行政院國家科學委員會專題研究計畫 成果報告
利用合成改良之癌症專一性啟動子進行肺癌之基因治療
計畫類別: 個別型計畫 計畫編號: NSC93-2314-B-002-237- 執行期間: 93 年 08 月 01 日至 94 年 07 月 31 日 執行單位: 國立臺灣大學醫學院外科 計畫主持人: 陳晉興 共同主持人: 柯文哲 報告類型: 精簡報告 處理方式: 本計畫可公開查詢中 華 民 國 94 年 10 月 17 日
行政院國家科學委員會專題研究計畫 成果報告
利用合成改良之癌症專一性啟動子進行肺癌之基因治療
計畫類別: 個別型計畫 計畫編號: NSC 93-2314-B-002-237 執行期間: 93 年 08 月 01 日 至 94 年 07 月 31 日 執行單位: 國立臺灣大學醫學院外科 計畫主持人: 陳晉興 共同主持人: 柯文哲 報告類型: 精簡報告 處理方式: 本計畫可公開查詢Abstract:
Key words: lung cancer, gene therapy, survivin, GRP, BIK
Non-small-cell lung cancer (NSCLC) is one of the leading causes of cancer death in Taiwan and numerous developed countries. No obvious improvement in survival can be achieved in recent years although various efforts have been made. As the pathogenesis and biology of tumors are becoming clear, gene therapy could be an attractive modality for the treatment of lung cancer. Most gene therapy approaches currently use nonspecific, nonselective prokaryotic promoters such as the CMV promoter that can be expressed at high levels in normal cells, potentially contributing to toxicity. An alternative approach would be to use a tissue- or tumor-specific promoter to limit the spectrum of cells that express the gene therapy construct. In the previous studies, we have shown in vitro and in vivo that the activation of the survivin promoter is lung cancer specific. In this study, we choose another promoter, the GRP-78 (glucose-regulated protein 78) promoter to evaluate its future application in lung cancer gene therapy. Our preliminary data showed that the GRP-78 promoter was cancer-specific in cell line studies. To enhance its activity, the CMV and GRP enhancers were added at the upstream of the GRP-78 promoter. It appeared that both the CMV enhancer and GRP enhancer could increase the activity of the GRP promoter, respectively, while the cancer-specificity remained unchanged. By using this modified GRP-78 promoter, we showed that the activity and cancer specificity of this promoter is better than the CMV promoter in both cell lines and xenograft animal studies. The promising results of this study indicate the potential use of the modified GRP-78 promoter in cancer gene therapy.
Introduction:
Lung cancer is one of the leading causes of death worldwide1, 2. Survival has not substantially increased recently, although vigorous efforts have been made to improve it. As the pathogenesis and biology of tumors are becoming clear, gene therapy could be an effective alternative for the treatment of lung cancer.
Most gene therapy approaches currently use nonspecific, nonselective prokaryotic promoters such as the simian virus 40 (SV40) and cytomegalovirus (CMV) promoters
that can be expressed at high levels in normal cells, potentially contributing to toxicity. This is particularly true when cytotoxic or strongly apoptotic genes are used. For cancer-specific expression of cytotoxic or apoptotic genes, an better approach would be to use a tissue- or cancer-specific promoter to limit the spectrum of cells that express the gene therapy construct3, 4.
Examples possible for such therapy are the survivin and GRP-78 (glucose-regulated protein-78) promoters. Survivin is a member of the IAP (inhibitor of apoptosis) family. Survivin is overexpressed in common cancers but not in normal adult tissues 5-7. Our recent research also showed that the Survivin promoter was more than 200 times more cancer specific than the cytomegalovirus (CMV) promoter. In addition, the Survivin promoter can induce transgene expression of a mutant BIK in cancer cells and so suppressed the growth of cancer cells in vitro and in vivo8. However, the activity of the survivin promoter was only 10% to 30% of the CMV promoter, which may hamper the potential use of the Survivin promoter in future gene therapy.
GPR-78 is a member of GRPs, which functions as a molecular chaperone. The GRP-78 is constitutively expressed in the endoplasmic reticulum (ER) in most cells under normal growth conditions, and can be highly induced by stressed cellular environments such as low glucose, low oxygen, or reagents that disrupt the ER function9. The expression of GRP-78 was mainly regulated by transcription, and it was also reported that the mRNA and protein levels of GRP-78 was highly elevated in breast cancers than in normal breast tissues10.
MATERIAL AND METHODS: Cell lines:
purchased from the American Type Culture Collection and cultured according to the vendor’s instruction.
2. Human normal cell lines of the lung (WI-38) and breast (184A1) were purchased from the American Type Culture Collection and cultured according to the vendor’s instruction for comparison.
3. Other cancer cell lines of different organs such as colon (HT-29), cervix (Hela), pancrease (Capan-1) and breast (MDA-435) were used for comparison.
Construction of vectors:
Various different luciferase constructs were generated by using the pGL3-basic-luciferase vector (Promega, commercially available).
To determine whether the modified GRP-78 promoter activity is higher in NSCLC cell lines than in normal cell lines, dual luciferase reporter assay by transient transfection of DNA-liposome was used:
GRP promoter LUC pGRP-Luc
GRP promoter LUC pGG-Luc
GRP enhancer
GRP promoter LUC pGMG-Luc GRP enhancer CMV enhancer
BglII 531 bp HindIII
KpnI NheI 310 bp
CMV promoter LUC pCMV-Luc
GRP promoter LUC
CMV enhancer NheI 453 bp XhoI
pMG-Luc
GRP promoter pGMG-BIKDD GRP enhancer CMV enhancer BikDD
Lipids (Albaster, AL). Cells were cultured for 24h in sex-well plates with RPMI or DMEM/F12 medium with 10% FBS until 60-70% confluence is reached. The DNA-liposome was added into the cultured plates at a ratio of 1.5 ug of DNA/well. Cotransfection with 0.15 ug of pRL-TK was performed as a control for transfection efficiency. After 24-36 hours, cells were washed with PBS and lysed by 250ul of luciferase Cell Culture Lysis Reagent (Promega). Lysed cells were scraped into Eppendorf tubes and then centrifuged at 12,000 rpm to remove cell debris. Twenty microliters of the supernatant and 50 ul of Luciferase Assay Reagnet (Promega) were added at the same time. The luciferase activity of each reporter construct was measured by luminometer. Luciferase activity was normalized to the transfection efficiency determined by the pRL-TK luciferase assay.
To determine whether BIKDD driven by the modified GRP-78 promoters has preferential killing effect of cancer cells than the CMV promoter in vitro:
The cells were cotransfected with 3 ug/well of tested or controlled BIK plasmids and 0.3 ug/well of CMV-GFP. Forty hours after transfection, the cells expressing GFP were selected by fluorescence-activated cell sorter (FACS) assay, and the apoptotic portion of these GFP-expressed cells was assessed by flow cytometric detection of sub-G1 DNA content. The apoptotic versus nonapoptotic cells were counted in each cell line.
To determine whether the modified GRP-78 promoter activity is higher in tumor tissue than in normal tissues:
Athymic nude mice (nu/nu, 6-8 weeks of age, 18-22 g) were inoculated with the human NSCLC cell line H1299 subcutaneously at a dose of 5x106 cells/mouse. When the tumor reaches a size of 4-5 mm2, the animals were randomized into groups with 5 mice in each group. The mice received a single i.v. injection of
liposome-promoter-luciferase, 50 ug of DNA/mice. Forty-eight hours after injection, the tumors and other organs were harvested, and the protein extracted and analyzed for luciferase activity.
To determine whether BIKDD driven by the modified GRP-78 promoter has a better killing effect of tumor than the CMV promoter in vivo:
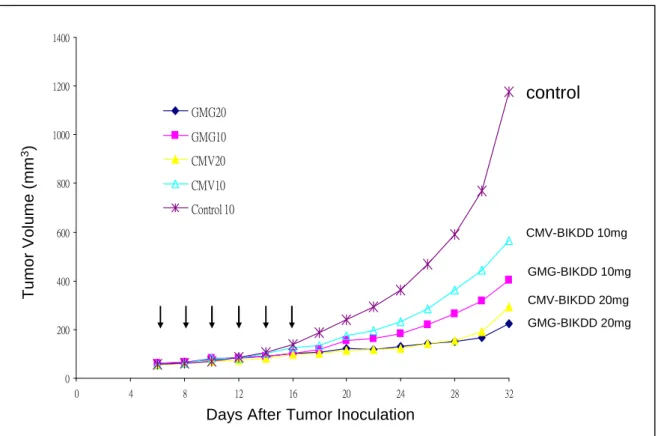
Athymic nude mice (nu/nu, 6-8 weeks of age, 18-22 g) were inoculated with the human NSCLC cell line H1299 subcutaneously at a dose of 5x106 cells/mouse. When the tumor reaches a size of 4-5 mm2, the animals were randomized into groups with 5 mice in each group. The mice in the treatment group were repeated i.v. injections of the modified BIKDD constructs, 50 ug of DNA/mice. The mice in the control group received the same dose of liposome-CMV-BIKDD. The tumor size was recorded every other day. The accumulated tumor volume and survival of the mice between the groups were compared.
Results:
The GRP-78 promoter activity was highly activated in cancer cell lines than in normal cell lines.
0 2000 4000 6000 HT-29 Hela H143 5 H322 H129 9 WI-3 8 184A 1 HBE4 /E6E 7 E6E7 Cancer cells
Normal or immortalized cells
D u a l L u c if e ra s e R a ti o
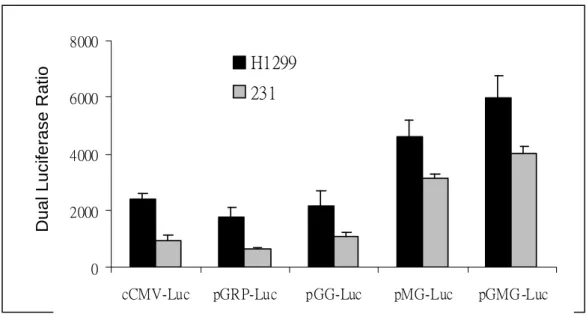
To increase the transgene expression of the GRP-78 promoter, the CMV and GRP enhancers were used to modify the survivin and GRP-78 promoter. We have constructed various plasmids by combination of these DNA fragments and founded that the CMV and GRP enhancers can substantially increase the activity of the GRP promoter
The killing effects of pGMG-BIKDD is significantly better than the pCMV-BIKDD
Fig. 3. The pGMG-BIKDD kills more cancer cells than the pCMV-BIKDD in H1299 and 22B cell lines. D u a l L u c if e ra s e R a ti o 0 2000 4000 6000 8000
cCMV-Luc pGRP-Luc pGG-Luc pMG-Luc pGMG -Luc H1299
231
Fig. 2. The activity of the GRP-78 promoter was significantly increased by adding the GRP and CMV enhancers.
0 10 20 30 40 50 60 70 80 90 100 H1299 22B T r e atm e n ts % o f v ia b il it y CM V- BIK D D GM G- BIK D D
To test the anti-cancer activity in vivo, the xenograft animal model was used. We found that the anti-cancer effects of pGMG-BIKDD was significantly better than the pCMV-BIKDD.
Discussion:
Discussion:
In this study, we found that the GRP-78 promoter is also a cancer-specific promoter. Moreover, we found that the GRP enhancer and the CMV enhancer can increase the activity of the GRP-78 promoter while the cancer-specificity remains unchanged. After modification, the modified GRP-78 promoter carries a better activity and cancer specificity than the CMV promoter. By comparison, we have shown that the GMG-BIKDD inhibit tumor growth more potently than the CMV promoter. In the near future, the modified GRP-78 construct may be used in lung cancer patients to evaluate it’s potential used in cancer gene therapy.
control CMV-BIKDD 10mg GMG-BIKDD 10mg CMV-BIKDD 20mg GMG-BIKDD 20mg 0 200 400 600 800 1000 1200 1400 0 4 8 12 16 20 24 28 32 GMG20 GMG10 CMV20 CMV10 Control 10 T u m o r V o lu m e (m m 3)
Days After Tumor Inoculation
REFERENCES:
1.Leading causes of death from cancer, Taiwan area, 1997. Health and vital statistics. Taiwan area, R.O.C. 1997 p90.
2.Boring CC, Squire TS, Tong T: Cancer statistics 1993. Ca Cancer J Clin 43:7-26, 1993.
3.Gibbs JB. Mechanism based target identification and drug discovery in cancer research. Science 2000;287:1969-73.
4.Robertson MW III, Wang M, Siegal GP, et al. Use of a tissue-specific promoter for targeted expression of te herpes simplex virus thymidine kinase gene in cervical carcinomal cells. Cancer Gene Ther. 1998;5:331-6.
5.Alteiri DC. Validating Survivin as a cancer therapeutic target. Nature Rev. Cancer 2003;3:46-54.
6.Ambrosini G, Adida C, Altieri DC. A novel anti-apoptosis gene, Survivin, expressed in cancer and lymphoma. Nature Med. 1997;3:917-21.
7.Tamm I, Wang Y, Sausville E, et al: IAP-family protein Survivin inhibits caspase activity and apoptosis induced by Fas(CD95), Bax, caspases, and anticancer drugs. Cancer Res. 58;5315-5320, 1998.
8.Chen JS, Liu JC, Shen L, et al. Cancer-specific expression of the survivin romoter and its potential use in gene therapy. Manuscript in preparation.
9.Lee AS. The glucose-regulated proteins: stress induction and clinical applications. Trends Biochem Sci. 26:504-10, 2001.
10.Fernandez PM, et al. Overexpression of the glucose-regulated stress gene GRP78 n malignant but not benign human breast lesions. Breast Cancer Res. Treat. 59, 15-26. 1996.