行政院國家科學委員會專題研究計畫 成果報告
人類上皮脂肪酸結合蛋白基因與肺癌細胞轉移能力之相關
性研究
計畫類別: 個別型計畫
計畫編號: NSC91-2314-B-002-229-
執行期間: 91 年 08 月 01 日至 92 年 07 月 31 日
執行單位: 國立臺灣大學醫學院內科
計畫主持人: 鄭之勛
報告類型: 精簡報告
處理方式: 本計畫可公開查詢
中 華 民 國 92 年 10 月 31 日
行政院國家科學委員會研究計畫成果報告 NSC-91-2314-B-002-229
人類上皮脂肪酸結合蛋白基因與肺癌細胞轉
移能力之相關性研究
主持人 鄭之勛 計畫中文摘要 癌細胞之轉移為癌症病患的主要死因之一,發現 癌細胞轉移相關基因仍為目前癌症研究的一大課題。 在先前的研究中,利用微陣列及肺癌轉移模式細胞株 (CL1系列),已經能成功地篩選出許多與癌轉移相關的 基因,其中部份已定性出為新的癌轉移相關基因,甚 至發現其具臨床轉移上的相關性。本研究計畫所要研 究的標的為人類上皮脂肪酸結合蛋白基因FABP5。經 由微陣列篩選檢驗,發現此轉譯此蛋白質之基因的 mRNA 在肺癌腫瘤細胞株的表現,轉移能力較低的細 胞株表現較少,並隨著在轉移能力的增強而有逐步且 明顯的增加。另由聚合脢鏈反應之結果,亦顯示此基 因之表現與肺癌腫瘤細胞株系列之轉移能力可能有正 相關性。本研究計畫中,我們由轉移能力較強的人類 肺癌細胞株CL1-5中選殖FABP5 基因並導入較不具轉 移能力的細胞株CL1-0中以增加其FABP5 的表現量, 藉以產生穩定的選殖細胞株,並以此細胞株進行人體 外之細胞侵襲能力檢定、細胞生長能力鑑定以及轉移 能力檢定,以分析此基因在癌轉移促進的能力。我們 預期在較不具轉移能力的細胞株 CL1-0中過度增加其 FABP5 的表現量會導致細胞侵襲能力與轉移能力的 增加。 關鍵詞: 癌細胞轉移,微陣列,人類上皮脂肪酸結合 蛋白 計畫英文摘要Metastasis remains the major cause of death from cancer. It may be of importance that discovering novel genes may elucidate the detailed mechanism of cancer metastasis. In the previous studies, by using cDNA microarray, we have identified several invasion/ metast- asis-related genes from established lung cancer cell model of metastasis and some them have been considered as novel genes in invasion/metastasis and correlated closely with clinical metastasis. The target of this study is one of the potential metastasis-related genes, human epidermal fatty acid-binding protein (E-FABP) gene FABP5. The results from microarray confirmed that FABP5 mRNA expression was in a lower level in less metastatic cell line such as CL1-0 and was greatly
increased in the highly metastatic cell lines such as CL1-5
and CL1-5-F4. Reverse-transcriptase polymerase chain
reaction (RT-PCR) also showed that the expression of FABP5 mRNA increase was correlated with the metastatic ability of human lung cancer cell lines. In this proposed study, we performed molecular cloning of the 437-base paired, full length cDNA of FABP5 from highly metastatic CL1-5 cell line onto a mammalian
expression plasmid vector, which was then be transfected into a less metastatic human lung adenocarcinoma cell line, CL1-0. The cell line was analyzed the metastasis-
promoting function of FABP5 by in vitro invasion assay,
cell growth assay, and in vitro metastasis assay. We expect that overexpression of FABP5 in the less metastatic human lung adenocarcinoma cell line will enhance their activity of invasiveness and metastasis. Keywords: metastasis, microarray, human fatty acid- binding protein
Introduction
Lung cancer has become one of the leading causes of cancer deaths in Taiwan[1]. The five-year survival rate was only 13% in all patients regardless of their stages of disease at the time of diagnosis[2]. Metastasis is common in patients with lung cancer, especially for small cell lung cancer and adenocarcinoma[3]. Metastasis is a complex process and consists of a series of linked, sequential and selective steps. Invasion of the paren- chyma by cancer cells, usually regarded as the first and probably the most crucial step of metastasis, may involve a series of biological changes concerning the interactions between cancer cells and the surrounding micro- environment[4].
It has been believed that during the development of a malignant tumor, some of the tumor cells may acquire new biologic characteristics, mutations with expression of metastasis-promoting genes or loss of function of metastasis-suppressing genes. A progression toward a more aggressive behavioral pattern may be seen during the progression of tumors, and the ability of invasion may become apparent late in the course of cancer pro- gression.
Several genes that are differentially expressed in the cell lines with different metastatic potential have been identified. One candidate gene was identified as human epidermal fatty acid-binding protein (FABP5).
The intracellular fatty acid-binding proteins (FABPs) are members of a multigene family encoding ~15-kDa proteins, which bind a hydrophobic ligand in a non-covalent, reversible manner[5-7]. The functions of these proteins within cells have remained elusive. Although the in vitro binding of a fatty acid has been analyzed extensively, the in vivo function is less well defined. It has been hypothesized that owing to the low solubility of fatty acids in an aqueous environment the FABPs are required to facilitate their solubilization. Additional proposed functions include facilitating the influx of fatty acids across the plasma membrane, transport of the fatty acids within the cell, determining compartmentalization for storage, modulation of activity of enzymes involved in fatty acid metabolism and protection from detergent-like effects of fatty acids. Currently there are nine different members of FABPs, with strikingly similarly defined functions. Because many of these proteins have been cloned from various organisms, and from different cell or tissue types, each has been named accorading to the location studied. Several types of cells or tissues have been reported to express the epidermal Fabp5, including lens, adipose, mammary and endothelial cells, stratified epithelia of epidermis and tongue, stomach, heart, brain, liver, spleen, muscle, lung, intestine, bone marrow, renal medulla,
testis, urithelium and retina[8-9]. Binding studies have been used to elucidate ligands for FABP5; these include long-chain fatty acids but do not include retinoid acid[10].
FABP5 has been shown to be upregulated in a number of altered states with disturbed lipid profiles. It is highly upregulated in benign papillomas, carcinomas[11], skin warts, and psoriatic keratinocytes[12]. It is upregulated in bladder transitional-cell carcinomas[13] and peripheral nerve trauma. However, the specific func- tions for FABP5 in the various cell types remains unde- termined. Recently in a studies comparing the mRNA expression of benign and malignant human prostate and breast cell lines, the human cutaneous FABP has been shown to express higher levels of mRNAs, and trans- fection of a C-FABP expression construct into the benign, non-metastatic rat mammary epithelia line Rama 37 with subsequent inoculation produced a higher significant number of metastases[14]. Further studies showed that expression of the endogenous vascular endothelial growth factor (VEGF) gene was increased by 3.8-5.2 fold in the C-FABP-transfected cells[15].
Reverse transcriptase – polymerase chain reaction (RT-PCR) also showed that the relative expression of FABP5 is increased in the more invasive and metastatic cell line (CL1-F4 and CL1-5) compared with less invasive
cell lines (CL1-0 and CL1-1). We therefore speculate that
FABP5 may be a metastasis/invasion associated gene in human lung adenocarcinoma and may be of prognostic importance.
In this study, we underwent molecular cloning and characterization of FABP5 involving metastasis/invasion in lung adenocarcinoma cell lines. Assay the metastasis and invasion ability of CL cell lines after transfection of sense and anti-sense cDNA of FABP5 was performed subsequently.
Materials and Methods
Cell culture
A panel of human lung adenocarcinoma cell lines, CL1-0, CL1-1, CL1-5, and CL1-5-F4, in ascending order of
invasiveness, was established in our laboratory[16]. Cells were grown as previously described11. Before functional
assays, only 0.02% EDTA will be used to avoid damage- ing cell-surface antigens.
Isolation of total RNA
Total cellular RNA was extracted from cultured cells as previously described, using a modified guanid- ium thiocyanate-phenol- chloroform extration method by using RNAzol B (Biotecx Laboratories, Inc., Texas, USA). After centrifugation at 12000 g for 15 minutes, it will be washed with cold 75% ethanol and solubilized in TE buffer and quantitated spectrophotometrically.
Northern hybridization
After the RNA from tumor cell lines was extracted, it was used for electrophoresis. Each lane on 1% agarose formaldehyde gel was loaded with 20 µg of total RNA, and after electrophoresis the gel was blotted onto a Hybond M membrane (Amersham). The membrane was hybridized with DIG-labled DNA probes synthesized
using the Rediprime DNA labeling system (Amersham). Hybridization was performed in 5X SSPE, 10X Denhardt’s solution and 100g/ml salmon sperm DNA, 50% deionized formamide for 18 hours at 42℃, and the filters was washed twice in 1X SSC for 15 minutes at room temperature and 0.1X SSC and 0.1% SDS for 30 minutes at 60℃. The amount of RNA in each lane was visualized with hybridization to GAPDH or Gβ-like protein. Densitometric analysis was performed using a phosphorImager (Molecular Dynamics)
Molecular cloning of full-length cDNA of FABP5
RNA from CL1-5 cells was reversely transcribed
with SuperScript II reverse transcriptase (Life Techono- logies, Inc.) and random hexamer. cDNA encoding the entire human FABP5 coding region was amplified by polymerase chain reaction (PCR). Primer sequences are as follows: 5’ primer = 5’-CACCATGGCCAGTTCA-3’ (nucleotides 45-62); 3’ primer = 5’-CCTGTCCAAAGT- GATGATGG-3’ (complementary to nucleotides 481-462). The reaction mix was denatured at 94℃ for 30 seconds, annealed at 55℃ for 3 minutes; this cycle was repeated 30 times. The 437-base pair FABP5 cDNA fragment was cloned into a TA vector, according to the manufacturer’s instructions (pGEM-T-Easy cloning kit; Promega Corp., Madison, WI), and sequenced with an autosequencer (Model ABI 377; PE Applied Biosystem, Foster City, CA). pCI-neo-FABP5 was created by inserting nucleo- tides 45-462 of the FABP5 cDNA between the EcoRI and NotI sites of a pCI-neo mammalian expression vec- tor (Promega Corp.) and used for transfection and ex- pression of FABP5 in CL1-0 cells. Nucleotides 45-462 of
the FABP5 cDNA was inserted into the PstI nad EcoRI sites of a pRSET C prokaryotic expression vector (In- vitrgen Corp., Carsbad, CA) to construct pRSET-FABP5, which was used to produce the protein to immunize mice.
Transfection and selection
PCI-neo plasmids (5µg) was transfected into 70% confluent CL1-0 cells with 20 U of LipofectAMINE re-
agent (Life Technologies, Inc.) as described previous- ly[17]. Other CL1-0 cells were transfected with the
pCI-neo vector containing no insert (mock transfected) as a control. Gentamicin was added to 500 µg/mL for the selection of stable transfectants. Se;ectopm medium will be changed every 3 days for a 3-week period. Clones of resistant cells were isolated and allowed to proliferate for further characterization. Integration of transfected plas- mid DNA into chromosomal DNA was confirmed by Northern blot analysis. For transient Transfection, 70% confluent cultures of CL1-5 and CL1-0 cells were
transfected with the pEGFP-FABP5 plasmid as above. Forty-eight hours later, living cells were examined directly and photographied with a Zeiss Axiphot epifluo- rescence microscopy equipped with an MRC-1000 laser scanning confocal imaging system (Bio-Rad Labora- tories, Rockville Center, NY).
In vitro invasion assay
The invasive activity of transfected clones will be examined by use of a membrane invasion culture system, in which a polycarbonate membrane with 10-µm pores
(Nucleopore Corp., Pleasanton, CA) coated with Matr- igel (Collaborative Biomedical, Becton Dickinson Lab- ware) at 5 mg/mL will be placed between the upper and lower well plates of a membrane invasion culture system chamber. Cells will be suspended in RPMI-1640 medium containing 10% NuSerum, and 2.5×104 cells will be
placed into each upper well of the chamber. After incu- bation for 48 hours at 37℃, cell that had invaded through the coated membrane will be removed from the lower wells with 1 mM EDTA in PBS and dot blotted onto a polycarbonate membrane with 3-µm pores. Blotted cells will be stained with propidium iodide (Sigma Chemical Co., St. Louis, MO), and the number of cells in each blot will be counted under a microscope at a magnification of ×50 by use of Analytical Imaging Station software package (Imaging Research Inc., ON, St. Catharines, Canada). Each experiment will be performed three times, and each sample will be assayed in triplicate.
Cell growth assay
Cell growth will be measured with the modified 3- (4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bro- mide (MTT) assay[18]. Four thousand cells and 100 µL of culture medium per well of a 96-well plate will be incubated for up to 4 days, and 10 µL of MTT (5mg/mL) will be added to each well and incubated for 4 hours at 37℃. The reaction will be stopped by adding 100 μL of 0.04 N HCl in isopropanol to each well, with vigorous mixing to solubilize colored crystals produced by this reaction. The absorbance at 570 nm relative to the absorbance at 630 nm, as the reference wavelength, will be measured by a multiwell scanning spectrophotometer. Cell viability will be examined by trypan blue dye exclusion. Each data point is the average of six deter- minations, and each experiment will be repeated at least three times.
In vitro experimental metastasis
Intravenous metastasis experiment will be used to characterize the lung cancer cell lines transfected with FABP5 (sense transfectant), anti-FABP5 (antisense trans- fectant), and control (mock transfectant). Six-week-old SCID mice will be used for experimental IV metastasis. The cells will be cultured in RPMI + 10% FCS with a short treatment with 0.25% trypsin and 5 mM EDTA, washed 3 times in PBS. The cells will be counted, re- suspended ad adjusted to 107 cells/0.1 mL concentra-
tion. Each group consists of 5 SCID mice and for each mouse 106 cells/0.2 mL will be injected through the tail
vein. Six to 8 weeks after the injections, the mice will be sacrificed and the lungs and livers will be removed and fixed with formalin for histological examination or fro- zen at -70℃ for further experiment.
RESULTS
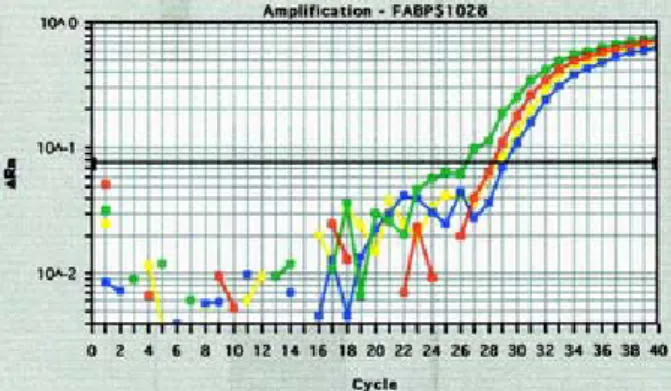
We have performed molecular cloning of the full-length DNA fragment of Fabp5 and have transfected it into a panel of cell lines successfully. The following figure 1 represents one of the stable inducible clone using Tet-off system, containing inducible Fabp5. Real- time reverse-transcription polymerase chain reaction (RT-PCR) of the mRNA expressions of the FABP5 was
performed to compare the gene expression between various cell lines transfected with Fabp5 as well as the parental cell lines. Its expression in this cell line is shown that compared with the parental cell line CL1-0 (the most
right curve) and a cell line that was transfected with vector only (the right second surve), the induced expres- sion of FABP5 mRNA in the Tet-Off cell line (the most left curve) showed an about 4-fold increased of express- ion.
Figure 1. mRNA expression of Fabp5 detected by real-time RT-PCR.
The protein expression of the transfected Fabp5 was also detected in the stable clones of the cell lines containing a vector with a Tet-Off system. (Figure 2). The FABP expression in the transfected gene was detected by an antibody targeting on the HA tag as a fusion protein with FABP. It shows that the current dosage of tetracycline used in this study did not com- pletely suppress the expression of FABP-HA, despite apparent difference. He are currently titrating the dosage of tetracycline to achieve more complete suppression of the transfected FABP5 for further investigations. Trans- fection of Fabp5 to other cancer cell lines was also started recently to find possible more suitable cell lines that may express higher level of FABP5.
Figure 2. Expression of the FABP5-HA fusion protein detected by Westen-blotting.
REFERENCES
1. Magrath I and Litvak J. Cancer in developing countries: opportunity and hcallange. J Natl Cancer
Inst 1993;85:862-866.
2. Pass HI, Mitchell JB, Johnson DH, et al. Lung can- cer, priciples and practaives in: Pass HI ed., Lippin- cott-Raven Press, New York 1996.
3. Feinstein AR, Gelfman NA, Yesner R. The diverse effects of histopathology on manifestations and out-
come of lung cancer. Chest 1974;66:225-229. 4. Liotta LA, Rao CN, Wewer UM. Biochemical inter-
actions of tumor cells with the basement membrane.
Ann Rev Biochem 1986;55:1037-1057.
5. Glatz JF and van der Vusse GJ. Cellular fatty acid- binding proteins: their function and physiological significance. Prog Lipid Res 1996;35:243-82. 6. Bernkohr DA et al. Intracellular lipid-binding pro-
teins and their genes. Annu Rev Nutr 1997;277-303. 7. Coe NR and Bernlohr DA. Physiological properties
and functions of intracellular fatty acid-binding proteins. Biochim Biophys Acta 1998;1391:287-306. 8. Krieg P et al. Tumor-specific overexpression of a
novel keratinocyte lipoid-binding protein. Identification and characterization of a cloned sequence activated during multistage carcinogenesis in mouse skin. J Biol Chem 1993;268:17362-17369. 9. Jaworski C and Wistow G. LP2, a differenttiation-
associated lipid-binding protein expressed in bovine lens. Biochem J 1996;320:49-54.
10. Siegenthaler G et al. Purification and characterizat- ion of the human epidermal fatty acid-binding pro- tein: localization during epidermal cell differentiat- ion in vitro and in vivo. Biochem J 1994;302:363- 371.
11. Krieg P et al. Tumor promoters induce a transient expression of tumor-associated genes in both basal and differentiated cells of the mouse epidermis.
Carcinogenesis 1988;9:95-100.
12. Madsen P et al. Molecular cloning and expression of a novel keratinocyte protein (psoriasis-associated fatty acid-binding protein [PA-FABP]) that is highly upregulated in psoriatic skin and that shares simi- larity to fatty acid-binding proteins. J Invest
Dermatol 1992;99:299-305.
13. Osterggard M et al. Proteome profiling of bladder squamous cell carcinomas: identification of markers that define their degree of differentiation. Cancer
Res 1997;57:4111-4117.
14. Jing C, Beesley C, Foster CS, et al. Identificatino of the messenger RNA for human cutaneous fatty acid- binding protein as a metastasis inducer. Cance Res 2001;60:2390-2398.
15. Jing C, Beesley C, Foster CS, et al. Human cutan- eous fatty acid-binding protein induces metastasis by up-regulating the expression of vascular endothelial growth factor gene in rat Rama 37 model cells.
Cancer Res 2001;61:4357-4364.
16. Chen JJ, Peck K, Hong TM et al. Global analysis of gene expression in invasion by a lung cancer model.
Cancer Res 2001;61:5223-5230.
17. Hong TM, Yang PC, Peck K, et al. Profiling the downstream genes of tumor suppressor PTEN in lung cancer cells by complementary DNA micro- array. Am J Respir Cell Mol Biol 2000;23:355-363. 18. Denizot F, Lang R. Rapid colorimetric assay for cell
growth and survival: modifications to the tetra- zolium dye procedure giving improved sensitivity and reliability. I Immunol Methods 1986;89:271- 277.