Garlic: Health Benefits and Impacts
Chia-Wen Tsai1, Haw-Wen Chen1, Le-Yen Sheen2, Chong-Kuei Lii1*
1Department of Nutrition, China Medical University, Taichung, Taiwan
2Institute of Food Science and Technology, National Taiwan University, Taipei, Taiwan
Supported by CMU97-134 and CMU98-CT-21
Correspondence: Dr. Chong-Kuei Lii, Department of Nutrition, China Medical University, Taichung, Taiwan; email: cklii @mail. cmu. edu.tw ; Fax: +886-4-22062891.
Abstract
Recent years have seen an increasing emphasis on foods and food components in disease prevention. Garlic (Allium sativum L.), one of the best-researched herbal remedies, holds a unique position in history, traditionally employed to treat infection, cold, diabetes, heart disease, and a host of other disorders. Clinically, it has been evaluated for lowering blood pressure, cholesterol and/or glucose concentration, as well as for prevention of arteriosclerosis and cancer. Epidemiologically, garlic consumption inversely correlates with risk of oral, stomach, esophageal, colon, and prostate cancers. In addition, biological activities of garlic including antibacterial, antithrombotic, antioxidant, immunomodulatory, and antidiabetic actions and drug metabolism modulation have been extensively investigated. Here we briefly summarize recent findings on garlic and its sulfur-containing compounds in preventing cardiovascular diseases and cancer, along with modulating drug-metabolizing enzymes and membrane transporter activities. Finally, garlic safety and drug interaction are discussed. Keywords: Garlic; Cardiovascular disease; Cancer; Drug metabolism; Food-drug interaction;
Chemicals and bioactive components of garlic
Unique flavor and health promoting functions of garlic are generally attributed to rich content of sulfur containing compounds: i.e., alliin, γ-glutamylcysteine, and their derivatives. Processing of a fresh and intact garlic bulb by crushing, grinding or cutting induces release of the vacuolar enzyme alliinase, which very quickly catalyzes alliin to allicin [1, 2]. Yet allicin is a very unstable compound, soon rearranged and transformed into numerous of lipid-soluble sulfur containing by-products, mostly diallyl disulfide (DADS), also into diallyl sulfide (DAS), diallyl trisulfide (DATS), allylmethyl trisulfide, and diallyl tetrasulfide [1]. These compounds emit strong odors and are kept in garlic oil. Under appropriate conditions, allicin can be transformed into other lipid-soluble products such as ajoene and vinyldithiin. Ajoene is
identified as a principal product in garlic extract prepared by using ether as a solvent [3]. In contrast to the processes stated above, alternative pathways occur in case of different garlic storage. An aging process by immersing intact or sliced raw garlic in alcohol or vinegar for several months results in sulfur-containing compounds in this aged product dramatically different from that in garlic oil. This aging process is supposed to cause considerable loss of allicin. Meanwhile, with the action of γ-glutamyltranspeptidase, the other sulfur-containing precursor γ-glutamylcysteine is transformed into water-soluble S-allylcysteine (SAC) and the
subsequent metabolites, including S-allylmercaptocysteine (SAMC) and S-methylcysteine. Unlike oily sulfur compounds, these water-soluble compounds are odorless but have more delicate and less characteristic flavor [4].
In addition to sulfur containing compounds as stated above, garlic is also rich in trace elements. In raw garlic, amounts of zinc, manganese, copper, selenium, and iodine are 556.1, 446.9, 143.3, 5.5 and 2.5 g, respectively, in 100 g fresh weight [5]. Protein content in raw garlic ranges from 2.6 to 3.0%, depending on garlic variety. Average content of free amino acids is 2.13%. Concentrations of dietary fiber, total tocopherols in raw garlic are 2310 and 103.1 mg/100 g fresh weight, respectively. Ascorbic and total polyphenols levels are 73.6 and 1.9 mg in 100 g dry weight [6]. Over 70 fatty acids were determined, with linoleic (46-53%), palmitic (20-23%), oleic (4-13%) and -linolenic acids (3-7%) most abundant, accounting for 80% of total lipids [7].
Garlic preparations and supplements
Because of the complex chemistry of garlic, variations in processing methods can yield quite different preparations. Raw garlic homogenate, major preparation of garlic, is the most common form of garlic consumption, allicin the main compound present in fresh raw garlic homogenate. Currently, many garlic supplements are marketed, with garlic oil, powder, and aged extract most popular. Garlic oil is mostly obtained by steam distillation, with a yield
around 2.5-3 g per kg fresh garlic. In garlic oil, DAS, DADS, and DATS, differing in number of sulfur atoms, and allylmethyl sulfide are four most abundant volatile allyl sulfides [8]. Garlic power is generated from garlic cloves dehydrated and pulverized into powder. Due to heat deactivation of alliinase during dehydration, the major active constitutes of garlic powder is alliin and a small amount of oil-soluble sulfur compounds. To overcome the strong and irritant odor and possible side effects of raw garlic and garlic oil, including growth retardation and destruction of gut microflora, an “aging’ process has been applied to garlic. Aged garlic is prepared by soaking of whole or sliced garlic cloves in alcohol or vinegar solution for 6-20 months, which removes several irritant sulfur containing compounds and also stabilizes some unstable compounds such as allicin [4, 9]. Water-soluble compounds SAC and SAMC are the most abundant sulfur-containing components; trace amounts of oil-soluble allyl sulfides exist in aged garlic. In contrast to odoriferous garlic oil and raw garlic, garlic powder and odorless aged garlic product are currently the most popular garlic supplements on the market.
Garlic and cardiovascular disorders
Cardiovascular disease is a common human chronic disease, and it is the leading cause of morbidity and mortality in the United States [10]. Etiology of cardiovascular disorders is multifactorial: e.g., hypercholesterolemia, hypertension, diabetes mellitus, familial heredity, hyperhomocysteinemia, increase of oxidative damage, smoking as well demonstrated risk factors [11]. Due to accompanying inflammation in the plague, cardiovascular disorders are regarded as chronic inflammation related diseases [11]. Increase ofproduction and release of inflammatory mediators, such as reactive oxygen species (ROS), tumor necrosis factor (TNF)-, interleukin (IL)-6, arachidonic acid metabolites, and nitric oxide is noted in the atherosclerotic lesion [12]. This results in greater expression of adhesion molecules, including P-selectin, E-selectin, vascular cell adhesion molecule (VCAM), intercellular adhesion molecule (ICAM), and monocyte chemotactic protein-1 on cell surfaces of monocytes and
leukocytes and vascular endothelial cells, which accelerates the adherence of monocytes and leukocytes on the vascular endothelium and subsequent transmigration into the subendothelial space. Within the intima, activated macrophages release reactive oxygen species, scavenge oxidized LDL (oxLDL), become foam cells, and lead to development of the fatty streak in the early stage of atherosclerosis [13, 14]. This explains why phytochemicals with anti-chronic inflammation, hypolipidemia, and antioxidation properties are thought capable of alleviating incidence of atherosclerosis.
Garlic has been regarded as a potent anti-atherogenic food [15]. Its lowering of blood cholesterol is believed largely due to a reduction in LDL-cholesterol [16, 17], which may be due to inhibition of hepatic hydroxymethylglutaryl-CoA reductase activity by alliin and allicin [18]. Over the past decade, several intervention studies and systemic meta-analysis reviews had investigated effectiveness of garlic prevention against cardiovascular disease and its properties (Table 1). In a double-blinded placebo-controlled randomized study with 51 coronary heart disease patients, results indicated that 12-mo treatment with 300 mg/d garlic powder significant decreases total cholesterol and LDL-cholesterol [19]. A reduction of 32.9 and 27.3 mg/dl in LDL-cholesterol by garlic was observed in men and women, respectively. Similar reduction of total cholesterol and LDL-cholesterol and increase of HDL-cholesterol were also reported in hypercholesterolemic adults, who were administered with 10 g/d garlic extract for 4 mo or 5 g/d raw garlic for 6 wks or 600 mg/d garlic powder for 12 wks [20-22]. Oily macerate of garlic (1620 mg/day for 30 d) was found to significantly lower levels of total cholesterol, LDL-cholesterol, and triglycerides in 70 hypertensive adults [23]. Still, Burggraaf and colleagues reported that a 12-wk of 2.1 g/d garlic powder administration does not change lipid profiles in overweight subjects with normal blood lipid [24]. Recently, a meta-analysis including 29 trials revealed garlic supplementation markedly reducing blood total cholesterol ( 0.19; 95 % CI 0.33, 0.06 mmol/L) and triacylglyceride levels ( 0.11; 95 % CI 0.19, -0.06 mmol/L) but exhibiting no significant effect on LDL- or HDL-cholesterol [25]. Similar
reduction on blood total cholesterol and triglycerides appeared in systemic reviews [26, 27]. Inconsistent clinical evidence warrants more study before reaching compelling conclusions.
Garlic is reported to prevent cardiovascular disease by multiple effects, one of which is inhibition of platelet aggregation. A single intravenous dose of aqueous extracts of garlic (10-100 mg/kg) dose-dependently inhibits blood thromboxane B2 concentration in rabbits [28]. Maximum inhibition of thromboxane B2 occurred at 0.5 h post injection and lasted to 6 h. In another study, oral administration with aqueous extract of fresh garlic inhibited cyclooxygenase activity in rabbit platelets, results in suppressing thromboxane formation and blood aggregation [29]. Similarly, in eight males (40 to 50 years old) consumption of one clove crushed garlic daily for 16 wks resulted in 80% reduction of serum thromboxane B2 levels [30]. Allicin and allicin-derived thiosulfinates are recognized as major compounds responsible for antithrombotic activity of garlic [31]. Besides inhibition of cyclooxygenase activity, other possible mechanisms for inhibiting platelet aggregation by garlic include suppressing intraplatelet Ca2+ mobilization, increasing cAMP and cGMP levels, increasing platelet-derived nitric oxide production, and reducing platelet binding to fibrinogen [32].
Garlic protection against cardiovascular diseases has been partly attributed to its potent anti-inflammatory activity [33]. Ethyl acetate-soluble fraction of garlic is proven effective in inhibiting nuclear factor B (NF-κB) activation as well as expression of cyclooxygenase 2 and inducible nitric oxide synthase in IL-3-dependent murine pro-B cells Ba/F3 through the Toll-like receptor-dependent pathway [34]. Thiacremonon, a novel organosulfur compound of garlic, inhibits 12-O-tetradecanoylphorbol-13-acetate-induced ear edema in ICR mice and carrageenan- and mycobacterium butyricum-induced inflammatory and arthritic responses in paws of Sprague-Dawley rats [35]. Garlic oil reportedly suppresses 1-chloro-2,4-dinitrobenzene (CDNB)-induced contact hypersensitivity as determined by ear swelling [36]. After 30-d administration of 600 mg/kg garlic powder, increase of interferon and decrease of IL-4 in phytohemagglutinin activated splenocytes were noted, suggesting garlic treatment
may favor a Th2 or humoral immune response [37]. Recently, 1,2-vinyldithiin was reported to significantly suppress IL-6 and monocyte chemoattractant protein 1 secretion by macrophage-secreted factors stimulated human preadipocytes isolated form subcutaneous adipose tissue of nonobese young women [38]. DAS was reported to prevent COX-2 upregulation and PGE2 secretion in primary human synovial fibroblasts and articular chondrocytes induced by IL-1 and monosodium urate crystal and ameliorates crystal induced synovitis potentially through NF-B signaling pathway [39].
The presence of proinflammatory cytokines initiates numerous physiological changes on vessel walls, such as enhanced adhesion of leukocytes to endothelium. Recent in vitro study indicated that the chloroform extract of aged black garlic attenuates TNF-α-induced VCAM-1 expression via NF-κB dependent pathway in human umbilical vein endothelial cells (HUVEC), hence decreasing adhesiveness of monocytes to endothelial cells [40]. In primary human coronary artery endothelial cells, aqueous extract of garlic (0.25-4.0 mg/ml) dose-dependently curbs ICAM-1 and VCAM-1 expression induced by IL-1α [41]. When stimulated with oxLDL, DADS and DATS suppress VCAM-1 and E-selectin expression in HUVEC and subsequent adhesion of HL-60 to endothelial cells [42]. Taken together, though lacking solid clinical evidence that garlic protection of blood vessels can be attributed to its anti-inflammation property, potent anti-inflammatory action of garlic and its sulfur-containing compounds obtained from in vitro and animal studies support the potential value of garlic in preventing atherogenesis.
Evidence indicates garlic also acting to maintain vascular tone and cardiac functions. Experiments on laboratory animals and investigations of humans proved diets supplemented with garlic as restoring endothelial functions. Allicin is believed the active component of raw garlic protecting coronary endothelial function and vasoreactivity in pulmonary hypertensive rats [43]. Enhancement of nitric oxide synthase activity and greater nitric oxide production
partly explained this hypotensive action. Our recent work demonstrated DADS and DATS protect endothelial nitric oxide’s synthase activity and production against oxLDL insult to endothelial cells [44] attributable partly to their mediation of phosphatidylinositol 3-kinase/protein kinase B signaling and prevention of eNOS degradation [44]. SAC supplementation reduces incidence of stroke in stroke-prone spontaneously hypertensive rats [45] and lowers mortality and infarct size in an acute myocardial infarction model induced by coronary artery ligation [46). In a streptozotocin-induced diabetic animal experiment, rats were orally administered with 0-100 mg/kg/d garlic oil for consecutive 16 d; streptozotocin-induced cardiac contractile dysfunction and apoptosis was markedly improved by garlic oil [47]. In a hypercholesterolemic animal experiment, rats were fed with 1.0% garlic and 0.5% turmeric supplemented diet for 10 wks. Enhanced vasorelaxation in aortic ring response to adenosine, acetylcholine, and isoproterenol, along with attenuation of contractile response to 5-hydroxytryptamine, were seen in animals of garlic and turmeric supplemented diet, thus lowering blood pressure [48]. In a randomized, placebo-controlled, cross-over design with 15 angiographically proven coronary artery disease patients, brachial artery flow mediated endothelium-dependent dilation was improved by aged garlic extract [49]. Similar to aged garlic extract, garlic oil in a dose of 250 mg/d for 2 mo has demonstrably improved both systolic and diastolic blood pressure in 20 hypertensive patients [13].
Garlic and cancer
The past few decades have seen many epidemiological studies of the correlation between garlic consumption and incidence of cancer; inverse relationship has emerged. Setiawan and colleagues observed negative dose-response relationship between monthly intake of garlic and risk of stomach cancer in Shanghai and Qingdao, China [50]. A recent study found odds ratios among persons with high versus low intakes of garlic and onions that correlated with starkly reduced risk of colorectal adenoma [51]. Persons who consume a high proportion of garlic
decreasing susceptibility to stomach and colon cancers is also reported [52]. Based on the US Food and Drug Administration’s evidence based review system for scientific evaluation of the risk of diverse types of cancers, 19 human studies reveal garlic’s anti-tumorigenic potential in stomach, colonrectal, breast, lung, and endometrial cancers. Very limited evidence supports a relation between garlic consumption and reduced risk of colon, prostate, esophageal, larynx, oral, ovary, or renal cell cancers [53].
Several human intervention studies plot garlic’s anti-carcinogenic traits. In a preliminary double-blind, randomized clinical trial using high-dose aged garlic extract (2.4 mL/d) as active treatment and low-dose aged garlic extract (0.16 mL/d) as control was performed on 51 patients with colorectal adenomas-precancerous lesions of the large bowel [54]. After 12 mo of treatment, 37 patients (19 in the active, 18 in control group) completed the study, with size and number of colon adenomas in the high-dose group significantly suppressed (P=0.04). An earlier double-blind intervention study of 5,033 subjects (2,526 in the intervention and 2,507 in the control group) was performed in China. A dose of 200 mg/d DATS in combination with 100 g/d selenium was taken by the intervention group each month for three years. Results showed that the DATS offered protection against gastric cancer for males [55]. In this study, it is interesting to note that no such protection appears in females.
Numerous studies on animal models found in literature were carried out using either garlic extract or individual garlic-derived compounds. Development of the aflatoxin B1- or diethylnitrosamine-induced liver cancer in rats was limited by fresh garlic [56] and garlic oil [57]; the latter protected against ferric nitrilotriacetate-induced kidney cancer growth in rats [58]. DADS suppressed 7,12-dimethylbenzo[a]anthracene (DMBA)-induced rat mammary tumor [59]. DAS and DATS protected against DMBA-, phorbol ester-, and benzo[a]pyrene-induced skin tumorigenesis in mice [60-63]; DATS inhibited growth of PC-3 human prostate cancer xenografts in male nude mice [64]. Similarly, ajoene significantly inhibited B16/BL6 melanoma growth and metastasis to lung in C57BL/L mice [65]. Aside from oil-soluble
organosulfur compounds, water-soluble SAC inhibited growth and malignant progression of highly metastatic human non-small-cell lung carcinoma in nude mice [66].
While the precise mechanism of anticancer efficacy of garlic is still not clear, molecular action such as regulation of cell proliferation, increase of tumor apoptosis, blocking cell cycle, inhibited carcinogen activation, increase of phase II drug metabolizing enzymes, enhanced anti-oxidation capacity, change of proteasome-dependent protein degradation, and modulation of immune response have been proposed and extensively probed in recent years (Table 2).
In many cancer cells, garlic organosulfur compounds display potential for suppressing growth of cancer cells and leading to cell cycle arrest. DAS increases accumulation of sub-G1 DNA and concomitant accumulation of cells in the G2/M phase in a dose-dependent manner in human anaplastic thyroid carcinoma cells [67]), as well as in human colon cancer cells [68]. DAS, DADS, and DATS further exhibited differential effects on lowering of cyclin-dependent kinase-Cdk7 and raising cyclin B1 protein levels in J5 human liver tumor cells, thus arresting cells in the G2/M phase [69]. Among those lipid-soluble allyl sulfides, which differ in number of sulfur atoms, DATS revealed better growth inhibition of human melanoma A375 cells and skin basal cell carcinoma cells than DADS and DAS did [70]. Induction of apoptosis and cell cycle arrest by garlic allyl sulfides are also reported in different types of cancer cells: e.g., human lung adenocarcinoma [71], glioblastoma [72], prostate cancer [73, 74], neuroblastoma [75], gastric cancer [76], bladder cancer [77], colon cancer [78], and mammary cancer [79].
Garlic organosulfur compounds resulting in cell cycle arrest and apoptotic process can be linked to modulation of several key elements in cellular signal transduction. It has been demonstrated that DATS-induced apoptosis of human prostate cancer cells PC3 involves c-Jun N-terminal kinase (JNK) and extracellular-signal regulated kinase (ERK)-mediated phosphorylation of Bcl-2 [80]. Inactivation of Akt signaling pathway also likely plays a role in DATS-induced mitochondrial translocation of BAD and caspase-mediated apoptosis in
PC3 and DU145 human prostate cancer cells [81]. Likewise, DATS arrest of DU145 cells in G2/M phase effected by hyperphosphorylation of Cdc25C [82] and delayed cdk1 translocation into nucleus [83], as well as by oxidative modification of -tubulin in human colon cancer cells, which impedes polymerization of tubulin [84]. Similar interruption on tubulin polymerization is reported by treating SW480 and NIH3T3 fibroblasts with SAMC; it subsequently arrests cells in mitosis and triggers JNK1 and caspase-3 signaling pathways leading to apoptosis [85]. In B16F-10 melanoma, apoptosis DADS-induced correlates with mitochondria-dependent pathway by up-regulating p53 and caspase-3 while down-regulating NF-B-mediated Bcl-2 activation [86]. Recently, both extrinsic and intrinsic death pathways were shown as involved in allicin induction of apoptosis in gastric SGC-7901 cancer cells [87].
Garlic organosulfur compounds may also act on epigenetic and exert its anti-carcnogenic activity. Histone acetylation notably increases in colonocytes isolated from DADS-treated rats and also in erythroleukemia cells from SAMC-treated mice, suggesting histone deacetylase as the target of garlic allyl compounds [88, 89]. In addition to DADS, other garlic organosulfur compounds have been tested and allyl mercaptan, metabolite of DADS, exerts the most potent histone acetylase inhibitor in assays with HeLa nuclear extracts, lysates from human colon cancer cells, or purified human histone deacetylase 8 [78, 90]. Allyl mercapton inhibition of histone deacetylase activity results in increasing histone acetylation and Sp3 transcription factor binding to the p21WAF1 gene promoter region, elevating p21 expression and cell cycle arrest of HT29 colon cancer cells [90]. Enzyme kinetics assays further reveals inhibition of allyl mercatpan on histone deacetlase through a competitive mechanism (Ki = 24 μM)[90].
Evidence indicates tumor invasion and metastasis as suppressed in the presence of garlic and its organosulfur compounds. DATS administration retards growth of PC-3 human prostate cancer xenograft in athymic mice [64] and prevents progression to invasive carcinoma and lung metastasis in transgenic adenocarcinoma of mouse prostate (TRAMP)
[91]. In in vitro experiments, DADS suppressed invasion of human prostate cancer LNCaP cells is attributed to inhibition of matrix metalloproteinase (MMP)-2 and -9 activity and to tightening of tight junctions [92]. Garlic suppression on tumor invasion may also be attributed to its action on E-cadherin expression. SAC and SAMC restoration of E-cadherin expression suppresses prostate cancer cell proliferation and invasion [93]. This increase of E-cadherin expression and inhibition of cell proliferation are also noted in oral squamous cancer CAL-27 cells in the presence of SAC [94]. Invasive activities of colorectal cancer SW480 and SW620 cells are inhibited by aged garlic extract, while aged garlic extract has no effect on invasion of HT29 cells, suggesting anti-invasive action of aged garlic extract as cancer cell-dependent [95]. In the presence of ajoene, human leukemia HL60 cells were arrested in the G2/M phase; both trypsin- and chymotrypsin-like proteasome catalytic activities inhibited [96].
Taken together, most animal and cell studies suggest garlic as a potent chemopreventive agent for several types of cancers by inhibiting cell proliferation, arresting cell cycle, inducing cell apoptosis, as well as blocking invasion and metastasis.
Garlic and detoxification system
Cancer chemopreventive effect of garlic organosulfur compounds is believed associated with modulation of carcinogen metabolism, including effects on Phase I and II detoxification enzymes. Phase I enzymes, mainly cytochrome P450 (CYP), detoxify a variety of endogenous and exogenous chemicals and activate many carcinogens [97]. Phase II enzymes catalyze conjugation of Phase I metabolites to various water-soluble molecules like glutathione (GSH), glucuronic acid or sulfate, accelerating metabolite excretion rate. Efficacy of DAS, DADS, and DATS on transcriptional regulation of Phase I and II detoxification enzyme expression positively correlates with suppression of aflatoxin B1- and benzo[a]pyrene-induced liver and forestomach neoplastic formation in mice and rats [98, 99]. Decreased 7,12-dimethylbenzo[a] anthracene-induced DNA adduct formation in rat mammary tissue by DADS [59], protection
against benzo[a]pyrene-induced skin tumorigenesis and micronucleated reticulocyte formation in mice by DAS [63], suppression of aflatoxin B1-induced DNA breaks by allicin, DAS, DADS, and SAC in HepG2 cells [100] can also be explained by their effectiveness on modulating metabolism of carcinogens.
Among CYP isozymes, decrease of CYP 2E1 activity and protein levels is reported in rats fed a diet containing 5% garlic powder [101]. This down-regulation of CYP 2E1 by garlic suppresses hepatic preneoplasia formation in rats initiated with diethylnitrosamine [102]. Glycidamide, active metabolite of acrylamide, formation in rat liver tissues diminishes via inhibiting CYP 2E1 by DAS [103]. In addition to DAS, reduction of CYP 2E1 activity and expression result from garlic oil, DADS and allyl methyl sulfide [104, 105]. In contrast to down-regulation of CYP 2E1, isozymes CYP1A1, CYP1A2, CYB2B1, and CYP3A2 activity as well as protein and mRNA levels are up-regulated by garlic organsosulfur compounds. Dosing rats with 200 mg/kg DAS and allyl methyl sulfide raise CYP1A1, CYP1A2, and CYP3A2 protein levels in a time-dependent manner, a rise noted 24 h post treatment [105]. A dose-dependent increase of rat liver CYP 1A1, 2B1, and 3A1 activities and gene transcription also emanates from garlic oil (30-200 mg/kg body weight), probably combined effect of three major allyl sulfides DAS, DADS, and DATS, in the garlic oil [104, 106, 107].
Besides acting on the stage of gene transcription, garlic constituents may bind to CYP and change its enzyme activity. Using human liver microsomes, activity of CYP 2C9, 2C19, 3A4, 3A5, and 3A7, but not 2D6, is inhibited by incubating with garlic oil or extracts of fresh garlic, garlic powder, or aged garlic [108]. In the case of CYP 2E1, diallyl sulfone and diallyl sulfoxide, metabolites of DAS, act as suicide substrates [109]; this inhibited CYP 2E1 activity explains partly the DAS action on attenuating acetaminophen- and carbon tetrachloride- and ischemic-reperfusion-induced toxicity in rat livers [110, 111].
Phase II detoxification enzymes are known to play a key role in accelerating excretion rate of numerous xenobiotics. Induction of Phase II enzymes like glutathione S- transferase
(GST), epoxide hydrolase (EH) , UDP-glucuronyl transferase (UGT), sulfotransferase, and NAD(P)H quinone oxidoreductase 1 (NQO1) is considered a crucial mechanism protecting organisms against chemical insult. It is thus reasonable to speculate that the inducibility of phytocompounds on Phase II enzymes associated with their efficacy in chemoprevention [112, 113]. GST is among the most important Phase II enzymes, its vital role in cancer prevention supported by finding 7,12-dimethylbenzanthracene-induced skin cancer significantly elevated in form of GST (GSTP)-null mice [114]. Increase of GST activity by garlic organosulfur compounds including allyl methyl trisulfide, allyl methyl disulfide, DATS and DAS strongly correlate with inhibition of benzo[a]pyrene-induced forestomach neoplasia [115]. Effects of DAS, DADS, and DATS on transcriptional regulation of GST enzyme expression are also positively correlated with their suppression of aflatoxin B1- and benzo[a]pyrene-induced liver and forestomach neoplastic formation [98, 99]. SAC decrease of 7,12-dimethylbenz[a] anthraxcene-induced hamster buccal pouch carcinogenesis is also accompanied by enhanced GST activity and GSH level [116].
Up-regulation of Phase II detoxification enzyme gene transcription involves a series of signaling pathways and transcriptional factors. Among those, the pivotal role of nuclear factor E2-related factor 2 (Nrf2) is well documented [112, 117]. Activation and binding of Nrf2 to promoter antioxidant response element/electrophile response element (ARE/EpRE) increases transcription of GST, NQO1, UGT, and sulfotransferase. By treating with garlic organosulfur compounds, Nrf2 nuclear translocation is increased and NQO1 expression is up-regulated in HepG2 cells and mice [118, 119). Increased hepatic NQO1 and GST activity helps attenuate carbon tetrachloride-induced liver injury in rats orally dosed with 500 mol/kg DATS for 5 consecutive days [120].
Dozens of organosulfur compounds have been identified in garlic products; they appear to vary in biological activity. It is interesting to ask what chemical characteristic of those garlic-derived compounds determines modulatory potency on drug metabolism. Evidence
from structure-function relationship studies indicate number of both allyl groups and sulfur atoms of each organosulfur compound as determining factors in transcription of Phase I and II enzymes. To Phase II detoxification enzymes, numbers of sulfur atoms and allyl groups positively correlate with potency on enhancing gene transcription. DATS displays the best induction of NQO1, follows by DADS; DAS has only minor effect [118]. Compare to DATS, DADS at a 10-fold higher dose (100 mol/kg) increase expression of GST and NQO1 in rat liver; DAS does not [121]. Similar findings (DATS>DADS>DAS) are reported on induction of pi form of GST (GSTP) in rat liver [107, 122]. Increase of UGT activity is also noted in HepG2 cells treated with DAS, dipropyl sulfide (DPS), and DADS; effective concentration of DAS and DPS (50 M) is much higher than that of DADS (2.5 M) [123]. Feeding rats a diet with 5% garlic powders markedly raises hepatic UGT activity in an alliin content-dependent manner [101]. Comprehensive study to examine effect of allyl sulfides DAS, DADS, DPS, and dipropyl disulfide (DPDS) on hepatic, renal, intestinal, and pulmonary Phase II enzymes GST, EH, UGT, and NQO, was performed by Guyonett et al. [124]. After oral dosing Wistar rats with 1 mmol/kg of each compounds for four consecutive days, DADS exerts the greatest inducibility on all Phase II detoxification enzymes, with pulmonary EH activity unchanged. In addition, induction on NQO activity is seen in DADS-treated animals only. DAS, DPS, and DPDS increase of GST and EH activities are only noted in liver. Later, increase of hepatic GST and NQO1 expression and activity by allyl-containing compounds was demonstrably greater than that of propyl-containing ones: i.e., DADS>DPDS and DATS>dipropyl trisulfide [118, 125, 126]. These findings suggest garlic alk(en)yl sulfides exerting differential induction potency on Phase II enzymes and such induction as tissue-specific.
As for Phase I enzymes, garlic components with allyl side chain have better inducibility on most CYP isozyme expression than propyl or methyl-containing ones. However, effect of sulfur atom number on CYP expression differs from their action on Phase II enzymes: garlic compounds with higher number of sulfur atoms display less inducibility [105, 107, 115, 122,
126]. This discrepancy suggests the regulatory mechanism of garlic organosulfur compounds on Phase II and CYP isozymes as different; precise active mechanism warrants further study.
Garlic and antioxidation
Oxidative stress is a state wherein balance between radicals generated and free radical or oxidant scavenging capacity of the endogenous antioxidant system is disrupted. Oxidative stress is documented as involved in pathogenesis of chronic diseases, including cardiovascular disorder and cancer. Hence, compounds with antioxidant properties may be used to prevent oxidative stress-mediated diseases [127]. Numerous studies have demonstrated garlic and its organosulfur compounds as potent antioxidants by displaying radical-scavenging activity and modulating cellular antioxidant enzyme activity. Aged garlic extract and SAC are shown to scavenge ROS and protect endothelial cells injury by oxLDL [128] and PC12 neuron cells damage by H2O2 [129]. Garlic extract is proven as effective as N-acetylcysteine in lessening ROS formation and GSH depletion induced by acetaminophen in rat primary hepatocytes [130]. Garlic pretreatment with 1 g/kg for 5 weeks reduces iron-catalyzed lipid peroxidation by lowering malondialdehyde level in rat liver and colon, along with enhancing status of antioxidants [131]. Likewise, garlic reduces iron-induced cell proliferation and autophagy and protects mitochondrial membrane by lowering iron storage in the liver [131]. Garlic oil is effective in reducing tributyltin-induced oxidative damage in mice and human amniotic cells [132], plus decreasing sodium nitrite-induced neurotoxicity in rats [133]. The aforementioned garlic protection against oxidant-induced damage can be attributed to increase in superoxide dismutase, GSH reductase, -glutamate cysteine ligase, and GST activities and also in GSH production [133-135]. Activated Nrf2 demonstrably plays a key role in garlic enhancement on both antioxidant defense capability and drug metabolism enzymes, as described above [134].
Garlic antioxidation properties have been ascertained in animal disease models. In the fructose-induced metabolic syndrome rat model, aqueous garlic extract attenuates oxidative
stress and prevents vascular remodeling by suppressing NAD(P)H-oxidase [136]. In Type 2 diabetes db/db/ mice, consumption of 5% freeze-dried aged black garlic for seven weeks significantly raises superoxide dismutase, catalase, and glutathione peroxidase activity and lessens lipid peroxidation in the liver [137]. In streptozotocin induced-diabetes rats, garlic oil helps normalize impaired antioxidant status [138]. Less neuron damage accompanied by increased synaptophysin and pre-synaptic protein synaptosomal associated protein of 25kDa (SNAP25), a member of soluble N-ethylmaleimide sensitive factor attachment protein receptors playing a key role in pre-synaptic vesicle fusion and exocytose, have been seen in Alzheimer’s APP transgenic (Tg) mice treated with 2% aged garlic extract containing diet and its active component SAC (20 mg/kg diet) [129]. SAC also reduces lipid peroxidation and superoxide radical production and elevates Cu-Zn-superoxide dismutase activity in 1-methyl-4-phenylpyridinium-induced parkinsonism in mice [139].
In recent years, several human intervention studies have examined antioxidant potency of garlic in humans. Two months of garlic oil (250 mg/d) supplementation starkly reduced oxLDL and 8-iso-prostaglandin F2 levels, these accompanied by significant decline in both systolic and diastolic blood pressures, in hypertensive patients [13]. Similar drop in oxLDL production is reported by dosing 70 hypertensive adults with 1620 mg/d oily macerate of garlic for 30 d [23]. In double-blind placebo-controlled study, plasma oxLDL level sharply fell in those administered for one year with 200 mg/d aged garlic extract combined with multi-micronutrients (folic acid, B6, B12, and L-arginine), as compared to controls [140]. Taken together, these results suggest potent antioxidant properties of garlic in delaying onset and development of cardiovascular disease, cancer, diabetes, and neurodegenerative diseases due to imbalance between free radical production and antioxidant defense.
Garlic and drug interaction
membrane transporter levels in liver, lung, kidney, and intestinal tissues. This raises some possibility of garlic supplementation causing food-drug interaction and change therapeutic efficacy of applied drugs. To resolve this question, in vitro and in vivo experiments have multiplied in recent years. Increased human immunodeficiency virus (HIV) protease inhibitor ritonavir toxicity is reported in AIDS patients co-administered with garlic [141]. This can be explained, at least in part, by inhibiting ritonavir excretion rate: e.g., allicin reported to inhibit p-glycoprotein-mediated efflux of ritonavir in Caco-2 cells [142]. However, by examining the permeability via rat jejunum and Caco-2 cell monolayer, aged garlic extract raises saquinavir and darunavir efflux [143]. Lately, higher darunavir efflux by aged garlic extract is noted in rat liver slices and isolated hepatocytes, whereas saquinavir efflux decreases [144]. The authors propose competitive binding to the same binding sites and positive cooperative effect for distinct binding places as likely responsible for garlic altering efflux of saquinavir and darunavir, respectively (144). Greater multidrug resistance-associated Protein 2 expression is also reported in kidney brush-border membranes by DADS, but not by SAC [145].
For in vivo models, pharmacokinetics of diuretic drug hydrochlorothiazide in rats are calculated following three-week garlic homogenate administration. Results show that garlic homogenate increases bioavailability and half-life while decreasing hydrochlorothiazide clearance [146]. Concomitantly, diuretic effect of hydrochlorothiazide is increased by garlic homogenate. Enhancement on antihypertensive and cardioprotective efficacy of captopril in rats by garlic homogenate and SAC was also reported in a later work [147]. In our laboratory, effect of garlic oil on the pharmacokinetcis of atorvastatin has been determined recently. Rats were orally administered with 50 mg/kg garlic oil for five consecutive days, then a single dose of atorvastatin (10 mg/kg) was given. Rise of p-glycoprotein levels in liver and 3A1/2 activity in both intestinal and liver tissue appear negatively corelated to area under the curve (AUC) of plasma concentration of atorvastatin and its metabolite 2-OH atorvastatin (unpublished data).
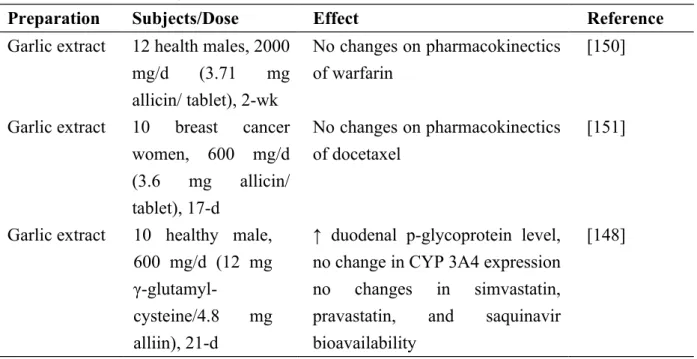
and changes their therapeutic efficacy. To date, limited research has been done (Table 3). In a clinical trial with 10 healthy subjects, 600 mg garlic extract was given daily for 21 d; results indicate garlic extract increasing intestinal p-glucoprotein expression and decreasing AUC of plasma concentration of saquinavir [148]. Our study evaluated pharmacokinetics of two hypocholesterolemic drugs, simvastatin and pravastatin, with their AUC not changed. Due to anti-thrombotic activity, garlic ranks among the most widely used herbal medicines, typically ingested by people receiving warfarin [149]. Change in pharmacokinetics of warfarin by garlic has been determined in clinical trial involving 12 healthy male volunteers. Results show plasma concentration-time profile of warfarin and platelet aggregation unaltered fy warfarin co-administered with garlic (2 g/d) for 2 wks [150]. Garlic influence on pharmacokinetics of docetaxel was also rated in 10 women with metastatic breast cancer [151], treated with 30 mg/m2 docetaxel given weekly for 3 of 4 wks. Three days after the initial dose of docetaxel, patients received 600 mg of garlic twice daily for 12 consecutive days. Results indicated that the clearance of docetaxel and additional pharmacokinetic parameters of docetaxel including peak concentration, AUC, and half-life are not affected. Although garlic had no significant effect on pharmacokinetics of docetaxel, these authors found patients with CYP3A5*3C/*3C genotype having lower mean AUC ratio than those with CYP3A5*1A/*1A [151]. This finding suggests genetic background as a determining factor in the outcome of garlic and drug interaction. Understanding the genotype of each tested individual may help in evaluating whether garlic interacts with drug and changes its therapeutic efficacy.
Although the results remain inconsistent and contradictory, based on garlic potency on modulating drug-metabolizing enzymes and membrane transporters activity and expression, possibility of garlic affecting drug therapeutic efficacy cannot be excluded. To clarify whether garlic interacts with drug metabolism and alters its pharmacokinetics, more well-designed study is warranted.
Safety of garlic
Consumed for hundreds of years, garlic is regarded as a safe food. Yet in addition to possible interaction with drugs as cited above, several health risks are reported for excess consumption of garlic or for those contacting it in the workplace. Gastrointestinal tract injury and allergic reactions spawned by garlic in particular attract concern. Increased exfoliation of gastric surface epithelial cells in healthy subjects is reported by intragastric infusion of single dose of raw garlic over 0.75 g [152]. By injecting 0.5 ml of raw garlic juice into ligated duodenum of rats, injury on duodenal mucosa lining followed 2 h exposure, with severe damage including ulcers and bleeding resulting after 24 h [153]. Damage to stomach and intestine may account for decrease of body weight by aqueous extract of garlic (300 or 600 mg/kg/d for 21 d) and garlic oil (200 mg/kg, thrice/wk for 6 wk) in rats [36, 154]. In a chronic toxicity test, however, no differences in body weight gain and in the urinary, haematological, serological, and histological examinations were observed in Wistar rats given garlic extract at doses of 2 g/kg for 5 time/wk for 6 mo [155]. These inconsistencies require more careful experimental design to clarify whether garlic displays adverse effect on gastrointestinal tract and growth, for instance, differences in garlic species and garlic preparations and also dosage tested merit consideration.
Over the past decades, the allergenic potential of garlic is well recognized. Case reports of allergic reactions―e.g., contact dermatitis, asthma, urticarial, pemphigus anaphylaxis―are known to associate with garlic use [156]. Allergic contact dermatitis to garlic was initially reported in 1950; to date, most cases of it appear in chefs and housewives in frequent contact with garlic [157-161]. Among Fernández-Vozmediano, et al.’s thirteen curry chefs, four tested DADS-positive, all showing dermatitis of the non-dominant hand, with hyperkeratosis and fissuring of the thumb, index, and middle finger [162]. Allergy on hands in the case of a 58-year-old male taking garlic to treat hyperlipidemia also relates to use of garlic tablets [163]. Based on such evidence, garlic is classified as Type I allergen [159], with allergens
identified as diallyl disulfide, allylpropyl disulfide, allylmercaptan, and allicin [164].
Conclusions
Past decades have seen myriad studies, especially in vitro and animal types, addressing protection of garlic against cardiovascular disease and cancer. This protection can arise from its diversified biological activities: enhanced antioxidant defense, lower blood lipids, inhibited blood aggregation, enhanced cancer cell cycle arrest/apoptosis, inhibited invasion and/or metastasis, modulating drug metabolism and/or immune response. Still, results observed in human clinical and intervention studies are inconsistent. Risk of garlic interacting with drugs attracts increasing interest, especially for the elderly and those with chronic disease. To understand actual health benefits and impact of garlic, further experiments are warranted.
Table 1. Garlic supplementation on human cardiovascular disorders.
Preparation Subjects/Dose Effect Reference
Garlic powder 51 CVD patients, 300
mg/day, 12-mo
↓ total cholesterol and LDL-C [165]
42 mild hypercholesterolemic men, 600 mg/d, 12-wk
↓ total cholesterol and LDL-C ↑ HDL-C
[22] 90 normollipidemic and
overweight adults, 2.1g/d, 12-wk
No changes in blood total cholesterol, LDL-C, and TG
[24]
Raw garlic 30 hypercholesterolemic
adults, 5 g/d, 42-d
↓ total cholesterol and TG ↑ HDL-C
[21]
Garlic extract 23 hypercholeterolemic adults
(13 with hypertension), ~10 g/d, 4-mo
↓ total cholesterol, LDL-C, and TG ↑ HDL ↓ SBP and DBP [166] Aged garlic extract
11 healthy adults, methionine-induced hyperhomocysteinemia, 4 mL/d, 6-wk ↑ NO and endothelium-derived hyperpolarizing factor [167] 65 intermediate CVD risk patients, 250 mg/d
co-administered with B vitamins (B12, B6, and folic acid) and
L-arginine, 12-mo
↓ total cholesterol, LDL-C, and homocysteine
↑ HDL
[168]
Garlic oil 20 hypertensive patients, 250
mg/d, 2-mo ↓ SBP, DBP, and oxLDL [13] Oil-macerated garlic 70 hypertensive adults, 1620 mg/d, 30-d
↓ total cholesterol, LDL-C, and TG
[23]
DBP, diastolic blood pressure; HDL-C, high density lipoprotein cholesterol; LDL-C, low density lipoprotein cholesterol; oxLDL, oxidized LDL; SHP, systolic blood pressure; TG, triglycerides.
Table 2. Mechanisms of garlic anti-cancer actions 1. Induce apoptosis/arrest cell cycle
2. Block invasion/metastasis 3. Suppress cell proliferation 4. Inhibit carcinogen activation 5. Enhance antioxidation
6. Decrease histone deacetylase activity 7. Interrupt tubulin polymerization 8. Change proteasome activity
Table 3. Garlic and drug interactions
Preparation Subjects/Dose Effect Reference
Garlic extract 12 health males, 2000 mg/d (3.71 mg allicin/ tablet), 2-wk
No changes on pharmacokinectics of warfarin
[150]
Garlic extract 10 breast cancer women, 600 mg/d (3.6 mg allicin/ tablet), 17-d No changes on pharmacokinectics of docetaxel [151]
Garlic extract 10 healthy male, 600 mg/d (12 mg
γ-glutamyl-cysteine/4.8 mg alliin), 21-d
↑ duodenal p-glycoprotein level, no change in CYP 3A4 expression no changes in simvastatin, pravastatin, and saquinavir bioavailability
Literature cited
1. Amagase H, Petesch BL, Matsuura H, Kasuga S, Itakura Y. Intake of garlic and its bioactive components. J Nutr. 2001;131:955S-62S.
2. Iciek M, Kwiecien I, Wlodek L. Biological properties of garlic and garlic-derived organosulfur compounds. Environ Mol Mutagen. 2009;50:247-65.
3. Block E, Ahmad S, Jain MK, Crecely RW, Apitz-Castro R, Cruz MR. The chemistry of alkyl thiosulfate esters. 8. (E,Z)-Ajoene: a potent antithrombotic agent from garlic. J Am Chem Soc. 1984;106:8295-6.
4. Kodera Y, Suzuki A, Imada O, Kasuga S, Sumioka I, Kanezawa A, et al. Physical, chemical, and biological properties of s-allylcysteine, an amino acid derived from garlic. J Agric Food Chem. 2002;50:622-32.
5. Gorinstein S, Drzewiecki J, Leontowicz H, Leontowicz M, Najman K, Jastrzebski Z, et al. Comparison of the bioactive compounds and antioxidant potentials of fresh and cooked Polish, Ukrainian, and Israeli garlic. J Agric Food Chem. 2005;53:2726-32.
6. Gorinstein S, Leontowicz H, Leontowicz M, Namiesnik J, Najman K, Drzewiecki J, et al. Comparison of the main bioactive compounds and antioxidant activities in garlic and white and red onions after treatment protocols. J Agric Food Chem. 2008;56:4418-26.
7. Tsiaganis MC, Laskari K, Melissari E. Fatty acid composition of Allium species lipids. J Food Compost Anal. 2006;19:620-7.
8. Yu TH, Wu CM, Liou YC. Volatile components from garlic. J Agric Food Chem. 1989;725-30.
9. Morihara N, Ushijima M, Kashimoto N, Sumioka I, Nishihama T, Hayama M, et al. Aged garlic extract ameliorates physical fatigue. Biol Pharm Bull. 2006;29:962-6.
10. Anderson RN. Deaths: leading causes for 1999. Natl Vital Stat Rep. 2001;49:1-87. 11. Ross R. Atherosclerosis--an inflammatory disease. N Engl J Med. 1999;340:115-26. 12. Zmijewski JW, Moellering DR, Le Goffe C, Landar A, Ramachandran A, Darley-Usmar VM. Oxidized LDL induces mitochondrially associated reactive oxygen/nitrogen species formation in endothelial cells. Am J Physiol Heart Circ Physiol. 2005;289:H852-61.
13. Dhawan V, Jain S. Effect of garlic supplementation on oxidized low density lipoproteins and lipid peroxidation in patients of essential hypertension. Mol Cell Biochem. 2004;266:109-15.
14. Keaney JF, Jr. Atherosclerosis: from lesion formation to plaque activation and endothelial dysfunction. Mol Aspects Med. 2000;21:99-166.
15. Gorinstein S, Jastrzebski Z, Namiesnik J, Leontowicz H, Leontowicz M, Trakhtenberg S. The atherosclerotic heart disease and protecting properties of garlic: contemporary data. Mol Nutr Food Res. 2007;51:1365-81.
16. Adler AJ, Holub BJ. Effect of garlic and fish-oil supplementation on serum lipid and lipoprotein concentrations in hypercholesterolemic men. Am J Clin Nutr. 1997;65:445-50.
17. Jain AK, Vargas R, Gotzkowsky S, McMahon FG. Can garlic reduce levels of serum lipids? A controlled clinical study. Am J Med. 1993;94:632-5.
18. Sangeetha T, Darlin Quine S. Preventive effect of S-allyl cysteine sulfoxide (alliin) on cardiac marker enzymes and lipids in isoproterenol-induced myocardial injury. J Pharm Pharmacol. 2006;58:617-23.
19. Sobenin IA, Pryanishnikov VV, Kunnova LM, Rabinovich YA, Martirosyan DM, Orekhov AN. The effects of time-released garlic powder tablets on multifunctional cardiovascular risk in patients with coronary artery disease. Lipids Health Dis. 2010;9:119. 20. Durak I, Kavutcu M, Aytac B, Avci A, Devrim E, Ozbek H, et al. Effects of garlic extract consumption on blood lipid and oxidant/antioxidant parameters in humans with high blood cholesterol. J Nutr Biochem. 2004;15:373-7.
21. Mahmoodi M, Islami MR, Asadi Karam GR, Khaksari M, Sahebghadam Lotfi A, Hajizadeh MR, et al. Study of the effects of raw garlic consumption on the level of lipids and other blood biochemical factors in hyperlipidemic individuals. Pak J Pharm Sci. 2006;19:295-8.
22. Sobenin IA, Andrianova IV, Demidova ON, Gorchakova T, Orekhov AN. Lipid-lowering effects of time-released garlic powder tablets in double-blinded placebo-controlled randomized study. J Atheroscler Thromb. 2008;15:334-8.
23. Duda G, Suliburska J, Pupek-Musialik D. Effects of short-term garlic supplementation on lipid metabolism and antioxidant status in hypertensive adults. Pharmacol Rep. 2008;60:163-70.
24. van Doorn MB, Espirito Santo SM, Meijer P, Kamerling IM, Schoemaker RC, Dirsch V, et al. Effect of garlic powder on C-reactive protein and plasma lipids in overweight and smoking subjects. Am J Clin Nutr. 2006;84:1324-9.
25. Reinhart KM, Talati R, White CM, Coleman CI. The impact of garlic on lipid parameters: a systematic review and meta-analysis. Nutr Res Rev. 2009;22:39-48.
26. Silagy C, Neil A. Garlic as a lipid lowering agent--a meta-analysis. J R Coll Physicians Lond. 1994;28:39-45.
27. Warshafsky S, Kamer RS, Sivak SL. Effect of garlic on total serum cholesterol. A meta-analysis. Ann Intern Med. 1993;119:599-605.
28. Thomson M, Mustafa T, Ali M. Thromboxane-B(2) levels in serum of rabbits receiving a single intravenous dose of aqueous extract of garlic and onion. Prostaglandins Leukot Essent Fatty Acids. 2000;63:217-21.
29. Ali M. Mechanism by which garlic (Allium sativum) inhibits cyclooxygenase activity. Effect of raw versus boiled garlic extract on the synthesis of prostanoids. Prostaglandins Leukot Essent Fatty Acids. 1995;53:397-400.
30. Ali M, Thomson M. Consumption of a garlic clove a day could be beneficial in preventing thrombosis. Prostaglandins Leukot Essent Fatty Acids. 1995;53:211-2.
31. Cavagnaro PF, Camargo A, Galmarini CR, Simon PW. Effect of cooking on garlic (Allium sativum L.) antiplatelet activity and thiosulfinates content. J Agric Food Chem. 2007;55:1280-8.
32. Rahman K. Effects of garlic on platelet biochemistry and physiology. Mol Nutr Food Res. 2007;51:1335-44.
33. Borek C. Garlic reduces dementia and heart-disease risk. J Nutr. 2006;136:810S-2S. 34. Youn HS, Lim HJ, Lee HJ, Hwang D, Yang M, Jeon R, et al. Garlic (Allium sativum) extract inhibits lipopolysaccharide-induced Toll-like receptor 4 dimerization. Biosci Biotechnol Biochem. 2008;72:368-75.
35. Ban JO, Oh JH, Kim TM, Kim DJ, Jeong HS, Han SB, et al. Anti-inflammatory and arthritic effects of thiacremonone, a novel sulfur compound isolated from garlic via inhibition of NF-kappaB. Arthritis Res Ther. 2009;11:R145.
36. Liu C-T, Chen H-W, Sheen L-Y, Kung Y-L, Chen PC-H, Lii C-K. Effect of Garlic Oil on Hepatic Arachidonic Acid Content and Immune Response in Rats. J Agric Food Chem. 1998;46:4642-7.
37. Zamani A, Vahidinia A, Ghannad MS. The effect of garlic consumption on Th1/Th2 cytokines in phytohemagglutinin (PHA) activated rat spleen lymphocytes. Phytother Res. 2009;23:579-81.
38. Keophiphath M, Priem F, Jacquemond-Collet I, Clement K, Lacasa D. 1,2-vinyldithiin from garlic inhibits differentiation and inflammation of human preadipocytes. J Nutr. 2009;139:2055-60.
39. Lee HS, Lee CH, Tsai HC, Salter DM. Inhibition of cyclooxygenase 2 expression by diallyl sulfide on joint inflammation induced by urate crystal and IL-1beta. Osteoarthritis Cartilage. 2009;17:91-9.
40. Lee EN, Choi YW, Kim HK, Park JK, Kim HJ, Kim MJ, et al. Chloroform extract of aged black garlic attenuates Talpha-induced ROS generation, VCAM-1 expression, NF-kappaB activation and adhesiveness for monocytes in human umbilical vein endothelial cells. Phytother Res. 2011;25:92-100.
41. Rassoul F, Salvetter J, Reissig D, Schneider W, Thiery J, Richter V. The influence of garlic (Allium sativum) extract on interleukin 1alpha-induced expression of endothelial intercellular adhesion molecule-1 and vascular cell adhesion molecule-1. Phytomedicine. 2006;13:230-5.
42. Lei YP, Chen HW, Sheen LY, Lii CK. Diallyl disulfide and diallyl trisulfide suppress oxidized LDL-induced vascular cell adhesion molecule and E-selectin expression through protein kinase A- and B-dependent signaling pathways. J Nutr. 2008;138:996-1003.
43. Sun X, Ku DD. Allicin in garlic protects against coronary endothelial dysfunction and right heart hypertrophy in pulmonary hypertensive rats. Am J Physiol Heart Circ Physiol. 2006;291:H2431-8.
44. Lei YP, Liu CT, Sheen LY, Chen HW, Lii CK. Diallyl disulfide and diallyl trisulfide protect endothelial nitric oxide synthase against damage by oxidized low-density lipoprotein. Mol Nutr Food Res. 2010;54 Suppl 1:S42-52.
45. Kim JM, Chang N, Kim WK, Chun HS. Dietary S-allyl-L-cysteine reduces mortality with decreased incidence of stroke and behavioral changes in stroke-prone spontaneously hypertensive rats. Biosci Biotechnol Biochem. 2006;70:1969-71.
46. Chuah SC, Moore PK, Zhu YZ. S-allylcysteine mediates cardioprotection in an acute myocardial infarction rat model via a hydrogen sulfide-mediated pathway. Am J Physiol Heart Circ Physiol. 2007;293:H2693-701.
47. Ou HC, Tzang BS, Chang MH, Liu CT, Liu HW, Lii CK, et al. Cardiac contractile dysfunction and apoptosis in streptozotocin-induced diabetic rats are ameliorated by garlic oil supplementation. J Agric Food Chem. 2010;58:10347-55.
48. Zahid Ashraf M, Hussain ME, Fahim M. Antiatherosclerotic effects of dietary supplementations of garlic and turmeric: Restoration of endothelial function in rats. Life Sci. 2005;77:837-57.
49. Williams MJ, Sutherland WH, McCormick MP, Yeoman DJ, de Jong SA. Aged garlic extract improves endothelial function in men with coronary artery disease. Phytother Res. 2005;19:314-9.
50. Setiawan VW, Yu GP, Lu QY, Lu ML, Yu SZ, Mu L, et al. Allium vegetables and stomach cancer risk in China. Asian Pac J Cancer Prev. 2005;6:387-95.
51. Millen AE, Subar AF, Graubard BI, Peters U, Hayes RB, Weissfeld JL, et al. Fruit and vegetable intake and prevalence of colorectal adenoma in a cancer screening trial. Am J Clin Nutr. 2007;86:1754-64.
52. Fleischauer AT, Arab L. Garlic and cancer: a critical review of the epidemiologic literature. J Nutr. 2001;131:1032S-40S.
53. Kim JY, Kwon O. Garlic intake and cancer risk: an analysis using the Food and Drug Administration's evidence-based review system for the scientific evaluation of health claims. Am J Clin Nutr. 2009;89:257-64.
54. Tanaka S, Haruma K, Yoshihara M, Kajiyama G, Kira K, Amagase H, et al. Aged garlic extract has potential suppressive effect on colorectal adenomas in humans. J Nutr. 2006;136:821S-6S.
55. Li H, Li HQ, Wang Y, Xu HX, Fan WT, Wang ML, et al. An intervention study to prevent gastric cancer by micro-selenium and large dose of allitridum. Chin Med J (Engl). 2004;117:1155-60.
56. Samaranayake MD, Wickramasinghe SM, Angunawela P, Jayasekera S, Iwai S, Fukushima S. Inhibition of chemically induced liver carcinogenesis in Wistar rats by garlic (Allium sativum). Phytother Res. 2000;14:564-7.
additives on aflatoxin-induced mutagenicity and hepatocarcinogenicity. Cancer Lett. 1997;115:129-33.
58. Agarwal MK, Iqbal M, Athar M. Garlic oil ameliorates ferric nitrilotriacetate (Fe-NTA)-induced damage and tumor promotion: implications for cancer prevention. Food Chem Toxicol. 2007;45:1634-40.
59. Song K, Milner JA. Heating garlic inhibits its ability to suppress 7, 12-dimethylbenz(a)anthracene-induced DNA adduct formation in rat mammary tissue. J Nutr. 1999;129:657-61.
60. Arora A, Kalra N, Shukla Y. Regulation of p21/ras protein expression by diallyl sulfide in DMBA induced neoplastic changes in mouse skin. Cancer Lett. 2006;242:28-36.
61. Kalra N, Arora A, Shukla Y. Involvement of multiple signaling pathways in diallyl sulfide mediated apoptosis in mouse skin tumors. Asian Pac J Cancer Prev. 2006;7:556-62. 62. Shrotriya S, Kundu JK, Na HK, Surh YJ. Diallyl trisulfide inhibits phorbol ester-induced tumor promotion, activation of AP-1, and expression of COX-2 in mouse skin by blocking JNK and Akt signaling. Cancer Res. 2010;70:1932-40.
63. Surh YJ, Kim SG, Liem A, Lee JW, Miller JA. Inhibitory effects of isopropyl-2-(1,3-dithietane-2-ylidene)-2- [N-(4-methylthiazol-2-yl)carbamoyl]acetate (YH439) on benzo[a]pyrene-induced skin carcinogenesis and micronucleated reticulocyte formation in mice. Mutat Res. 1999;423:149-53.
64. Xiao D, Lew KL, Kim YA, Zeng Y, Hahm ER, Dhir R, et al. Diallyl trisulfide suppresses growth of PC-3 human prostate cancer xenograft in vivo in association with Bax and Bak induction. Clin Cancer Res. 2006;12:6836-43.
65. Taylor P, Noriega R, Farah C, Abad MJ, Arsenak M, Apitz R. Ajoene inhibits both primary tumor growth and metastasis of B16/BL6 melanoma cells in C57BL/6 mice. Cancer Lett. 2006;239:298-304.
66. Tang FY, Chiang EP, Pai MH. Consumption of S-allylcysteine inhibits the growth of human non-small-cell lung carcinoma in a mouse xenograft model. J Agric Food Chem. 2010; 58:1156-64.
67. Shin HA, Cha YY, Park MS, Kim JM, Lim YC. Diallyl sulfide induces growth inhibition and apoptosis of anaplastic thyroid cancer cells by mitochondrial signaling pathway. Oral Oncol. 2010;46:e15-8.
68. Sriram N, Kalayarasan S, Ashokkumar P, Sureshkumar A, Sudhandiran G. Diallyl sulfide induces apoptosis in Colo 320 DM human colon cancer cells: involvement of caspase-3, NF-kappaB, and ERK-2. Mol Cell Biochem. 2008;311:157-65.
69. Wu CC, Chung JG, Tsai SJ, Yang JH, Sheen LY. Differential effects of allyl sulfides from garlic essential oil on cell cycle regulation in human liver tumor cells. Food Chem Toxicol. 2004;42:1937-47.
cancer cells through induction of DNA damage mediated G2/M arrest and apoptosis. J Agric Food Chem. 2010;58:7096-103.
71. Wu XJ, Hu Y, Lamy E, Mersch-Sundermann V. Apoptosis induction in human lung adenocarcinoma cells by oil-soluble allyl sulfides: triggers, pathways, and modulators. Environ Mol Mutagen. 2009;50:266-75.
72. Das A, Banik NL, Ray SK. Garlic compounds generate reactive oxygen species leading to activation of stress kinases and cysteine proteases for apoptosis in human glioblastoma T98G and U87MG cells. Cancer. 2007;110:1083-95.
73. Gunadharini DN, Arunkumar A, Krishnamoorthy G, Muthuvel R, Vijayababu MR, Kanagaraj P, et al. Antiproliferative effect of diallyl disulfide (DADS) on prostate cancer cell line LNCaP. Cell Biochem Funct. 2006;24:407-12.
74. Arunkumar A, Vijayababu MR, Srinivasan N, Aruldhas MM, Arunakaran J. Garlic compound, diallyl disulfide induces cell cycle arrest in prostate cancer cell line PC-3. Mol Cell Biochem. 2006;288:107-13.
75. Karmakar S, Banik NL, Patel SJ, Ray SK. Garlic compounds induced calpain and intrinsic caspase cascade for apoptosis in human malignant neuroblastoma SH-SY5Y cells. Apoptosis. 2007;12:671-84.
76. Li N, Guo R, Li W, Shao J, Li S, Zhao K, et al. A proteomic investigation into a human gastric cancer cell line BGC823 treated with diallyl trisulfide. Carcinogenesis. 2006;27:1222-31.
77. Wang YB, Qin J, Zheng XY, Bai Y, Yang K, Xie LP. Diallyl trisulfide induces Bcl-2 and caspase-3-dependent apoptosis via downregulation of Akt phosphorylation in human T24 bladder cancer cells. Phytomedicine. 2010;17:363-8.
78. Druesne N, Pagniez A, Mayeur C, Thomas M, Cherbuy C, Duee PH, et al. Diallyl disulfide (DADS) increases histone acetylation and p21(waf1/cip1) expression in human colon tumor cell lines. Carcinogenesis. 2004;25:1227-36.
79. Lei XY, Yao SQ, Zu XY, Huang ZX, Liu LJ, Zhong M, et al. Apoptosis induced by diallyl disulfide in human breast cancer cell line MCF-7. Acta Pharmacol Sin. 2008;29:1233-9.
80. Xiao D, Choi S, Johnson DE, Vogel VG, Johnson CS, Trump DL, et al. Diallyl trisulfide-induced apoptosis in human prostate cancer cells involves c-Jun N-terminal kinase and extracellular-signal regulated kinase-mediated phosphorylation of Bcl-2. Oncogene. 2004;23:5594-606.
81. Xiao D, Singh SV. Diallyl trisulfide, a constituent of processed garlic, inactivates Akt to trigger mitochondrial translocation of BAD and caspase-mediated apoptosis in human prostate cancer cells. Carcinogenesis. 2006;27:533-40.
82. Xiao D, Herman-Antosiewicz A, Antosiewicz J, Xiao H, Brisson M, Lazo JS, et al. Diallyl trisulfide-induced G(2)-M phase cell cycle arrest in human prostate cancer cells is
caused by reactive oxygen species-dependent destruction and hyperphosphorylation of Cdc25C. Oncogene. 2005;24:6256-68.
83. Herman-Antosiewicz A, Kim YA, Kim SH, Xiao D, Singh SV. Diallyl trisulfide-induced G2/M phase cell cycle arrest in DU145 cells is associated with delayed nuclear translocation of cyclin-dependent kinase 1. Pharm Res. 2010;27:1072-9.
84. Hosono T, Fukao T, Ogihara J, Ito Y, Shiba H, Seki T, et al. Diallyl trisulfide suppresses the proliferation and induces apoptosis of human colon cancer cells through oxidative modification of beta-tubulin. J Biol Chem. 2005;280:41487-93.
85. Xiao D, Pinto JT, Soh JW, Deguchi A, Gundersen GG, Palazzo AF, et al. Induction of apoptosis by the garlic-derived compound S-allylmercaptocysteine (SAMC) is associated with microtubule depolymerization and c-Jun NH(2)-terminal kinase 1 activation. Cancer Res. 2003;63:6825-37.
86. Pratheeshkumar P, Thejass P, Kutan G. Diallyl disulfide induces caspase-dependent apoptosis via mitochondria-mediated intrinsic pathway in B16F-10 melanoma cells by up-regulating p53, caspase-3 and down-up-regulating pro-inflammatory cytokines and nuclear factor-kappabeta-mediated Bcl-2 activation. J Environ Pathol Toxicol Oncol. 2010;29:113-25. 87. Zhang W, Ha M, Gong Y, Xu Y, Dong N, Yuan Y. Allicin induces apoptosis in gastric cancer cells through activation of both extrinsic and intrinsic pathways. Oncol Rep. 2010;24:1585-92.
88. Druesne-Pecollo N, Chaumontet C, Pagniez A, Vaugelade P, Bruneau A, Thomas M, et al. In vivo treatment by diallyl disulfide increases histone acetylation in rat colonocytes. Biochem Biophys Res Commun. 2007;354:140-7.
89. Lea MA, Rasheed M, Randolph VM, Khan F, Shareef A, desBordes C. Induction of histone acetylation and inhibition of growth of mouse erythroleukemia cells by S-allylmercaptocysteine. Nutr Cancer. 2002;43:90-102.
90. Nian H, Delage B, Pinto JT, Dashwood RH. Allyl mercaptan, a garlic-derived organosulfur compound, inhibits histone deacetylase and enhances Sp3 binding on the P21WAF1 promoter. Carcinogenesis. 2008;29:1816-24.
91. Singh SV, Powolny AA, Stan SD, Xiao D, Arlotti JA, Warin R, et al. Garlic constituent diallyl trisulfide prevents development of poorly differentiated prostate cancer and pulmonary metastasis multiplicity in TRAMP mice. Cancer Res. 2008;68:9503-11.
92. Shin DY, Kim GY, Kim JI, Yoon MK, Kwon TK, Lee SJ, et al. Anti-invasive activity of diallyl disulfide through tightening of tight junctions and inhibition of matrix metalloproteinase activities in LNCaP prostate cancer cells. Toxicol In Vitro. 2010;24:1569-76.
93. Chu Q, Ling MT, Feng H, Cheung HW, Tsao SW, Wang X, et al. A novel anticancer effect of garlic derivatives: inhibition of cancer cell invasion through restoration of E-cadherin expression. Carcinogenesis. 2006;27:2180-9.
94. Tang FY, Chiang EP, Chung JG, Lee HZ, Hsu CY. S-allylcysteine modulates the expression of E-cadherin and inhibits the malignant progression of human oral cancer. J Nutr Biochem. 2009;20:1013-20.
95. Matsuura N, Miyamae Y, Yamane K, Nagao Y, Hamada Y, Kawaguchi N, et al. Aged garlic extract inhibits angiogenesis and proliferation of colorectal carcinoma cells. J Nutr. 2006;136:842S-6S.
96. Xu B, Monsarrat B, Gairin JE, Girbal-Neuhauser E. Effect of ajoene, a natural antitumor small molecule, on human 20S proteasome activity in vitro and in human leukemic HL60 cells. Fundam Clin Pharmacol. 2004;18:171-80.
97. Guengerich FP, Shimada T. Oxidation of toxic and carcinogenic chemicals by human cytochrome P-450 enzymes. Chem Res Toxicol. 1991;4:391-407.
98. Guyonnet D, Belloir C, Suschetet M, Siess MH, Le Bon AM. Mechanisms of protection against aflatoxin B(1) genotoxicity in rats treated by organosulfur compounds from garlic. Carcinogenesis. 2002;23:1335-41.
99. Hu X, Benson PJ, Srivastava SK, Mack LM, Xia H, Gupta V, et al. Glutathione S-transferases of female A/J mouse liver and forestomach and their differential induction by anti-carcinogenic organosulfides from garlic. Arch Biochem Biophys. 1996;336:199-214. 100. Belloir C, Singh V, Daurat C, Siess MH, Le Bon AM. Protective effects of garlic sulfur compounds against DNA damage induced by direct- and indirect-acting genotoxic agents in HepG2 cells. Food Chem Toxicol. 2006;44:827-34.
101. Le Bon AM, Vernevaut MF, Guenot L, Kahane R, Auger J, Arnault I, et al. Effects of garlic powders with varying alliin contents on hepatic drug metabolizing enzymes in rats. J Agric Food Chem. 2003;51:7617-23.
102. Park KA, Kweon S, Choi H. Anticarcinogenic effect and modification of cytochrome P450 2E1 by dietary garlic powder in diethylnitrosamine-initiated rat hepatocarcinogenesis. J Biochem Mol Biol. 2002;35:615-22.
103. Taubert D, Glockner R, Muller D, Schomig E. The garlic ingredient diallyl sulfide inhibits cytochrome P450 2E1 dependent bioactivation of acrylamide to glycidamide. Toxicol Lett. 2006;164:1-5.
104. Chen HW, Tsai CW, Yang JJ, Liu CT, Kuo WW, Lii CK. The combined effects of garlic oil and fish oil on the hepatic antioxidant and drug-metabolizing enzymes of rats. Br J Nutr. 2003;89:189-200.
105. Davenport DM, Wargovich MJ. Modulation of cytochrome P450 enzymes by organosulfur compounds from garlic. Food Chem Toxicol. 2005;43:1753-62.
106. Chen HW, Yang JJ, Tsai CW, Wu JJ, Sheen LY, Ou CC, et al. Dietary fat and garlic oil independently regulate hepatic cytochrome p(450) 2B1 and the placental form of glutathione S-transferase expression in rats. J Nutr. 2001;131:1438-43.
oil and its three major organosulfur components on the hepatic detoxification system in rats. J Agric Food Chem. 2002;50:378-83.
108. Foster BC, Foster MS, Vandenhoek S, Krantis A, Budzinski JW, Arnason JT, et al. An
in vitro evaluation of human cytochrome P450 3A4 and P-glycoprotein inhibition by garlic. J
Pharm Pharm Sci. 2001;4:176-84.
109. Brady JF, Ishizaki H, Fukuto JM, Lin MC, Fadel A, Gapac JM, et al. Inhibition of cytochrome P-450 2E1 by diallyl sulfide and its metabolites. Chem Res Toxicol. 1991;4:642-7.
110. Shaik IH, George JM, Thekkumkara TJ, Mehvar R. Protective effects of diallyl sulfide, a garlic constituent, on the warm hepatic ischemia-reperfusion injury in a rat model. Pharm Res. 2008;25:2231-42.
111. Yang CS, Chhabra SK, Hong JY, Smith TJ. Mechanisms of inhibition of chemical toxicity and carcinogenesis by diallyl sulfide (DAS) and related compounds from garlic. J Nutr. 2001;131:1041S-5S.
112. Jana S, Mandlekar S. Role of phase II drug metabolizing enzymes in cancer chemoprevention. Curr Drug Metab. 2009;10:595-616.
113. Tan XL, Spivack SD. Dietary chemoprevention strategies for induction of phase II xenobiotic-metabolizing enzymes in lung carcinogenesis: A review. Lung Cancer. 2009;65:129-37.
114. Henderson CJ, Smith AG, Ure J, Brown K, Bacon EJ, Wolf CR. Increased skin tumorigenesis in mice lacking pi class glutathione S-transferases. Proc Natl Acad Sci USA. 1998;95:5275-80.
115. Sparnins VL, Barany G, Wattenberg LW. Effects of organosulfur compounds from garlic and onions on benzo[a]pyrene-induced neoplasia and glutathione S-transferase activity in the mouse. Carcinogenesis. 1988;9:131-4.
116. Balasenthil S, Nagini S. Protective effects of S-allylcysteine on hepatic glutathione and glutathione-dependent enzymes during hamster cheek pouch carcinogenesis. J Biochem Mol Biol Biophys. 2002;6:13-6.
117. Na HK, Surh YJ. Modulation of Nrf2-mediated antioxidant and detoxifying enzyme induction by the green tea polyphenol EGCG. Food Chem Toxicol. 2008;46:1271-8.
118. Chen C, Pung D, Leong V, Hebbar V, Shen G, Nair S, et al. Induction of detoxifying enzymes by garlic organosulfur compounds through transcription factor Nrf2: effect of chemical structure and stress signals. Free Radic Biol Med. 2004;37:1578-90.
119. Fisher CD, Augustine LM, Maher JM, Nelson DM, Slitt AL, Klaassen CD, et al. Induction of drug-metabolizing enzymes by garlic and allyl sulfide compounds via activation of constitutive androstane receptor and nuclear factor E2-related factor 2. Drug Metab Dispos. 2007;35:995-1000.