Taipei Medical University Institutional Repository:Item 987654321/4214
112
0
0
全文
(2) 目錄 中文摘要…………………………………………………….2 英文摘要…………………………………………………….4 縮寫………………………………………………………….6 壹、緒論………………………………………………….....8 貳、材料與方法 實驗材料……………………….……………………….24 實驗方法…………………………….………………….31 參、結果…………………………………………………....44 肆、討論…………………………………………………....60 伍、結論…………………………………………………....69 陸、圖表…………………………………………………....70 柒、參考文獻……………………………………………....99. 1.
(3) 中文摘要 有許多證據顯示誘導出誘發型一氧化氮合成酵素(iNOS)的表現以 及一氧化氮(NO)的產生對於抗病毒、免疫調節以及抗腫瘤情況是具有 幫助的。但假如在不對的地方及時間誘導出來,則會造成許多人類疾 病相關的病理生理病變。在中樞神經系統當中,一氧化氮的大量增加 會造成一些神經發炎性及神經退化性疾病的發生,例如:多發性硬化 症(multiple sclerosis)、愛滋痴呆症候群(HIV dementia)、腦部局部缺 血 、 帕 金 森 氏 症 (Parkinson's disease) 以 及 阿 滋 海 默 症 (Alzheimer's disease)等。然而在以上的病理情況發生下,中樞神經系統中小膠質 細胞會快速地被過度活化,而在本篇研究中,我們則會加以探討氧化 態低密度脂蛋白(ox-LDL)與 phenytoin 對於小膠質細胞產生誘發型一 氧化氮合成酵素之影響機轉。 氧化態低密度脂蛋白對於動脈粥狀硬化的發生扮演著很重要的 角色,而 ox-LDL 透過與清道夫受體 A 型的高度親合力以及內化作用 之後,進而影響一些慢性發炎疾病的發生。經由我們的結果證實, ox-LDL 可以誘發小膠質細胞產生大量的一氧化氮,並發現 ox-LDL 也能誘發大量 iNOS 蛋白及其 mRNA 的表現,代表著這 ox-LDL 對於 iNOS 的影響可能在基因表現層面。因此我們對於 ox-LDL 在 iNOS 的表現機轉上做進一步的探討,利用不同抑制劑的觀察,發現誘導出 iNOS 蛋白表現主要是經由 p38 MAPK 的活化,並且其活化的 p38 能 進入細胞核內,促使轉錄因子 ATF-2 的磷酸化。綜合以上結論,ox-LDL 活化小膠質細胞,主要是經由活化 p38 來活化轉錄因子 ATF-2,進而 啟動 iNOS 的基因表現,最終使得 iNOS 蛋白及 NO 大量產生。另一 2.
(4) 方面也發現 ox-LDL 所引發的訊息傳導與 dynamin 有著相當的關係, 即透過 endocytosis 抑制劑可以降低 ox-LDL 所誘發的 iNOS 蛋白表 現。在相關的受體方面,發現 ox-LDL 可能與 fucoidan 所結合的受體 並不相同,由於 fucoidan 並無法誘發小膠質細胞產生 iNOS 蛋白表現。 phenytoin 為一種抗痙攣藥物,臨床上用來治療癲癇相當有效。近 來有文獻指出,在腦脊髓炎與多發性硬化症中,phenytoin 能夠改善 其中小膠質細胞活化的情況,而目前對於此相關機轉還不甚清楚。我 們從實驗結果中顯示,phenytoin 能夠降低經由 LPS 刺激小膠質細胞 所產生的 NO,並且不影響其細胞存活率。此外,我們還發現 phenytoin 能夠抑制 LPS 刺激小膠質細胞所產生的 iNOS 蛋白表現,而且是有 濃度依賴性的抑制效果。在機轉方面,我們證實 phenytoin 並不能有 效抑制經 LPS 活化所造成 IκB-α抑制蛋白的降解,因此,我們推論 phenytoin 的抑制作用可能不是經由 NF-κB 這條路徑。. 3.
(5) Abstract Accumulating evidence indicates that induction of inducible nitric oxide synthase (iNOS) expression and hence nitric oxide (NO) production. has. been. described. to. have. beneficial. antiviral,. immunomodulatory, and antitumoral affect. However, induced at the wrong place or at the wrong time, iNOS has detrimental consequences and seems to be involved in the pathophysiology of different human diseases. As such, elevated levels of NO produced within the central nervous system (CNS) are associated with the pathogenesis of neuroinflammatory and neurodegenerative human diseases such as multiple sclerosis, HIV dementia, brain ischemia, trauma, Parkinson's disease, and Alzheimer's disease. However, the overactivation of microglia can occur rapidly after the above pathologies of the CNS. In this study, we have observed that oxidized low-density lipoprotein (ox-LDL) and phenytoin affect microglial functions in association with the mechanisms of iNOS expression in microglial cells. It is well documented that ox-LDL have been shown to play a crucial role in atherosclerosis. The class A scavenger receptor (SR-A) mediates the high affinity binding and internalization of ox-LDL implicated in the development of chronic inflammation. Our results demonstrate that ox-LDL stimulates NO production in microglial cells, and it also stimulates. both. protein. concentration-dependent. levels. manner,. and. mRNA. indicating. that. of this. iNOS. in. oxidative. components may affect iNOS gene expression. Therefore, we 4.
(6) investigated the molecular mechanism of ox-LDL on iNOS expression. Using different pharmacological inhibitors, we found the induction of iNOS by p38 mitogen-activated protein kinase activation. Besides, we even more found that activated p38 in the nucleus induced phosphorylation of activating transcription factor-2 (ATF-2). Taken together, our data clearly demonstrated that ox-LDL-induced microglial cells production of NO and iNOS was mediated mainly by phosphorylation of p38 and ATF-2. On the other hand, we also found that dynamin inhibitor decreased iNOS production in ox-LDL stimulated microglial cells, and fucoidan, SR-A ligand, couldn’t stimulate iNOS production in microglial cells. Phenytoin is an anticonvulsant drug which can be useful in the treatment of epilepsy clinically. The recent report indicated that sodium channels are important for activation of microglia in encephalomyelitis (EAE) and multiple sclerosis (MS), but the anti-inflammatory mechanisms remain poorly understood. We showed here that phenytoin decreased NO production in LPS-stimulated microglial cells, without affecting cell viability. Besides, we even more found that phenytoin suppressed the expression of iNOS protein induced by LPS in a dose dependent way. Further, we found that phenytoin couldn’t inhibit the LPS-induced IκB-α degradation suggesting that this drug might not affect LPS-induced activation of NF-κB by abrogating the LPS-induced IκB-α degradation.. 5.
(7) 縮寫 ApoB-100. apolipoproteinB-100. AP-1. activator protein-1. ATF-2. activating transcription factor-2. BSA. bovine serum albumin. COX-2. cyclo-oxygenase-2. CREB. cAMP response element binding protein. C/EBP. CCAAT-enhancer box binding protein. DMSO. dimethylsulphoxide. EAE. encephalomyelitis. ecNOS. endothelial constitutive nitric oxide synthase. ERK. extracellular signal-regulated kinases. FBS. fetal bovine serum. GM-CSF. granulocyte-macrophage colony stimulating factor. 4-HNE. 4-hydroxynonenal. HIV. human-immunodefficiency virus. ICAM-1. ntercellular adhesion molecule-1. IFN-γ. interferon-γ. IκB. inhibitor of NF-κB. IKK. IκB kinase. IL-1β. interleukin -1β. iNOS. inducible nitric oxide synthase. IRF-1. interferon regulatory factor-1. JNK. c-Jun N-terminal kinase. LDL. low-density lipoprotein. LOX-1. lectin-like ox-LDL receptor-1. LPS. lipopolysaccharide. MAPKs. mitogen-activated protein kinases. MCP-1. monocyte chemotactic protein-1. M-CSF. macrophage colony stimulating factor 6.
(8) MD-2. myeloid differentiation protein-2. MDA. malondialdehyde. MKK. mitogen-activated protein kinases kinase. MS. mutiple sclerosis. ncNOS. neuronal constitutive nitric oxide synthase. NEMO. nuclear factor κ-B essential modulator. NF-κB. nuclear factor κ-B. NLS. nuclear localization signal. NO. nitric oxide. Oct. octamer factor-1. Ox-LDL. oxdized low density lipoprotein. PDGF. platelet derived growth factor. PUFAs. polyunsaturated fatty acids. RNS. reactive nitrogen species. ROS. reactive oxygen species. RT-PCR. reverse transcription-polymerase chain reaction. SAPK. stress-activated protein kinases. SDS-PAGE. sodium dodecyl sulfate-polacrylamide gellectrophoresis. SR-A. scavenger receptor of class A. SREC. scavenger receptor expressed by endothelial cells. STAT. signal transducer and activator of transcription. TLR4. toll-like receptor 4. TNF-α. tumor necrosis factor-α. VCAM-1. vascular cell adhesion molecule-1. 7.
(9) 壹、緒論 一、小膠質細胞與神經退化性疾病之關係 中樞神經系統主要由神經元細胞和神經膠質細胞組成,而神經元 細胞在大腦皮層腦細胞中只佔了 15%,神經膠質細胞則佔了 85%。 而其中神經膠質細胞可分為星狀膠質細胞(astrocytes)、小膠質細胞 (microglia) 及 寡 樹 突 膠 質 細 胞 (oliogodendrocytes) 。 星 狀 膠 質 細 胞 (astrocytes)含量最多,佔了 80%左右,而小膠質細胞只佔了神經膠質 細胞的 12%左右,但卻是主要的免疫細胞。小膠質細胞為大腦中的長 駐型巨噬細胞(resident macrophage-like),散佈於中樞系統中,功能上 與周邊組織的巨噬細胞類似[1],對於免疫調節、外來微生物及腫瘤 細胞的防禦作用和組織修復皆扮演著很重要的角色。 在健康的大腦中,小膠質細胞處於休息狀態(Resting state),此時 細胞型態為本體較小且長而突起出現分支,稱之為分支狀小膠質細胞 (ramified microglia),但休息狀態並不代表是不活化狀態,它們仍具有 高度活性,利用顯微鏡發現其分支不斷有偽足狀突起形成球狀頂端, 讓小膠質細胞來隨時偵測以應付環境的改變,而這些分支也會直接與 星狀細胞、神經細胞和血管接觸,成為大腦皮質細胞中重要的溝通管 道 [2-4]。當中樞神經系統發生創傷或微生物入侵時,小膠質細胞會 被活化,此時細胞型態為本體較大而突起較短,稱之為阿米巴狀小膠 質細胞(ameboid microglia),成為活化狀態(Activated state)[5],此種型 態的小膠質細胞通常伴隨下列現象發生:包括細胞增生、移動、吞噬 作用增加,釋放大量細胞激素和化學激素(Chemokines)等發炎相關物 質,如 tumor necrosis factor-α (TNF-α)、interleukin -1β (IL-1β)、 8.
(10) chemokines、 prostaglandins、 COX-2 、活性氧族群 (reactive oxygen species, ROS)及活性氮族群(reactive nitrogen species, RNS),如一氧化 氮(nitric oxide, NO)等[6-8]。而這些物質可能直接具有神經毒性,或 間接地促使各類免疫細胞再釋放毒性物質,導致二度神經損傷[9]。 對於中樞系統而言,小膠質細胞適當的活化是一種保護作用,但 是錯誤的調節機制卻會造成嚴重的傷害,因此腦部神經發炎反應對於 中樞神經系統疾病的病程上扮演著很重要的角色[7]。而許多文獻指 出慢性發炎反應是導致 dopaminergic neurons 死亡與神經退化性疾病 發生的重要因素之一[10;11]。其中參與腦部發炎反應的細胞主要有兩 種神經膠質細胞(glia cells),分別為小膠質細胞與星狀膠質細胞,又 以小膠質細胞最為主要。由於小膠質細胞被認為是腦內免疫系統的第 一道防線,當腦部受到微生物、病毒感染或創傷時,小膠質細胞會快 速活化[12],而此現象在許多神經退化性疾病的病程中也都可以觀察 到,例如:多發性硬化症 (multiple sclerosis)、愛滋痴呆症候群 (HIV dementia)、腦部局部缺血、帕金森氏症 (Parkinson's disease)以及阿滋 海默症 (Alzheimer's disease)等[11-13]。因此,如何調控小膠質細胞的 活化與發炎物質的產生,對於神經退化疾病是個值得去深入探討的研 究方針。. 二 、 氧 化 低 密 度 脂 蛋 白. (Oxidized Low-Density. Lipoprotein,ox-LDL) 之病理角色 氧化態低密度脂蛋白即為低密度脂蛋白(low-density lipoprotein, LDL)受到脂質過氧化的產物。LDL 為人類脂蛋白依比重分類的其中 一種,密度約為 1.019-1.063 g/ml,其組成由約 2700 多個脂肪酸,而 9.
(11) 其 中 有 一 半 為 多 元 不 飽 和 脂 肪 酸 (polyunsaturated fatty acids, PUFAs),和脫輔基蛋白(apolipoproteinB-100, ApoB-100),構成一個圓 球 狀 的 大 粒 子 , 球 心 為 膽 固 酯 (cholesterol ester) 與 三 酸 甘 油 脂 (triglycerides)所組成的親脂性核心,平均直徑長為 18-25 nm,分子量 約為 250-280 萬 Dalton[14]。而其相關的脂蛋白受體會辨認 LDL 上的 ApoB-100 蛋白並與之結合,進而使細胞進行胞飲作用,藉此來進行 LDL 之分解代謝及應用。 LDL 在體內能幫助運送膽固醇至身體各組織,主要功能在運送磷 脂,由於 LDL 含有大量的膽固醇,約佔血漿總膽固醇的 60-70%,所 以當代謝速率差時,血漿濃度則會過高(160-180 mg/ml)造成過多膽固 醇滲入血管壁中,使得動脈粥狀硬化的發生率增加,因此 LDL 血中 濃度過高對於動脈粥狀硬化的發生視為一危險因子,有文獻指出降低 血中膽固醇的量可預防動脈粥狀硬化疾病的發生[15]。LDL 中的多元 不飽和脂肪酸(PUFAs)在自由基或其他氧化劑(如:金屬離子 Cu2+、Fe2+ 等)的作用下,會先形成不飽和脂肪酸自由基(R‧),進而氧化成脂過 氧基(ROO‧),最後形成過氧化脂質(ROOH)與另一不飽和脂肪酸自 由基(R‧),因而引起一連串的自由基連鎖反應,造成大量的脂質過 氧化(lipid peroxidation),而這些脂質過氧化物最終被分解成多種產 物,包括丙二醛(malondialdehyde, MDA)、4-羥烯酸(4-hydroxynonenal, 4-HNE)等,並且會與 LDL 的 ApoB-100 上的離胺酸(lysine)殘基結合, 形 成 所 謂 的 氧 化 態 低 密 度 脂 蛋 白 (ox-LDL)[16] 。 ox-LDL 由 於 ApoB-100 結構受到改變,故不再受原 LDL 受體所辨識,而改為與清 道夫受體(scavenger receptor)有高度的親和性[17]。有文獻指出當內皮 細胞受到 ox-LDL 的刺激過後,發現其 Lectin-like ox-LDL receptor-1 10.
(12) (LOX-1)的 mRNA 表現量有顯著的增加,此外在平滑肌細胞、巨嗜細 胞及單核球細胞中還發現有其他 ox-LDL 的受體,包括:SR-A、 SR-BⅠ、CD36 及 CD68 等[18;19]。 而 ox-LDL 對於不同細胞也就有著不同的影響,例如對於內皮細 胞則能刺激其分泌黏附分子(adhesion molecules),包括有 E-selectin、 VCAM-1 (vascular cell adhesion molecule-1)及 ICAM-1 (intercellular adhesion molecule-1),使得單核球細胞黏附於內皮後移行入內膜。此 外 ox-LDL 還 會 促 使 細 胞 表 現 單 核 球 細 胞 趨 化 性 蛋 白 (monocyte chemotactic protein-1, MCP-1)及巨噬細胞聚落刺激因子(macrophage colony stimulating factor, M-CSF),來促使單核球細胞分化成巨噬細 胞,而這些巨噬細胞利用清道夫受體將 ox-LDL 噬入,而形成所謂的 泡沫細胞(foam cells),最終大量泡沫細胞堆積在血管內膜下方,此過 程為促成動脈粥狀硬化的重要因素[20;21]。對於平滑肌細胞,ox-LDL 能誘導其產生血小板生長因子(platelet derived growth factor, PDGF), 來促進其移行與增生[22],此現象也被認為是促成阻塞性血管疾病的 病灶,而此類疾病一般好發於主動脈、冠狀動脈、頸動脈以及腦部動 脈,其中在腦部又以缺血性中風最為主要。此外,ox-LDL 更可誘發 慢性發炎反應,並在許多腦部發炎及退化性疾病扮演相當重要的角色 [23;24]。進來文獻也指出 ox-LDL 本身與急性冠狀動脈症候群及急腦 部梗塞發作有相當的關係[25;26],並發現有降低冠狀動脈血流的作用 [27],以及造成微血管通透度與白血球黏滯性之增高[28]。另外對於 小膠質細胞與星狀膠質細胞,ox-LDL 則能增加其活性氧族群(reactive oxygen species, ROS)的釋放,藉此以影響其增生、細胞活化及細胞激 素的釋放,因此 ox-LDL 可作為腦部發炎反應的一種指標性物質 11.
(13) [24;29;30],但由於活化機轉還不明,故本篇論文以此作為刺激劑, 以了解與小膠質細胞活化間的影響與分子機轉。. 三、脂多醣體(Lipopolysaccharide,LPS)與發炎反應之關係 脂 多 醣 體 (lipopolysaccharide, LPS) 或 稱 為 細 菌 性 內 毒 素 (endotoxin),位於革蘭氏陰性菌(Gram-negative)之細胞壁上,並且是 最早發現與發炎反應有關的主要成分之ㄧ。其主要結構可分為兩部 分,分別為lipid A與多醣(polysccharides),而Lipid A為LPS的重要毒 性來源[31]。LPS屬於大分子,所以較不易通過血腦屏障,但當遇到 發炎的情況時,血腦屏障則會被破壞,使得LPS得以進入血腦屏障來 活化小膠質細胞,因此常使用LPS來作為誘導大腦產生發炎反應的實 驗模式[32]。過去研究也有指出,在巨噬細胞[33]、大鼠動脈平滑肌 細胞[34]及肺動脈平滑肌細胞[35]等細胞中,單以LPS或合併使用 IL-1β、TNF-α等cytokines,能夠增加iNOS基因轉錄的作用。而當利 用LPS、cytokines與amyloid β刺激小膠質細胞時,過程中會經由活化 蛋白激酶(protein kinase)、酪胺激酶(tyrosine kinase)、MAPK等路徑, 進而活化某些轉錄因子(transcription factor),例如:nuclear factor-κB (NF-κB)、cAMP response element binding protein (CREB)及activator protein-1 (AP-1)等,藉此來產生大量的iNOS、COX-2及cytokines等發 炎物質[36;37]。當這些發炎cytokines過度表現時,便會導致循環功能 的障礙、組織的破壞、細菌性腦膜炎及高致死率的敗血性休克[38]。 目前已知與LPS相關的受體包含三個部份,分別為CD14、Toll-like receptor 4 (TLR4)與myeloid differentiation protein-2 (MD-2)。當細胞 (如:macrophage或monocytes)受到LPS刺激時,LPS便會與LPS-binding. 12.
(14) protein (LBP)在plasma中結合,並且第一時間被細胞表面的CD14吸 引,然後直接與TLR4和coreceptor MD-2結合[39],接著再將訊息往下 傳給adapter molecule MyD88啟動一連串訊息傳遞,最後便活化轉錄 因子,以進行基因的轉錄及轉譯作用[40]。. 四、一氧化氮(Nitric Oxide)在腦部疾病之病理介紹 一氧化氮是含一未配對電子的自由基,屬於活性氮族群(reactive nitrogen species, RNS)之ㄧ,此分子即不穩定,在氧存在下會快速代 謝成亞硝酸鹽(nitrite)和硝酸鹽(nitrate)。在哺乳動物細胞中,NO 是最 小的產物之一,也是半衰期極短的分子。在 1988 年,Garthwaite 等 學者在腦細胞中發現有一氧化氮的存在,並且認為是神經性的訊息傳 導因子[41],因此,NO 是中樞與周邊系統很重要的訊息傳遞物質, 而對於大腦的免疫系統扮演著很重要的角色[42]。當腦部受到創傷或 感染時,會導致局部釋放大量前發炎因子,包括腫瘤壞死因子(tumor necrosis factor-α, TNF-α) 、 干 擾 素 γ(interferon- γ) 及 介 白 素 -1β(interleukin-1β)等,當作用於小膠質細胞與星狀膠質細胞時,會促 進其誘導型一氧化氮合成酶(inducible nitric oxide synthase, iNOS)的 表現,進而引發 NO 的產生[43;44]。而釋放出的這些 NO、cytokines 以及 reactive oxygen species (ROS)等皆參與神經退化的過程[45;46]。 許多文獻亦指出高量的 NO 會對神經細胞、小膠質細胞及星狀膠質細 胞產生毒性[1;47]。 NO 的生理作用依合成的量、時間及被合成的位置來決定,例如 在過去發現低量的 NO 具有抗腫瘤與抗生素的作用,但當細胞激素誘 發所產生高量的 NO 則具有細胞毒性。此外,NO 能使細胞膜上的脂 13.
(15) 質過氧化,由 NO 和 superoxide 可以形成相當有破壞性的過氧亞硝基 物(peroxynitrite, ONOO-),它能造成 DNA 斷裂、脂質氧化和蛋白 nitration [48],而 nitration 的蛋白質,例如:α-synuclein 正是神經退 化性疾病腦部病變斑塊區的構成單元,即是帕金森氏症(Parkinsin’s disease)患者腦部斑塊(Lewy body)的主要成分[49]。在哺乳動物體內還 具有許多重要的生理與病理作用,包括擴張血管、抑制血小板凝集、 抑制細胞增生、作為神經傳導物質,以及與神經退化性疾病等病理狀 況有關,並在免疫系統發炎反應中扮演重要角色[50;51]。 NO 在體內之生合成路徑,主要是由一氧化氮合成酵素(nitric oxide synthase, NOS)經由三種受質:L-arginine、NADPH 與 O2,加 上五種輔助因子:FAD、FMN、calmodulin、(6R)-tetrahydrobiopterin (BH4)及 heme 的作用,催化產生 NO 及 L-citrulline[52]。目前可將 NOS 粗 略 區 分 為 兩 種 不 同 形 式 : 結 構 型(constitutive)與誘導型 (inducible)兩類一氧化氮合成酵素。兩類 NOS 相似之處是結構上均 含有 calmodulin-, FMN-及 NADPH-binding domains。結構形一氧化 氮合成酵素(cNOS)平時就存在著,包括(1)神經性結構型一氧化氮合 成酵素(neuronal constitutive NOS, ncNOS, or type Ⅰ),主要存在於 中樞與周邊系統,主要與腦部學習與記憶有關,以及(2)內皮性結構 型一氧化氮合成酵素(endothelial constitutive NOS, ecNOS or type Ⅲ),主要存在於血管內皮細胞,主要與血管擴張有關。而這兩種 酵素為鈣離子依賴型(Ca2+-dependent)酵素,其活性需要藉由細胞內 鈣離子濃度的變化來調節[53]。另一種誘導型一氧化氮合成酵素 (iNOS or type Ⅱ),則須由 endotoxin、proinflammatory cytokine,例 如 TNF-α、interleukin-1 (IL-1)和 interferon-γ (IFN-γ)等,及各種訊息 14.
(16) 傳導來誘導產生,其活性不須依賴鈣和 calmodulin 的存在,為非鈣 離子依賴型(Ca2+-independent)酵素,主要是以轉錄因子(transcription factor)來調節其酵素活性[54]。通常 cNOS 釋放少量且短時間的 NO,但 iNOS 卻能造成大量且長時間的 NO[53]。iNOS 在中樞神經 系統中,主要存在於小膠質細胞及星狀膠質細胞中,並在 lipopolysaccharide (LPS)、cytokines、以及 amyloid β處理下會產生 大量 iNOS[36;37;44]。不僅如此,在其他中樞神經系統中,如:神 經細胞、血管內皮細胞、血管平滑肌細胞等,皆與小膠質細胞一樣 具有 iNOS 的表現[55;56]。 iNOS 蛋白質分子量大小約 130 kDa,其基因序列主要位於 17 號染色體[57]。目前已知調控 iNOS 基因上游的轉錄因子包含有 AP-1 (activating protein-1)、 NF-κB 、 IRF-1 (interferon regulatory factor-1)、 C/EBP (CCAAT-enhancer box binding protein) 、 CREB (cAMP–responsive element binding protein)及 Oct (octamer factor-1) 等[54],顯示 iNOS 的表現可藉由多種轉錄因子的調控。而在以大 鼠膠質細胞的實驗中,發現調節其 iNOS 表現的轉錄因子主要有 CREB、C/EBP 及 ATF-2 (activating transcription factor-2),並且主要 是經由 mitogen activated protein kinase (MAPK)此訊息傳遞路徑 [58]。由於 NO 特殊的生理活性,當其濃度過高會產生不必要的生 理作用,例如在阿茲海默症的過程的病程中,會引發過量的 iNOS 表現而產生大量的 NO,因而導致嚴重的神經毒性[59]。動脈粥狀 硬化損傷的發炎部位增加時,常伴隨 iNOS 活性及表現的增加,同 時也增加 NO 代謝產物 peroxynitrite (ONOO-)的產生。Peroxynitrite 是大量 NO 與超氧陰離子反應所形成的產物,具有相當毒性,具有 15.
(17) 誘發低密度脂蛋白(LDL)氧化、促進血小板凝集作用,因而促進動 脈粥狀硬化的病程[60;61],因此如何調控 iNOS 的表現進而控制 NO 的產生一直是廣為研究的課題。在本實驗中,也將對 ox-LDL 與 phenytoin 在這方面做探討。. 六、基因表現之相關訊息傳遞路徑 (1)Mitogen-activated protein kinase (MAPKs) Mitogen-activated protein kinases (MAPKs)屬於serine/threonine kinases 酵素,會受到生長因素或環境因素等影響。主要的mammalian MAPKs 包括四個路徑,分別主要的上游路徑:(a) extracellular signal-regulated kinases (ERK1與ERK2);(b) c-Jun N-terminal kinase (JNK1, JNK2與 JNK3) ; (c) ERK5 (big MAP kinase 1 [BMK1] 與 MAPK7) (d) p38-MAPK (p38-MAPKα、p38-MAPKβ、p38-MAPKγ [ERK6, SAPK3] 和p38-MAPKδ [SAPK4])[62]。而其中ERK pathway主要會受到生長因 子的活化且與細胞的生長、增生與分化有關[63],另外JNK與p38 MAPK也已知為stress-activated protein kinases (SAPK),因此它們容易 受到壓力的刺激(如:osmotic shock)、紫外光的照射、LPS以及一些細 胞激素(如:TNF-α)等因素而活化,進而啟動一些基因調控機轉,通 常與細胞的存活、凋亡、transformation、進化、遷移與免疫之活化有 關[64;65]。以上種種的刺激因子易造成一種或多種MAPK kinase (MKK)刺激Thr與Tyr的區域磷酸化而產生活性,進而translocate至細 胞核,促使轉錄因子(AP-1為主)磷酸化,進而啟動活化與基因表現(其 中包括調節iNOS等基因表現)[66]。例如在TGF-β-activating kinase (TAK)-1的刺激之下,主要會活化MKK3/MKK6-p38 MAPK訊息路 16.
(18) 徑,進而抑制cyclin D1的基因表現,而達到抑制細胞增生的效果,主 要 由 於 cyclin D1 有 促 進 細 胞 週 期 進 程 的 的 功 用 。 假 若 經 由 活 化 MKK1-p44/p42 MAPK訊息路徑,則會促進cyclin D1的基因表現,進 而達到細胞增生的效果[67]。另有文獻指出,LDL對於血管平滑肌細 胞能夠經由產生H2O2活化p38 MAPK,進而促使AP-1及NF-κB轉錄因 子活化,使得IL-8大量產生[68]。另一方面,已知ox-LDL對於單核細 胞及內皮細胞皆能誘發IL-8的產生[69;70]。不僅如此,LDL及ox-LDL 誘導內皮細胞產生PGE2,主要途徑也是經由活化p38 MAPK以及活化 NF-kB與CREB轉錄因子,進而產生COX-2蛋白表現的結果[71]。而對 於ox-LDL誘發內皮細胞大量表現MMP-1蛋白及其活性,發現p38 MAPK的活化可能扮演著很重要的角色[72]。在ox-LDL接受體的表現 方面,透過heparin-binding epidermal growth factor-like growth factor (HB-EGF)與平滑肌細胞的實驗發現,Lectin-like ox-LDL receptor-1, (LOX-1)的表現主要可能經由ERK及p38 MAPK活化的路徑[73]。對於 ox-LDL誘導巨噬細胞的增生,發現主要是經由granulocyte/macrophage colony-stimulating factor (GM-CSF)的產生,進而使其Ras-或RhoA-p38 MAPK路徑的活化[74]。綜合以上結果,瞭解p38 MAPK路徑的活化 與ox-LDL誘發相關發炎反應有著相當重要的關係。 p38為訊息傳遞路徑MAPK中的一個激酶,其活化態主要是經由上 游激酶MAP kinase kinases (MKKs)將其threonine及tyrosine磷酸化而 成,而主要的MKKs有兩種,分別為MKK3及MKK6。但也有文獻指 出有些p38 MAPK訊息傳遞路徑則不是經由MKKs所活化,而是經由 TAB1 (transforming growth factor-β-activated protein kinase 1 (TAK1)-binding protein),此活化途徑主要發生在LPS、TNF及CpG刺. 17.
(19) 激B細胞細胞株[75]。此外,MKKs的上游則有MKK kinase (MAP3K) 參與其中,主要包括有TAK1、ASK1/MAPKKK5、DLK/MUK/ZPK、 與 MEKK4,假若MAP3K過度表現則會活化p38及JNK兩條路徑,這 也是為何兩條路徑經常共同活化的主要原因[76]。而MAP3K所啟動下 游的磷酸化主要經由Rho家族中的低分子量GTP結合蛋白(包括Rac1 與Cdc42),例如Rac1主要能與MEKK1或MLK1結合,而Cdc42則只能 與MLK1結合。對於p38路徑的調節主要經由MAP kinase phosphatase (MKP) 家 族 來 對 激 酶 去 磷 酸 化 , 例 如 serine/ threonine protein phosphatase type 2C (PP2C)主要是對人類MKK6與MKK4做負向調 控,以抑制MAP kinase HOG1路徑的進行[77]。至於p38 MAPK活化 後所結合的受質,除了之前所述的AP-1等轉錄因子外,還有MAP kinase-activated protein kinase 2 (MAPKAPK2或MK2)、small heat shock protein 27 (HSP27) 、 lymphocytespecific protein 1 (LSP1) 及 tyrosine hydroxylase等蛋白。因此,p38 MAPK除了啟動基因表現外還 有 許 多 功 能 , 例 如 受 質 MK2 就 能 進 一 步 的 磷 酸 化 tristetraprolin (TTP),以影響p38 mRNA的穩定[78]。 而Activator protein-1簡稱AP-1,屬於轉錄因子(transcription factors) 其中ㄧ個重要成員,其組成包括ATF (ATF2、LRF1/ATF3、B-ATF、 JDP1與JDP2)、Fos (c-Fos、FosB、Fra-1與Fra-2)、Jun (c-Jun、JunB與 JunD)與Maf (c-maf、MafA、MafB、MafG/F/K與Nrl),這些蛋白會調 控細胞之增生與凋亡[79]。在轉錄(transcription)之過程中,AP-1的活 化會受到MAPKs調控,尤其是JNK與p38都會使c-Jun磷酸化而產生活 性,造成許多特定基因表現,如iNOS與細胞激素的產生[66]。此外, 對於活化小膠質細胞產生相關發炎因子,MAPKs訊息傳遞路徑皆扮. 18.
(20) 演著很重要的角色[80]。而ox-LDL對於平滑肌細胞所造成的細胞凋亡 或增生,皆發現與MAPKs路徑的活化有相當的關係[81;82]。因此在 接下來的實驗中也將再探討此路徑對於ox-LDL的重要性。 (2)Nuclear factor-κB (NF-κB) NF-κB 為 一 種 轉 錄 因 子 , 首 先 被 發 現 於 B 細 胞 內 可 以 和 immunoglobulin κ light-chain enhancer上一小段DNA序列結合而命名 [83]。之後便發現在所有細胞中皆有此轉錄因子的表現,且和許多蛋 白基因的調控有著密切的關係。NF-κB主要由一系列稱作REL大家族 的 protein 所 組 成 , 使 得 不 同 NF-κB 有 著 共 同 的 REL domain (REL homology domains, RHD)[84],這段約300個胺基酸所組成的區域提供 NF-κB進行DNA結合、或和抑制性蛋白IκB (inhibitor of NF-κB)交互作 用的位置。其主要組成物分為7種:RELA/p65, c-REL, RELB, p105, p50, p100及p52,其中只有RELA/p65, c-REL, RELB具有carboxy-terminal transactivation domain (TAD) 。 所 構 成 的 NF-κB 其 中 之 ㄧ 可 以 為 RELA/p65, c-REL, RELB, p50或p52,而另外再搭配相同(homodimer) 或不同(heterodimer)之REL家族,於RHD上組合而成的一個二元體 (dimer) , 最 常 見 的 NF-κB 種 類 為 RELA/p65 及 p50 所 形 成 之 heterodimer。因此藉由不同組成之NF-κB,其所能調控之蛋白質基因 也就有其特異性,如RELA–RELA homodimer可以調控IL-8、type-VII collagen、intercellular adhesion molecule-1,RELA/p65–p50可以調控其 自身之抑制性蛋白(inhibitor of NF-κB α, IκB-α)、IL-8、IL-6、TNF-α、 granulocyte-macrophage. colony. stimulating. factor. (GM-CSF) 、. human-immunodefficiency virus-1 (HIV-1)、MMP等之表現。而這些蛋. 19.
(21) 白質的表現不論在正常生理(如免疫、細胞之存活、cell cycle、細胞黏 附等)或疾病上(關節炎、癌症、發炎)均扮演了很重大的角色。 NF-κB在平常不受刺激的細胞中會和其抑制性蛋白IκB-α結合, 而所形成之complex可以在核中與細胞質中來回穿梭(shuttle),不過由 於穿出核外之效率較大,因此淨結果仍使complex大都在細胞質中 [85]。IκB-α可以遮蔽NF-κB上之nuclear localization signal (NLS),使 得NF-κB無法translocate進入核內與DNA結合。NF-κB可以被許多刺激 而活化,這包括各種pro-inflammatory cytokines、生長因子、DNA damaging agent及病毒蛋白等[85]。而藉由TNF-α所刺激之NF-κB活化 路徑已被徹底研究出來:當接受TNF-α的刺激後,各種不同的接合蛋 白(adaptor)將會由細胞質中被吸引至細胞膜[86],隨後便會吸引並活 化由NF-κB essential modulator (NEMO, 又稱IKKγ)、IKKα (ΙΚΚ1)及 ΙKKβ (ΙΚΚ2) kinase所組成的IκB-kinase (IKK) complex[87],活化後之 IKK complex會磷酸化IκB-α Ser32及Ser36之位置,而使IκB-α被標記 上ubiquitin,隨後便會被26S proteasome所水解,因而使NF-κB上之 nuclear localization signal (NLS)曝露而translocate進入細胞核中。在核 內之NF-κB會佔住一個或多個含有一般性(consensus) κB-enhancer之 序列(5’-GGGRNYYYCC-3’,R為purine,Y為pyrimidine,而N為任意 核苷酸),招來並結合許多co-activators而刺激目標基因的表現。 另外最近則發現NF-κB本身所進行之修飾,亦可影響其所調節之 基因表現。這主要可分為NF-κB本身直接進行的後轉譯修飾(post translation modification)以及其目標基因上之histone所進行之修飾;這 些作用在核內所造成的結果可以決定NF-κB轉錄反應之強度及持續 時間(duration),其中有RELA/p65之磷酸化或acetylation、目標基因之. 20.
(22) histone磷酸化或acetylation[84]。舉例來說:(1) 目前已知RELA/p65 之磷酸化對於其抑制型蛋白IκB-α的親和力較低[88],p65之磷酸化也 可 以 更 有 效 地 迫 使 transcriptionally repressive HDAC (histone deacetylase) complexes離開以和目標DNA結合(如p50-HDAC1,於平時 未刺激狀時其會和DNA上之κB enhancer binding site結合阻止其啟動) [89] 。 (2) 磷 酸 化 RELA/p65 可 加 強 其 和 p300/CBP [CREB (cyclicAMP-response element)-binding protein]之結合[90],而p300/CBP可當 作轉錄因子及transcriptional activator之間的橋樑,促進基因之表現, 屬於transcriptional co-activators。(3) 在p65不同胺基酸上進行磷酸化 可以造成不同transcriptional cofactors之吸引而有不同之基因表現。 最後核內活化之NF-κB會在適當作用後被自身所轉錄合成出的 IκB-α結合而帶出核外,以終結NF-κB之轉錄作用。此外有文獻指出, ox-LDL對於NF-κB的訊息傳遞路徑有著雙面的影響,本身能夠活化 NF-κB路徑,而引發proimflammatory基因的表現,另一方面,也能抑 制經由發炎物質(如:LPS)所誘發的NF-κB[91]。而已知NF-κB的活 化,在LPS引發iNOS表現的反應中扮演相當重要角色[92]。. 六、Penytoin 之簡介 Phenytoin,結構式:C15H11N2NaO2,分子量:274.26,為治療局 部性發作與全身性強陣攣性發作最有效藥物之一。主要作用機制為抑 制細胞膜上鈉離子的流動[93],而達到神經細胞膜的穩定,進而降低 腦部的抽畜及延長心臟房室傳導的效果。此外,phenytoin亦可以誘發 肝臟cytochrome P450氧化酵素的活性[94]。由於phenytoin與血中白蛋 白的結合率相當高,也因此在低白蛋白血症(hypoalbuminemia)、腎臟. 21.
(23) 衰竭、肝衰竭及併用其他可能延長phenytoin代謝的藥物情況下,較易 產生毒性。而中毒的嚴重性依據其血中濃度的高低有所不同,當濃度 高達50-70 mg/l時,便可能造成死亡。因此在使用phenytoin上須做例 行性的藥物濃度監測,尤其是使用在預防頭部創傷或接受外科手術後 的抽畜情況[95]。另有文獻指出,phenytoin能夠改善多發性硬化症 (multiple sclerosis)及腦脊髓炎(encephalomyelitis)所造成的神經性發炎 反應[96],因此,本實驗將以此藥物對於小膠質細胞活化的影響做進 一步的機轉探討。. 七、研究目的 無論是在神經發炎性疾病或是神經退化性疾病發展進程上,小膠 質細胞的活化於這些病程中都扮演著舉足輕重的角色。而當小膠質細 胞活化的過程中,則會釋放大量 NO 及 cytokines (如:TNF-α、IL-1、 IL-6)等發炎物質,這些發炎介質已被證實與一些神經發炎性疾病 (如:多發性硬化症)有關。氧化態低密度脂蛋白不僅對於動脈粥狀硬 化的生成及惡化具嚴重的影響力,與 endotoxin (如:LPS)皆會刺激或 誘導 iNOS 產生,但過度的 iNOS 產生時,便會造成各種不同的生理 與病理傷害。故本篇論文則是以小鼠小膠質細胞(BV-2)為實驗細胞, 利用氧化態低密度脂蛋白(ox-LDL)與 LPS 來探討 microglia 釋放 NO 以及表現 iNOS 的作用機轉,並藉以了解中樞性藥物 phenytoin 對 LPS 刺激所誘發的 iNOS 活性及表現之影響性。且更進一步地研究 phenytoin 在細胞訊息傳遞路徑中,如:NF-κB 機制之影響性。藉由 這些研究來讓目前醫療對於一些神經發炎性及退化性疾病的治療有 更近一步的幫助。. 22.
(24) 八、實驗設計 本實驗利用小鼠小膠質細胞株 murine microglial cell line (BV-2) 為本實驗之細胞株,以氧化態低密度脂蛋白或細菌內毒素當刺激劑來 處理細胞,另一方面亦會加入不同濃度之中樞性藥物 phenytoin,利 用藥理或分子生物性實驗方法觀察刺激劑及藥物對於所誘發的 iNOS 活性表現之作用機轉。. 23.
(25) 貳、材料與方法 實驗材料: 一、實驗細胞 小鼠小膠質細胞株 murine microglial cell line (BV-2)由台灣大學林琬 琬教授所提供,為帶有 v-raf/v-myc oncogene 的反轉錄病毒所感染 primary microglia cells 所得的細胞株,BV-2 已被證實其細胞型態與功能和 primary microglia cells 相似[97],因此本實驗以 BV-2 cell line 探討其 ox-LDL 及 phenytoin 對 iNOS 表現之訊息傳遞機制。. 二、實驗藥品 1. 下列藥品為 Amersham 公司(Piscataway, NJ)之產品: Protein molecular weight markers (Prestain markers) Triton X-100 2. 下列藥品為 Amersham Life Science 公司(Cleveland, OH)之產品: Sodium dodecylsulfate (SDS) Tris (hydroxymethyl)-aminomethane (Tris-base) 3. 下列藥品為 Bio-Rad 公司(Hercules, CA)之產品: Agarose Ammonium persulfate (APS) Glycine 24.
(26) Protein assay dye reagent concentrate 4. 下列藥品為 Gibco BRL 公司(Grand Island, NY)之產品: Fetal bovine serum (FBS) Penicillin/ Streptomycin/ L-Glutamine Dulbecco’s Modified Eagle Medium (with phenol red) Dulbecco’s Modified Eagle Medium (without phenol red) Total RNA isolation reagent (TRIzol Reagent) 5. 下列藥品為 Calbiochem 公司(La Jolla, CA)之產品: PD98059, ERK inhibitor SP600125, JNK inhibitor LY294002, phosphatidylinositol 3-kinase inhibitor SB203580, p38 inhibitor 6. 下列藥品為 Sigma 公司(st. Louis, MO)之產品: Aprotinin Albumin, Human Anisomycin, p38 activator Brij 35 solution (30 % w/v) Bromophenol blue Albumin, Bovine Bovine serum albumin (BSA) β-Mercaptoethanol. 25.
(27) Brilliant Blue G-Colloidal Concentrate Brij 35 solution (30 % w/v) Bromophenol blue Coomassie brilliant blue Diethyl pyrocarbonate (DEPC) Dimethyl sulfoxide (DMSO) Dithiothretiol (DTT) Ethylenediaminetetraacetic acid (EDTA·2Na) Epoxide, Cholesterol 5α, 6α-epoxide, (C27H46O2) 7-Keto, 5-cholesten-3β-ol-7-one, (C27H44O2) Isoamyl alcohol Lipopolysaccharide (LPS) Leupetin N-(2-Hydroxyethyl) piperazine-N’-(2-ethanesulfonic acid) (HEPES) Phenylmethylsulfonyl fluoride (PMSF) Phenytoin, (C15H11N2NaO2) Sodium orthovandate Sodium pyrophosphate Tetrazolium dye 3- (4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT) Triol, Cholestane-3β, 5α, 6β-triol, (C27H48O3) 7. 下列藥品為 Pharmacia Biotech.公司(Uppsala, Sweden)之產品: Glycerol. 26.
(28) Polyoxyethylenesorbitan monolaurate (Tween 20) N, N, N, N, -Tetramethylethylethylenediamine (TEMED) 8. 下列藥品為 Wako 公司(Osaka, Japan)之產品: Acetic acid Calcium chloride, Dihydrate Chloroform Ethanol Glycerol Methanol Sodium Hydrogen Carbonate Sodium Hydroxide 9. 下列藥品為 MDBio Inc.公司(Taipei, Taiwan)之產品: 40 % Acrylamide/Bis solution DNA markers TBE solution 5X 10. 下列藥品為其他公司之產品: 2-Propanol (Showa) EtBr:Ethidium Bromide (BDH) Gelatin (Kanto, Japan) Dynasore, dynamin inhibitor Sodium fluoride (Fluka). 27.
(29) 三、套組試劑 (Kit) Enhanced chemiluminescence (ECL) Western blotting detection reagent NE-PERTM nuclear and cytoplasmic extraction reagents (PIERCE, USA) Lipid Hydroperoxide Assay Kit (Cayman chemical, USA) Nitric Oxide Quantitation Kit (Active Motif, USA) SUPERSCRIPT One-Step (Invitrogen, USA). RT-PCR. with. PLATINUN. Taq. 四、抗體 (Antibody) iNOS Rabbit monoclonal Antibody (Santa Cruz, USA) p38 MAP Kinase Mouse monoclonal Antibody (Cell Signaling, USA) Phospho-p38 Rabbit polyclonal Antibody (Cell Signaling, USA) Phospho-ATF-2 Rabbit polyclonal Antibody (Cell Signaling, USA) ATF-2 Rabbit monoclonal Antibody (Cell Signaling, USA) α-tubulin Mouse monoclonal Antibody (NeoMarkers, USA) IkB-α Rabbit polyclonal Antibody (Santa Cruz, USA) Anti-mouse Ig, HRP linked whole antibody (Amersham, USA) Anti-rabbit Ig, HRP linked whole antibody (Amersham, USA). 五、引子 (Primer) 28.
(30) 1. 購自鉅生(Genemed Synthesis, USA)公司: iNOS sense: 5’-GTGCTGAAGGACACACTAAAGAAGA -3’ iNOS antisense: 5’-TTGCCATCCTCTCAAAGTTGTAGG -3’ 2. 購自生工(MDBio Inc.)公司: β-actin sense: 5’-TGGAATCCTGTGGCATCCATGGAAAC-3’ β-actin antisense: 5’-TAAAACGCAGCTCAGTAACAGTCCG-3’. 六、耗材 Biodyne® B Precut Nylon Membranes (PIERCE, U.S.A.) Blotting paper (Amersham, U.S.A.) Cassette (Okamoto, Japan) Developer and replenisher (Kodak, U.S.A.) Fixer and replenisher (Kodak, U.S.A.) Polyvinylidene fluoride microporous membrane (PVDF; Hybond-P, Amersham, U.S.A.) Scientific Imaging film (Kodak, U.S.A.) Lab-Tek® 8 well chamber slide system (Nalge Nunc International U.S.A.) 6 well plates、12 and 24 well plates、75 cm2 culture flask 等細胞培 29.
(31) 養耗材(Costar, USA or Orange, Europe) Eppendorf、96 well-ELISA plates、cuvette 及 0.2 ml 薄壁微量 PCR 管等實驗耗材(Corning , U.S.A.)。. 30.
(32) 實驗方法: 一、小膠質細胞株 BV-2 之培養 (murine microglial cell line) BV-2 cells 為 murine microglial cell line,此細胞株由 Dr. V. Bocchini at University of Perugia (Perugia Italy)所建立,本實驗由台灣大學林琬 琬教授所提供。將此細胞株之抗凍管由液態氮中取出,快速置於 37℃ 之水浴槽(water bath)中解凍 1 分鐘後,迅速移到無菌操作台(laminar flow)內取出細胞放入含有 4~6 ml Dulbecco’s modified Eagle medium (DMEM)培養液之 75 cm2 培養瓶中,此 DMEM 培養液外加 10 %去活 性之胎牛血清(heat-inactivated fetal bovine serum, FBS)、2 mM 之 L-glutamine、100 U/ml penicillin 及 100 μg/ml streptomycin。取 20 μl 以細胞計數器(Hemocytometer)計算細胞濃度並加入適量 DMEM 培養 液,使其細胞濃度介於 4 × 105 ~ 1 × 106 cells/ml。將培養瓶置於 37℃ 恆溫箱(incubator)中,以條件 95 % air 及 5 % CO2 的混合氣體,濕度 70 %的環境中培養。當細胞增殖至接近飽和時,以細胞計數器測量細 胞之數量,約 1 × 106 ~ 2 × 106 cells/ml 時,便以 1 : 1 的比例稀釋做細 胞之繼代培養(subculture)。因 BV-2 細胞為半貼附型細胞,。故更換 時需先將培養液吸掉,再加入適量 DMEM 培養液至 75 cm2 培養瓶 中,以敲打培養瓶底部以及用 pipette 溫和地以培養液沖下細胞後, 將含細胞之培養液吸至 50 ml 離心管中,在室溫下以轉速 1000 rpm (Kubota Corporation®, 5910)離心 5 分鐘。離心後,吸除上清液再加入 新鮮培養液培養;大約每隔二至三天便需更換新鮮培養液。細胞於實 驗前需更換成含 1 % FBS 之 DMEM。. 二、低密度脂蛋白之純化(Low-Density Lipoprotein) 31.
(33) 實驗前須先配製兩種密度之 NaBr buffer,分別為 112.9%及 107.1%,首先將血液 60 ml 平均分入試管中,並加入等比例的 Sodium Citrate (human plasma : Sod. Citrate = 9 : 1),於室溫下以轉速 1710 g 離心 10 分鐘。離心後,將最上層的 PRP (platelet rich plasma)平均分 入六個試管中,於室溫中再以轉速 3045 g 離心 10 分鐘。最後取得上 層較佳的 PPP (platelet poor plasma),於八支高速離心管中各加入 4 ml PPP、密度 107.1%之 NaBr 1 ml 以及 0.2 M 之 EDTA‧2 Na 50 μl,混 合均勻後置於冰上,並秤重以便於高速離心時平衡之擺放,於 4℃下 以轉速 41K rpm (Beckman L-90K Ultracentrifuge)離心 16 小時。離心 後,用 200 μl pipette 以旋轉方式將高速離心管中上層乳白色的 VLDL 吸除,每管吸除 2 ml VLDL 後,加入密度 112.9%之 NaBr 2 ml,混合 均勻後置於冰上,並秤重以便於高速離心時平衡之擺放,於 4℃下以 轉速 41K rpm (Beckman L-90K Ultracentrifuge)離心 20 小時。離心後, 用 200 μl pipette 以旋轉方式取得高速離心管中上層之黃色懸浮液 LDL,將每管取得約 1 ml LDL 置於有蓋試管中存放,收集完成後, 將 LDL 放入濕潤之透析膜中,用夾子夾住並綁住浮標,放入 4 L 有 加入 EDTA‧2 Na 之 PBS (KH2PO4 0.2 g/L、KCl 0.2 g/L、Na2HPO4 1.15 g/L、NaCl 8 g/L、EDTA‧2 Na 2.9776 g/4L)中,於加入 EDTA‧2 Na 之 PBS 中透析三次,每次 24 小時,再於無加入 EDTA‧2 Na 之 PBS 中透析兩次,每次 12 小時。透析完成後,將 LDL 取出並用 0.22 μm filter 過濾之,將過濾完之 LDL 以試管充氮氣避光儲存於 4℃。. 三 、 氧 化 態 低 密 度 脂 蛋 白 之 配 製 (Oxidized Low-Density Lipoprotein). 32.
(34) 氧化低密度脂蛋白前須先定量其濃度,先將 2 mg/ml 之 albumin bovine (BSA)與 0.2 N NaOH 反應 10 分鐘並稀釋成 1 mg/ml (取 150 μl 之 2 mg/ml BSA 與 150 μl 之 0.2 N NaOH 充分混合)置於 eppendorf 中, 再以二次水分別稀釋成濃度為 0.5 mg/ml、0.2 mg/ml、0.1 mg/ml,做 為 standard。同樣地,LDL 也必須與 0.2 N NaOH 反應 10 分鐘並稀釋 成 1/2 原液濃度 (取 100 μl 之 LDL 與 100 μl 之 0.2 N NaOH 充分混合) 置於 eppendorf 中,再以二次水稀釋成 3/8 原液濃度。將兩種濃度之 LDL 連同標準品各取 100 μl 加入玻璃管中,並加入 2 ml 之 Dye reagent concentrate (Bio-Rad)稀釋液 (Bio-Rad : d2H2O = 1 : 3)均勻混合並反 應 5 分鐘,再從中取定量置於 cuvette 中,接著使用 UV/VIS 光譜儀 (UV/VIS Spectrophotometer,V-530)以 595 nm 之波長偵測每一個 cuvette 之吸光值,所得之吸光值以線性迴歸方式換算成濃度。測得 LDL 之濃度後,於每試管中加入 1 ml 之 LDL,以 PBS 稀釋成 0.5 mg/ml,再加入 10 μl/ml 之硫酸銅溶液 (6.24 mg/50ml d2H2O)混合均 勻,於 37℃下以銅與空氣搖晃氧化 24 小時。氧化完成後,將 ox-LDL 取出並用 0.22 μm filter 過濾之,將過濾完之 ox-LDL 以試管充氮氣避 光儲存於 4℃。另一方面,為了區別 ox-LDL 中的小分子與整體的作 用,因此將氧化後的 ox-LDL 利用 Amicon Ultra-4 Centrifugal Filter Devices-10,000 MWCO (molecular weight cut-off 10 KDa)在室溫下以 轉速 4000 g 離心 10 分鐘,收集下方的過濾液 (<10 KDa),以試管充 氮氣避光儲存於 4℃。. 四、BV-2 存活率測定(MTT assay) 此實驗目的是為了測試藥物是否對於 BV-2 具有細胞毒性,以得. 33.
(35) 知其抑制效果是否起因於其細胞之毒性作用。所以利用存活細胞的粒 線體酵素 dehydrogenase 將 MTT 試劑還原成 formazan 紫色結晶,再 用 DMSO 將結晶溶解,亦即藉由細胞之代謝活性來作為細胞是否存 活之依據,而測得之溶解吸光值等於相對細胞存活率,故存活細胞越 多,其吸光值越高。針對 BV-2 細胞存活率的測定須在實驗前先將培 養液更換成含 1 % FBS 之 DMEM (without phenol red),將定量的 BV-2 細胞(2 × 105 cells/ml)分置於 12 well 的培養盤,放入 incubator 內 24 小時後,以不同濃度的藥物處理細胞培養 22.5 小時後,加入 0.55 mg/ml MTT,於 37℃下 incubation 1.5 小時,再將每 well 內的液體吸 至 eppendorf,於室溫下以轉速 1431 g 離心 10 分鐘。離心後,去除上 清液,加入 1 ml dimethylsulfoxide (DMSO)到每個 eppendorf 中,混合 均勻後,再將液體加回到 12 well 中,於室溫下避光輕搖 15 分鐘,待 well 內殘餘細胞完全溶解後,再從每 well 中各取出 50 μl 置入含有 150 μl DMSO 之 96 well-ELISA plates,充分均勻後用酵素免疫分析儀 (MRX microplate reader, USA),以 550 nm 之波長偵測每一個 well 之 吸光值。而存活百分率之運算如下: 加藥處理之吸光値 Resting 之吸光値. × 100%. 五、細胞內蛋白質分離與定量(Intracellular protein isolation and quantity) 將 實 驗 處 理 後 之 細 胞 , 以 外 加 protease inhibitor (10 μl/ml aprotinin、1 mM PMSF 和 10 μl/ml leupeptin)及 2 mM Dithiothreitol (DTT) 之 lysis buffer (50 mM HEPES、5 mM EDTA‧2 Na、50 mM. 34.
(36) NaCl 與 1 % Triton X-100)將細胞溶破,在 4℃下靜置 30 分鐘,每 5 分鐘 vortex 15 秒,隨後於 4℃下以轉速 12879 g 離心 20 分鐘,吸取 上清液,定量蛋白質。若進行有關 IκBα等磷酸化蛋白質之測定時, 其 lysis buffer 則必須再多加入 phosphatase inhibitor (10 mM sodium fluoride、1 mM sodium orthoranadate 和 5 mM sodium pyrophosphate)。 蛋白質定量前,先將 2 mg/ml 之 albumin bovine (BSA)稀釋成 0.1 mg/ml (取 40 μl 之 2 mg/ml BSA 與 760 μl 二次水充分混合)置於 eppendorf 中,再以二次水分別稀釋成濃度為 5 μg/ml、10 μg/ml、15 μg/ml、20 μg/ml、25 μg/ml、30 μg/ml、35 μg/ml,做為 standard。並取蛋白質樣 品 2 μl 與二次水 798 μl 均勻混合,連同標準品全數加入 200 μl Dye reagent concentrate (Bio-Rad)快速均勻混合,再從中分別取 200 μl 置 於 96-well ELISA plates 中,接著使用酵素免疫分析儀(MRX microplate reader, U.S.A.)以 550 nm 之波長偵測每一個 well 之吸光值。所得之吸 光值以線性迴歸方式換算成濃度,測定後,樣品分裝保存於-80℃備 用。. 六、西方點墨法 (Western blotting) 將實驗處理取得之已定量的細胞內蛋白質成份以 5 : 1 的體積比 例加入 6 倍 sample loading dye (350 mM Tris-base、30 % Glycerol、350 mM SDS、175 μM Bromophenol blue 與 600 mM DTT 調成 pH 6.8)充 分混勻後,95℃加熱 5 分鐘,使蛋白質 denature 後,快速置於冰上至 少 5 分鐘,以免回溫過程中酵素影響蛋白質,最後在 4℃下以轉速 2236 g 離心 5 分鐘後備用。再以 10 % polyacrylamide gel 於 running buffer (25 mM Tris-base、192 mM Glycine 和 0.1 % SDS 調成 pH 8.3)下,每行分. 35.
(37) 析 80 μg (± 5 μg) 之蛋白質(針對 iNOS),並以 125 V/80 mA 進行電泳 分離。隨後將膠片置於 transfer buffer (1 M Tris-base、20 % methanol 和 150 mM glycine 調成 pH 8.3)中,以 70 V/300 mA 進行電泳轉漬 3 小 時 , 使 膠 片 上 之 蛋 白 質 轉 移 至 Nitrocellulose membrane (NC ; Hybond-C)或 Polyvinylidene fluoride microporous membrane (PVDF ; Hybond-P)表面,隨後將轉漬膜浸潤在 25℃下的 blocking buffer (5 % non-fat milk、10 mM Tris-base、100 mM NaCl 和 0.1 % Tween 20 調成 pH 7.5)中,約 1 小時後,以 TBST (10 mM Tris-base、100 mM NaCl 和 0.1 % Tween 20 調成 pH 7.5)清洗轉漬膜三次,每次 10 分鐘,之後 加入一級抗體( primary antibody),(如:iNOS、phospho-p38、total-p38、 total IκB-α、phospho-ATF-2、α-tubulin 等),於室溫中搖晃作用 2-4 小時。再用 TBST 清洗轉漬膜四次,每次 7 分鐘,之後再加入標記有 horseradish perosidase (HRP)的二級抗體(secondary antibody),於室溫 下反應 1~2 小時,再以 TBST 清洗轉漬膜四次,每次 7 分鐘。最後使 用冷光反應劑 Enhanced chemiluminescence (ECL) Western blotting detection reagent 使底片感光,以用來偵測蛋白質量的表現情形。最後 將成像後的底片掃瞄輸入電腦,以影像分析軟體(Bio-1D version 99) 做分析處理,以 resting 亮度當作 1,其餘 band 之亮度以其相對倍數 表現之。. 七、RNA 萃取與定量(RNA extract and quantity ) 實驗前將培養液更換成含 1 % FBS 之 DMEM (without phenol red),將定量的 BV-2 細胞(5 × 105 cells/ml)分置於 6 well 的培養盤中, 並以不同濃度的 ox-LDL 處理細胞,培養 6 小時後離心,吸除上清液,. 36.
(38) 接著加入 1 ml TRIzol reagent (total RNA isolation reagent, Invitrogen) 溶破細胞,用 tip 來回吸排數次並快速移至 1.5 ml eppendorf 中,室溫 下靜置 5 分鐘待細胞溶解,在 4℃下以轉速 12035 g 離心 10 分鐘。離 心後,取出 900 μl 上清液到新的 eppendorf,靜置 5 分鐘後,每管加 入 200 μl chloroform,劇烈震盪 15 秒,再靜置於室溫下 3 分鐘。接下 來在 4℃下轉速 10432 g 離心 15 分鐘。離心後,小心吸取上清液至新 的 eppendof 中,並加入 0.5 ml 的 isopropyl alcohol 於室溫下靜置 10 分鐘,使 RNA 析出,再以 4℃轉速 10432 g 離心 10 分鐘。離心後, 將上清液去除,留住管底的凝膠樣沉澱物(gel-like pellet)。再加入 1 ml 75 %之酒精(以 0.01 % DEPC-H2O 及 99.5 %無水酒精配製)清洗凝膠 樣沉澱物。將每管輕微振盪後,再以 4℃轉速 7085 g 離心 5 分鐘。離 心後立刻風乾凝膠樣沉澱物 20-30 分鐘,再以 70 μl DEPC-H2O 溶解 RNA。取少量至 microcuvette 中,用 GeneQuant Pro 分析儀以 260/280 nm 之波長來偵測樣品之純度及濃度。測定後將 RNA 溶液分裝置於 -80℃中保存備用。. 八、反轉錄-聚合酵素連鎖反應(Reverse transcriptionpolymerase chain reaction,RT-PCR) 利 用 Super Script One-Step RT-PCR with PLATINUN Taq (Invitrogen)之套組,依照說明書指示進行 RT-PCR 實驗。 Component. Volume/50 μl. 2X Reaction Mix Template RNA 37. Final concentration. 25 μl. 1X. 2 μl. 10 pg - 1 μg.
(39) Sense Primer. 0.5 μl. 0.2 μM. Anti-sense Primer. 0.5 μl. 0.2 μM. RT/Taq(RT enzyme) Mix. 1 μl. 0.2 μM. Autoclaved dH2O. 21μl. 將以上所列之 reagents 依序加入至 0.2 ml 薄壁微量 PCR 反應管, 《注意:上述過程均在冰上操作,且反應管需事先滅過菌並 pre-chilled》輕微 vortex 混合均勻,然後在冰上輕敲,使 vortex 時沾 於管壁上的 sample 沉降,最後置入 PCR 反應器(GeneAmp® PCR System 2400)中依照下列的反應條件來進行 RT-PCR: (1) cDNA 的反轉錄合成 iNOS : 50℃,30 分鐘 → RNA 反轉錄成 cDNA 94℃,2 分鐘 → 與 RNA 分離成為單股 cDNA β-actin : 50℃,30 分鐘 → 94℃,2 分鐘 (2) PCR amplification (放大) iNOS : 95℃,20 秒 → denature template,確定 cDNA 打開 57℃,30 秒 → anneal primers,primers 黏上 cDNA 72℃,30 秒 → cDNA synthesis,複製 cDNA 總反應 35 cycles 38.
(40) β-actin : 95℃,20 秒 → 47℃,30 秒 → 72℃,20 秒 總反應 30 cycles (3) Final extension 72℃,5 分鐘 → 讓最後一個 cycle 的 cDNA 完成 降到 4℃ → 避免 cDNA 壞掉 最後將 cDNA 存放於 4℃中備用,-80℃可長期保存. 九、膠體電泳(Agarose gel electrophoresis) 將實驗處理後取得放大之 cDNA 產物以 4 : 1 的體積比例加入 5 倍 sample loading dye (25 % Glycerol 和 0.25 % Bromophenol blue)充分 混合均勻,將每一管 sample loading 在 1.5 %洋菜膠片(含 0.5 μg/ml ethidium bromide,具螢光染色 cDNA 之能力),於 0.5 % TBE buffer 下,以 150 V/40 mA 條件進行電泳分離。iNOS 與β-actin 之 mRNA 經 由反轉錄後之 cDNA 分別為 311 bp 與 288 bp。最後將膠片置於影像 分析系統(Vilber Lourmat,France),以 CCD 拍攝並將圖檔存入磁片 中,再將磁片所存之影像於電腦中以影像分析軟體(Bio-1D version 99) 進行影像分析處理。iNOS cDNA 之相對含量需先以β-actin 做適當校 正,實驗結果的表示以相對此數值表示,最後以 resting 亮度當作 1, 其餘 band 之亮度以其相對倍數表現之。. 十、細胞核內及核外蛋白質分離與定量(Nuclear extracts preparation,NE-PERTM) 39.
(41) 將定量的 BV-2 細胞(3 × 105 cells/ml,8 ml/well)種植於 10 cm dish 中[細胞數固定 2 × 106 cell 時,packed cell volume (PCV)大約 20 μl], 經過處理後收集細胞於離心管,去除上清液,加 500 μl cold-PBS 清 洗 pellet 並移至新的 0.5 ml pre-chilled eppendorf,然後在 4℃下以轉速 500 g 離心 3 分鐘。離心後,去除上清液並盡量吸乾,同時檢查 packed cell volume (PCV),先用手將 pellet 打散後(click)再加入 200 μl ice-cold CER-I,並以最大振數 vortex 15 秒,在 4℃下靜置 10 分鐘,加入 11 μl ice-cold CER-II,vortex 5 秒,4℃下靜置 1 分鐘,再 vortex 5 秒,之 後於 4℃下以轉速 16000 g 離心 5 分鐘。離心後,上清液即為 cytoplasmic extracts。將上清液移出後,click eppendorf 使 pellet 鬆散, 再加入 100 μl ice-cold NER,每 10 分鐘 vortex 15 秒,總共 40 分鐘, 然後在 4℃下以轉速 16000 g 離心 10 分鐘,上清液即為 nuclear extracts。最後將上清液分裝到一新的 pre-chilled eppendorf 並保存於 -80℃,並做蛋白質定量。. 十一、脂質過氧化(Lipid Hydroperoxide,LPO)含量測定 將氧化及未氧化之 LDL 利用 Extract R(10 mg/1.5 ml methanol)等 比例(sample:Extract R = 1:1)稀釋成適當濃度置於 2 ml pre-chilled eppendorf,vortex 混合均勻後,加入 1 ml 冰的 chloroform,再 vortex 混合均勻,然後在 4℃下以轉速 1500 g 離心 5 分鐘。離心後,取最底 層之 chloroform 約 0.7 ml 至另一新的 1.5 ml pre-chilled eppendorf,此 即為 extract。接著取 0.5 ml extract 與 0.45 ml chloroform-methanol 混 和液(chloroform-methanol = 2:1)均勻混合於新的 2 ml pre-chilled eppendorf 中,再加入 50 μl chromogen (FTS Reagent 1:FTS Reagent 2. 40.
(42) = 1:1) vortex 混於均勻後,於室溫下靜置 5 分鐘。在脂質過氧化定 量前,將 lipid hydroperoxide standard (HP)以 chloroform-methanol 混和 液分別稀釋成濃度為 0 nmol、0.5 nmol、1 nmol、2 nmol、3 nmol、4 nmol、5 nmol,做為 standard,再加入 50 μl chromogen vortex 混於均 勻後,於室溫下靜置 5 分鐘。接著分別取樣品及標準品各 1 ml 加入 石英之 cuvette 中,使用 UV/VIS 光譜儀 (UV/VIS Spectrophotometer, V-530)以 500 nm 之波長偵測每一個 cuvette 之吸光值。所得之吸光值 以線性迴歸方式換算成濃度,即可了解各管 LDL 之氧化程度。. 十二、一氧化氮(NO)含量測定 將定量的 BV-2 細胞(2 × 105 cells/ml,1 ml/well)培養於 12 well plate 不含酚紅之 DMEM 培養液中,處以藥物後 24 小時,將先前之 培養液取出 70 μl 和 20 μl reconstituted cofactors solution 、10 μl reconstituted nitrate reductase solution 混合於 96 well plate 中,以速度 150 rpm 搖晃 1 分鐘後,於室溫下避光作用 30 分鐘。再依序加入 50 μl Griess reagent A 和 50 μl Griess reagent B 於每個 well 中,於室溫下避 光作用 20 分鐘。使用 Griess reagent 反應之成色產物來測定細胞釋放 nitrite 的量。另一方面,對於 ox-LDL 處理後的培養液,則須利用 Amicon Ultra-4 Centrifugal Filter Devices-10,000 MWCO (molecular weight cut-off 10 KDa)在室溫下以轉速 4000 g 離心 10 分鐘,收集下方 的過濾液 (<10 KDa),再進行上方的步驟。最後反應完的溶液,利用 microplate reader 以 550 nm 波長測其吸光值,並藉由 sodium nitrite 為 標準曲線,以線性回歸方式計算每一 well 中 nitrite 含量。Nitrite 是 NO 的穩定氧化終產物,所以測量培養液中 nitrite 含量表示相對 NO. 41.
(43) 的含量。. 十三、免疫螢光染色(Immunofluoresence Staining) 將定量的 BV-2 細胞(2 × 105 cells/ml,300 μl/well)種植於 8 well chamber,實驗前 12 小時將培養液更換成含 1 % FBS 之 DMEM,利 用 ox-LDL 及 Anisomycin 刺激 30 與 60 分鐘,反應完後立即置於冰 上,接著用冰的 PBS 400 μl/well 清洗兩次,清洗後再加入 4 % paraformaldehyde 400 μl/well,於 4℃中反應 30 分鐘。反應完後吸除 paraformaldehyde,以室溫之 PBS 400 μl/well 清洗三次,每次 5 分鐘, 清洗後再加入 0.1 % Triton X-100 400 μl/well,於 4℃中反應 15 分鐘。 反應完後吸除 Triton X-100,以室溫之 PBS 400 μl/well 清洗三次,每 次 5 分鐘,清洗後再加入 3 % BSA 400 μl/well,於室溫中反應 30 分 鐘。反應完後吸除 BSA,以室溫之 PBS 400 μl/well 清洗三次,每次 5 分鐘,清洗後再加入一級抗體(primary antibody),(如:phospho ATF-2、 α-tubulin 等)於室溫中作用 1 小時,再用 PBS 清洗 3 次,每次 5 分鐘, 之後再加入標記有 FITC 的二級抗體(secondary antibody),於室溫下避 光反應 1 小時,再以 PBS 清洗 3 次,每次 5 分鐘。反應完後即拆掉 chamber 之分隔壁,於載玻片上滴上一滴 antifade mount media,接著 用蓋玻片緩慢放於載玻片上,以倒立顯微鏡(Nikon®, Eslipse TS100) 觀察之,觀察後避光儲存於 4℃。. 十四、質體轉染(plasmid transfection ) 將定量的 BV-2 細胞(2 × 105 cells/ml/well)種植於 6 well 中,實驗 前將培養液更換成 OPTI-MEM 1 ml,準備相同 well 數之 eppendorf 並裝入 50 μl OPTI-MEM,分別將 vector、mutant MKK3、mutant MKK6 42.
(44) 及 mutant p38 等 DNA 取 0.5 μg (DNA (μg):Lipofectamine (μl) = 1: 2)加入裝有 OPTI-MEM 之 eppendorf 中,均勻 mix 10 下。再另外準備 相 同 well 數 之 eppendorf 並 裝 入 50 μl OPTI-MEM 和 1 μl Lipofectamine,於室溫反應 5 分鐘(不可超過 25 分鐘)。將含有 DNA 之 OPTI-MEM 緩慢地沿液面加入稀釋的 Lipofectamine 中,均勻 mix 10 下後,於室溫反應 20 分鐘。接著均勻加入 well 中,將 6 well plate 置於 37℃恆溫箱(incubator)中,以條件 95 % air 及 5 % CO2 的混合氣 體,濕度 70 %的環境中反應 6 小時。反應完成後,將培養液更換成 含 10 % FBS 之 DMEM,即可進行細胞實驗。. 十五、數據分析 本實驗之數據結果皆以實驗次數之平均值±標準誤差(mean ± S.E.) 表示。根據 Student’s t-test 作為統計分析,若 P<0.05,則表示有意 義的差異。. 43.
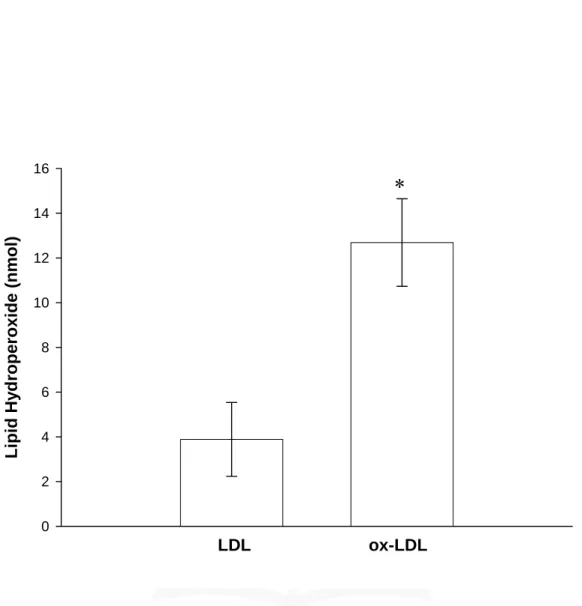
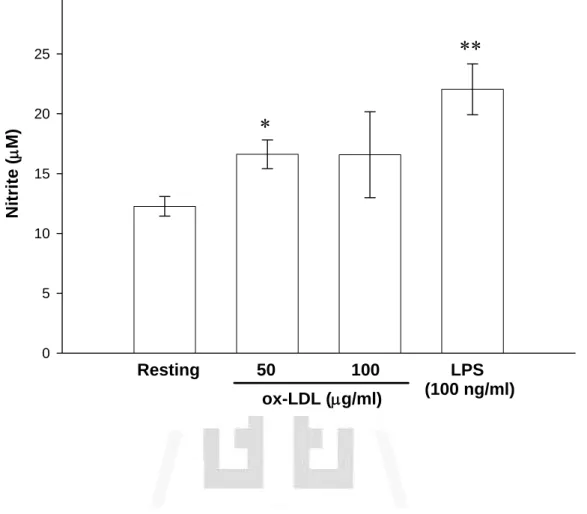
(45) 叁、結果 一、 探討 ox-LDL 對於活化小膠質細胞株 BV-2 細胞誘導 NO 合成之影響 本實驗首先是要探討 ox-LDL 是否能活化 BV-2 細胞並產生相關發 炎介質(如一氧化氮;nitric oxide, NO)。實驗所使用的 ox-LDL 主要為 銅氧化之 ox-LDL (其條件以 5 μM Cu2+在 37℃下處理 24 小時)。為了 進一步了解 ox-LDL 的氧化程度,利用 LPO assay 對各次所分出的 LDL 與 ox-LDL 進行測定,由 Fig. 1 顯示,ox-LDL 較 LDL 的氧化程度的 確有意義明顯的增加,且統計上具顯著的差異性(LDL:3.9 ± 1.7 nmole 與 ox-LDL:12.7 ± 2.0 nmole)。隨後分別以不同濃度的 ox-LDL (50 及 100 μg/ml) 處理 BV-2 細胞並培養 24 小時,取細胞上清液部分來進行 實驗,由 Fig. 2 顯示,在未加刺激劑的情形下(Resting),其上清液只 偵測到濃度為(12.3 ± 0.8 μM)的 nitrite 累積量。而以 ox-LDL (50 及 100 μg/ml)刺激 24 小時後,發現其上清液中 nitrite 累積量皆有增加,雖然 只有 ox-LDL (50 μg/ml)的刺激量在統計上有顯著的差異性,但從數據 上仍可大致看出皆有高於 Resting 的趨勢(50 μg/ml:16.6 ± 1.2 μM 與 100 μg/ml:16.6 ± 3.6 μM)。另一方面 LPS (100 ng/ml)作為正向對照 組,的確也能明顯刺激 nitrite 累積量之生成(22.0 ± 2.1 μM)。由以上 結果得知,ox-LDL 無論在 50 及 100 μg/ml 兩個濃度上皆可活化 BV-2 細胞,並且會明顯誘發 NO 的產生。. 二、 探討 ox-LDL 對於 BV-2 細胞的細胞存活率之影響 本實驗利用 MMT assay 測定 ox-LDL 對於細胞存活率之影響。由 44.
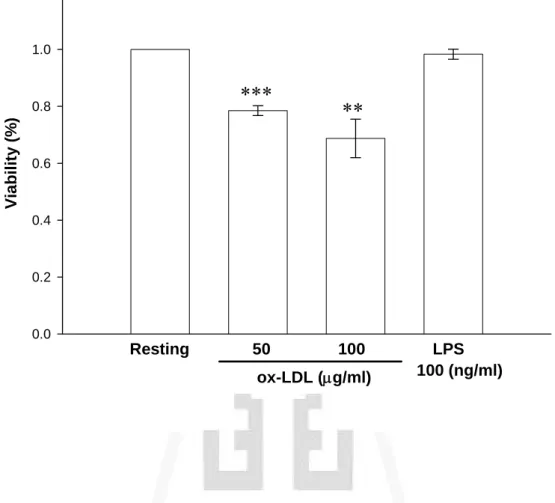
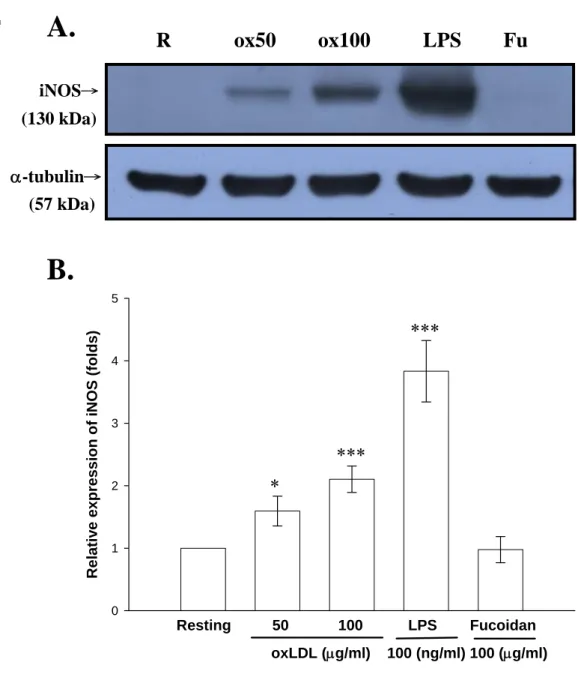
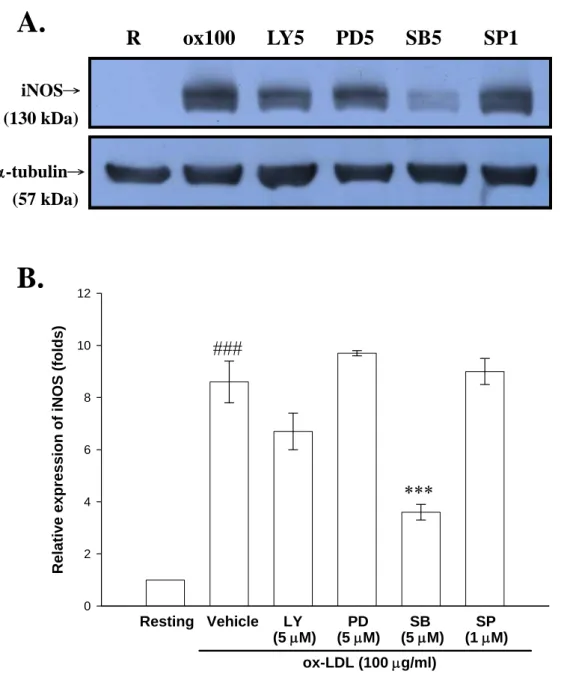
(46) 實驗結果發現,雖然 ox-LDL 分別在濃度為 50 及 100 μg/ml 的刺激 下,細胞存活率皆具有統計意義的下降,但整體而言其細胞存活率皆 可維持在約 70%(ox-LDL, 50 μg/ml:78.5 ± 1.8%及 100 μg/ml:68.8 ± 6.8%, n = 3),即 ox-LDL 並不會嚴重影響細胞的存活。因此,本實驗 所使用之 ox-LDL 濃度範圍內對細胞皆不具嚴重毒性反應。另一方 面,LPS (100 ng/ml)對於細胞存活率的影響則不具統計上的差異(98.3 ± 1.8%, n = 4)(Fig. 3)。. 三、 探討 ox-LDL 及 A 型清除接受體制效劑(Fucoidan)對於 活化 BV-2 細胞誘導 iNOS 產生之影響 由之前的實驗結果已證實 ox-LDL 的確能誘導 BV-2 細胞增加 NO 的產生,為了瞭解 ox-LDL 對 NO 的促進作用是否藉由增加其細胞內 iNOS 的蛋白質表現,我們利用西方點墨法(Western blot)以分析 ox-LDL 對細胞內 iNOS 蛋白質表現的影響。 將 BV-2 細胞(2 × 105 cell/ml)處理並培養 24 小時後,取其細胞內 萃取物(cell lysate)進行實驗,由 Fig. 4A 顯示,在未加刺激劑的情形 下(Resting),其細胞萃取物只偵測到非常微量的 130 kD 之 iNOS 蛋白 表現量(Resting,1.0 ± 0.0 fold)。而以 ox-LDL (50 及 100 μg/ml)刺激 24 小時後,發現其細胞萃取物中 iNOS 蛋白表現量有顯著的增加(Fig. 4A),並且隨著 ox-LDL 濃度的增加,iNOS 的蛋白表現量也逐漸提高, 且統計上有顯著的差異性(ox-LDL, 50 μg/ml:1.6 ± 0.2 folds 及 100 μg/ml:2.1 ± 0.2 folds, n = 3-6;Fig. 4B)。另一方面利用 LPS (100 ng/ml) 作為正向對照組,的確也能明顯刺激 iNOS 之生成(3.8 ± 0.5 folds, n = 6;Fig. 4A and B)。而在 fucoidan (100 μg/ml)的刺激方面,觀察到其 45.
(47) 細胞萃取物中 iNOS 蛋白表現量並沒有顯著的增加,即代表與未刺激 組(Resting)沒有太大差異(1.0 ± 0.2 folds, n = 3;Fig. 4A and B)。我們 也發現,即使 fucoidan 的濃度高達 200 μg/ml 時,細胞萃取物中 iNOS 蛋白表現量仍沒有顯著的增加(1.0 ± 0.2 folds, n=3)(data not shown)。 代表 fucoidan 對於 BV-2 細胞的確無法刺激其 iNOS 的蛋白表現。. 四、 探討特定訊息路徑抑制劑對於 ox-LDL 活化 BV-2 細胞 誘導 iNOS 產生之影響 由以上結果得知,ox-LDL 可以活化 BV-2 細胞增加 NO 及 iNOS 的表 現。為了進一步了解其中的刺激機制,我們使用了多種訊息抑制劑, 包括有 phosphatidylinositol 3-kinase inhibitor (LY 294002)、MAP kinase kinase (MEK) inhibitor (PD 98059)、p38 MAPK inhibitor (SB 203580) 及 JNK inhibitor (SP 600125),進而觀察 ox-LDL 誘發 iNOS 表現的訊 息傳遞路徑。由 Fig. 5A and B 顯示,以 ox-LDL 處理後可誘發 iNOS 達 8.6± 0.7 folds (n = 3)。然而各抑制劑如 LY (5 μM)、PD (5 μM)及 SP (1 μM)對於 ox-LDL 所誘發的 iNOS 表現並無明顯的影響(LY:6.7 ± 0.7 folds;PD:9.7 ± 0.1 folds 與 SP:9.0 ± 0.5 folds)。但是 SB (5 μM) 卻能明顯的抑制 ox-LDL 所誘發的 iNOS 表現(3.6 ± 0.3 folds, n = 3)。 於是初步推論 ox-LDL 可能是藉由 p38 之 Mitogen-activated protein kinase (MAPKs)的訊息傳遞路徑來以增加 iNOS 在 BV-2 細胞中的表 現。. 五、 探討各訊息路徑抑制劑對於 BV-2 細胞的細胞存活率之 影響. 46.
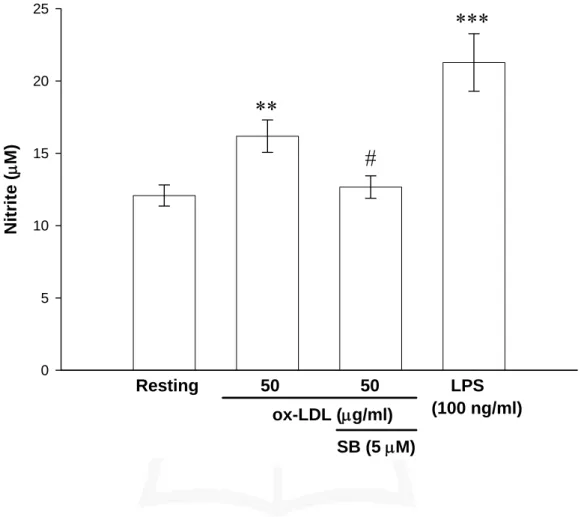
(48) 本實驗利用 MMT assay 來測定對於細胞存活率之影響。由 Fig. 6 實驗結果發現,在 ox-LDL 100 μg/ml 的刺激下,會稍微降低細胞存活 率(80.5 ± 2.9%),但不會過於嚴重,而分別加入了 LY 294002 (5 μM)、 SP 600125 (5 μM)、SB 203580 (5 μM)及 PD 98059 (1 μM)後,細胞存 活率反而有些微的上升(LY:92.7 ± 3.7%;PD:95.3 ± 2.6%;SB:94.3 ± 3.7%及 SP:94.3 ± 2.0%)。. 六、探討 SB 203580 對 BV-2 細胞中以 ox-LDL 誘發 NO 的 影響 由前面的實驗結果可以知道,p38 抑制劑(SB 203580)可明顯抑制 ox-LDL 活化 BV-2 細胞產生 iNOS 蛋白表現。因此,我們想了解 NO 的產生是否也會受到抑制劑 SB 203580 的影響。由 Fig. 7 的結果顯 示,當未給予 ox-LDL 刺激的情形下,只有偵測較少量的 nitrite 累積 量(Resting:12.1 ± 0.7 μM)。以 ox-LDL (50 μg/ml)處理 24 小時後,nitrite 的累積量很明顯地的增加,且統計上具顯著的差異性(16.2 ± 1.1 μM, n = 3)。而給予抑制劑 SB 203580 (5 μM)之後,可以觀察到 ox-LDL 所 誘發 nitrite 的累積量有明顯的受到抑制,與 ox-LDL 的刺激量相比呈 現有意義的差異(12.7 ± 0.8 μM, n = 3)。而 LPS 為正向對照組,在濃 度為 100 ng/ml 的刺激下,nitrite 的累積量的確能很明顯地被誘發出 來,與空白組(Resting)相比亦呈現有意義的差異(21.3 ± 2.0 μM, n = 3)。. 七、探討 SB 203580 對 BV-2 細胞中以 ox-LDL 誘發 iNOS mRNA 的影響. 47.
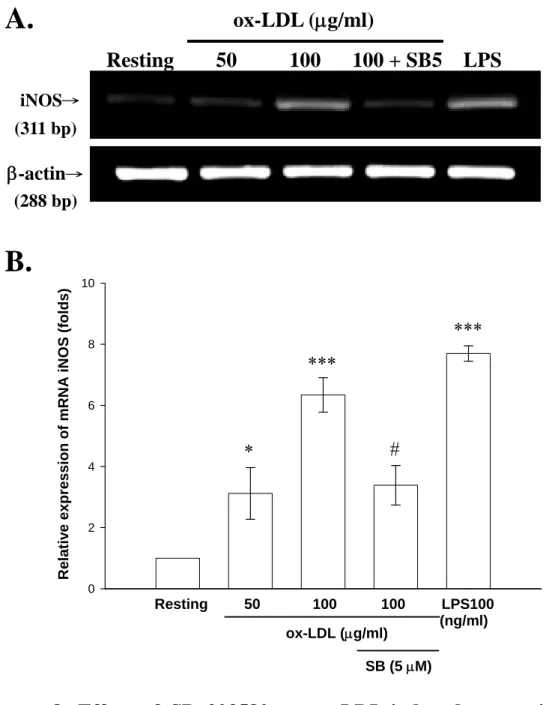
(49) 由前面的實驗結果可以知道,SB 203580 可明顯抑制 ox-LDL 所 誘發 BV-2 細胞的 iNOS 蛋白表現。因此,我們接著觀察 iNOS 基因 的轉錄過程中是否也會受到抑制劑 SB 203580 的影響。BV-2 細胞內 轉 錄 出 的 mRNA 是 經 由 萃 取 total RNA 過 程 之 後 , 以 reverse transcriptase reaction (RT-PCR)的方法完成 cDNA 分析。使用 iNOS 與 β-actin 的專一性引子(primers)來進行放大 iNOS 與β-actin 之 cDNA 產 物,並以 1.5 %的洋菜膠(agarose gel)進行電泳分析。由 Fig. 8A 的結 果顯示,分別以 ox-LDL (50 及 100 μg/ml)處理 6 小時後,iNOS mRNA 的表現皆很明顯地被誘發出來。隨著 ox-LDL 濃度的增加,iNOS mRNA 的表現量也隨之增高,與空白組(Resting)相比各別約增加為 3.1 倍及 6.3 倍(ox-LDL, 50 μg/ml:3.1 ± 0.8 folds 及 100 μg/ml:6.3 ± 0.6 folds, n = 4;Fig. 8B)。而給予抑制劑 SB 203580 (5 μM)之後,可以觀 察到 ox-LDL 所誘發 iNOS mRNA 的表現有明顯的抑制的效果(Fig. 8A)。其 SB 203580 抑制 ox-LDL 所誘發 iNOS mRNA 表現抑制百分 比約為 46 % (數值為 3.4 ± 0.6 folds, n = 4)(Fig. 8B)。而 LPS 為正向對 照組,在濃度為 100 ng/ml 的刺激下,iNOS mRNA 的表現的確能很 明顯地被誘發出來,與空白組(Resting)相比約增加為 7.7 倍(7.7 ± 0.3 folds, n = 4)(Fig. 8A and B)。至於β-actin 為 internal control,抑制劑對 其無明顯影響。. 八、 比較 SB 203580 對於 ox-LDL 及 Anisomycin 活化 BV-2 細胞誘導 iNOS 產生之影響 由前面的實驗結果可以得知,SB 203580 可明顯抑制 ox-LDL 所 誘發 BV-2 細胞的 iNOS 蛋白表現及 iNOS mRNA 的表現。為了進一. 48.
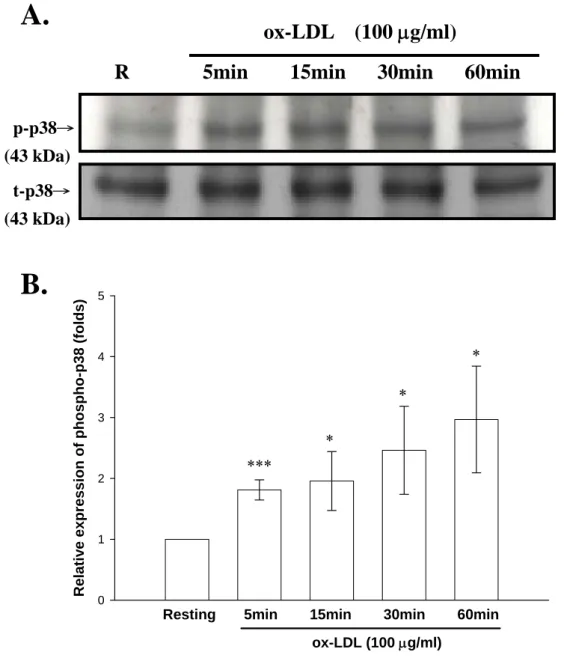
(50) 步確認其中的作用機制,我們使用已知的 p38 活化劑 anisomycin 以與 ox-LDL 的刺激做比較。由 Fig. 9 顯示,分別以 ox-LDL (100 μg/ml) 及 anisomycin (50 nM)處理 24 小時後,iNOS 的表現皆很明顯地被誘 發出來,與空白組(Resting)相比各別約增加為(ox-LDL:7.4 folds 與 anisomycin:4.2 folds)。而給予抑制劑 SB 203580 (5 μM)之後,可以 觀察到 ox-LDL 及 anisomycin 所誘發 iNOS 的表現皆有明顯的抑制的 效果。其 SB 203580 抑制 ox-LDL 所誘發 iNOS 表現抑制百分比為 53 % (SB,3.5 folds)。而抑制 anisomycin 所誘發 iNOS 表現抑制百分比 為 55 % (anisomycin,1.9 folds)。. 九、探討 ox-LDL 對於活化 BV-2 細胞引發 p38 MAPKs 訊息 傳遞路徑活化之作用 Mitogen-activated protein kinases (MAPKs)訊息傳遞過程中,必須 經過自身的磷酸化才能成為具有活性的分子,進而轉移(translocate) 進入核內,而活化相關的轉錄因子(transcription factor)(例如 AP-1、 ATF-2 等),再驅動特定基因的表現。本實驗利用西方點墨法(Western blot)評估 ox-LDL 是否會促進 p38 訊息傳遞分子磷酸化。在 Fig. 10A 顯 示 , 當 未 給 予 ox-LDL 刺 激 的 情 形 下 , 只 有 偵 測 的 很 微 量 的 phospho-p38 表現(Resting:1.0 ± .0.0 fold)。當以 ox-LDL (100 μg/ml) 刺激 BV-2 細胞後,分別在 5、15、30 與 60 分鐘的情形下測定,發現 在刺激 5 分鐘時,phospho-p38 表現量就有很明顯的增加,並且隨著 時間增長,phospho-p38 表現量也隨之增加,在 60 分鐘時達到最大量, 統計上也皆為有意義的差異(5 min:1.8 ± 0.2 folds;15 min:2.0 ± 0.5 folds;30 min:2.5 ± 0.7 folds 及 60 min:3.0 ± 0.9 folds, n = 3;Fig.. 49.
(51) 10B)。. 十、探討透析 ox-LDL(10 KDa 以下)對於活化 BV-2 細胞誘 導 iNOS 產生之影響 本實驗所使用之透析 ox-LDL,即利用 Amicon Ultra-4 Centrifugal Filter Devices-10,000 MWCO (molecular weight cut-off 10 KDa)將原 ox-LDL 所過濾而得之溶液。將 BV-2 細胞(2 × 105 cell/ml)處理並培養 24 小時後,取其細胞內萃取物(cell lysate)進行實驗,由 Fig. 11A 顯示, 在未加刺激劑的情形下(Resting),其細胞萃取物只偵測到非常微量的 130 kD 之 iNOS 蛋白表現量(Resting:1.0 ± 0.0 fold)。而以 ox-LDL (50 μg/ml)刺激 24 小時後,發現其細胞萃取物中 iNOS 蛋白表現量有顯著 的增加(Fig. 11A),且統計上有顯著的差異性(2.0 ± 0.2 folds, n = 3;Fig. 11B)。另外以 10 KDa 以下之 ox-LDL 溶液刺激 24 小時後,發現其細 胞萃取物中 iNOS 蛋白表現量同樣有顯著的增加(Fig. 11A),且統計上 有顯著的差異性,但與原 ox-LDL 刺激相比仍較低(1.5 ± 0.1 folds, n = 3;Fig. 11B)。另一方面利用 LPS (100 ng/ml)作為正向對照組,的確 也能明顯刺激 iNOS 之生成(3.1 ± 0.3 folds, n = 3;Fig. 11A and B)。. 十一、探討 epoxide、7-keto 及 triol 對於活化 BV-2 細胞誘導 iNOS 產生之影響 由前面的實驗結果得知,小分子的 ox-LDL 也能夠活化 BV-2 細胞 誘導 iNOS 的表現。因此,本實驗進一步利用三個已知的膽固醇氧化 產物,分別來觀察其對 BV-2 細胞的刺激情況。由 Figure 12A 顯示, 當 未 給 予 任 何 刺 激 的 情 形 下, 只 有 偵 測 的 很 微 量 的 iNOS 表現. 50.
數據
+7


相關文件
- In the current lesion, the epithelioid cells were positive for S100 protein and no sustentacular cells were identified, effectively ruling out PGL as the diagnosis. -
Finally, the immunohistochemical results in the present case highlights that Bcl-2 negativity in granular cells indicating an apoptotic process, CD-68 positivity in granular
Its main tool is the stem cells that are seeded on the surface of biomaterials (scaffolds), in order to create a biocom- plex. Several populations of mesenchymal stem cells are found
Histologically VX is distinguished from other lesions by presence of large numbers of foam cells, and limited to the connective tissue papillae.. The foam cells on
Diagnosis is confirmed by histopathology showing the presence of Antoni A(cellular fascicular) and Antoni B (myxoid; vacuolated) cells, nuclear palisading, whirling of cells
stained negative for a-smooth muscle actin because the main body of the tumor consisted of epithelioid or epithe- lial cells that contained few positive smooth muscle cells.. We
Stain (-) because the main body of the tumor consisted of epithelioid or epithelial cells that contained few positive smooth muscle cells. No clear ductal structures or
They could also successfully form single-cell derived colonies (Fig.. Pyogenic granuloma-derived cells. A) After the initial seeding, many cells attached to the plates in an
