolism in man and in animal models.Br J Nutr 1:S163-8.
Bastie CC, Zong H, Xu J, Busa B, Judex S, Kurland IJ and Pessin JE (2007) Integrative metabolic regulation of peripheral tissue Fatty Acid oxidation by the SRC kinase family member fyn. Cell Metab 5:371-81.
Browning JD and Horton JD (2004) Molecular mediators of hepatic steatosis and liver injury. J Clin Invest 114:147-152.
Cho IJ, Lee C and Ha TY (2007) Hypolipidemic effect of soluble fiber isolated from seeds of cassia tora Linn. In rats fed a high-cholesterol diet. J Agric Food Chem 55 : 1592-6.
Dominguez-Diez A, Fernandez-Escalante JC and Pons-Romero F (2001) Gene expression of tumor necrosis favtor alpha and TNF-receptor, p55 and p75, in nonalcoholic steatoheapatitis patients. Hepatology 34 :1158-63.
Chan PT, Fong WP, Cheunq YL, Huanq Y, Ho WK and Chen ZY (1999) Jasmine green tea epicatechins are hypolipidemic in hamsters (Mesocricetus auratus) fed a high fat diet. J Nutr 129: 1094-101.
Cani PD, Dewever C and Delzenne NM (2004) Inulin-type fructans modulate gastrointestinal peptides involved in appetite regulation (glucagon-like peptide-1 and ghrelin) in rats. Br J Nutr 92:521-6. Dulloo AG, Duret C, Rohrer D, Girardier L, Mensi N, Fathi M, Chantre P
and Vandermander J (1999) Efficacy of a green tea extract rich in catechin polyphenols and caffeine in increasing 24-h energy expenditure and fat oxidation in humans. Am J Clin Nutr 72:1232-4. Dulloo AG, Seydoux J, Girardier L, Chantre P and Vandermander J (2000)
Green tea and thermogenesis: interactions between catechin-polyphenols, caffeine and sympathetic activity. Int J Obes Relat Metab Disord 24: 252-8.
Daubioul CA, Horsmans Y, Lambert P, Danse E and Delzenne NM (2005) Effects of oligofructose on glucose and lipid metabolism in patients with nonalcoholic steatohepatitis: results of a pilot study. Eur J Clin Nutr 59:723-6.
Folch J, Lees M and Sloane Stanley GH (1957) A simple method for the isolation and purification of total lipids from animal tissue. J Biol Chem 226:497-509
Fiorini RN, Donovan JL, Rodwell D, Evans Z, Chenq G, May HD, Milliken CE, Markowitz JS, Campbell C, Haines, JK, Schmidt MG and Chavin KD (2005) Short-term administration of (-)-epigallocatechin gallate reduces hepatic steatosis and protects against warm hepatic ischemia/reperfusion injury in steatotic mice. Liver Transpl 11:298-308.
Frank J, Budek A, Lundh T, Parker RS, Swanson JE, Lourenco CF, Gago B, Laranjinha J, Vessby B and Kamal-Eldin A (2006) Dietary
and protect the vitamin from oxidation in vitro. J Lipid Res 47:2718-25.
Gelman L, Feige JN and Desvergne (2007) Molecular basis of selective PPAR gamma modulation for the treatment of Type 2 diabetes. Biochim Biophys Acta 16.(in press)
Guarner F (2005) Inulin and oligofructose: impact on intestinal diseases and disorders. Br J Nutr 1:S61-5.
Grundy SM, Ahrens EH and Miettinen TA (1965) Quantitative isolation and gas-liquid chromatographic analysis of total fecal bile acid. J Lipid Res 6:397-410.
Harlander JC, Kwo PY, Cumming OW (2001) Atorvastatin for the treatment of NASH.Gastroenterology 120:539-44.
Hidaka H, Tashiro Y and Eida T (1991) Proliferation of bifidobacteria by oligosaccharides and their useful effect on human health. Bifidobacteria Microflora 10:65-79.
Hata Y, Hara T, Oikawa T, Yamamoto M, Hirose N, Nagashima T, Torihama N, Nakajima K, Watabe A and Yamashita M (1983) The effects of fructooligosaccharides against hyperlipidemia. Geriatr Med 21:156-67.
Hugo Abei (1947) Catalase in vitro. B Chance Acta Chem Scand 236:121-5.
Ikeda I, Hamamoto R, Uzu K, Imaizumi K, Naqao K, Yanaqita T, Suzuki Y, Kobayashi M and Kakuda T (2005) Dietary gallate esters of tea catechins reduce deposition of visceral fat, hepatic triacylglycerol, and activities of hepatic enzymes related to fatty acid synthesis in rats. Biosci Biotechnol Biochem 69:1049-53.
James OFW and Day CP (1998) Non-alcoholic steatohepatitis (NASH): a disease of emerging identity and importance. J Hepatol 29:495-501. Jin Y, Jin CH and Row KH (2006) Separation of catechin compounds
from different teas. Biotechnol J 1:209-13.
CYP2E1-dependent toxicity. Free Radic Biol Med 36:359-70.
Ji LL, Li FY, Luo Y, Ma XP and Chen Zl (2007) Free radicals in Carassius auratus liver: their generation and oxidative stress induced by 2,4,6-trichlorophenol. Ying Yong sheng Tai Xue Bao.18:129-32.
Kurihara H, Fukami H, Asami S, Toyoda Y, Nakai M , Shibata H and Yao XS (2004) Effects of oolong tea on plasma antioxidative capacity in mice loaded with restraint stress assessed using the oxygen radical absorbance capacity (ORAC) assay. Biol Pharm Bull 27:1093-8.
Kuo KL, Weng MS, Chiang CT, Tsai YJ, Lin-Shiau SY and Lin JK (2005) Comparative studies on the hypolipidemia and growth suppressive effects of oolong, black, pu-erh and green tea leaves in rats. J Agric Food Chem 53: 480-9.
Kondo K, Kurihara M, Miyata N, Suzuki T and Toyoda M (1999) Mechanistic Studies of Catechins as Antioxidants against Radical Oxidation. Archives of Biochemistry and Biophysics 362:79–86. Kuo SI and Noh SK (2007) Green tea as inhibitor of the intestinal
absorption of lipid: potential mechanism for its lipid-lowering effect. J Nutr Biochem 18:179-83.
Kumar KS and Malet PF (2000) Nonalcoholic steatohepatitis. Mayo Clin Proc 75:733-39.
Kaur N and Gupta AK (2002) Applications of inulin and oligofructose in health and nutrition. J Biosci 27:703-14.
Lieber CS, Leo MA, Mak KM, Xu Y, Cao Q, Ren C, Ponomarenko A and Decarli LM (2004) Model of Nonalcoholic steatohepatitis. Am J Clin Nutr 79:350-1.
Ludwig J, Viggiano TR, McGill DB and Oh BJ (1980) Nonalcoholic steatohepatitis. Mayco Clinical Proc 55:434-38.
Letteron P, Fromenty B, Terris B, Degott C and Pessayre D (1996) Acute and chronic hepatic steatosis lead to in vivo lipid peroxydation in mice. J Hepatol 24:200-208.