The
NBS1 Genetic Polymorphisms and the Risk
of the Systemic Lupus Erythematosus in Taiwanese Patients
Ying-Ju Lin
&Yu-Ching Lan
&Lei Wan
&Chung-Ming Huang
&Cheng-Wen Lin
&Kai-Chung Hsueh
&Da-Yuan Chen
&Ting-Hsu Lin
&Fuu-Jen Tsai
Received: 29 December 2009 / Accepted: 17 May 2010 / Published online: 23 June 2010 # Springer Science+Business Media, LLC 2010
Abstract
Introduction Systemic lupus erythematosus (SLE), a
multi-systemic autoimmune disease, is characterized by the
production of a range of autoantibodies against nuclear
constituents and other self-antigens. The studies in DNA
repair deficiencies in SLE patients have been recently
investigated.
Aims Few studies have been conducted on DNA repair
gene polymorphisms and their role in autoimmune diseases.
Our study purpose was to examine and compare NBS1
genotype distributions in a group of Taiwanese SLE
patients and controls in Taiwan.
Patients and Methods Participants were Taiwanese SLE
patients and healthy controls. We studied associations
among NBS1 polymorphisms
—rs1061302, rs709816, and
rs1805794—considering clinical features for the entire
group and stratified subgroups. No statistically significant
differences between the patients and controls were noted.
However, we observed significant decreases in Ht1-GGG,
Ht2-AAC, and Ht3-AGC in the SLE patients (Ht1-GGG,
OR=0.26, 95% CI: 0.16–0.41; Ht2-AAC, OR=0.30, 95%
CI: 0.17–0.53; Ht3-AGC, OR=0.35, 95% CI: 0.19–0.71)
and significant increases in Ht4-AAG, Ht5-AGG, and
Ht8-GGC among the SLE patients. Combined, these results
suggest an association between NBS1 genetic
polymor-phisms and Taiwanese SLE patients.
Keywords Systemic lupus erythematosus . NBS1 .
genetic polymorphism
Introduction
Systemic lupus erythematosus (SLE), a multisystemic
autoimmune disease, is characterized by the production of
a range of autoantibodies against nuclear constituents and
other self-antigens [
1
–
3
]. Since SLE is a multifactorial
disease, complex interactions among genetic, hormonal,
and environmental factors play important roles in
deter-mining its induction and development [
4
,
5
]. The immune
complex damages multiple organ systems and presents
numerous clinical manifestations [
6
–
9
]. Arthritis, serum
Ying-Ju Lin and Yu-Ching Lan made equal contributions to this work. Y.-J. Lin
:
L. Wan:
D.-Y. Chen:
T.-H. Lin:
F.-J. Tsai (*)Department of Medical Research, China Medical University Hospital, No.2 Yuh-Der Road,
Taichung 404, Taiwan
e-mail: [email protected] Y.-J. Lin
:
L. Wan:
F.-J. TsaiSchool of Chinese Medicine, China Medical University, Taichung, Taiwan
Y.-C. Lan
Department of Health Risk Management, China Medical University,
Taichung, Taiwan C.-M. Huang
Division of Immunology and Rheumatology, China Medical University Hospital, Taichung, Taiwan
C.-W. Lin
Department of Medical Laboratory Science and Biotechnology, China Medical University,
Taichung, Taiwan K.-C. Hsueh
Children’s Medical Center, China Medical University Hospital, Taichung, Taiwan
autoantibodies, glomerulonephritis, joint pain, skin rash,
and vasculitis commonly develop in combination with one
or more symptoms observed in SLE patients [
10
].
An important clinical factor is autoimmunity to
double-stranded DNA and nucleosomes that present diagnostic and
pathogenic impacts. Although DNA is poorly immunogenic,
antigenicity may be induced by reactive oxygen species,
drugs, and exposure to UV light [
11
–
14
]. These
environmen-tal factors may trigger altered DNA conformation or damage
to/breaks in DNA bases, resulting in apoptotic bodies. Once
produced, damaged DNA is normally recognized and
repaired by complex mechanisms [
15
,
16
]. DNA repair
system deficiencies have been investigated in SLE patients
[
17
–
23
], and among other findings researchers have reported
that (a) their peripheral blood neutrophils display increased
DNA damage [
17
], (b) DNA damage repair is delayed by
ionizing radiation or oxidative damage in cells [
18
,
19
], and
(c) DNA repair pathway genes are downregulated in
peripheral blood lymphocytes [
20
,
21
].
The NBS1 gene contains 16 exons and is located on
chromosome band 8q21.3 [
24
,
25
]. One NBS1 gene product,
nibrin (p95, NBS1), is a key regulator of the MRE11/
RAD50/nibrin (M/R/N) protein complex that is involved in
DNA double-strand break repair, telomere maintenance,
immunoglobulin class switching, meiotic recombination,
and DNA damage response [
26
,
27
]. Nibrin directs the
M/R/N complex to DNA damage sites and promotes DNA
binding and nuclease activity. In addition, nibrin is required
for ataxia telangiectasia mutated (ATM) kinase activation
and the related downstream ATM phosphorylation of targets
that are critical to the cell cycle [
28
,
29
].
To date, few studies have been conducted on DNA repair
gene polymorphisms and their role in autoimmune diseases
[
30
–
33
]. Our study purpose was to examine and compare
NBS1 genotype distributions in a group of Taiwanese SLE
patients and controls in Taiwan. It is interesting to carry out
an association study for a better understanding of the role of
NBS1 in SLE.
Patients and Methods
Patients The study sample consisted of 164 SLE patients
and 176 healthy individuals recruited from China Medical
University Hospital in Taichung, Taiwan. All of the patients
met American Rheumatism Association criteria for SLE
classification [
10
]. Data collection was partly determined
by patient availability, diagnoses of inactive or active
disease, and whether or not they were undergoing treatment
with steroids. All samples for genomic DNA isolation were
collected by venipuncture. Members of the control group
were identified through routine health examinations. All
participants signed informed consent forms. The study
TableI PCR Primer and Probe Sequences of NBS1 Genetic Polymorphisms that W ere Used in this Study Gene Name Polymorphisms Exon/ Intron Location in the Protein SNP Database ID Nucleotide Change Reporter 1Dye Allele Reporter 1 Quencher Reporter 2 Dye Allele Reporter 2 Quencher Context Sequence NBS1 Glu185Gln 5 BRCT domain rs1805794 C/G VIC C NFQ F A M G NFQ AA TTTGTGGAGGCTGCTTCTTGGACT [C/G] AACTGCTTTCAGGAA TTCAGT AAAA Asp399Asp 10 central region rs709816 A/G VIC A NFQ F A M G NFQ TGCAGGACTCCTTT ACAGTGGGTGC [A/G] TCTTGTGAAAGCA TTCTGAA TTTTT Pro672Pro 13 MRE1 1 binding domain rs1061302 A/G VIC A NFQ F A M G NFQ A T T AAAAACTCT ACTTCCAGAAA TCC [A/G] TCTGGCA T AAA TGA TGA TT A TGGTC The genotyping method was designed by T aqMan® Genotyping Assays (Applied Biosystems) BRCT breast cancer C-terminal
design was approved by the Human Subjects Committee of
the Institutional Review Board of China Medical University
Hospital.
Genomic DNA Extraction and Genotyping Genomic DNA
was extracted from peripheral blood leukocytes according
to standard protocols (Genomic DNA Kit, Qiagen).
Biallelic NBS1 genetic polymorphisms were detected using
TaqMan(R) Genotyping Assays (Applied Biosystems;
Table
I
).
Statistical Analysis Genotype and allelic frequency
dis-tributions for the targeted polymorphisms in both SLE
patients and controls were analyzed using a chi-squared
test. Allelic frequencies are expressed as percentages of
total alleles. Odds ratios (OR) were calculated for
genotype and allelic frequencies (95% confidence
inter-val [CI]). Haplotypes were inferred from unphased
genotype data using the Bayesian statistical method in
the Phase 2.1 software program [
34
,
35
]. All statistical
tests were performed using SPSS 12.0 for Windows XP
(SPSS, Inc., Chicago, IL, USA). The p values were
adjusted by using Bonferroni’s correction. Statistical
significance was considered as p value <0.016 for allele
and genotype frequency analysis data. Statistical
signifi-cance was considered as p value <0.00625 for haplotype
analysis data.
Results
The genotypes of NBS1 (rs1061302), NBS1 Asp399Asp
(rs709816), and NBS1 Glu185Gln (rs1805794) were
identified by probe hybridization with corresponding
primers (Table
I
). Allele and genotype frequencies are
presented in Table
II
. As shown, genotype distributions
were consistent with the Hardy–Weinberg equilibrium.
Statistically significant differences in allele and genotype
frequencies were not observed between the patients and
controls.
Haplotype frequencies were estimated using the three
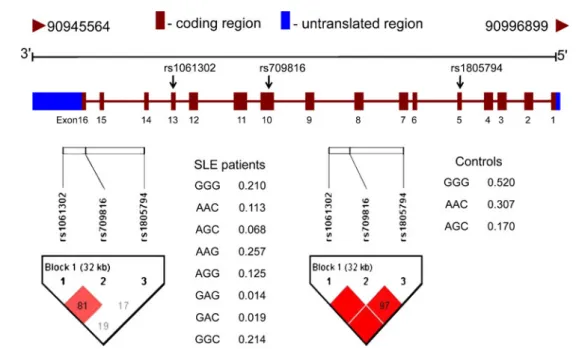
genetic polymorphisms (allele frequency >5%; Fig.
1
and
Table
III
). Of the eight observed haplotypes, six were
present in both SLE patients and control individuals. The
frequency of the most common haplotype (Ht1-GGG) in
the control group was 51.98%, compared to 21.75% for
the SLE patients (Table
III
). According to results from a
haplotype-specific analysis, Ht1-GGG, Ht2-AAC, and
Ht3-AGC were identified as
“protective” (Ht1-GGG, p<
0.001, OR = 0.26, 95% CI: 0.16–0.41; Ht2-AAC, p<0.001,
OR = 0.30, 95% CI: 0.17–0.53; Ht3-AGC, p<0.001, OR=
0.35, 95% CI: 0.19
–0.71). Ht4-AAG, Ht5-AGG, and
Ht8-GGC were presented as
“at-risk” (Ht4-AAG, p<0.001,
OR = 104.06, 95% CI: 6.42–1,685.70; Ht5-AGG, p<0.001,
OR = 2524.06, 95% CI: 7.96 × 10
−6–8×10
11; Ht8-GGC, p <
0.001). Results from our analysis of associations between
SLE patient clinical feature profiles and various
haplo-types suggested that there were no significant associations
(not shown).
Discussion
In this study, our findings suggest that (a) Taiwanese
individuals with the NBS1 haplotypes Ht1-GGG, Ht2-AAC,
and Ht3-AGC are at lower risk of developing SLE and (b)
Polymorphisms SLE Controls p value Odds ratio (95% CI)
Number (%) Number (%) NBS1 Pro672Pro (rs709816) A 149 (54.4) 169 (48.0) 0.114 1.29 (0.94–177) G 125 (45.6) 183 (52.0) 1 AA 41 (29.9) 35 (19.9) 0.121 1.70 (0.88–3.26) AG 67 (48.9) 99 (56.3) 0.98 (0.56–1.73) GG 29 (21.2) 42 (23.9) 1 NBS1 Asp399Asp (rs709816) A 105 (38.3) 109 (31.0) 0.054 1.39 (0.99–1.93) G 169 (61.7) 243 (69.0) 1 AA 16 (11.7) 18 (10.2) 0.060 1.57 (0.74–3.37) AG 73 (53.3) 73 (41.5) 1.77 (1.10–2.86) GG 48 (35.0) 85 (48.3) 1 NBS1 Glul85Gln (rs1805794) C 135 (41.2) 163 (47.9) 0.078 0.76 (0.56–1.03) G 193 (58.8) 177 (52.1) 1 CC 24 (14.6) 34 (20.0) 0.174 0.55 (0.28–1.06) CG 87 (53.0) 95 (55.9) 0.71 (0.43–1.17) GG 53 (32.3) 41 (24.1) 1
Table II Allele and Genotype Frequencies of NBS1 Gene Polymorphisms in Taiwanese SLE Patients and Controls
Allele frequency was compared between SLE and controls by
2×2χ2tests. Genotype frequency
was compared between SLE and
controls by 3×2χ2 tests. The
p values were adjusted by using
Bonferroni’s correction. Statistical
significance was considered as p value <0.016 (0.053) CI confidence interval
individuals with the NBS1 haplotypes Ht4-AAG,
Ht5-AGG, and Ht8-GGC are at higher risk of developing
SLE. Combined, these results suggest an association
between NBS1 genetic polymorphisms and Taiwanese
SLE patients.
To date, few studies have been conducted on DNA repair
gene polymorphisms and their role in autoimmune diseases
[
30
–
33
]. In the present study, we found an association
between SLE and NBS1 haplotypes. Interpreting these
results is limited by the lack of research on links between
NBS1 genetic variants, autoimmune diseases, and related
clinical manifestations. However, a possible explanation for
our results is that insufficient DNA damage repair may
contribute to SLE immune dysfunction. More detailed
studies are required to determine which molecular
mecha-nisms are controlled by NBS1 genetic variants.
The genetic variants from DNA repair genes have been
investigated in cancers [
30
,
32
,
36
–
38
]. These genetic
polymorphisms commonly found in DNA repair genes have
been suggested for altering protein function and deficits in
repair capacity, resulting in genetic instability and failure to
maintain normal cell proliferation. SNPs in NBS1 are
Table III Haplotype Frequencies of NBS1 Gene Between SLE Patients and Controls
Haplotype rs1061302 rs709816 rs1805794 SLE (%) Control (%) p value Odds ratio (95% CI)
Ht1 G G G 21.75% 51.98% <0.001 0.26 (0.16–0.41) Ht2 A A C 11.79% 30.67% <0.001 0.30 (0.17–0.53) Ht3 A G C 6.63% 17.05% <0.001 0.35 (0.19–0.71) Ht4 A A G 23.04% 0.29% <0.001 104.06 (6.42–1,685.70) Ht5 A G G 12.58% 0.01% <0.001 2,524.06 (7.96×10−6–8×1011) Ht6 G A G 1.50% 0.01% 0.106 178.71 (1.87×10−5–1.71×109) Ht7 G A C 1.78% 0.00% 0.075 — Ht8 G G C 20.93% 0.00% <0.001 —
—indicates not applicable, CI confidence interval
Order of SNPs comprising the NBS1 haplotypes: rs1061302, rs709816, and rs1805794. Percentages may not add to 100% because of the presence of rare haplotypes not presented in this table. The p values were adjusted by using Bonferroni’s correction. Statistical significance was considered as p value <0.00625 (0.05/8)
Fig. 1 Results of SNP association study of the NBS1 genetic polymorphisms on chromosome 8q21.3. Upper schematic showing the NBS1 gene which contains 16 exons (from chromosome 8: 90,945,564 to 90,996,899) were used for genotyping in 164 Taiwanese SLE patients and the 176 individuals from the general population of Taiwan with Han Chinese ethnic background for SNP
association. Lower schematic showing haplotype blocks for control and SLE patients. The blocks were constructed based on the confidence interval approach using the HAPLOVIEW software [50]. The red denotes the haplotype blocks and the white represents evidence of recombination
associated with certain types of cancer. For example,
Glu185Gln (rs1805794) is associated with increased risks
for lung cancer, breast cancer, and leukemia [
39
–
44
]. The
functional relevance of this polymorphism is unknown.
However, its location within the breast cancer C-terminal
domain may be related to some effect on protein function
[
27
]. Lu and colleagues suggested that the homozygous CC
genotype and heterozygous GC genotype may contribute to
sporadic breast cancer in young non-Hispanic white women
[
43
]. The heterozygous GC genotype carriers had an
increased risk of lung cancer in China [
44
]. In addition,
no significant difference of this polymorphism but some
specific haplotypes of the NBS1 gene was observed in
leukemia [
42
]. Few studies have been conducted so far to
study the association of NBS1 haplotypes with SLE. Our
analysis on the NBS1 haplotype provided additional
information on these polymorphisms as markers of genetic
susceptibility and a more efficient method for assessing the
genetic susceptibility of a candidate gene than any of the
polymorphisms. Individuals with SLE are known to have
increased susceptibility to lung cancer, breast cancer, and
hematological malignancies [
45
–
47
]. Our findings suggest
that SLE and certain types of cancer share similar
pathogenic pathways during disease development (e.g.,
DNA repair system deficiencies caused by genetic variants
of functional SNPs in DNA repair genes). Furthermore,
DNA repair mechanisms are central to genetic
recombina-tion during lymphocyte maturarecombina-tion [
48
,
49
]. Further studies
are required to determine the role of DNA repair genes in
the relationship between SLE and cancer. It is our hope that
these findings will assist in that effort.
In summary, to our knowledge, this is the first evidence
of an association between common NBS1 genetic variants
and SLE. According to our results, haplotypes containing
variant alleles from the three studied NBS1 polymorphisms
may cause increased susceptibility to SLE.
Acknowledgments This project was supported by grants from China Medical University (CMU97-CMC-005), China Medical University Hospital (DMR-97-102), and the Republic of China National Science Council (NSC97-2320-B-039-023-MY3).
References
1. Mortensen ES, Fenton KA, Rekvig OP. Lupus nephritis: the central role of nucleosomes revealed. Am J Pathol. 2008;172 (2):275–83.
2. Fenton KA, Rekvig OP. A central role of nucleosomes in lupus nephritis. Ann N Y Acad Sci. 2007;1108:104–13.
3. Yasutomo K. Pathological lymphocyte activation by defective clearance of self-ligands in systemic lupus erythematosus. Rheumatology (Oxford). 2003;42(2):214–22.
4. Tsao BP. The genetics of human systemic lupus erythematosus. Trends Immunol. 2003;24(11):595–602.
5. Rosenstein BS, Rosenstein RB, Zamansky GB. Repair of DNA damage induced in systemic lupus erythematosus skin fibroblasts by simulated sunlight. J Invest Dermatol. 1992;98(4):469–74. 6. Kotzin BL. Systemic lupus erythematosus. Cell. 1996;85(3):303–
6.
7. Nath SK, Kilpatrick J, Harley JB. Genetics of human systemic lupus erythematosus: the emerging picture. Curr Opin Immunol. 2004;16(6):794–800.
8. Robak E, Sysa-Jedrzejowska A, Robak T, Smolewski P. Peripheral blood lymphocyte apoptosis and circulating dendritic cells in patients with systemic lupus erythematosus: correlation with immunological status and disease-related symptoms. Clin Rheumatol. 2006;25 (2):225–33.
9. Kyttaris VC, Katsiari CG, Juang YT, Tsokos GC. New insights into the pathogenesis of systemic lupus erythematosus. Curr Rheumatol Rep. 2005;7(6):469–75.
10. Hochberg MC. Updating the American College of Rheumatology revised criteria for the classification of systemic lupus erythema-tosus. Arthritis Rheum. 1997;40(9):1725.
11. Cooke MS, Mistry N, Wood C, Herbert KE, Lunec J. Immunoge-nicity of DNA damaged by reactive oxygen species—implications for anti-DNA antibodies in lupus. Free Radic Biol Med. 1997;22(1– 2):151–9.
12. Zacharias W, Koopman WJ. Lupus-inducing drugs alter the structure of supercoiled circular DNA domains. Arthritis Rheum. 1990;33(3):366–74.
13. Cerboni B, Morozzi G, Galeazzi M, Bellisai F, Micheli V, Pompucci G, et al. Poly(ADP-ribose) polymerase activity in systemic lupus erythematosus and systemic sclerosis. Hum Immunol 2009;70(7):487–91.
14. Alam K, Moinuddin, Jabeen S. Immunogenicity of mitochondrial DNA modified by hydroxyl radical. Cell Immunol. 2007;247 (1):12–7.
15. Zhou BB, Elledge SJ. The DNA damage response: putting checkpoints in perspective. Nature. 2000;408(6811):433–9. 16. Friedberg EC. DNA damage and repair. Nature. 2003;421
(6921):436–40.
17. Courtney PA, Crockard AD, Williamson K, Irvine AE, Kennedy RJ, Bell AL. Increased apoptotic peripheral blood neutrophils in systemic lupus erythematosus: relations with disease activity, antibodies to double stranded DNA, and neutropenia. Ann Rheum Dis. 1999;58(5):309–14.
18. McCurdy D, Tai LQ, Frias S, Wang Z. Delayed repair of DNA damage by ionizing radiation in cells from patients with juvenile systemic lupus erythematosus and rheumatoid arthritis. Radiat Res. 1997;147(1):48–54.
19. McConnell JR, Crockard AD, Cairns AP, Bell AL. Neutrophils from systemic lupus erythematosus patients demonstrate in-creased nuclear DNA damage. Clin Exp Rheumatol. 2002;20 (5):653–60.
20. Mandel M, Gurevich M, Pauzner R, Kaminski N, Achiron A. Autoimmunity gene expression portrait: specific signature that intersects or differentiates between multiple sclerosis and systemic lupus erythematosus. Clin Exp Immunol. 2004;138(1):164–70. 21. Pereira E, Tamia-Ferreira MC, Cardoso RS, Mello SS,
Sakamoto-Hojo ET, Passos GA, et al. Immunosuppressive therapy modulates T lymphocyte gene expression in patients with systemic lupus erythematosus. Immunology. 2004;113(1):99–105.
22. Lawley PD, Topper R, Denman AM, Hylton W, Hill ID, Harris G. Increased sensitivity of lymphocytes from patients with systemic autoimmune diseases to DNA alkylation by the methylating carcinogen N-methyl-N-nitrosourea. Ann Rheum Dis. 1988;47 (6):445–51.
23. Palmer RG, Smith-Burchnell CA, Dore CJ, Denman AM. Sensitivity of lymphocytes from patients with systemic lupus erythematosus to the induction of sister chromatid exchanges by
alkylating agents and bromodeoxyuridine. Ann Rheum Dis. 1987;46(2):110–3.
24. Carney JP, Maser RS, Olivares H, Davis EM, Le Beau M, Yates 3rd JR, et al. The hMre11/hRad50 protein complex and Nijmegen breakage syndrome: linkage of double-strand break repair to the cellular DNA damage response. Cell. 1998;93 (3):477–86.
25. Matsuura S, Tauchi H, Nakamura A, Kondo N, Sakamoto S, Endo S, et al. Positional cloning of the gene for Nijmegen breakage syndrome. Nat Genet. 1998;19(2):179–81.
26. Matsuura S, Kobayashi J, Tauchi H, Komatsu K. Nijmegen breakage syndrome and DNA double strand break repair by NBS1 complex. Adv Biophys. 2004;38:65–80.
27. Kobayashi J, Antoccia A, Tauchi H, Matsuura S, Komatsu K. NBS1 and its functional role in the DNA damage response. DNA Repair (Amst). 2004;3(8–9):855–61.
28. Uziel T, Lerenthal Y, Moyal L, Andegeko Y, Mittelman L, Shiloh Y. Requirement of the MRN complex for ATM activation by DNA damage. EMBO J. 2003;22(20):5612–21.
29. Lee JH, Paull TT. Direct activation of the ATM protein kinase by the Mre11/Rad50/Nbs1 complex. Science. 2004;304 (5667):93–6.
30. Bassi C, Xavier D, Palomino G, Nicolucci P, Soares C, Sakamoto-Hojo E, et al. Efficiency of the DNA repair and polymorphisms of the XRCC1, XRCC3 and XRCC4 DNA repair genes in systemic lupus erythematosus. Lupus. 2008;17(11):988–95.
31. Hur JW, Sung YK, Shin HD, Park BL, Cheong HS, Bae SC. TREX1 polymorphisms associated with autoantibodies in patients with systemic lupus erythematosus. Rheumatol Int. 2008;28 (8):783–9.
32. Koyama A, Kubota Y, Shimamura T, Horiuchi S. Possible association of the X-ray cross complementing gene 1 (XRCC1) Arg280His polymorphism as a risk for rheumatoid arthritis. Rheumatol Int. 2006;26(8):749–51.
33. Graessler J, Verlohren M, Graessler A, Zeissig A, Kuhlisch E, Kopprasch S, et al. Association of chondromodulin-II Val58Ile polymorphism with radiographic joint destruction in rheumatoid arthritis. J Rheumatol. 2005;32(9):1654–61.
34. Stephens M, Donnelly P. A comparison of bayesian methods for haplotype reconstruction from population genotype data. Am J Hum Genet. 2003;73(5):1162–9.
35. Stephens M, Smith NJ, Donnelly P. A new statistical method for haplotype reconstruction from population data. Am J Hum Genet. 2001;68(4):978–89.
36. Goode EL, Ulrich CM, Potter JD. Polymorphisms in DNA repair genes and associations with cancer risk. Cancer Epidemiol Biomark Prev. 2002;11(12):1513–30.
37. Jiang J, Zhang X, Yang H, Wang W. Polymorphisms of DNA repair genes: ADPRT, XRCC1, and XPD and cancer risk in genetic epidemiology. Methods Mol Biol. 2009;471:305–33. 38. Masutani M, Nakagama H, Sugimura T. Poly(ADP-ribosyl)ation
in relation to cancer and autoimmune disease. Cell Mol Life Sci. 2005;62(7–8):769–83.
39. Hsu HM, Wang HC, Chen ST, Hsu GC, Shen CY, Yu JC. Breast cancer risk is associated with the genes encoding the DNA double-strand break repair Mre11/Rad50/Nbs1 complex. Cancer Epidemiol Biomark Prev. 2007;16(10):2024–32.
40. Smith TR, Levine EA, Freimanis RI, Akman SA, Allen GO, Hoang KN, et al. Polygenic model of DNA repair genetic polymorphisms in human breast cancer risk. Carcinogenesis. 2008;29(11):2132–8.
41. Ryk C, Kumar R, Thirumaran RK, Hou SM. Polymorphisms in the DNA repair genes XRCC1, APEX1, XRCC3 and NBS1, and the risk for lung cancer in never- and ever-smokers. Lung Cancer. 2006;54(3):285–92.
42. Mosor M, Ziolkowska I, Januszkiewicz-Lewandowska D, Nowak J. Polymorphisms and haplotypes of the NBS1 gene in childhood acute leukaemia. Eur J Cancer. 2008;44(15):2226–32.
43. Lu J, Wei Q, Bondy ML, Li D, Brewster A, Shete S, et al. Polymorphisms and haplotypes of the NBS1 gene are associ-ated with risk of sporadic breast cancer in non-Hispanic white women < or =55 years. Carcinogenesis. 2006;27(11):2209–16. 44. Lan Q, Shen M, Berndt SI, Bonner MR, He X, Yeager M, et al. Smoky coal exposure, NBS1 polymorphisms, p53 protein accumulation, and lung cancer risk in Xuan Wei, China. Lung Cancer. 2005;49(3):317–23.
45. Gayed M, Bernatsky S, Ramsey-Goldman R, Clarke A, Gordon C. Lupus and cancer. Lupus. 2009;18(6):479–85.
46. Gadalla SM, Amr S, Langenberg P, Baumgarten M, Davidson WF, Schairer C, et al. Breast cancer risk in elderly women with systemic autoimmune rheumatic diseases: a population-based case-control study. Br J Cancer. 2009;100(5):817–21.
47. Bernatsky S, Joseph L, Boivin JF, Gordon C, Urowitz M, Gladman D, et al. The relationship between cancer and medication exposures in systemic lupus erythaematosus: a case-cohort study. Ann Rheum Dis. 2008;67(1):74–9.
48. Stavnezer J, Guikema JE, Schrader CE. Mechanism and regula-tion of class switch recombinaregula-tion. Annu Rev Immunol. 2008;26 (26):261–92.
49. Jolly CJ, Cook AJ, Manis JP. Fixing DNA breaks during class switch recombination. J Exp Med. 2008;205(3):509–13. 50. Barrett JC, Fry B, Maller J, Daly MJ. Haploview: analysis and
visualization of LD and haplotype maps. Bioinformatics. 2005;21 (2):263–5.