國 立 交 通 大 學
生物科技學院
生化工程研究所
碩士論文
探討血漿中 Haptoglobin: 生理功能及純化方法
Studies of Plasma Haptoglobin: Physiologic Significance and Purification
研究生: 岳 中 曦
指導教授: 毛仁淡 教授
誌 謝
自大二暑假就進入了毛教授的實驗室,大學畢業後繼續兩年的碩士生涯, 在這裡四年的時光,學習到除了科學上的新知外,更讓我感受到團隊合作以及與 人相處的重要性。要感謝許多人,一路上幫助我、陪伴我、扶持我,以及讓我成 長很多的人。 首先要感謝的是一路陪我走來的父母:岳成武先生及吳國玉女士,不管是在 物質以及精神層面都給我了很大的支持與幫助,當我遇到挫折時,一路給我的鼓 勵與關心,沒有他們就沒有今天傑出的我,我將這份榮耀獻給他們。接著要感謝 的是我的指導教授毛仁淡教授,這四年來,不僅在於科學上及論文上的指導,更 教導我待人接物及人生規劃的原則,平時也不忘關心我們的日常生活,就像生命 中第二的父母一樣。所謂一日為師終生為父,他給予我的諄諄教誨,我必定會謹 記在心。除此之外也要感謝張純銘教授、戴鏗碩教授、王耀宏教授在口試及論文 上的建議與指正,許多寶貴的意見讓學生獲益良多。 在實驗室要感謝財木學長、繼峰學長、文亮學長、以祥學長、明誌學長在實 驗上的教導及生活上的幫助,特別是繼峰學長自我進入實驗室後就一直不斷給予 我科學方面的教導直到我畢業。也要感謝怡芢學姊、俊瑩學長、珮真學姐、文昭 學姊、立品學姊、佩茹學姊。接著是我唯一的好同學冠佑,很高興可以跟你們一 起奮鬥,一起努力走過來。在學弟妹中,感謝為實驗室帶來一股歡笑的乃齊,還 有實驗室之花小祕書善琦,以及大學部小函、偉庭、威延、怡安以及佩宇,很高興能在與你們一起打拼,除此之外,文穎、馨儀及我的男朋友漢平獻上最深的感 謝。
這四年來,我真的成長很多,感謝上蒼給予我許多挫折以及磨鍊,讓我更成 熟懂事,謝謝你們。
探討血漿內haptoglobin:生理功能及純化方法 學生: 岳中曦 指導教授:毛仁淡 博士 國立交通大學生化工程所碩士班
摘要
人類haptoglobin 分成三種表現型:1-1,2-1 和 2-2。 先前,我們利用血紅素亦或是 單株抗體(8B1-3A)所製作之親合性管柱純化之 Hp 仍有大量 apoA-Ι 所污染。針對不純的 問題,我們發展出一套利用親合性管柱之新式純化流程。首先,我們製作及純化單株抗 體抗α 次單元(3H8)。血漿通過親合性層析管柱與抗體結合後,利用含 0.12 M NaCl 之 phosphate 緩衝液(pH 7.4)沖洗後,在使用含 0.04%十二烷基磺酸鈉緩衝液(pH 7.4)再次沖 洗結合於管柱上的最主要的雜蛋白質apoA-Ι。最後,以含 0.1%十二烷基磺酸鈉緩衝液(pH 11)將 Hp 沖提下來,並以 1M Tris-HCl 緩衝液(pH 6.8)中和。純化之 Hp 具有高純度去除 apoA-Ι 之污染,並且仍具有與血紅色結合之生化功能。純化之 Hp 三種表現型的純度皆 可到達95%以上。此新式純化方式具有高效率及高純度。其設計原理及最佳化過程皆詳 述於內文中。 猪Hp 是一個急性蛋白質,它於血漿中的濃度在發炎或感染時會急速升高。Hp 之主 要功能為負責與血漿中游離血紅素結合以抑制其氧化壓力。我們研究台灣蘭嶼迷你猪 (n=43)的 Hp 表現型,發現它們皆為同型二聚體的結構(β-α-α-β)相近於人類 Hp1-1 表現 型。利用西方墨點法及凝膠過濾管柱分析,我們發現有25%的蘭嶼迷你猪血漿中具有極 低或是沒有Hp 表現(<0.05 mg/ml)。其 cDNA 轉譯成胺基酸之序列與普通家猪(NP_999165)比較分析後發現 identity 高達 99.7%。蘭嶼迷你猪(n=43)於血漿中之平均濃度為 0.21 ± 0.25 mg/ml,明顯低於普通家猪 0.78 ± 0.45 mg/ml。另外,25%的蘭嶼迷你猪其血漿 Hp 濃度低於0.05 mg/ml。再者,具低濃度與具高濃度之迷你猪其血紅素新陳代謝速率並無 顯著差異。迷你猪其心血管系統近似於人類,因此被視為重要臨床前實驗動物,可用於 動脈硬化、心導管及心臟移植方面的試驗。然而,迷你猪是否可作為發炎相關疾病之實 驗動物仍未知。因此,此研究對於蘭嶼迷你猪作為發炎相關疾病之實驗動物提供了完整 的參考性價值。
Studies of haptoglobin: physiological significance and purification.
Student: Chung-Hsi Yueh Advisor: Dr. Simon J.T. Mao National Chiao Tung University, Institute of Biochemical engineering.
Abstract
Human haptoglobin (Hp) is classified as three phenotypes: Hp 1-1, 2-1, and 2-2. Previously, we have isolated this protein by affinity columns using either hemoglobin or monoclonal antibody (mAb) prepared against Hp β-chain (clone 8B1-3A). The isolated Hp from both methods, however, contaminates plasma apolipoprotein A-I (apoA-I). In the present report, we developed a novel affinity column procedure using a mAb prepared against α-chain of Hp (clone 3H8) for Hp purification. Plasma was first chromatographed onto the column followed by a normal wash with a buffer containing 0.12 M NaCl, 0.02 M phosphate, pH 7.4 (PBS). The bound proteins were then pre-washed with a 0.04% sodium dodecyl sulfate (SDS)-PBS, pH 7.4, to remove the low-affinity bound apoA-I from Hp. Finally, the bound Hp was eluted with a 0.1% SDS-PBS, pH 11, and collected in tubes containing 1 M Tris-HCl, pH 6.8. As a result, the isolated Hp was devoid of apoA-I and able to retain the biological function by forming an Hp-hemoglobin complex. The homogeneity of each isolated Hp 1-1, 2-1 or 2-2 was greater than 95% with a yield greater than 50%. The procedure described here is significantly improved in time consuming, recovery and purity. The rationale, design, and optimization for each step are described in detail.
Porcine haptoglobin (Hp) is an acute phase protein. Its plasma level increases significantly during inflammation and infection. One of the main functions of Hp is to bind free hemoglobin (Hb) and inhibits its oxidative activity. In the present report, we studied the Hp phenotype of Taiwanese Lanyu miniature pigs (TLY minipigs) (n=43) and found their Hp structure to be a homodimer (β-α-α-β) similar to human Hp 1-1. Interestingly, by Western blot and HPLC analysis, 25% of TLY possessed low or no plasma Hp level (<0.05 mg/ml). The Hp cDNA of those TLY was then cloned and the translated amino-acid sequence was analyzed. No sequences were found to be deficient showing a 99.7% identity with domestic pigs (NP_999165). The mean overall Hp level of TLY (0.21 ± 0.25 mg/ml) (n=43) determined by ELISA was markedly lower than that of domestics (0.78 ± 0.45 mg/ml) (p<0.001). Moreover, 25% of those TLY revealed a Hp level extremely low (<0.05 mg/ml). In addition, the Hb clearance rate did not reveal significant difference between TLY with high and extremely low level of Hp. Although TLY is a relevant animal model for pre-clinical studies such as atherosclerosis, restenosis, and heart transplantation due to the similarity of cardiovascular system between TLY and human, whether they can be used as a model for infectious diseases, using Hp as a marker has not been explored. The present study may provide a reference value for the future using TLY as an animal model for infectious disease.
Part Ι — Content
Page
Part Ι Content 5
Figure Content 7
Chapter 1 Introduction 11
Chapter 2 Materials and Methods 14
2.1. Materials 14
2.2. Preparation of ascetic fluid 14
2.3. Purification of Hp mAb 14
2.4. ELISA evaluation of dissociation of bound Hp from mAb 15
2.5. Effect of SDS on the dissociation of HDL from Hp 16
2.6. Preparation of antibody affinity column 16
2.7. Initial purification of three Hp phenotypes on immunoaffinity chromatography 17 2.8. Final affinity purification of bound Hp via pre-wash with 18
0.04% SDS-PBS, pH 7.4 2.9. Gel electrophoresis and densitometry 18
2.10. Western blot analysis 19
Chapter 3 Results 3.1. Evaluation of dissociation of bound Hp from immobilized mab 20 3.2. Effect of the concentration of SDS-PBS, pH 11, on the dissociation of bound 20
Hp from immobilized mAb
3.3. Initial purification of Hp1-1, 2-1, and 2-2 on immunoaffinity chromatography 21 3.4. Removal of apoA-I contamination 22 3.5. Final affinity purification of bound Hp via pre-wash with 0.04% SDS-PBS, 22
pH 7.4
3.6 Analysis of hemoglobin-binding property of isolated Hp 1-1, 2-1, and 2-2 23 Chapter 4 Discussion 24 Chapter 5 References 28
Figure Content
Page
Fig. 1. Schematic drawing of Hp phenotypes. 31 Fig. 2. Evaluation of dissociation of bound Hp from immobilized mAb at various 32
elution conditions.
Fig. 3. Effect of SDS concentrations on the dissociation of Hp and mAb. 33 Fig. 4. Elution profile of Hp on immunoaffinity column and contamination of apoA-I. 34 Fig. 5. Binding of HDL with Hp and its dissociation by SDS. 35
Fig. 6. Final purification of human Hp. 36
Part Ⅱ — Content
Page
Part Ⅱ Content 8
Figure content 10
Chapter 1 Introduction 39
Chapter 2 Materials and Methods 42
2.1. Materials 42
2.2. Preparation of plasma samples: 42
2.3. Phenotyping of Hp 43
2.4. Western blot 43
2.5. HPLC gel-filtration chromatography 44
2.6. Analysis of cDNA sequence of Hp from TLY minipigs 44
2.7. Determination of plasma Hp using ELISA 45
2.8. Clearance rate of Hb in TLY minipigs 46
Chapter 3 Results 3.1. Phenotyping of domestic pigs and TLY minipig 47
3.2. Evaluation of the presence of Hp in TLY minipigs by Western blot and HPLC 47 3.3. Analysis of porcine Hp mRNA expression in the leukocytes of TLY 48
3.4. Determination of plasma Hp of TLY and domestics using ELISA 48
Chapter 4 Discussion 50 Chapter 5 References 53
Figure Content
Page
Fig. 1. Schematic drawing of Hp phenotypes. 59
Fig. 2. Phenotyping of human and porcine Hp 60
Fig. 3. Western blot of porcine plasma Hp 61
Fig. 4. Typical chromatographic profiles of porcine plasma 62
Fig. 5. Expression of Hp mRNA in TLY using RT-PCR. 63 Fig. 6. Alignment of putative amino-acid sequence 64
Fig. 7. Plasma Hp concentrations in TLY and domestics. 65
Fig. 8. Population distribution (histogram) of Hp levels in TLY and domestics. 66
INTRODUCTION
Hp is known as an acute phase protein, its plasma level elevates in response to infection and inflammation. The normal concentrations of Hp in human plasma are relatively high ranging from 1.0 to 1.5 mg/ml. Clinically, it is a useful marker for some inflammatory related diseases [1-4].
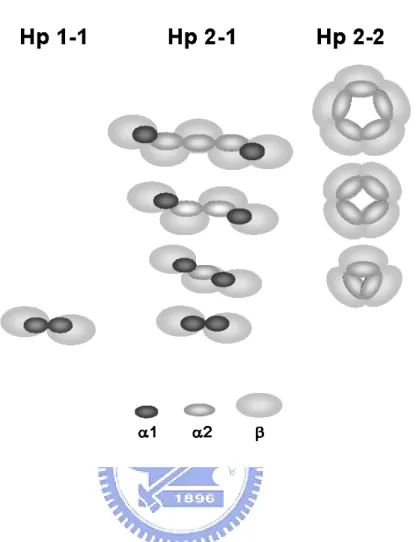
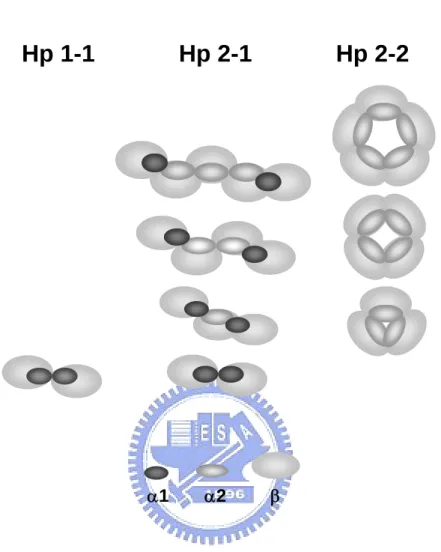
Human Hp is tetrameric arrangement joined by disulfide linkages among 2α and 2β chains (Fig. 1) [5-7]. According to the length of α chain (α1 or α2), it results in three phenotypes of Hp in the population, Hp 1-1, 2-1, and 2-2. All three phenotypes share the same 2β chains (each with about Mr 40,000 kDa including 245 amino acids containing about 30% carbohydrate in moiety), with different compositions in α chains. Homozygous Hp 1-1 or 2-2 contains identical α1 (including 83 amino acids, ~9 kDa) or α2 (including 142 amino acids, ~16.5 kDa) chains, respectively, whereas heterozygous Hp 2-1 contains each α1 and α2. Owing to an extra thio-group on the α2 chain of Hp 2-1 and 2-2 (not 1-1), they form large polymers of monomeric, trimeric, tetrameric, pentameric, hexameric, and even larger arrangement through the disulfide-linkages (Fig. 1).
The polymeric forms of Hp 2-1, 2-2 are more prone to the development of inflammatory-related cardiovascular disease than that of Hp 1-1. Presumably, this is due to the complicated structure of Hp 2-1 and 2-2 as it forms heterogeneous polymers, in which some of the biological entities are not fully expressed on the surface (Fig. 1). Recently, we
have demonstrated that Hp is an extremely potent antioxidant that directly inhibits low density lipoprotein (LDL) oxidation. The potency among the Hp phenotypes is ranked as: Hp1-1 > Hp2-1 > Hp2-2 > probucol > vitamin E [8]. Due to the difficult procedures for Hp purification, the structural and functional relationship among the Hp phenotypes has not yet been well established.
Previously, we established a simple gel-filtration purification method with high yield for porcine plasma Hp [9]. Since pigs only express one single phenotype of Hp, namely Hp 1-1, the molecular form of their Hp is simple and homogeneous. The same purification procedures are not practical for isolating human samples due to the heterogeneous nature of Hp 2-1 and 2-2 (Fig. 1). The most common procedures involved in the human Hp purification frequently suffer some drawbacks. Using hemoglobin-Sepharose affinity column, the harsh-elution condition (8 M urea) often causes dissociation of a hemoglobin subunit from the sepharose. Human plasma apoA-I of high density lipoprotein (HDL) appears to be a major contaminant [10]. Using immunoaffinity column chromatography previously established in our laboratory, human apoA-I is still coeluted with Hp. It needs another gel-filtration of high-performance liquid chromatography (HPLC) step to remove apoA-I [11]. The step is time-consuming and attenuates the yield of Hp [11]. The major focus of the present study was to eliminate the apoA-I contamination over the affinity column. We investigated the interaction between HDL and Hp and developed a new eluting buffer
system to remove the apoA-I prior to a final elution of Hp. The overall recovery of purified Hp using this procedure is greater than 50% with homogeneity about 97%.
MATERIALS and METHODS
2.1. MaterialsMonoclonal antibodies (mAb) against human Hp were produced using procedures previously described in our laboratory [12,13]. Rabbit anti-Goat IgG was purchased from Chemicon (Temecula, CA). CNBr activated Sepharose 4B was purchased from Pharmacia (Uppsala, Sweden). All other chemicals were purchased from Sigma (St. Louis, MO) without any further purification. The buffers used in this report were all filtered through a 0.45 mm filter before use.
2.2. Preparation of ascetic fluid
Adult mice were injected intraperitoneally 0.5 ml of pristane (2,6,10,14-tetramethyldecanoic acid). After 7 days, 106 hybridoma cells in 0.5 ml PBS were injected into intraperitoneal cavity of each mouse. Following another 7-14 days, the ascetic fluid was drawn by an 18-gauge needle attached to a 5-ml syringe. Each mouse produced about 5-10 ml of ascetic fluid in three batches. The collect fluid was incubated at 37℃ for 1 h and stored at 4 ℃for overnight followed by a centrifugation at 3000 g for 10 min to remove an oil layer and cell pellet.
Monoclonal antibodies prepared against human Hp were characterized according to the standard procedures established in our laboratory [12,13]. MAb 3H8 prepared against α-chain of Hp, which possessed the highest binding affinity to Hp, was selected for preparation of the affinity column. Briefly, 20 ml of ascetic fluid from the 3H8 monoclone was first precipitated by a 50% saturated ammonium-sulfate. The precipitate was dissolved in 5 ml PBS, followed by an extensive dialysis against 20 mM phosphate buffer (PB) at pH 7.2 to remove the remaining ammonium sulfate. The sample was then passed through a diethylaminoethanol (DEAE)-cellulose column equilibrated with 20 mM PB at pH 7.2, the pass-through fraction eluted with the equilibrium buffer yield about 100 mg of almost pure IgG fraction.
2.4. ELISA evaluation of dissociation of bound Hp from mAb
All the reactions including those for enzyme-linked immunoabsorbent assay (ELISA), apoA-I binding assay, preparation of affinity column, and column chromatography were conducted at room temperature, unless otherwise specified. In brief, mouse 3H8 mAb against human Hp (0.5 μg in 50 μl) was first immobilized onto a microtiter well followed by three washes to remove unbound 3H8. Each well was then blocked by 2% (w/v) dry milk in a buffer containing 0.02 M phosphate, 0.12 M NaCl, pH 7.4 (PBS). After three washes, 50 μl of Hp (5 ng/μl) in PBS were added and incubated for 1 h. Following washes to remove
the excess of Hp, the bound Hp were treated three times with various eluting reagents including a PBS, pH 11, 4 M urea-PBS, pH 11, and 0.01-2.5% SDS-PBS, pH 11. Final binding of Hp was determined using goat anti-Hp followed by a HRP conjugated anti-goat IgG. The microtiter plate was then developed with 100 μl tetramethylbenzidine (TMB) per well and stopped by adding 100 μl 1N HCl prior to read at 450 nm.
2.5. Effect of SDS on the dissociation of HDL from Hp
Human Hp was first immobilized (0.5 μg/50 μl) onto each microtiter well followed by three washes to remove unbound Hp similar to that described above. The wells were then blocked by 2% dry milk in PBS. After three washes, 50 μl of HDL (5 μg/ml) in PBS were added to each well and incubated for 1 h. After washes, the bound HDL was treated three times with SDS at various concentrations (0-0.1%) in PBS at pH 7.4. Determination of final bound HDL was conducted using goat anti-apoA-I followed by a HRP conjugated anti-goat IgG. The microtiter plate was developed with 100 μl TMB and stopped by adding 100 μl 1 N HCl prior to read at 450 nm.
2.6. Preparation of antibody affinity column
Briefly, 1 g of freeze-dried Sepharose (1 g of freeze-dried powder gave about 3.5 ml final volume of gel) was swollen and suspended in 1 mM HCl and immediately washed with 400
ml of 1 mM HCl solution within 15 min on a sintered glass filter. The gel was then washed with a coupling buffer containing 0.1 M NaHCO3 and 0.5 M NaCl, pH 8.3, and degassed.
About 15 ml (2 mg/ml) of dialyzed anti-Hp IgG in coupling buffer containing 0.1 M NaHCO3
and 0.5 M NaCl, pH 8.3 was slowly added into the gel (CNBr-activated Sepharose-4B), while gently rotating for 1 h at room temperature. After coupling, the gel was washed with 500 ml of coupling buffer to remove unbound materials via a sintered glass filter. The gel was then blocked with a buffer containing 0.1 M Tris-HCl, pH 8.0, for 2 h to saturate the remaining reactive-sites. The degassed gel was then washed with 3 cycles of 0.1 M acetate, pH 4 and 0.1 M Tris-HCl, pH 8, both containing 0.5 M NaCl. Finally, the gel was equilibrated in PBS and packed onto a 1.0 x 10 cm column with a bed volume of approximately 3.5ml.
2.7. Initial purification of three Hp phenotypes on immunoaffinity chromatography Initially, 1 ml of filtered human plasma of each Hp phenotype batch was loaded onto the affinity column (3 ml in bed volume). After which time, the column was washed with 40 ml PBS and eluted with 20-60 ml of a freshly prepared eluting buffer, either a PBS (pH 11), a 0.1% SDS-PBS (pH 11), or a 0.1% SDS-PBS (pH 7.4). Two ml of each fraction was collected in a tube containing 0.1 ml of 1 M Tris–HCl buffer, pH 6.8, to immediately neutralize the pH before a final dialysis in PBS. The pooled fraction of desired was extensively dialyzed against 4 l of PBS (with three changes) to remove SDS.
2.8. Final affinity purification of bound Hp via pre-wash with 0.04% SDS-PBS, pH 7.4 The isolation procedures were similar to that described above, except two steps involved. First, following 1 ml plasma flowed through the column and washed with 40 ml of PBS. The bound Hp was then washed again by 20 ml of 0.04% SDS–PBS, pH 7.4, to remove apoA-I that associated with Hp. Next, the bound Hp was eluted with 0.1% SDS-PBS, pH 11, as described. The fractions containing Hp were pooled and extensively dialyzed against 4 l of PBS (with three changes) to remove the SDS. Homogeneity of each Hp phenotype was then analyzed on SDS–PAGE containing 1% 2-mercaptoethanol.
2.9. Gel electrophoresis and densitometry
SDS–PAGE was performed according to Laemmli’s method with modifications by using 5% polyacrylamide (w/v) on the stacking gel as previously described [14]. Samples (typically 5 μg) for 15% SDS–PAGE were preheated at 100℃ for 10 min in a loading buffer (12mM Tris–HCl, pH 6.8, 0.4% SDS (w/v), 5% glycerol (v/v), 2.88mM of 2-mercaptoethanol, and 0.02% bromphenol blue (w/v)). The samples were run for about 1.5 h at 100V and stained using Coomassie brilliant blue R-250. Densitometric analysis of SDS–PAGE was performed using a Molecular Dynamics densitometer (Hercules, CA) for data acquisition and Image Quant software for integration and analysis.
2.10. Western blot analysis
Following the separation of proteins by SDS–PAGE, the gel was electrically transfer to a nitrocellulose-membrane attached with a 3MM filter paper pre-soaked in a transfer buffer containing 48 mM Tris–HCl, 39 mM glycine, 0.037% SDS (w/v), and 20% methanol (v/v), pH 8.3. The rest of the procedures for immunoblot was similar to that described previously [9,10].
RESULTS
In our previous studies, we have described mAb and hemoglobin affinity-column methods for the purification of human Hp [10,11]. Although these two methods, using PBS (pH11) as an eluent, were simple; the average Hp recovery was lower than 50% and an additional chromatographic step was need to remove the contaminated apoA-I. To improve the recovery and the purity of isolated Hp in the present report, the following steps described below were conducted.
3.1. Evaluation of dissociation of bound Hp from immobilized mAb
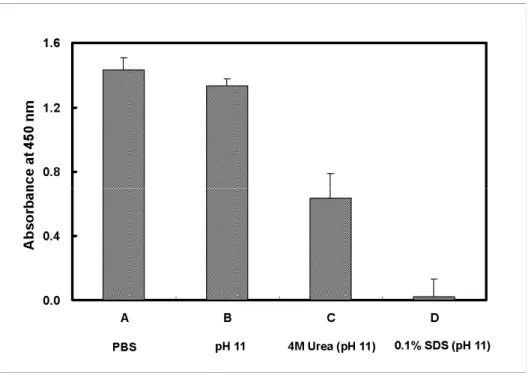
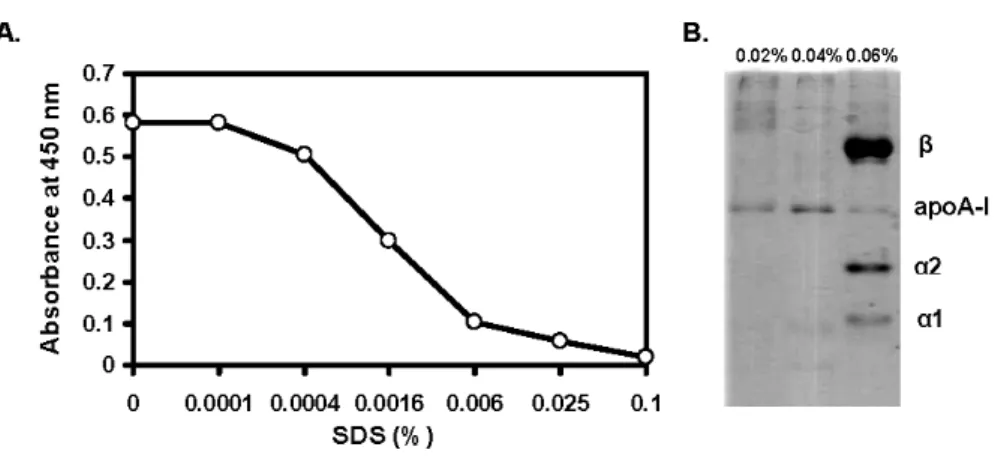
To optimize the elution conditions for the yield of Hp while isolating, we evaluated the Hp and mAb interaction on an ELISA plate to mimic the Hp binding on mAb affinity column. Various modified eluting buffers including PBS (pH 11), 4 M urea-PBS (pH 11), and 0.1% SDS-PBS (pH 11) were studied. Fig. 2 demonstrates that the dissociation of bound Hp from mAb was not completed using PBS, pH 11. However, the bound Hp was completely dissociated when using the same PBS buffer containing 0.1% SDS. Buffer containing 4M urea was only partially effective. The data suggest that SDS with pH 11 could be a useful reagent for eluting the bound Hp.
immobilized mAb
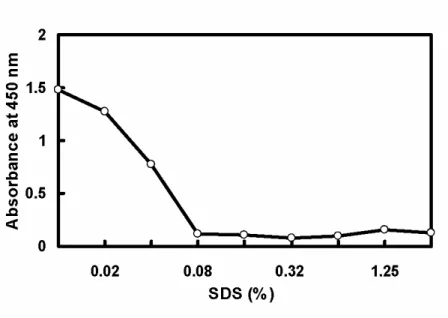
To optimize the concentration of SDS capable of dissociating Hp from immobilized mAb, serial diluted SDS was used for the assay. Fig. 3 shows that the SDS concentrations greater than 0.08% completely dissociated Hp from the mAb. Since the mAb was immobilized on the plate as a capture, it is worth mentioning that SDS at 0-2.5% did not cause the dissociation of immobilized mAb from the plate (data not shown).
3.3. Initial purification of Hp1-1, 2-1, and 2-2 on immunoaffinity chromatography
Fig. 4 shows the optimal conditions for the recovery of Hp phenotypes from the column. Overall, the Hp yield using SDS-PBS, pH 11 as an eluant was two times higher than that using PBS, pH 11 (a conventional method) or SDS-PBS, pH 7.4. Interestingly, eluting buffer of SDS with pH 7.4 delayed the Hp retention time. One of the explanations is that antibody-antigen interaction is mostly ionic dependent; SDS at neutral pH slowly alters the conformation of antigen or antibody via a protein-SDS micelle formation [15]. Further conformational change is induced at high pH resulting in the dissociation of antigen-antibody complex. Thereby, elution of Hp by SDS combined with pH 11 resulted in high recovery of Hp 1-1 (71%), 2-1 (68%), and 2-2 (85%) from the column (Table. 1). Nonetheless, the isolated Hp appeared to contaminate some apoA-I (Fig. 4D).
3.4. Removal of apoA-I contamination
ApoA-I is a major apolipoprotein residued in HDL [16]. It has been known for some time, apoA-I is a major contaminant in isolated Hp using variety of the methods [9,10]. To minimize the apoA-I contamination, we evaluated the HDL and Hp interaction on an ELISA plate immobilized with Hp. Fig. 5A reveals that apoA-I was able to bind Hp, but bound apoA-I was dissociated from Hp at SDS (pH 7.4) concentrations greater than 0.025%. In the next experiment, we mimicked the above condition directly on the affinity column. By SDS-PAGE, concentrations between 0.02 and 0.04% of SDS could wash bound apoA-I away from the column, but higher concentrations of SDS (≧0.06%) removed both apoA-I and Hp from the immobilized mAb (Fig. 5B). Likewise, 0.04% SDS-PBS (pH 11) also removed apoA-I and Hp (data not shown). The data indicate apoA-I can be selectively removed using 0.04% SDS-PBS, pH 7.4, prior to a final elution of Hp.
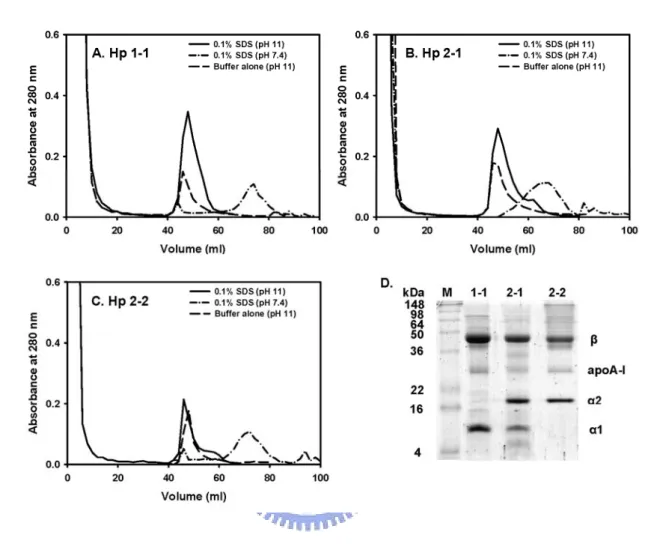
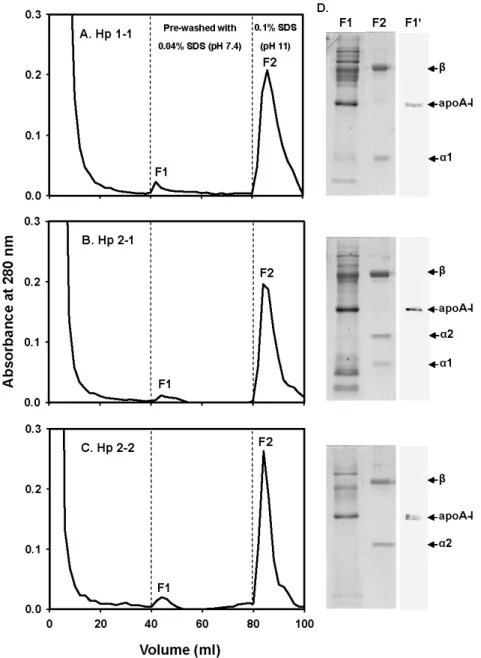
3.5. Final affinity purification of bound Hp via pre-wash with 0.04% SDS-PBS, pH 7.4 Finally, we used 0.04% SDS as a pre-wash for the removal of bound apoA-I from Hp over the affinity column. Fig. 6 shows a typical chromatographic profile for the plasma of Hp 1-1, 2-1, or 2-2. Following a flow through of plasma and washes with a PBS, pH 7.4, the column was then pre-washed with 0.04% SDS-PBS, pH 7.4 to remove bound apoA-I as well as other non-specifically bound proteins as fraction 1 (F1). Next, 0.1% SDS–PBS, pH 11
was used to elute bound Hp from mAb as fraction 2 (F2). Fig. 6 exhibits that pre-wash fraction (F1) had most of apoA-I contaminant as identified by a SDS-PAGE and confirmed by a Western blot analysis. The homogeneity of final isolated Hp was about 97% (Fig. 6, F2 fraction). The final recovery of isolated Hp 1-1, 2-1, and 2-2 was approximately of 56, 55, and 71%, respectively (Table 1).
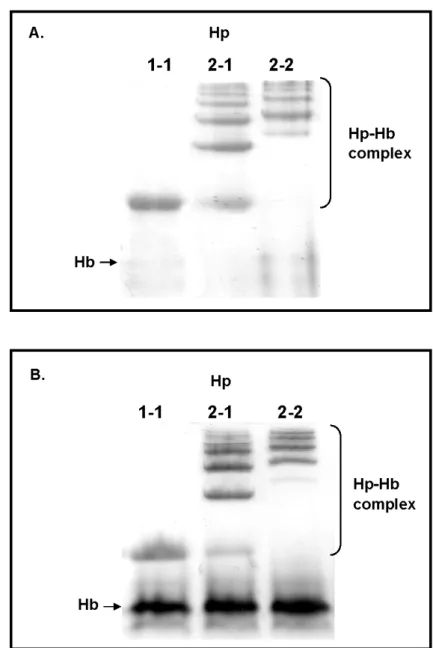
3.6. Analysis of hemoglobin-binding property of isolated Hp 1-1, 2-1, and 2-2
Fig. 7 reveals that final isolated Hp possessed the ability to form complex with Hb using a 7% native PAGE analysis. Each phenotype formed complex with Hb as a monomer (Hp 1-1) and polymers (Hp 2-1 and 2-2), while Hp 2-1 shared one completely identical monomer to Hp 1-1.
DISCUSSION
Clinically, Hp phenotypes are found to be related to several inflammatory diseases. For examples, polymeric form of Hp 2-1 or 2-2 is associated with the complications of myocardial infarction [17], kidney failure [18], and diabetics [19]. In a prospective study, human subjects with Hp 2-2 are at a 5-fold increased risk for the development of CAD as compared to those with Hp 1-1 [20]. The risk in heterozygous Hp 2-1 participates is intermediate [20]. Low levels of Hp are also found among HIV-1 seropositive patients with Hp 2-2 [21]. Due to the difficult purification procedures for each Hp phenotype, the relationship between the Hp levels and its affected diseases are rarely reported. One of the most difficulties is to isolate Hp 2-1 and 2-2 because of their heterogeneity with polymeric molecular forms (Fig. 1). On the other hand, it is essential to study the structural and functional relationship among the Hp phenotypes. Previously, Rademacher et al. [22] utilize the chicken hemoglobin–Sepharose affinity column to isolate human Hp; the harsh-elution condition (8M urea) causes the dissociation of a hemoglobin subunit from the Sepharose matrix. Meanwhile, human apoA-I appears to be another major contaminant. Travis et al. [23] employ Sephadex G-200 gel filtration, but the purified Hp is accompanied with large amounts of IgM and a-2 macroglobulin. Morimatsu et al. [24] provide a modified method using HPLC with anion-exchange, Sephacryl S-300, TSK Phenyl-5PW, and TSK DEAE-5PW columns together; the procedures however are time-consuming and the yield is relatively low.
In our previous studies, we utilized hemoglobin or Hp mAb (prepared against Hp β-chain;clone 8B1-3A) affinity column to isolate three phenotypes of Hp [10,11]. Both methods showed that there was a contamination of apoA-I, when eluting Hp by PBS, pH 11. To remove contaminated apoA-I, a re-chromatography on an HPLC Sephorose-12 gel-filtration was required [11]. For this reason, one major focus of this report was attempt to minimize the contamination of apoA-I. To test the possibility that plasma HDL (apoA-I containing lipoprotein) may directly bind to Hp, we immobilized purified Hp on an ELISA plate followed by the addition of excess amount of HDL to saturate its interaction with Hp, if any. Most interestingly, we found that HDL could bind Hp (Fig. 5), although the binding affinity between HDL and Hp was not readily known. Such binding is not non-specific, since Hp can inhibit the apoA-I-dependent lecithin:cholesterol acyl transferase (LCAT) activity in vitro [25], which plays a role in the reverse cholesterol transport [26]. The data suggest that HDL is associated with Hp in plasma, at least in part. Fortunately, the binding affinity seemed to be differentially lower than that of Hp-mAb. It was why 0.04% SDS-PBS (pH 7.4) only removed apoA-I from Hp, but not Hp from immobilized mAb (Fig. 5). But, SDS concentrations ≧ 0.06% or 0.04% SDS-PBS at pH 11 should be avoid for pre-wash as Hp could come off the column. In an early study [9], we proposed to use HDL depleted plasma (a bottom fraction obtained by ultracentrifugation at KBr d.1.21 g/ml) for Hp purification, assuming the contamination of plasma apoA-I could be eliminated. This
experiment was conducted in the present study. The isolated Hp still contained some apoA-I, but not as much as that of whole plasma (data not shown). Therefore, the use of HDL-deficient plasma for Hp purification may not be considered in the future.
SDS is used more often than any other detergent as an excellent denaturing or unfolding reagent [15,27]. It breaks mostly the quaternary and tertiary protein-protien interaction [28,29]. As such, at low SDS concentration (0.04%), pH 7.4, it dissociates the apoA-I from Hp. While at 0.1% of SDS, pH 11, it elutes bound Hp from the immobilized mAb over the affinity column. However, one concern is that SDS may alter the conformation of isolated Hp. Biswas and Das reported that α-Crystallin was able to refold to native structure after unfolding by SDS [29]. To address whether isolated Hp could refold closely to its native conformation, we monitored the structure of Hp 1-1 using a circular dichroic spectrophotometer. We found SDS-eluted Hp being slightly more disordered than that of Hp eluted without using SDS. Following extensive dialysis in 4 l PBS with three changes, the disordered structure, however, refolded to the original alpha-helical content (about 30% helix) as that values previously reported [11] (data not shown). Furthermore, we have shown that the formation of Hp-Hb complex is dependent on the overall conformation of Hp [8]. In the present study, the purified Hp following a dialysis could form complex with Hb (Fig. 7), suggesting that Hp have refolded to native form. Nevertheless, the purified Hp also retained its immunochemical property as determined by Western blot and ELISA (data not shown).
Taking together, the present method using SDS as an eluant has certain advantages. First, Hp can be purified in a predictable way by passing it through the immobilized mAb. Second, the bound apoA-I (or other non-specially bound proteins) is selectively removed prior to final elution of Hp. Third, the technique allows isolation of the polymeric form of Hp 2-1 and 2-2 (Fig. 7) unlike standard methods which may significantly lose part of them. Some standard methods which rely on different molecular masses or charges can distribute the component to different fractions. Fourth, the simple technique can be used for analytical purpose, for example, for the determination of polymeric forms of Hp that may be of important for the investigation of metabolism in pathological cases. Because SDS is used in the elution buffers, it is worth mentioning that a large volume of PBS (300 ml), pH 7.4, is required for the regeneration of the column, which is time consuming and considered to be one disadvantage of the present method.
REFERENCES
[1] M.T.M. Raijmakers, E.M. Roes, R.H.M. te Morsche, E.A.P. Steegers, W.H.M. Peters, J. Med. Genet. 40 (2003) 214.
[2] G. Engstrom, L. Stavenow, B. Hedblad, P. Lind, K.F. Eriksson, L. Janzon, F. Lindgarde, Diabetes 52 (2003) 442.
[3] S.M. Sadrzadeh, J. Bozorgmehr, Am J Clin Pathol. 121 (2004) S97
[4] M.S. Arredouani, A. Kasran, J.A. Vanoirbeek, F.G. Berger, H. Baumann, J.L. Ceuppens, Immunology. 114 (2005) 263
[5] M. Kristiansen, J.H. Graversen, C. Jacobsen, O. Sonne, H. Hoffman, S.K.A. Law, S.K. Moestrup, Nature 409 (2001) 198.
[6] N. Maeda, O. Smithies, Annu. Rev. Genet. 20 (1986) 81.
[7] D. Patzelt, G. Geserick, H. Schroder, Electrophoresis 9 (1988) 393.
[8] C.F. Tseng, C.C. Lin, H.Y. Huang, H.C. Liu, S.J.T. Mao, Proteomics 4 (2004) 2221. [9] S.J. Yang, S.J.T. Mao, J. Chromatogr. B, 731 (1999) 395.
[10] C.Y. Liau, T.M. Chang, J.P. Pan, W.L. Chen, S.J.T. Mao, J. Chromatogr. B, 790 (2003) 209.
[11] C.F. Tseng, H.Y. Huang, Y.T. Yang, S.J.T. Mao, Protein Expr. Purif. 33 (2004) 265. [12] W.L. Chen, W.T. Liu, M.C. Yang, M.T. Hwang, J.H. Tsao, S.J.T. Mao, J. Dairy Sci. 89
[13] W.L. Chen, M.T. Huang, H.C. Liu, C.W. Li, S.J.T. Mao, J. Dairy Sci. 87 (2004) 2720. [14] U.K. Laemmli, Nature 227 (1970) 680.
[15] W. Parker, P.S. Song, Biophys J. 61 (1992) 1435.
[16] S.J. Mao , J.P. Miller , A.M. Gotto Jr, J.T. Sparrow, J Biol Chem. 255 (1980) 3448. [17] M. Suleiman, D. Aronson, R. Asleh, M.R. Kapeliovich, A. Roguin, S.R. Meisel, M.
Shochat, A. Sulieman, S.A. Reisner, W. Markiewicz, H. Hammerman, R. Lotan, N.S. Levy, A.P. Levy, Diabetes. 54 (2005) 2802
[18] B. Zvi, A.P. Levy, Clin Lab. 52 (2006) 29
[19] I. Hochberg, A. Roguin, E. Nikolsky, P.V. Chanderashekhar, S. Cohen, A.P. Levy, Atherosclerosis 161 (2002) 441.
[20] R. Asleh, S. Marsh, M. Shilkrut, O. Binah, Circ. Res. 92 (2003) 1193.
[21] T.R. Zaccariotto, E.T. Rosim, D. Melo, P.M. Garcia, R.R. Munhoz , F.H. Aoki , M de Fatima Sonati. J Clin Pathol, 59 (2006) 550.
[22] B.E. Rademacher, W.J. Steele, Anal. Biochem. 160 (1987) 119. [23] J.C. Travis, B.G. Sanders, J. Exp. Zool. 180 (1972) 141.
[24] M. Morimatsu, B. Syuto, N. Shimada, T. Fujinaga, S. Yamamoyo, M. Saito, M. Naiki, J. Biol. Chem. 266 (1991) 11833.
[25] M. Balestrieri, L. Cigliano, M.L. Simone, B. Dale, P. Abrescia, Mol Reprod Dev. 59 (2001) 186.
[26] M.S. Spagnuolo, L. Cigliano, L.D. D'Andrea, C. Pedone, P. Abrescia, J Biol Chem. 280 (2005) 1193.
[27] V.I. Muronetz , T. Korpela, J Chromatogr B Analyt Technol Biomed Life Sci. 790 (2003) 53
[28] J. Bhattacharyya, K.P. Das, J. Disp. Sci. Technol, 20 (1999) 1163. [29] A. Biswas , K.P. Das, Protein J. 23 (2004) 529.
Fig. 1. Schematic drawing of molecular arrangement in huamn Hp phenotypes. Hp 1-1 possesses only the basic dimer (α1-β)2. While Hp 2-1 is comprised of many structures, a
dimer (α1-β)2, a trimer (α-β)3, and other linear polymers. Here, α represents both α1 and α2
chains. Hp 2-2 is comprised of a trimer (α2-β)3 and other cyclic polymers. Each α1, α2,
or β is 83, 142, or 245 amino acids in length, respectively. α2 is similar to α1, differing only by an additional insertion of a repeat identical to 3/4 of α1. Due to the extra Cys-74 in α2, Hp 2-1 and 2-2 form complicated polymers.
Fig. 2. Evaluation of dissociation of bound Hp from immobilized mAb at various elution conditions. To determine a simple and optional condition that is able to elute the Hp from immobilized mAb, Hp mAb was first immobilized on an ELISA plate followed by incubating 50 μg of purified Hp 2-1 in 50 μl PBS. After washing, bound Hp was then treated with various eluting buffers. (A) Total binding of Hp without eluting reagents; (B) PBS, pH 11; (C) 4 M urea-PBS, pH 11; (D) 0.1% SDS-PBS, pH 11. Binding of Hp was determined using goat anti-Hp followed by HRP conjugated anti-goat IgG for final development of chromogenicity. Each value represents a mean of triplicates ± SD.
Fig. 3. Effect of SDS concentrations (pH 11) on the dissociation of bound Hp from immobilized mAb. The remaining Hp was determined similar to that described in Fig. 2. Each point represents a mean of triplicates ± SD. Typical deviation is shown in the first three doses.
Fig. 4. Elution profile of bound Hp from immobilized mAb on immunoaffinity column (A-C) and contamination of apoA-I in isolated Hp (D). (A-C) One ml of human plasma was applied onto the immunoaffinity column and washed with a PBS, pH 7.4. The bound Hp was subsequently eluted with a 0.1% SDS-PBS, pH 11 and with a 0.1% SDS-PBS, pH 7.4. The overall data suggests that the eluting buffer containing 0.1% SDS at pH 11 results in high yield for Hp purification. (D) Analysis of isolated Hp by a 15% SDS-PAGE containing 1% 2-mercaptoethanol. It reveals that isolated Hp contaminates apoA-I.
Fig. 5. Binding of HDL with Hp and its dissociation by SDS. (A) Hp was first immobilized on an ELISA plate followed by incubating 50 μg of purified HDL in 50 μl PBS. After washing with PBS, the bound HDL dissociated from Hp by treating with SDS in a dose-dependent fasion. Binding levels of HDL were determined using goat anti-apoA-I followed by HRP conjugated anti-goat IgG similar to that described in Fig. 2. The data suggests that Hp associated apoA-I can be eliminated in the presence of SDS. (B) Eluant from affinity column, eluted with 0.02%, 0.04% and 0.06% SDS-PBS, pH 7.4, was analyzed by SDS-PAGE in the presence of reducing reagent. It suggests to avoid of using 0.06% SDS for apoA-I removal.
Fig. 6. Final purification of human Hp 1-1 (A), 2-1 (B), and 2-2 (C) from immunoaffinity column pre-washed with 0.04% SDS-PBS, pH 7.4. (A-C) Initially, 1 ml of human plasma was passed through the column with PBS, pH 7.4 and washed with the same buffer. The bound Hp was then pre-washed with 40 ml of 0.04% SDS-PBS, pH 7.4 to remove Hp associated apoA-I (F1). Finally, Hp was eluted from immobilized mAb by 0.1% SDS-PBS, pH 11 (F2). (D) Analysis of F1 and F2 on 15% SDS-PAGE containing 1% 2-mercaptoethanol and F1 on Western blot (F1'). The α and β subunits corresponding to each Hp phenotype are shown.
Fig. 7. Analysis of hemoglobin-binding property of isolated Hp 1-1, 2-1, and 2-2 (A) and Hp 1-1, 2-1, and 2-2 from human plasma (B) on 7% native-PAGE. Briefly, each isolated Hp (5 μg) or each plasma (6 μl) was incubated with hemoglobin (Hb) (5 μg) at room temperature for 30 min. Following electrophoresis, the gel was stained directly by a freshly prepared DAB solution containing 0.05% H2O2 as a developer for the endogenous peroxidase
INTRODUCTION
Porcine Hp is an acute phase protein with a molecular weight approximate 120 kDa [1-3]. It possesses an electrophoretic mobility and quaternary structure similar to human Hp 1-1 being a homodimer (αβ)2 that is linked by disulfide bridges [3, 4]. Due to its homogeneity,
porcine Hp can be purified by a one-step HPLC gel-filtration chromatography established in our laboratory [5].
Functions of Hp have been proposed comprehensively; one of the major functions is to scavenge and complex with plasma free hemoglobin (Hb) that possesses an oxidative toxicity via iron-containing hemes [6, 7]. The Hp-Hb complex is then metabolized through a cycteine-rich receptor (CD163) on macrophage [8]. It has the bacteriostasis ability by attenuating iron necessary for bacteria growth [9, 10]. Hp has also been proved to be an extremely potent antioxidant which directly inhibits Cu(Ⅱ) induced low density lipoprotein oxidation and reduces cell oxidative stress [11].
The plasma concentration of porcine Hp increases significantly during infection, inflammation, and tissue damage [12, 13]. Pigs with lameness, tail biting or diarrhea show a high level of Hp [14]. Natural infections of pigs with porcine reproductive and respiratory syndrome virus (PRRS) [15], or with Actinobacillus pleuropneumoniae [13], Mycoplasma hyorhinis [16] and Toxoplasma gondii [17] bacteria resulted in a significant elevation of their Hp levels. The protein level has therefore been used to identify both clinical and subclinical
diseases [1, 18] as well as to monitor the health status in pig production [14].
Human plasma Hp is classified as three phenotypes: 1-1, 2-1, and 2-2 (Fig. 1) [11, 19]. Hp 1-1 is a molecule of homodimer or (αβ)2, whereas 2-1 is comprised of multiple forms
including homodimer, trimer, tetramer, and other linear polymers. Hp 2-2, on the other hand, consists of trimer, tetramer, and other cyclic polymers. In non-human mammalians, both dimeric and polymeric forms of Hp exist. Thus far, polymeric forms analogous to human polymeric Hp are found only in ruminant families of Artiodactyla order [20]. TLY minipigs classified as Sus scrofa are aborigine in Taiwan with ears smaller than the other breeds. The body weight of TLY is usually lower than 25 kg during the first six months of age as compared with 80 kg of other domestic pigs [21]. Due to its low body weight and conserved population, TLY minipigs have been used as an experimental animal [21]. In the present study, we phenotyped the Hp of TLY minipigs. While phenotyping, we noticed that many of them possessed very low plasma levels of Hp. The cDNA nucleotide sequence of TLY Hp was also conducted to study whether there was a gene defect. It is of interest to observe that there is no direct relationship between the putative amino-acid sequence of Hp and its levels. Their amino-acid sequences are almost completely identical to that of domestic pigs (except Val-65ÆLeu-65). Finally, a noncompetitive ELISA was employed to determine their plasma Hp levels. In average, the mean Hp levels of TLY (0.21 ± 0.25 mg/ml) were significantly lower (4X) than that of domestics (0.78 ± 0.45 mg/ml) (p<0.001). Among
which, 25% of TLY revealed a level lower than 0.05 mg/ml. The present study may provide a reference value for the future use of TLY as an animal model for inflammatory, cardiovascular, and infectious disease studies.
MATERIALS and METHODS
2.1. MaterialsPurified porcine Hb and rat polyclonal antibodies against porcine Hp were prepared according to the methods previously described [5, 22]. Goat anti-rat IgG was purchased from Chemicon (Temecula, CA). Sepharose-12 HR column was obtained from Pharmacia (Uppsala, Sweden). Total RNA extraction kit was purchased from Geneaid (Taipei, Taiwan). All other chemicals were purchased from Sigma-Aldrich (St. Louis, MO) without any further purification. The buffers used in this report were all filtered through a 0.45-μm filter before use (Millipore, MA, USA).
2.2. Preparation of plasma samples
Forty-three TLY minipigs (23 sows and 20 boars) aged from 1 to 2 years were raised in Taitung Animal Propagation. Thirty-five domestics (12 Durac, 15 Landrace, and 8 Yorkshire) aged 6-8 months used in this study were from Animal Technology Institute, Taiwan. All of these pigs were reared in a natural lighting environment. Management and medical treatment were conducted according to the instruction established by National Science Council of Taiwan. Pigs were vaccinated against swineenzooticpneumonia, hog cholera, pseudorabies, atrophic rhinitis, foot and mouth disease, and Actinobacilluspleuropneumoniae. All selected animals were free of adverse signs of disease after a clinical examination in the
herd. Blood samples were collected via jugular puncture using a 25-gauge needle and added into the tubes containing 0.1% ethylenediaminetetraacetic acid (EDTA). Plasma was obtained by a centrifugation at 2,000 g for 20 min and stored at -20℃ until analysis.
2.3. Phenotyping of Hp
A 7% native-polyacrylamide gel electrophoresis (native-PAGE) was conducted according to the Laemmli’s method with the use of 5% polyacrylamide (w/v) as a stacking gel [23]. Eight μl of plasma was incubated with 16 μg of porcine or human Hb for 30 min at room temperature and subsequently mixed with a loading buffer [12 mM Tris–HCl, pH 6.8, 5% glycerol (v/v), 2.88 mM of 2-mercaptoethanol, and 0.02% bromphenol blue (w/v)] in a final volume of 15 μl. Following the electrophoresis, the gel was briefly washed and then immersed in freshly prepared 3,3’-diaminobendidine (DAB) solution (0.125 g of DAB dissolved in 0.5 ml DMSO with the addition of 30 ml deionized H2O containing 0.05% H2O2)
with gently shaking to develop the pattern of Hp-Hb complex.
2.4. Western blot
Following the separation of proteins by SDS–PAGE, the gel was electrically transferred to a nitrocellulose-membrane attached with a 3MM filter paper presoaked in a transfer buffer containing 48 mMTris–HCl, 39 mM glycine, 0.037% SDS (w/v) and 20% methanol (v/v), pH
8.3. The rest of the procedures for Western blot were similar to that described previously [5, 24]. Rat polyclonal antibody against β chain of porcine Hp was used for the determination of presence of Hp.
2.5. HPLC gel-filtration chromatography
Plasma of 3 individual pigs (1 Yorkshire and 2 TLY minipigs with normal and low Hp level) was used for the analysis of Hp elution profile. Each plasma was fractionated via a 50% saturated ammonium sulfate. After gentle stirring for 30 min at room temperature, the mixture was centrifuged at 10,000 g for 15 min at 4 . The supernatant was subsequently ℃ dialyzed against 4 l of PBS at 4 overnight with three changes. The sample was then ℃ chromatographed onto a Superose-12 HR column (30 x 1.0 cm) pre-equilibrated with PBS and conducted by a Waters 600 HPLC controller system (Millipore, MA, USA). The flow rate was 0.5 ml/min using PBS as a mobile phase.
2.6. Analysis of cDNA sequence of Hp from TLY minipigs
Total RNA was isolated from whole blood containing leukocytes using a Total RNA Mini Kit according to the manufacture’s instructions. An oligo(dT) 18mer was used for the reverse transcription step. The first strand cDNA was synthesized with M-MLV reverse transcriptase at 37°C for 50 min. A gene fragment coding for porcine Hp was amplified by
PCR using proofreading DNA-polymerase and oligonucleotide primers as previously described [25]. The primer design was based on the published nucleotide sequence of porcine Hp (NM_214000). The cDNA primers prepared were 5’-CGCGC CCTGG GAGCT GT-3’ (forward) and 5’-CAGTT GGCAG CTATG GTTGT CTGAA CCA-3’ (reverse). The PCR product was then cloned and transformed into Escherichia coli JM109 via TA-cloning kit (Invitrogen, Carlsbad, CA). After sequencing, the putative amino-acid sequence alignment between TLY minipig and NP_999165 was conducted. The identity/divergence percentage was analyzed by DNAstar software (DNASTAR, Madison, WI).
2.7. Determination of plasma Hp using ELISA
Porcine Hp concentration was determined by Pig Haptoglobin ELISA kit (Immunology Consultants Laboratory, OR, USA) according to the manufacture’s instructions. In brief, plasma samples were first diluted to 1:10,000 with a sample dilution buffer. The diluted samples of 100 μl or calibrator were then added to each microtiter well containing immobilized anti-Hp IgG followed by an incubation for 30 min at 24℃. After thorough washes, 100 μl of HRP-conjugated anti-porcine Hp IgG were added and incubated for 15 min. Finally, the plate was washed for three times and developed in 100 μl of 3, 3’, 5, 5’-tetramethylbenzidine (TMB) substrate for 15 min. The reaction was stopped by adding 100 μl of stop solution and read at 450 nm.
2.8. Clearance rate of Hb in TLY minipigs
Porcine Hb was purified using a procedure as previously described [24]. Sixty mg fluorescein isothiocyanate (FITC) dissolved in 120 μl dimethyl sulfoxide was gently mixed with 1 g of porcine Hb in 30 ml of NaHCO3 buffer (pH 9.0) (60 μg FITC per mg Hb) at 37℃
for 1 h. The reaction mixture was then dialyzed against 4 l of PBS at 4℃ overnight (in dark) with three changes. Before use, the dialyzed FITC-Hb was sterilized by filtrating through a 0.22-μm filter. The FITC-Hb was then infused into 3 TLY minipigs with low Hp level (0.043, 0.0064, 0.091 mg/ml) and 3 TLY with high level (~0.4 mg/ml) through ear vein at a dosage of 1.5 mg of Hb per kg body weight. For each TLY, approximately 2 ml of blood were collected into a vacuum tube containing 0.1% EDTA by venipuncture from the jugular vein at the time point of 5, 10, 20, 40, 60, 120, 240, 360 min. Because the fluorescence intensity was significantly influenced by pH, 800 μl of tested plasma samples were buffered to pH 7.4 by the addition 200 μl of 500 mM HEPES [26, 27]. The FITC-Hb standards ranged from 0 to 0.02 mg/ml were prepared by spiking desired amount of FITC-Hb into the plasma. Eventually, the intensity of fluorescence was measured at 485-nm excitation and read at 538-nm emission using a fluoresence spectrophotometer (HITACHI F-7000, Japan) to determine the levels of FITC-Hb remained.
RESULTS
3.1. Phenotyping of domestic pigs and TLY minipigWe identified their Hp phenotype based on the formation of Hp-Hb complex on a 7% native-PAGE. A typical example for the phenotyping among the domestics (n=35) and TLY (n=43) is shown in Fig. 2. Phenotype of each human Hp 2-1 or 2-2 was clearly demonstrated by the presence of polymeric forms. We were not able to observe any Hp polymers in a total of 78 pigs screened. Interestingly, we found some TLY whose Hp 1-1 levels were considerably lower than that of domestic pigs based on the Hp-Hb complex formation (Fig. 2). Hypothetically, some TLY minipigs might be of deficient Hp or be associated with structural alteration of Hp.
3.2. Evaluation of the presence of Hp in TLY minipigs by Western blot and HPLC To further determine the plasma Hp levels in those TLY minipigs, we analyzed their Hp content on a Western blot using a rat polyclonal antibody against porcine Hp β chain [5]. Approximately, 25% of TLY showed little or no Hp as compared to domestic pigs. A typical example of the Western blot is shown in Fig 3. To rule out a possible Hp structural change may attenuate its immunoreactiviety, chromatography using established gel-filtration (HPLC) method [5] was conducted to identify the presence of plasma Hp. Fig. 4 shows a typical chromatographic profile from a pig with “Western-blot negative” plasma, in which we
were unable to find the peak corresponding to Hp as compared to that normal TLY and domestics. Thus, the “absence” of Hp in some TLY was confirmed.
3.3. Analysis of porcine Hp mRNA expression in the leukocytes of TLY
Next, we cloned the cDNA of Hp from TLY who possessed low or no levels of plasma Hp. The reasons for this experiment were two folds. First, we attempted to address whether their Hp gene was genetically defect. Second, we sought for their putative amino-acid sequence and to compare with that of domestics. Using RT-PCR for Hp αβ, we demonstrated that those TLY with extremely low or no Hp levels (n=4) were able to express Hp mRNA in their leukocytes (Fig. 5). A typical example of putative amino-acid sequence of TLY deducted from nucleotide sequence of αβ chain was shown in Fig. 6. Alignment of the amino-acid sequence of TLY with that of domestic pig from Genbank of NCBI # NP_999165 shows that there is only one substitution at N-terminus (Val-65ÆLeu-65). It reveals that the divergence between TLY and domestics is less than 0.6% (Table 1).
3.4. Determination of plasma Hp of TLY and domestics using ELISA
Because TLY were able to express Hp mRNA and their sequences were almost identical within the minipigs, it suggests that the little or no Hp levels found in some minipigs were not due to the gene deficient. Hp concentrations were then measured using a noncompetitive
ELISA. The plasma Hp levels (mean ± SD) of TLY (n=43) and domestics (Durac, Landrace, and Yorkshire) (n=35) were 0.21 ± 0.25 and 0.78 ± 0.45 mg/ml, respectively. The levels of Durac (n=12), Landrace (n=15), and Yorkshire (n=8) were 0.60 ± 0.18, 0.78 ± 0.51, 1.05 ± 0.55 mg/ml, respectively (Fig. 7 and Table 2). The population distribution (histogram) of TLY and domestics according to Hp levels was depicted in Fig. 8. In TLY, 40% of their Hp values were lower than 0.1 mg/ml (0.046 ± 0.030 mg/ml), while 25% of them were lower than 0.05 mg/ml (0.026 ± 0.02 mg/ml).
3.5. Clearance rate of Hb in TLY minipigs
To determine the possible effect of low Hp level on the clearance rate of Hb, minipigs with normal (n=3) and low Hp level (n=3) were infused with FITC-labeled Hb (FITC-Hb). Prior to this experiment, the linear relationship between the concentration of FITC-Hb and intensity of fluorescence was established with a regression coefficient 0.98 (data not shown). The clearance rate of infused FITC-Hb was then determined over time by fluorescent intensity transcribed to the Hb concentration using the standard curve. In general, there was no significant difference in clearance rate of FITC-Hb between the two groups (p>0.1) within the tested period of time (Fig. 9) [28].
DISCUSSION
Porcine Hp consists of –SH covalently linked α and β chains similar to human Hp 1-1 phenotype as (αβ)2. In the absence of reducing reagent it shows a major band at 120 kDa on
the SDS-PAGE [1-3]. Interestingly, the αβ of dog, cat, and bear are joined by a non-covalent interaction [29-31]. In the present study, we analyzed the Hp phenotype of several species of porcine Hp including TLY and domestics (Durac, Landrace, and Yorkshire); all of them belong to Hp 1-1.
In newborn domestic piglets, the plasma Hp levels increased dramatically after weaning by 2-folds [32]. The increase is thought to be associated with pathogen induced infection. Infections of PRRS virus [15], Actinobacillus pleuropneumoniae [13], Mycoplasma hyorhinis [16], and Toxoplasma gondii [17] in adults also significantly increased Hp concentrations, but depending on the species of a pathogen. For example, Hp concentration may reach to 15-folds when infected with Streptococcus suis [33]. In general, for a conventional herd, the average plasma Hp level was approximately 0.9 mg/ml and for a SPF herd it was 0.3 mg/ml [33]. In the present study, 43 TLY from a no-declared SPF herd were all clinically healthy, but their plasma Hp levels (0.21 ± 0.25 mg/ml) were approximately 4-folds lower than that of domestics (0.78 ± 0.45 mg/ml) (p<0.001). About 25% of the TLY whose Hp levels were extremely low (<0.05 mg/ml). In human, there is an inverse relationship with the severity of hemolysis. Hypo- or anhaptoglobinemia are found in malaria or anemia patients associated
with hemolysis. However, we did not observe TLY herd being suffered from any hemolysis. It has been reported that some patients with hypohaptoglobinemia or anhaptoglobinemia (Hp 0 phenotype) are of genetic origin [34-36]. A silent allele Hp0 has been identified as a complete Hp gene deletion from the upstream of the promoter region of Hp gene [37, 38]. However, the Hp mRNA expressed in TLY with “hypohatoglobinemia” are similar to other normal minipigs (Table 1.) indicating that the low Hp levels of TLY are not associated with the Hp gene deficiency. In healthy cattle, the mean concentrations of plasma Hp is about 0.084 mg/ml [33], the low plasma Hp levels found in our minipigs may mimick to the characteristics of cattle.
While the mechanism involved in the low levels of Hp in TLY is not readily known, it should be mentioned that these minipigs were first found in Lanyu Island located at a separate entity near southeast part of Taiwan. In 1980, 4 boars and 16 sows were imported to Taiwan where 20 TLY were randomly mated and propagated. The coat of some offsprings appeared few white spots on solid black coat namely spotty Lanyu minipigs. Following the isolation of spotty minipigs, some piglets have some white coat in the offsprings. These offsprings are now propagated and used as an animal model in our laboratories [21]. Hypothetically, the large number of “hypohaptoglobinemia” in TLY minipigs might be a consequence of the close inbreeding.
macrophages or liver parenchymal cells to attenuate undesired glomerular filtration of Hb. Interestingly, the clearance rate of FITC-Hb in these pigs was almost the same as those TLY minipigs with high Hp level. The data suggests that a limited level of Hp might be sufficient for normal catabolism Hp. On the other hand, a report has shown that Hp knockout (-/-) mice were able to clear Hb by altered by-pass pathways utilizing liver, spleen, and kidney [39]. However during the severe intravascular hemolysis, Hp deficient mice suffered serious renal damage resulting in poor recovery and regeneration for renal tissue [28, 40].
TLY minipigs are originally characterized as dark black with a heavy coat, and their ears are smaller than those of most domestic breeds. They weigh about 25 kg when they are sexually matured as compared with 100 kg of other domestic pigs. The morphologic characteristics of TLY minipigs are distinct from other breeds, however, the amino-acid sequence of Hp of TLY is not divergent from those domestic species. There is only one amino-acid residue consistently converted (Val-65ÆLeu-65) with the identity of 99.7% and similarity of 100% as compared to that of domestics. Due to the similarity of cardiovascular system between TLY and human [21], it is conceivable that TLY can be a relevant animal model for pre-clinical studies such as atherosclerosis, restenosis, and heart transplantation. On the other hand, whether they can be used as a model for infectious diseases, using Hp as a marker, has not been explored. The present study showing the Hp levels of TLY may provide a reference value for the future use as an animal model for inflammatory diseases.
REFERENCES
[1] Heegaard, P.M.H., Klausen, J., Nielsen, J.P., González-Ramón, N., Piñeiro, M., Lampreave, F. and Alava, M.A. 1998. The porcine acute phase response to infection with Actinobacillus pleuropneumoniae. Haptoglobin, C-reactive protein, major acute phase protein and serum amyloid A protein are sensitive indicators of infection. Comp. Biochem. Physiol. B, Biochem. Mol. Biol. 119: 365–373.
[2] Lockhart, W.L., Chung, W.P. and Smith, D.B. 1972. Studies on the dissociation of porcine haptoglobin. Can. J. Biochem. 50: 775-781.
[3] Shim, B.S., Yoon, C.S., Oh, S.K., Lee, T.H. and Kang, Y.S. 1971. Studies on swine and canine serum haptoglobins. Biochem. Biophys. Acta. 243: 126-136.
[4] Hiss, S., Deszczka, S.K., Regula, G., Hennies, M., Gymnich, S., Peterson, B. and Sauerwein, H. 2003. Development of an enzyme immunoassay for the determination of porcine haptoglobin in various body fluids: Testing the significance of meat juice measurements for quality monitoring programs. Vet. Immunol. Immunopathol. 96: 73-82. [5] Yang, S.J. and Mao, S.J.T. 1999. A simple HPLC purification procedure for porcine
plasma haptoglobin. J. Chromatagr. B. 731: 395-402.
[6] Engström, G., Hedblad, B., Stavenow, L., Lind, P., Janzon, L. and Lindgärde, F. 2003. Inflammation-sensitive plasma proteins are associated with future weight gain. Diabetes 52: 2097–2101.
[7] Raijmakers, M.T.M., Roes, E.M., te Morsche, R.H.M., te Steegers, E.A.P. and Peters, W.H.M. 2003. Haptoglobin and its association with the HELLP syndrome. J. Med. Genet. 40: 214–216.
[8] Kristiansen, M., Graversen, J.H., Jacobsen, C., Sonne, O., Hoffman, H.J., Law, S.K. and Moestrup, S.K. 2001. Identification of the haemoglobin scavenger receptor. Nature 409: 198–201.
[9] Bullen, J.J. 1981. The significance of iron in infection. Rev. Infect. Dis. 3: 1127-1138. [10] Eaton, J.W., Brandt, P., Mahoney, J.R. and Lee J.T. 1982. Haptoglobin: A natural
bacteriostat. Science 215: 691-693.
[11] Tseng, C.F., Lin, C.C., Huang, H.Y., Liu, H.C. and Mao, S.J.T. 2004. Antioxidant role of human haptoglobin. Proteomics 4: 2221-2228.
[12] Eckersall, P.D., Saini, P.K. and McComb, C. 1996. The acute phase response of acid soluble glycoprotein, α-1-acid glycoprotein, ceruloplasmin, haptoglobin and C-reactive protein, in the pig. Vet. Immunol. Immunopathol. 51: 377-385.
[13] Hall, W.F., Eurell, T.E., Hansen, R.D. and Herr, L.G. 1992. Serum haptoglobin concentration in swine naturally or experimentally infected with Actinobacillus pleuropneumoniae. J. Am. Vet. Med. Assoc. 201: 1730-1733.
[14] Petersen, B., Lipperheide, C., Knura-Deszczka, S. and Ponsgen-Schmidt, E. 2001. Haptoglobin- a screening parameter for encompassing health management of the meat
production chain. Fleischwirtschaft Int. 1: 12-15.
[15] Asai, T., Mori, M., Okada, M., Uruno, K., Yazawa, S. and Shibata, I. 1999. Elevated serum haptoglobin in pigs infected with porcine reproductive and respiratory syndrome virus. Vet. Immunol. Immunopathol. 70: 143-148.
[16] Magnusson, U., Wilkie, B., Artursson, K. and Mallard, B. 1999. Interferon-alpha and haptoglobin inpigs selectively bred for high and low immune response and infected with Mycoplasma hyorhinitis. Vet. Immunol. Immunopathol. 68: 131-137.
[17] Jungersen, G., Jensen, L., Riber, U., Heegaard, P.M.H., Petersen, E., Poulsen, J.S., Bille-Hansen, V. and Lind, P. 1999. Pathogenicity of selected Toxoplasma gondii isolates in young pigs. Int. J. Parasitol. 29: 1307-1319.
[18] Harding, J.C., Baarsch, M.J. and Murtaugh, M.P. 1997. Association of tumor necrosis factor and acute phase reactant changes with post arrival disease in swine. Zentralbl. Veterinarmed. B. 44: 405-413.
[19] Yueh, S.C.H., Lai, Y.A., Chen, W.L., Hsu, H.H. and Mao, S.J.T. 2007. An improved method for haptoglobin 1-1, 2-1, and 2-2 purification using monoclonal antibody affinity chromatography in the presence of sodium dodecyl sulfate. J. Chromatagr. B. 845: 210-217.
[20] Katnik, I., Pupek, M. and Stefaniak, T. 1998. Cross reactivities among some mammalian haptoglobins studied by a monoclonal antibody. Comp. Biochem. Physiol. B, Biochem.
Mol. Biol. 119: 335-340.
[21] Lin, J.H., Chu, R.M., Yang, P.C., Weng, C.N., Lin, P.H., Liu, S.K. and Mao, S.J.T. 1999. Influence of age on the electrocardiographic waves in taiwanese Lan-Yu miniature pigs. Contemp Top Lab Anim Sci. 38: 36-41.
[22] Mao, S.J.T., Kazmar, R.E., Silverfield, J.C., Alley, M.C., Kluge, K. and Fathman, C.G. 1982. Immunochemical properties of human low density lipoproteins as explored by monoclonal antibodies. Binding characteristics distinct from those of conventional serum antibodies. Biochim. Biophys. Acta. 713: 365-374.
[23] Laemmli, U.K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227: 680-685.
[24] Liau, C.Y., Chang, T.M., Pan, J.P., Chen, W.L. and Mao, S.J.T. 2003. Purification of human plasma haptoglobin by hemoglobin-affinity column chromatography. J. Chromatogr. B. 790: 209-216.
[25] Lai, I.H., Tsai, T.I., Lin, H.H., Lai, W.Y. and Mao, S.J.T. 2007. Cloning and expression of human haptoglobin subunits in Escherichia coli: delineation of major antioxidant domain. Protein Expr. Purif. 52: 356-362.
[26] Lorenz, J.N. and Gruenstein, E. 1999. A simple, nonradioactive method for evaluating single-nephron filtration rate using FITC-inulin. Am. J. Physiol. 276: F172-177.
2004. Serial determination of glomerular filtration rate in conscious mice using FITC-inulin clearance. Am. J. Physiol Renal Physiol. 286: F590-596.
[28] Lim, S.K., Ferraro, B., Moore, K. and Halliwell, B. 2001. Role of haptoglobin in free hemoglobin metabolism. Redox. Rep. 6: 219-227.
[29] Kurosky, A., Hay, R.E. and Bowman, B.H. 1979. Canine haptoglobin: A unique haptoglobin subunit arrangement. Comp. Biochem. Physiol B. 62: 339-344.
[30] Mominoki, K., Tosa, N.N., Morimatsu, M., Syuto, B. and Saito, M. 1995. Haptoglobin in Carnivora: A unique molecular structure in bear, cat and dog haptoglobins. Comp. Biochem. Physiol. B, Biochem. Mol. Biol. 110: 785-789.
[31] Tosa, N., Morimatsu, M., Nakagawa, M., Miyoshi, F, Uchida, E., Niiyama, M., Syuto, B. and Saito, M. 1993. Purification and identification of a serum protein increased by anthelmintic drugs for Dirofilaria immitis in dogs. J. Vet. Med. Sci. 55: 27-31.
[32] Martin, M., Tesouro, M.A., Gonz Am, N., Pi, A. and Lampreave, F. 2005. Major plasma proteins in pig serum during postnatal development. Reprod. Fertil. Dev. 17: 439-445. [33] Chan, J.P.W., Chu, C.C., Fung, H.P., Chuang, S.T., Lin, Y.C., Chu, R.M. and Lee, S.L.
2004. Serum haptoglobin concentration in cattle. J. Vet. Med. Sci. 66: 43-46.
[34] Delanghe, J., Langlois, M. and De Buyzere, M. 1998. Congenital anhaptoglobinemia versus acquired hypohaptoglobinemia. Blood. 91: 3524.
American Negroes: evidence for a third allele Hp-2m. Am. J. Hum. Genet. 12: 160-169. [36] Manoharan, A. 1997. Congenital haptoglobin deficiency. Blood. 90: 1709a-1709.
[37] Koda, Y., Soejima, M., Yushioka, N. and Kimura, H. 1998. The haptoglobin gene deletion responsible for anhaptoglobinemia. Am. J. Hum. Genet. 62: 245-252.
[38] Koda, Y., Watanabe, Y., Soejima, M., Shimada, E., Nishimura, M., Morishita, K., Moriya, S., Mitsunaga, S., Tadokoro, K. and Kimura, H. 2000. Simple PCR detection of haptoglobin gene deletion in anhaptoglobinemic patients with antihaptoglobin antibody that causes anaphylactic transfusion reactions. Blood 95: 1138-1143.
[39] Kino, K., Mizumoto, K., Wanatabe, J. and Tsunoo, H. 1987. Immunohistochemical studies on hemoglobin-haptoglobin and hemoglobin catabolism sites. J. Histochem. Cytochem. 35: 381-386.
[40] Lim, S.K., Kim, H., Lim, S.K., bin Ali, A., Lim, Y.K., Wang, Y., Chong, S.M., Costantini, F. and Baumman, H. 1998. Increased susceptibility in Hp knockout mice during acute hemolysis. Blood. 92: 1870-1877.
Fig. 1. Schematic drawing of molecular arrangement in human Hp phenotypes. Hp 1-1 exclusively possesses only the basic dimer (α1β)2, whereas Hp 2-1 is comprised of
polymeric structures: dimer (α1β)2, trimer (α1β)(α2β)(α1β), and other linear polymers. Hp
2-2 is comprised of a trimer (α2β)3 and other cyclic polymers. α1, α2, and β are 83, 142, and
245 amino-acid residues in length, respectively. α2 is similar to α1, differing only by an additional insertion of a repeat identical to 3/4 of α1. Owing to the extra Cys-74 in α2, Hp 2-1 and 2-2 are able to form complicated polymers.
Hp 1-1 Hp 2-1 Hp 2-2
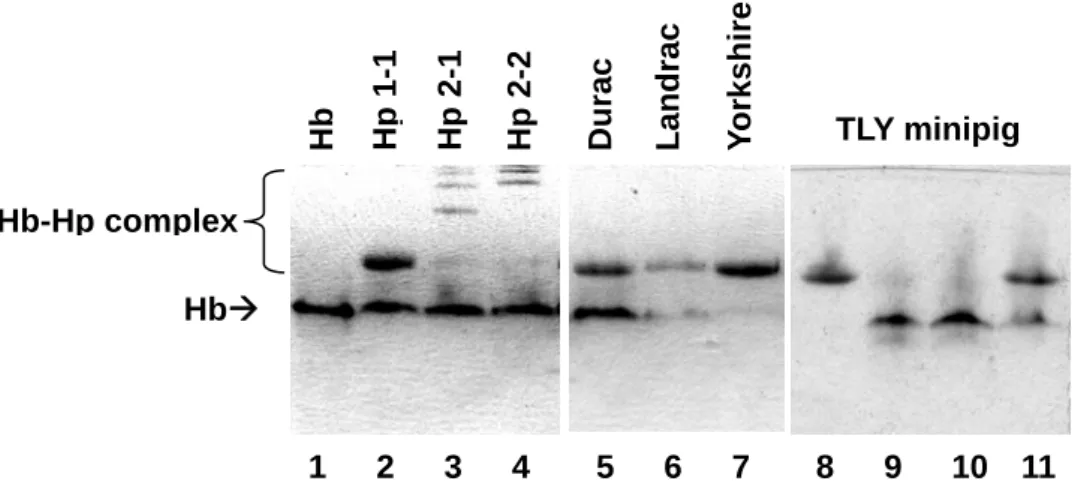
Fig. 2. Phenotyping of human and porcine Hp on a 7% native-PAGE. Briefly, each plasma (8 μl) was incubated with hemoglobin (Hb) (16 μg/4 μl) at room temperature for 30 min. Following electrophoresis, the gel was directly stained in a freshly prepared DAB solution containing 0.05% H2O2 as a developer for the endogenous peroxidase activity of
hemoglobin. Lane 1: Hb alone. Lanes 2-4: Human plasma Hp 1-1, 2-1, and 2-2 complexed with Hb. Lanes 5-7: Three species of domestic porcine plasma. Lanes 8-11: Four individuals of TLY minipigs plasma.
Hb HbÆ Hb-Hp complex Hp 1-1 Durac Landrac Yo rkshire TLY minipig 1 2 3 4 5 6 7 8 9 10 11 Hp 2-1 Hp 2-2
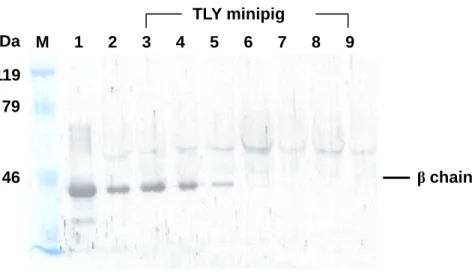
Fig. 3. Western blot of plasma Hp using rat polyclonal antibody prepared against porcine Hp. Porcine plasma (1 μl) and purified Hp (3 μg) containing 1% 2-mercaptoethanol was loaded on a 8% SDS–PAGE. After transferring and blocking, the separated proteins were incubated with rat antiserum against porcine Hp β chain, followed by an incubation with peroxidase-labeled goat anti-rat IgG. The membrane was then developed by a DAB solution containing 0.05% H2O2. Lane M: Molecular markers. Lane 1: Purified porcine Hp. Lane
2: Yorkshire plasma. Lanes 3-9: Seven individuals of TLY plasma. The weak bands showing a molecular weight greater than Hp β chain in each lane represent those from a non-specific binding between secondary antibody (goat anti-rat IgG) and porcine IgG.
kDa 46 119 79 β chain M 1 2 3 4 5 6 7 8 9 TLY minipig