Levels of endogenous polyamines and NaCl-inhibited growth of rice
seedlings
Chuan Chi Lin & Ching Huei Kao*
Department of Agronomy, National Taiwan University, Taipei, Taiwan, Republic of China (*author for correspondence)
Received 28 September 1994; accepted 21 November 1994
Key words: NaCl, Oryza sativa, polyamine, seedling growth
Abstract
The role of endogenous polyamines in the control of NaCl-inhibited growth of rice seedlings was investigated. Putrescine, spermidine and spermine were all present in shoots and roots of rice seedlings. NaCl treatment did not affect spermine levels in shoots and roots. Spermidine levels in shoots and roots were increased with increasing concentrations of applied NaCl. NaCl at a concentration of 50 mJ4, which caused only slight growth inhibition, drastically lowered the level of putrescine in shoots and roots. Addition of precursors of putrescine biosynthesis (L-arginine and L-ornithine) resulted in an increase in putrescine levels in NaCl-treated shoots and roots, but did not allow recovery of the growth inhibition of rice seedlings induced by NaCl. Pretreatment of rice seeds with putrescine caused an increase in putrescine level in shoots, but could not alleviate the inhibition effect of NaCl on seedling growth. The current results suggest that endogenous polyamines may not play a significant role in the control of NaCl-inhibited growth of rice seedlings.
Abbreviations: PUT - putrescine; SPD - spermidine; SPM - spermine.
1. Introduction
Aliphatic polyamines (PUT, SPD, and SPM), recently recognized as a new class of plant growth sub- stances [4], are present in all plants examined to date. Polyamine levels and plant growth rates have been positively correlated in a wide variety of conditions; high levels of polyamines are associated with rapidly growing tissues [ 1,4].
Several reports have shown that exogenous appli- cations of PUT can overcome the harmful effects of NaCl stress [6,9, lo]. In addition, carrot cells treated for 2 years in culture with 5 m&f PUT showed greater plating efficiency on a saline medium than cultures raised in the absence of PUT [8]. NaCl is known to inhibit the growth of rice seedlings [5, 9, 10, 111. It has been shown that PUT can reduce NaCl-induced inhibition of early seedling growth of rice [lo] and that endogenous levels of polyamines decreased in rice seedlings under NaCl stress [ 111. However, polyamine
accumulation in rice seedlings in response to NaCl has also been reported [2]. Furthermore, Krishnamurthy and Bhagwat [7] reported that PUT was accumulated in salt-sensitive rice cultivars and was significantly reduced in concentration in salt-tolerant ones under saline condition. In view of the contradictory results, the present investigation was conducted to determine the role of endogenous polyamines in the control of NaCl-inhibited growth of rice seedlings.
2. Materials and methods
Rice (Oryza sativa L., cv. Taichung Native 1) seeds were sterilized with 2.5% sodium hypochlorite for 15 min and washed thoroughly with distilled water. The seeds were then germinated in Petri dishes (20 cm) containing distilled water at 37 “C under dark condi- tion. After incubation of 1 -day, uniformly germinated seeds were selected and transferred to Petri dishes (9.0
Shoot
$
5 12 ; 8 4 0 50 100 150 NaCI, mMFig. 1. Effect of NaCl on seedling growth of rice. Seedling growth was measured after 5 days of treatment. Vertical bars represent standard errors (n = 4). Only those standard errors larger than the symbols are shown.
cm) containing two sheets of Whatman No. 1 filter paper moistened with 10 mL of distilled water or test solutions. The germinated seeds were allowed to grow at 27 “C in darkness and 3 mL of distilled water or test solutions was added to each Petri dish on day 3 of the growth Each Petri dish contained 20 germinated seeds. Each treatment was replicated 4 times. Fresh weight of shoot and root was measured after 5 days in darkness.
For polyamine extraction, shoot and root samples were homogenized in a motar and pestle in 5% per- chloric acid. Polyamine levels were determined using high performance liquid chromatography after benzoy- lation as described previously [3]. Polyamine levels were expressed as nmol per g fresh weight.
For all measurements, each treatment was repeated four times and all experiments were repeated three times. Similar results were obtained each time. The data reported here are from a single experiment.
Results and discussion
The growth of shoots and roots was followed by mea- suring the fresh weight of shoot and root. Figure 1 shows that the growth of seedlings decreased with
PUT
c
SPD 350 T , , ,‘I.--J
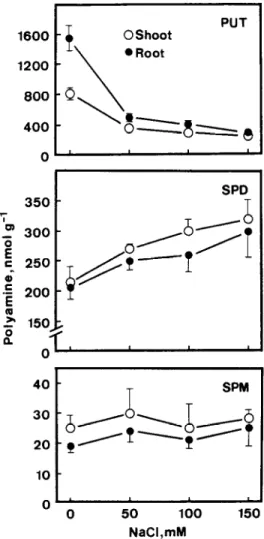
” 0 50 100 150 NaCl,mMFig. 2. Effect of NaCl on levels of polyamines in rice seedlings. Polyamines were determined after 5 days of treatment. Vertical bars represent standard errors (n = 4). Only those standard errors larger than the symbols are shown.
increase in concentration of NaCl. The growth of shoots and roots at 150 mM NaCl was reduced to 15 and 30%, respectively, of the control values. These results are, in general, consistent with those reported by Flowers and Yeo [5].
PUT, SPD, and SPM were present in both shoots and roots of rice seedlings. However, no detectable levels of cadaverine, a diamine usually found in the family Leguminosae, and diaminopropane, an oxida- tion product of naturally occurring polyamines, were observed in shoots and roots of rice seedlings. Figure 2 shows that NaCl treatment had no effect on the levels of SPM in shoots and roots of rice seedlings. The effect of NaCl on the levels of SPD in shoots and roots showed a distinct pattern. The levels of SPD in shoots
Shoot Root 1600 1200 800 400 0 -NaCI PUT +NaCI -NaCI PUT +NaCI 80 60 40 20 0 SPM 0 0124 0 0124 SPM L-Arginine,mM
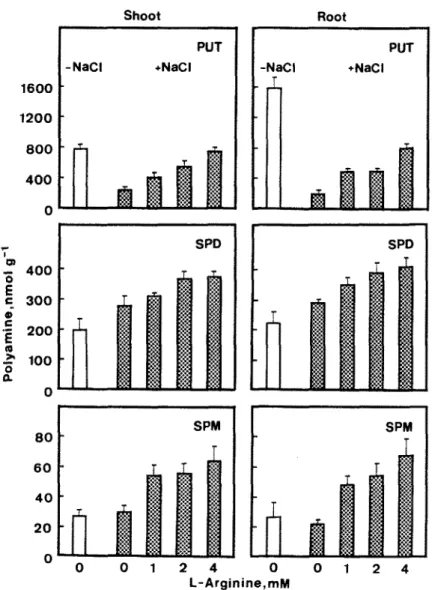
Fig. 3. Effect of L-arginine on polyamine levels in rice seedlings treated with NaC1. The concentration of NaCl was 150 mM. Polyamines were measured after 5 days of treatment. Vertical bars represent standard errors (II = 4).
and roots were found to increase with increasing NaCl concentrations (Fig. 2). All these results suggest that endogenous levels of SPD and SPM are not associated with growth inhibition of shoot and root in response to N&l.
NaCl treatment resulted in the decline of the levels of PUT in shoots and roots (Fig. 2). NaCl at the concen- tration of 50 m&f, which caused only slight growth inhi- bition, drastically lowered the levei of PUT in shoots and roots. As judged by the growth of rice seedlings in response to NaCl stress, the rice cultivar Taichung Native 1 used in this study is considered to be NaCl sensitive. However, our results are inconsistent with those reported by Krishnamurthy and Bhagwat [7] who
showed that PUT level accumulated in NaCl sensitive rice cultivars under saline condition.
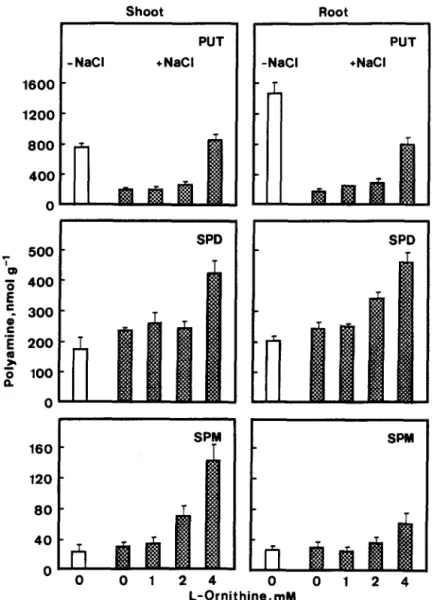
To characterize the role of endogenous polyamines in NaCl-inhibited growth of seedlings, precursors of the biosynthesis of PUT, such as L-arginine and L-omi- thine, were used to increase the level of intracellular PUT in NaCl-treated shoots and roots. L-Arginine treatment resulted in the increase of PUT, SPD and SPM levels in both shoots and roots of NaCl-treated seedlings (Fig. 3). The increase of PUT by L-arginine in NaCl-treated shoots was found to be in a concen- tration dependent manner (Fig. 3). L-Omithine at a concentration of 4 mM also increased PUT, SPD and SPM levels in NaCl-treated shoots and roots (Fig. 4).
Shoot Root SPD 7 500 $ 400 E 5 300 2 ‘E 200 s g 100 0 160 SPM 120 80 40 ‘0 0124 0 0124 L-Ornithine,mM
Fig. 4. Effect of L-omithine on polyamine levels in rice seedlings treated with NaCl. The concentration of NaCl was 150 mkf. Polyamines were measured after 5 days of treatment. Vertical bars represent standard errors (n = 4).
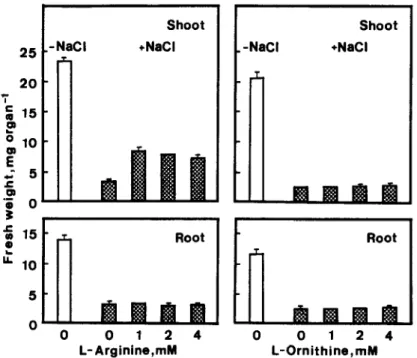
However, L-arginine and L-omithine did not reduce growth inhibition of roots under NaCl condition (Fig. 5) and L-omithine did not counteract the growth inhi- bition of shoots induced by NaCl (Fig. 5). L-Arginine, which increased PUT level in NaCl-treated shoots in a concentration dependent manner, alleviated growth inhibition of shoots induced by NaCl, but not in a con- centration dependent manner (Fig. 5).
To further characterize the role of PUT on the growth of shoot and root of rice seedlings in response to NaCl, we raised rice seedlings in the presence of PUT and tested its effect on growth inhibitionby NaCl. Rice seeds were soaked either in distilled water or PUT (1
mM) for 24 h in darkness and germinated seeds were transferred to distilled water or NaCl (150 mM) for 5 days. Seedlings raised in the presence of additional PUT contained high internal PUT in endosperm and shoots when compared with those raised in distilled water (Table 1). However, the growth of both shoots and roots in NaCl medium was not enhanced by pre- treating seeds with PUT (Fig. 6). Those results are inconsistent with those of Prakash and Prathapasenan [ 101, who demonstrated that pretreatment of rice seeds with PUT could alleviate the inhibitory effect of NaCl on seedling growth The large reduction in PUT in the PUT-treated seedlings placed in NaCl, compared with
Shoot +NaCI -NaCI Shoot +NaCI 0 0 0124 L-Arginine,mM 0 0124 L-Ornithine,mM
Fig. 5. Effect of L-arginine and L-omithine on NaCl-inhibited growth of rice seedlings. The concentration of NaCl was 150 mA4. The growth of rice seedlings was measured after 5 days of treatment. Vertical bars represent standard errors (n = 4).
Table I. Effect of PUT pretreatment on the level of PUT in endosperm, shoots and roots of rice seedlings. Seeds were pre- treated with water or PUT (1 mA4) for 24 h and then transferred to water or NaCl (150 mA4). The levels of PUT in endosperm was determined after 24 h of pretreatment and those in shoots and roots were determined after 5 days of treatment
Treatment PUT (nmol g-l)
Endosperm Shoot Root
Water, 24 h 25.4 f 2.6 - - PUT, 24 h 73.9f1.4 - Water -+ Water - 755 l 68 1079 f 85 PUT + Water - 1043 f 45 1104f44 Water + NaCl - 238 f 24 223 f 15 PUT+ NaCl - 3196 7 245f31 H20 - H,O ImMPUT- H,O H20 --cl50 mM NaCl ImMPUT-150mM NaCl ET! Shoot 0 5 10 15 20 25 H20 -150 mM NaCl ImMPUT-150mMNaCl Root
PUT-treated, water controls is most probably due to the effect of NaCl on metabolism of PUT. Further work is required to clarify this issue.
Taking all data into account, we conclude that the changes in polyamine levels are not associated with the growth inhibition induced by NaCl. The decline in PUT level seems to be part of the overall expression of NaCl effect on rice seedlings.
0 4 8 12
Fresh weight, mg organ-’ Fig. 6. Effect of PUT pretreatment on NaCl-inhibited growth of rice seedlings. Seeds were pretreated with water or PUT (1 mM) for 24 h and then transferred to water or NaCl (150 m&l) for 5 days. Mean f SE, 4 replicates.
Acknowledgement
This work was supported financially by the National Science Council of the Republic of China (NSC83- 0409-B002-034) to C.H.K.
References
1. Altman A (1989) Polyamines and plant hormones. In: Bachrach U and Heimer YM (eds) The Physiology of Polyamines, Vol. 1, pp 121-145. Boca Raton: CRC Press 2. Basu R, Maritra N and Ghosh B (1988) Salinity results
in polyamine accumulation in early rice (Oryza safiva L.) seedlings. Aust J Plant Physiol 15: 777-786
3. Chen CT and Kao CH (199 1) Senescence of rice leaves XXIX. Ethylene production, polyamine level and polyamine biosyn- thetic enzyme activity during senescence. Plant Sci 78: 193- 198
4. Evans PT and Malmberg RL (1989) Do polyamines have roles in plant development? Annu Rev Plant Physiol Plant Mol Biol 40: 235-269
5. Plowers TJ and Yeo AR (1981) Variability in the resistance of sodium chloride salinity within rice varieties. New Phyto188: 363-372
6. Krishnamurthy R (1991) Amelioration of salinity effect in salt tolerant rice (Oryza s&vu L.) by foliar application of putrescine. Plant Cell Physio132: 699-703
7. Krishnamurthy R and Bhagwat KS (1989) Polyamines as mod- ulators of salt tolerance in rice cultivars. Plant Physiol 91: 500-504
8. Mengoli M, Pistocchi R and Bagni N (1989) Effect of long-term treatment of carrot cell cultures with millimolar con- centrations of putrescine. Plant Physiol Biochem 27: l-8 9. Prakash L and Prathapasenan G (1988) Effect of NaCl salinity
and putrescine on shoot growth, tissue ion concentration and yield of rice (Oryza sariva L. GR3). J Agric Crop Sci 160:
325-334
10. Prakash L and Prathapasenan G (1988) Putrescine reduced NaCl-induced inhibition of germination and early seedling growth of rice (Oryza safiva L.). Aust J Plant Physiol 15: 761-767
11. Prakash L, Dutt M and Prathapasenan G (1988) NaCl alters contents of nucleic acids, protein, polyamines and the activity of agmatine deiminase during germination and seedling growth of rice (Oryza sariva L.). Aust J Plant Physiol 15: 769-776