Distribution of the Freshwater Prawns (Macrobrachium Bate,
1868) in Taiwan in Relation to Their Biogeographic Origins
台灣島上淡水沼蝦屬分布與其生物地理起源之關係
Rung-Tsung Chen
1, 2, Shi-Tsang Chang
1, *, Ming-Fon Yeh
1, Hung-Po Chen
1,
Te-Hung Chen
1, Chu-Fa Tsai
1and Wann-Nian Tzeng
2陳榮宗
1, 2張世倉
1, *葉明峰
1陳宏柏
1陳德鴻
1蔡住發
1曾萬年
21Endemic Species Research Institute, Jiji, Nantou, Taiwan 2Institute of Fisheries Science, National Taiwan University, Taipei, Taiwan
1行政院農業委員會特有生物研究保育中心 南投縣集集鎮民生東路 1 號 2國立台灣大學漁業科學研究所 台北市羅斯福路四段 1 號
* Corresponding author: stchang@tesri.gov.tw
* 通訊作者:stchang@tesri.gov.tw
Abstract
Using data collected from the freshwater prawn inventory survey of 2000-2008 and historical data obtained from literature prior to 2000, distribution patterns of 15 species of the Macrobrachium prawns in Taiwan were examined. They were able to divide into two groups: the east-coast group and the island-wide group. The east-coast group contained 11 amphidromous species: M. equidens, M. mammillodactylus,
M. latidactylus, M. gracilirostre, M. lepidactyloides, M. lar, M. placidulum, M. jaroense, M. esculentum, M. latimanus and M. australe. They were found in the east coast, some of them in its adjacent northern
and southern regions. They were the Southeast Asia origins and dispersed to Taiwan mainly through the Philippines, and showed close association with the Kuroshio Current in the dispersion. The island-wide group contained 4 species: M. japonicum, M. formosense, M. asperulum and M. nipponense. The former species was postulated to have evolved within the island of Taiwan, while the latter three species were originated from the China mainland. They were common in streams around the island. M. japonicum and
M. formosense that have been considered to be amphidromous showed the distribution patterns fairly
similar to that of non-obligatory amphidromous M. nipponense and even to that of landlocked M.
asperulum, rather than to those of the amphidromous species of the east-coast group. The evidences
suggest that these two species might not be the amphidromous but the non-obligatory amphidromous, but a further study is needed for the confirmation. The distribution patterns of the Macrobrachium prawns in Taiwan are resulted from their life cycle modes, adaptability to local environments at present and biogeographic origins and dispersal routes in the past evolutionary history.
摘
要
台灣 15 種淡水沼蝦在島上的分布範圍,可區分為東岸型及全島型等 2 種分布類型。東岸型 均為兩側洄游性物種,分別為 Macrobrachium equidens、M. mammillodactylus、M. latidactylus、M.
gracilirostre、M. lepidactyloides、M. lar、M. placidulum、M. jaroense、M. esculentum、M. latimanus
及 M. australe 等 11 種,牠們分布在台灣的東部,部分物種亦分布在鄰接的北部及南部地區,其中
M. australe 則另分布在台灣的西北部。這些物種皆來自東南亞島嶼,並經由菲律賓來到台灣,其在
島上的分布及擴散來台的途徑與黑潮關係密切。全島型包括 4 種,分別為 M. japonicum、M.
formosense、M. asperulum 及 M. nipponense,前者在台灣種化形成,後 3 種則源自於中國大陸。
牠們廣泛分布於台灣的溪流,其中 M. japonicum 及 M. formosense 為兩側洄游性,其在島上的分 布型態與兼具兩側洄游的 M. nipponense 及陸封性的 M. asperulum 極為相似,卻與同為兩側洄游性 的東岸型物種之分布極為不同,顯示 M. japonicum 及 M. formosense 或許並非全然為兩側洄游性, 而是如 M. nipponense 為兼具兩側洄游性的物種,此點需要進一步的研究證實。台灣淡水沼蝦在 島上的分布型態,是演化過程中物種地理起源、擴散來台途徑及其對環境適應能力等因素所造成。
Key words: biogeography, distribution pattern, Macrobrachium, Taiwan
關鍵詞:生物地理、分布型態、沼蝦屬、台灣
收件日期:98 年 2 月 6 日 接受日期:98 年 6 月 22 日
Received: February 6, 2009 Accepted: June 22, 2009
Introduction
Freshwater prawns (Macrobrachium Bate,
1868) of Taiwan are highly diverse, rich in amphidromous forms, but poor in endemic and land-locked forms (Hwang and Yu 1982; Shy and
Yu 1998; Cai and Jeng 2001; Cai and Shokita 2006; Chen et al. 2009). The fauna is a mixture of the species of various biogeographic origins and more similar to those of the Philippines Islands and the Ryukyu Islands than that of the China mainland (Chen et al. 2009).
In the previous study (Chen et al. 2009), we examined the distribution patterns of 15 known species of Macrobrachium of Taiwan in the Indo-Pacific region and their relationships based on the phylogenies reconstructed from the nuclear 28S rDNA gene. For the 13 amphidromous species, their biogeographic origins and dispersion routes to Taiwan were hypothesized to be: 1) in the eastern region of southeast Asia islands through the Philippines for M. australe, M. lar, M. latimanus,
M. gracilirostre, M. jaroense, M. esculentum, M. lepidactyloides and M. placidulum; 2) in the
western region of southeast Asia islands through the Philippines and/or China mainland for M.
equidens, M. latidactylus and M. mammillodactylus;
3) in China mainland for M. formosense, and 4) on the island of Taiwan for M. japonicum. For the remaining two species, M. asperulum, a land-locked prawn, and M. nipponense, a non-obligatory amphidromous prawn, were originated from China mainland.
This study continued the previous study (Chen et al. 2009) to examine the distribution of the Macrobrachium prawns on the island of Taiwan and to determine whether there are relationships between the distribution and their biogeographic origins and past dispersion routes.
Material and Methods
During the period from 2000 to 2008, we made an inventory survey of freshwater prawns
in inland waters of Taiwan. A 8-volt backpack electrofishing gear (Yeh et al. 2000), 30 cm x 10 cm baited shrimp traps (Chen et al. 2003), and various types of hand nets (Short 2000) were used. A total of 3,382 individuals belonging to 15 species of Macrobrachium were collected at 662 sites. An exception was M. shaoi that was extremely rare and found only in a tributary of Shuangchi River in the northern Taiwan (Cai and Jeng 2001) was not collected in this study. Most of the prawns were collected by the shrimp traps, but a lot of large individuals were captured by electrofishing. Most of the collections were fixed in 15% formalin water solution, preserved in 75% ethyl-alcohol water solution, and deposited at the Endemic Species Research Institute, Jiji, Nantou, Taiwan. The data were stored in Microsoft office Access 2003 for database setting. Also, distribution data of the prawns prior to 2000 were obtained from literatures (Hwang and Yu 1982; Shy 1994; Shy
et al. 1996) and compared to those from our
2000-2008 survey. The distribution map of each species of the prawns was constructed with SuperGIS version 2.2.
Results
Based on field data collected from our 2000-2008 survey and distribution data obtained from literature (Hwang and Yu 1982; Shy 1994; Shy et al. 1996), the distribution patterns of 15 species of the Macrobrachium prawns in Taiwan were plotted in Figs. 1-4. They were divided into two groups: 1) the east-coast group and 2) the island-wide group:
The east-coast group
species. They were distributed in the east coast with some in adjacent northern and/or southern regions and absent in the western region (Figs. 1-3). They corresponded to the species of three groups with different biogeographical origins and dispersion routes hypothesized by Chen et al. (2009). They were the Eastern Southeast Asia group (M. esculentum, M. gracilirostre, M. jaroense,
M. lepidactyloides and M. placidulum), the
Indo-West Pacific group (M. australe, M. lar and M.
latimanus), and the Western Southeast Asia group
(M. equidens, M. latidactylus and M.
mammillo-dactylus). The former two groups were found in
the Philippines but not in China mainland, and thus, they were postulated to originate from the eastern region of Southeast Asia islands and dispersed to Taiwan through the Philippines. The latter group was found in both the Philippines and China mainland and assumed to originate from the western region of Southeast Asia islands and dispersed to Taiwan through the Philippines and/ or China mainland (Chen et al. 2009).
For the Eastern Southeast Asia group, M.
esculentum, M. jaroense and M. placidulum were
found to be rare and occurred only in streams along the east coast (Fig. 1), whereas M. gracilirostre and M. lepidactyloides were common and found not only in the east coast but also in the southwestern region (Fig. 2A-B). The Indo-West Pacific group,
M. lar and M. australe were common and found
in the east coast, southwestern and northwestern region of the island (Fig. 2C-D), while M. latimanus was rare and occurred in streams of southwestern region and some in the east coast (Fig. 1). For the Western Southeast Asia group, M. equidens was common and occurred in the east coast and also in both southwestern and northwestern regions of the island, while M. latidactylus and M.
mammillodactylus were rare and occurred only
in the east coast (Fig. 3).
The island-wide group
The island-wide group contained four species:
M. asperulum, M. nipponens, M. formosense and M. japonicum. They occurred in streams around
the island (Fig. 4), differing from those of the east coast group (Fig. 1-3). The former three species are postulated to originate from the China mainland, while the latter species is an autochthonous prawn that was evolved within the island of Taiwan (Chen et al. 2009). The four species have been considered to have different life cycle modes: M.
asperulum as a landlocked prawn (Shokita 1977;
Lin et al. 1988; Shy 1994; Shy et al. 1996; Liu et
al. 2007; Mashiko and Shy 2008), M. nipponense
as a non-obligatory amphidromous prawn (Shy
et al. 1987; Shy 1994; Shy et al. 1996; Mashiko
and Shy 2008), and M. formosense and M. japonicum as the amphidromous prawns (Shy et al. 1990; Shy 1994; Shy et al. 1996; Suzuki and Kusamura 1997; Liu et al. 2007; Mashiko and Shy 2008).
Distributions of the freshwater prawns in Taiwan are closed related to their biogeographical origins and modes of their life cycles. The east coast group contained amphidromous prawns of the tropical Southeast Asia islands origins, whereas the island-wide group contained land-locked, non-obligatory amphidromous, and amphidromous prawns of the East Asia origins (Taiwan or China mainland).
Discussion
The east-coast group
Although 11 amphidromous prawns of the east-coast group came from three different
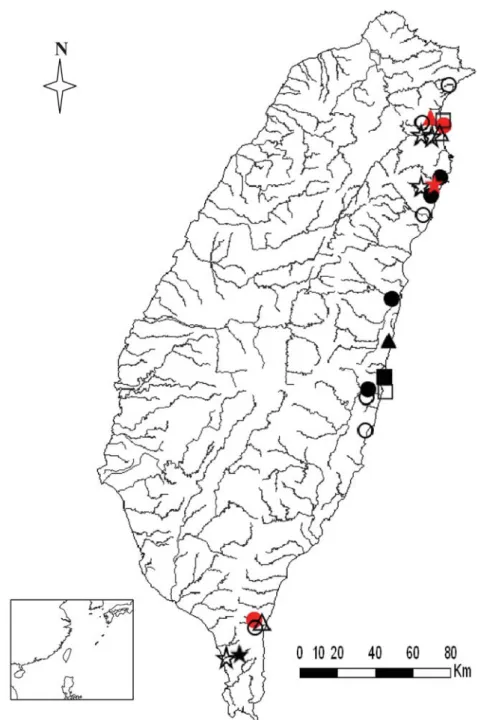
Fig. 1. Distributions of four rare species of the east coast group of Macrobrachium in Taiwan: M.
placidulum (solid circles, 2000-2008 survey; open circles, Shy 1994; red solid circles, Hwang and Yu
1982); M. jaroense (solid triangles, 2000-2008 survey; open triangles, Shy 1994; red solid triangles, Hwang and Yu 1982); M. esculentum (solid squares, 2000-2008 survey; open squares, Shy 1994); and
Fig. 2. Distributions of four common species of the east coast group of Macrobrachium in Taiwan: A.
M. gracilirostre; B. M. lepidactyloides; C. M. lar; D. M. australe (solid circles, 2000-2008 survey; red
Fig. 3. Distributions of common M. equidens (solid circles, 2000-2008 survey; open circles, Shy 1994;
red solid circles, Hwang and Yu 1982) and two rare species: M. latidactylus (solid triangles, 2000-2008 survey; open triangles, Shy 1994; red solid triangles, Hwang and Yu 1982); M. mammillodactylus (solid stars, 2000-2008 survey; open stars, Shy 1994) of the east-coast group in Taiwan.
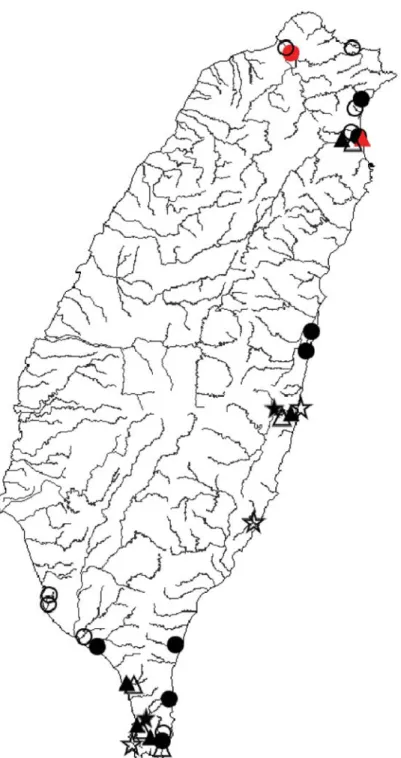
Fig. 4. Distributions of four species of the island-wide group of Macrobrachium in Taiwan: A. M.
japonicum; B. M. formosense; C. M. nipponense; D. M. asperulum (solid circles, 2000-2008 survey; red
phylogenetic lineages and biogeographic origins in the tropical Southeast Asia islands (Chen et al. 2009), they showed fairly similar distribution pattern in Taiwan. They were distributed in the downstream section of the rivers close to the mouths in the east coast (Figs. 1-3). Apparently, these prawns of Southeast Asia islands origins still shared fairly similar life cycle modes and habitat requirements in Taiwan.
Streams in the east coast of Taiwan have high gradients as the mountain ranges approach closely to the coast. The stream water flows directly into coastal waters, forming a small blackish water zone outside the stream mouth rather than within the mouth. Larval development of these amphidromous prawns must be completed in the coastal waters, suggesting that the larvae may require high salinity and be highly tolerant to it, even to sea water.
There is the warm Kuroshio Current flowing northwardly closely to the shore along the east coast of Taiwan. The larvae of these amphidromous prawns have a chance to be transported northwardly by the current. It seems that the Kuroshio Current might play a role in the northward dispersion of these amphidromous prawns from Southeast Asia islands to Taiwan, even to Japan. According to this line of the reduction, M. equidens, M.
latidactylus and M. mammillodactylus of the
Western Southeast Asia group that are considered to have dispersed to Taiwan through the Philippines and/or China mainland (Chen et al. 2009) might be more likely through the Philippines than through the China mainland.
When the Kuroshio Current hits the southern tip of Taiwan, its main stem flows along the east coast of the island and a small western branch enters the Taiwan Strait and flows along the southwestern coast of the island (Lin et al. 1992;
Liang et al. 2003). This may explain the reason that these tropical prawns are often also found in the southwestern region.
During the Pleistocene glaciation, the western region of Taiwan was connected to the China mainland with the land bridge. The region was undoubtedly inhabitable for amphidromous prawns that requires brackish waster for larval development. Since the last glaciation, the land bridge has been transformed into Taiwan Strait, but the western region has still remained inhabitable for these tropical amphidromous prawns. This may be due to the cold China mainland current that flows southwardly along the west coast from China mainland in winter.
The island-wide group
For the four species of the island-wide group, the distribution patterns of M. japonicum and M.
formosense (Fig. 4A-B) that are considered to be
amphidromous (Shy et al. 1990; Shy 1994; Shy
et al. 1996; Suzuki and Kusamura 1997; Liu et al. 2007) differed greatly from those of the
amphidromous species of the east-coast group (Fig. 1-3) but were fairly similar to that of M.
nipponense that is non-obligatory amphidromous
and M. asperulum that is landlocked (Fig. 4C-D). The life cycles of M. japonicum and M.
formosense are somewhat similar to that of M. nipponense. They have small eggs and general
nine zoea stages (Ogasawara et al. 1979). In Japan the three species are found to spend their long pelagic zoeal stage in estuarine brackish waters, and then juveniles of M. formosense and M.
japonicum migrate upstream to freshwater (Ogasawara et al. 1979; Shokita 1979; Shy et al. 1990), whereas individuals of M. nipponense remain in the estuaries (river mouths) to complete
their life cycle, but some populations are found in coastal or inland freshwater lakes, resulting from a recent shift in its habitat from estuaries to inland freshwaters (Mashiko 1990; Mashiko and Numachi 2000). Such shift to freshwater form from amphidromous form have been well documented for other freshwater shrimp Paratya australiensis Kemp, 1917 in Australia (Williams 1977; Hancock and Bunn 1977; Walsh and Mitchell 1995; Cook
et al. 2006), and also many species of fishes and
invertebrates (Lee and Bell 1999; Lee 1999; Taylor and McPhail 1999; Waters and Wallis 2001; Raeymaekers et al. 2005; Cook et al. 2006). The shift of amphydromy to freshwater form plays an important role in diversification of many freshwater fauna in evolution.
The above evidences may suggest that M.
japonicum and M. formosense might not be
amphidromous but non-obligatory amphidromous like M. nipponense. A further study is needed to confirm the life cycle modes of those two species. Mashiko and Shy (2008) used molecular clock to estimate the time of speciation events and suggest that M. shokitai endemic to the Irimoto Island of the southern Ryukyu differentiated from
M. asperulum of Taiwan approximately 1.0 million
years ago, and M. nipponense from M. formosense 0.48 million years ago. According to Lee (2006) the uplift rate of Taiwan Central Range was initially slow at a rate of < 1 mm/year from 6 million years ago to 1 million years ago. Since then the rate has increased to 4-10 mm/year. The mountains started to build from the north toward the south at a rate of 60-90 km/million years.
M. asperulum of China mainland might arrive
in the west coast of Taiwan before the time of the rapid uplift of the Central Mountain Range 1 million years ago at the time when the east coast
remained in connection to the west coast with shallow valleys. This might enable the species to disperse across the island to the east coast. M.
nipponense and M. formosense arrived in the west
coast of Taiwan from China mainland apparently latter than that of M. asperulum, but still enable them to disperse to the east coast. The similar cases of cross island dispersions are reported for the freshwater fish Varicorhinus barbatulus and freshwater crab Candidiopotamon rathbunae (Wang et al. 2004; Shin et al. 2006).
The distribution patterns of the Macrobrachium prawns in Taiwan are related to their biogeographic origins and dispersal routes in the evolutionary history, their life cycle modes, and their adaptability to local climatic and environmental conditions at the present.
Acknowledgements
This study was financial supported by the Endemic Species Research Institute. The authors are grateful to Messrs R. C. Jang, M. H. Shen, K. W. Wang, Y. R. Syu and Y. H. Yu for field collections.
Literature Cited
Bate, C. S. 1868. On a new genus, with four new species of freshwater prawns. Proceedings of the Royal Society of London 1868: 363-368. Cai, Y. and M. S. Jeng. 2001. On a new species of Macrobrachium Bate, 1868 (Decapoda, Palaemonidae) from northern Taiwan. Crus-taceana 74 (3): 275-283.
Cai, Y. and S. Shokita. 2006. Report on a collection of freshwater shrimps (Crustacea: Decapoda: Caridea) from the Philippines, with descriptions of four new species. The Raffles Bulletin of
Zoology 54 (2): 245-270.
Chen, R. T., C. F. Tsai and W. N. Tzeng. 2009. Freshwater prawns (Macrobrachium Bate, 1868) of Taiwan with special references to their biogeographic origins. Journal of Crustacean Biology 29(2): 232-244. Chen, R. T., P. H. Ho and H. H. Lee. 2003.
Distribution of exotic freshwater fishes and shrimps in Taiwan. Endemic Species Research 5(2): 33-46.
Cook, B. D., A. M. Baker, T. J. Page, S. C. Grant, J. H. Fawcett, D. A. Hurwood and J. M. Hughes. 2006. Biogeographic history of an Australian freshwater shrimp, Paratya australiensis (Atyidae): The role life history
transition in phylogeographic diversification. Molecular Ecology 15: 1083-1093.
Hancock, M. A. and S. E. Bunn. 1997. Population dynamics and life history of Paratya
australiensis Kemp, 1917 (Decapoda: Atyidae)
in upland rainforest streams, south-eastern Queensland, Australia. Marine and Freshwater Research 48: 361-369.
Hwang, J. J. and H. P. Yu. 1982. Studies on the freshwater shrimps of the genus Macrobrachium (Crustacea: Decapoda: Palaemonidae) from Taiwan. Quarterly Journal of the Taiwan Museum 25: 157-180.
Lee, C. E. 1999. Rapid and repeated invasions of freshwater by the copepod Eusytemora affinis. Evolution 53: 1423-1434.
Lee, C. E. and M. A. Bell. 1999. Causes and consequences of recent freshwater invasions by saltwater animals. Trends in Ecology and Evolution 14: 284-288.
Lee, Y. H., C. C. Chen, T. K. Liu, H. C. Ho, H. Y. Lu and W. Lo. 2006. Mountain Building Mechanism in Southern Central Range of
Taiwan Orogeny Belt-from Accretionary Wedge Deformation to Arc-Continental Collision. Earth and Planetary Science 252: 413-422.
Liang, W. D., T. Y. Tang, Y. J. Yang, M. T. Ko and W. S. Chuang. 2003. Upper-ocean currents around Taiwan. Deep-Sea Research Part II 50: 1085-1105.
Lin, S., J. Y. Shy and H. Yu. 1988. Morphological observation on the development of larval
Macrobrachium asperulum (Von Martens,
1868) (Crustacea, Decapoda, Palaemonidae) reared in the laboratory. Journal of the Fisheries Society of Taiwan 15(2): 8-20.
Lin, C. T., C. Z. Shyu and W. H. Shin. 1992. The Kuroshio Fronts and Cold Eddies off Northeastern Taiwan Observed by NOAA-AVHRR Imageries. Terrestrial, Atmospheric and Oceanic Sciences 3(3): 225-242. Liu, M. Y., Y. X. Cai and C. S. Tzeng. 2007.
Molecular systematics of the freshwater prawn genus Macrobrachium Bate, 1868 (Crustacea: Decapoda: Palaemondiae) infaerred from mtDNA sequences, with emphasis on East Asian species. Zoological Studies 46 (3): 272-289.
Mashiko, K. 1990. Diversified egg and clutch sizes among local populations of the freshwater prawn Macrobrachium nipponense. Journal of Crustacean Biology 10: 306-314. Mashiko, K. and K. Numachi. 2000. Derivation
of populations with different-sized eggs in the palaemonid prawn Macrobrachium nipponense. Journal of Crustacean Biology
20: 118-127.
Mashiko, K. and J. Y. Shy. 2008. Derivation of four morphologically affiliated species of
with divergent reproductive characteristics in Northeastern Asia. Journal of Crustacean Biology 28(2): 370-377.
Ogasawara, Y., S. Koshio and Y. Taki. 1979. Response to salinity in larvae from three local populations of the freshwater shrimp,
Macrobrachium nipponense. Bulletin of the
Journal Society of Scientific Fisheries 45(8): 937-943.
Raeymaekers, J. A. M., G. E. Maes, E. Audenart and F. A. M. Volckaert. 2005. Detecting Holocene divergence in the anadromous-freshwater three-spined stickleback
(Gaste-rosteus aculeatus) system. Molecular Ecology
14: 1001-1014.
Shin, H. T., H. C. Hung, C. D. Schubart, C. A. Chen and H. W. Chang. 2006. Intraspecific genetic diversity of the endemic freshwater crab Candidiopotamon rathbunae (Decapoda, Brachyura, Potamidae) reflects five million years of the geological history of Taiwan. Journal of Biogeography 33: 980-989. Shokita, S. 1977. Abbreviated metamorphosis of
land-locked fresh-water prawn, Macrobrachium
asperulum (Von Martens, 1868) from Taiwan.
Annotations Zoologicae Japoneses 50: 110-122. Shokita, S. 1979. The distribution and speciation of the inland water shrimps and prawns from the Ryukyu Islands-II. Bulletin of the College of Science, University of the Ryukyus 28: 93-278. Short, J. W. 2000. Systematics and biogeography of Australian Macrobrachium (Crustacea: Decapoda: Palaemonidae) - with descriptions of other new freshwater Decapoda. Ph. D. dissertation. The University of Queensland. Queensland, Australia.
Shy, J. Y., H. G. Lai and H. P. Yu. 1987. Morphological observation on the development of larval
Macrobrachium nipponense (De Haan)
(Crus-taceaa, Decapoda, Palaemonidae) reared in the laboratory. Journal of the Fisheries Society of Taiwan 14(1): 1-14.
Shy, J. Y., Y. E. Tsou and H. P. Yu. 1990. Morphological observation on the larval de-velopment of Macrobrachium formosense (Crustacea: Decapoda: Palaemonidae). Journal of the Fisheries Society of Taiwan 17(1): 21-34. Shy, J. Y. 1994. Taxonomy, distribution and ontogeny of freshwater shrimps and crabs in Taiwan. Ph. D. dissertation. National Taiwan Ocean University. Taiwan.
Shy, J. Y., H. T. Lai and H. P. Yu. 1996. On the freshwater shimps and crabs from the three southwestern counties Yunlin, Chiayi and Tainan of Taiwan. Annual of Taiwan Museum 39: 201-246.
Shy, J. Y. and H. P. Yu. 1998. Freshwater shrimps of Taiwan. National Museum of Marine Biology and Aquarium, Pingdong, Taiwan. Suzuki, H. and T. Kusamura. 1997. Reexamination
of the diagnostic characters of two freshwater palaemonid prawns, Macrobrachium nipponense (De Haan, 1849) and M. formosense Bate, 1868 (Decapoda, Caridea) from Japan. Crustaceana 70: 831-839.
Taylor, E. B. and J. D. McPhail. 1999. Evolutionary history of an adaptive radiation in species pairs of three-spine sticklebacks (Gasterosteus): Insights from mitochondrial DNA. Biological Journal of the Linnean Society 66: 271-291. Walsh, C. J. and B. D. Mitchell. 1995. The freshwater shrimp Paratya australiensis (Kemp, 1917) (Decapoda: Atyidae) in estuaries of south-western Victoria, Australia. Marine and Freshwater Research 46: 959-965. Wang, J. P., H. D. Lin, S. Huang, C. H. Pan, X.
L. Chen and T. Y. Chiang. 2004. Phylogeography of Varicorhinus barbatulus (Cyprinidae) in Taiwan based on nucleotide variation of mtDNA and allozymes. Molecular Phylogenetics and Evolution 31: 1143-1156.
Waters, J. M. and G. P. Wallis. 2001. Cladogenesis and loss of the marine life-history phase in freshwater galaxiid fishes (Osmeriformes: Galaxiidae). Evolution 55: 587-597. Williams, W. D. 1977. Some aspects of the ecology
of Paratya australiensis (Crustacea: Decapoda: Atyidae). Australian Journal of Marine and Freshwater Research 28: 403-415.
Yeh, M. F., J. J. Chiou and T. W. Lee. 2000. Distribution of fishes in Chinshoei Creek in relation to elevation and stream gradient. Endemic Species Research 2: 34-43.