Elsevier Editorial System(tm) for Fertility and Sterility Manuscript Draft
Manuscript Number: F and S8110R2
Title: Androgen Excess Down-regulates Connexin43 in a Human Granulosa Cells Line Article Type: Correspondence
Section/Category:
Keywords: Connexin43, androgen, human granulosa cells Corresponding Author: Pao-Lin Kuo, MD
Corresponding Author's Institution: National Cheng Kung University, College of Medicine, Tainan, Taiwan.
First Author: Cheng-Hsuan Wu, M.D.
Order of Authors: Cheng-Hsuan Wu, M.D.; Jyuer-Ger Yang, M.S.; Jiann-Jou Yang, Ph.D.; Yueh-Min Lin, M.D.; Hong-Der Tsai, M.D.; Ching-Yuang Lin, M.D. Ph.D.; Pao-Lin Kuo, M.D.
Abstract: Objective: To investigate the effects of androgen excess on connexin43 (Cx43) expression in human granulosa cells in vitro.
Design and Setting: Controlled experimental study in a Medical Center
Intervention(s): Human granulosa cell line (HO-23) was treated by various dosages of
dihydrotestosterone (DHT). The effect of DHT treatment on granulosa cell Cx43 was assessed by using Western blot. Gap junctional intercellular communication (GJIC) between granulosa cells was
investigated by using the scrape-loading and dye transfer. Androgen receptor antagonist, flutamide, was used to test the specificity of the observed androgen responses.
Main Outcome Measure(s): Cx43 protein expression following DHT treatment.
Result(s): Treatment of the HO-23 cells with DHT showed a dose-dependent decrease in Cx43 protein expression. Flutamide significantly blocked the inhibitory effects of DHT on Cx43 expression. DHT-treated cells demonstrated a decreased enhancement of GJIC as assessed by dye transfer experiments. Conclusion(s): High level androgen reduces Cx43 expression and impairs GJIC between human
granulosa cells through androgen receptors. It may impair folliculogenesis, induce follicular atresia and lead to ovulatory dysfunction.
2010/6/16
Alan H. DeCherney, M.D.
Editor-in- Chief
Fertility and Sterility
Department of Obstetrics and Gynecology
UCLA School of Medicine
10833 Le Conte Avenue, 24-153 CHS
Los Angeles, California 90095-1740
Dear Prof. DeCherney:
We would like to resubmit our manuscript entitled “Androgen Excess
Down-regulates Connexin43 in a Human Granulosa Cells Line” for publication in
your cerebrated journal, Fertility and Sterility.
We have answered the reviewers’ and editorial’s comments in the revision as
below:
Editorial’s comment:
1) We had corrected
Reference 4.
2)
Our abstract had been shortened to two sentences.
Reviewer’s comment:
1) We had corrected into “induce” and “lead”. (
Abstract, lines 30, 31)
2)
We change mammalians into mammalian. (Page 10, line 39)
3)
Figure 1-B, bottom: change B+D+ into B+D+F
In addition, we declare that the material contained in this manuscript has not
been published, or is being submitted elsewhere. There is no interest conflict in this
study.
With best wishes,
Looking forward to hearing from you soon, preferably by E-mail or by Fax.
Pao-Lin Kuo, M.D.
Dept. of Obstetrics and Gynecology, National Cheng Kung University Hospital
138 Victory Road, Tainan 70428, Taiwan
E-mail: [email protected] Fax: +886-6-2766185
Cover Letter1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
F & S 8110 decline and resubmit non-highlighted
Running title: Androgen excess down-regulates Cx43 in granulosa cells
*Manuscript1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
Androgen Excess Down-regulates Connexin43 in a Human Granulosa Cells Line
Cheng-Hsuan Wu M.D.,
a,dJyuer-Ger Yang M.S.,
aJiann-Jou Yang Ph.D.,
cYueh-Min Lin M.D.,
bHong-Der Tsai M.D.,
a. Ching-Yuang Lin M.D. Ph.D.,
e,fPao-Lin Kuo M.D.
ga
Department of Obstetrics and Gynecology ,
bDepartment of Pathology ,
Changhua Christian Hospital, Changhua,
c
Department of Biomedical Science,
dInstitute of Medicine, Chung Shan
Medical University, Taichung,
eChildren‟s Medical Center,
fGraduate Institute of Clinical Medical Science,
China Medical University, Taichung,
gDepartment of Obstetrics and Gynecology , National Cheng Kung University
Hospital, Tainan, Taiwan, R.O.C
*To whom correspondence should be addressed at:
Ching-Yuang Lin, M.D., Ph.D.,
Children‟s Medical Center, China Medical University Hospital,
2 Yuh-Der Road, Taichung, Taiwan.
TEL: +886-4-22052121 ext 2077 FAX: +886-4-22032798
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
Pao-Lin Kuo,M.D.
Department of Obstetrics and Gynecology ,
National Cheng Kung University Hospital,
138 Sheng-Li Road, Tainan, Taiwan 704
Tel: +886-6-2353535 ext 5221
Fax: +886-6-2766185
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
Capsule:
Androgen excess reduces Cx43 expression and impairs GJIC between human
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
By using a cell model, we found high level androgen reduces connexin 43 expression
and impairs gap junction intercellular communication between human granulosa cells
through androgen receptors. This finding suggests that high level androgen may
impair folliculogenesis which in turn leads to ovulatory dysfunction in polycystic
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
Androgen excess is the central defect in polycystic ovarian syndrome (PCOS) patients and androgen excess is believed to be involved in the pathogenesisof follicular development arrest (1,2). Jakimiuk et al. (3) have shown that women with PCOS express elevated levels of 5α reductase mRNA in the granulosa cells. Thus, it is conceivable that the deleterious effects
of androgens in the ovary may be mediated through the conversion to 5α reduced metabolites.
Numerous literatures have shown that normal folliculogenesis relies on the bidirectional talk between granulosa cells and oocytes (4, 5). By facilitating the transfer of ions and small molecules from cell to cell, gap junction intercellular communication (GJIC) between granulosa cells and oocytes plays a critical role in folliculogenesis and oogenesis (6, 7). Connexins (Cx) are membrane-spanning proteins that assemble to form the intercellular channels of gap junctions. At least20 rodent and 21 human connexins have been identified and namedaccording to their molecular weights (8). Gap junctionsbetween ovarian granulosa cells contain predominantly Cx43 which is present at all stages of follicle development(6, 7); while Cx37 is expressed at the oocyte surface in oocyte-somatic cell gapjunctions, with little if any contribution from Cx43 (9; 10). Using chimeric ovaries, Gitten and Kidder (11) demonstrated that Cx37 is required for oocytes but not granulosa cells development; while Cx43 is required for granulosa cells but not oocytes development. The expression level of Cx43 protein is increased as the follicle grows and matures and decreased during follicular atresia (22). Taken together, these data suggested that Cx43 gap junction communication is criticalfor ovarian folliculogenesis.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
hormone, regulate expression of connexins. However, to the best of our knowledge there have
been no reports regarding the effectof androgen on Cx43 expression in human granulosa cells.
We hypothesize that decreased Cx43 expression and GJIC activity are induced by high levels
of androgens in granulosa cells in PCOS ovaries, resulting in impaired folliculogenesis and
anovulation - a hallmark of PCOS. We treated the human granulosa cell line (HO-23) cultured
in vitro with different concentrations of dihydrotestosterone (DHT), a 5α reduced metabolite
of testosterone that has the highest affinity for the androgen receptor (AR) , to investigate the
Cx43 protein levels and to evaluate the effect of androgen on connexins expression in human
granulosa cells.
The HO23 human immortalized luteinized granulosa cell line was provided by Dr. Abraham
Amsterdam, Weizmann Institute of Science, Rehovot 76100, Israel (13). Granulosa cells were
maintained at 37°C in 5% CO2 /air in Dulbecco‟s minimal essential medium (DMEM)/Ham's
F12 (1:1), supplemented with 5% fetal calf serum and antibiotics (100 IU/ml penicillin and
100 μg/ml streptomycin). To test the effect of DHT on Cx43 expression, cells (2 x 106) were
seeded on 100 mm culture dishes and incubated for 24 hours. The media were removed, and
the cells were reincubated in fresh media with different concentrations of DHT. The cells
were harvested for protein assays at 24 hours.
Protein content in cell lysate was determined by the bicinchoninic acid (BCA) protein assay
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
polyvinylidene difluoride membrane (Bio-Rad Laboratories, Ca, USA). The membranes were
blocked with 5% nonfat dry milk in Tris-buffered saline (TBS) for 1 hours and incubated with
mouse anti-human Cx43 antibody (Sigma-Aldrich Co. , USA) (1:3000) in 0.1% nonfat dry
milk overnight at 4°C. After three washes in TBS containing 0.1% Tween-20, the membranes
were incubated with goat anti-mouse IgG (Chemicon, USA) (1:5000) conjugated with
alkaline phosphatase (Sigma-Aldrich) for 1 hour. The membranes were washed again as
above, applied to the enhanced chemilunimescence (ECL) detection kit (Amersham, UK), and
exposed to X-ray film (BioMAx films, Eastman Kodak Company, Rochester, NY, USA. To
assess the amount of Cx43 protein loaded, filterswere treated with 2% SDS and 100 mM β-mercaptoethanol in 62.5 mM Tris-HCl (pH 6.8) for 30 min at 60°C and reprobed for
β-actin. Films were scanned, and the optical density of the bandswas measured with Scion Image. Data shown are representativeof at least three independent experiments with similar
results.
Assessment of GJIC was determinedusing the scrape-loading and dye transfer (SLDT)
technique with a fluorescent dye, LuciferYellow (LY, Sigma, USA) (14). Briefly, human
granulosa cells, cultured as described above, were washed thoroughlywith PBS. SL was
performed applying three cuts on cell monolayerwith a surgical scalpel, and then a
mixture of 1% LY and 1% of rhodamine dextran in PBS was added to the cells.The cells
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
fluorescence. The cells were then fixed with 4% paraformaldehyde and photographed with a
fluorescent microscope equipped with a camera. Cells that receivedthe LY from the
scrape-loaded cells were considered as communicating.The dye-coupled cell layers on either
sides of the scrape were counted to evaluate the GJIC.
Data are expressed as mean ± SEM of at least three independent experiments performed at
different time points. All data were analyzed using one-way analysis of variance (ANOVA).
If the differences were significant, a Dunnett post hoc test was used for post-ANOVA
multiple comparisons. Statistical significance was determined asP < 0.05.
The effect of DHT on Cx43 protein expression in response to 8-Br-cAMP was examined by
Western blot analysis. 8-Br-cAMP, an analogue of cyclic AMP, is known to activate
adenylate cyclase and increase the adenosine cAMP pool in the granulosa cells and
up-regulates the Cx43 expression (15). A previous study showed the range of DHT in
follicularfluid of normal population is 0.77 ± 0.11 ng/ml (16). To demonstrate the effect of
androgen excess on Cx43 expression, cultured human granulosa cells were treated with
increasing doses of DHT at 0, 1, 10, or 100 ng/ml. Western blot analysis of DHT-treated
granulosa cell protein samples demonstrated decreasing amount of Cx43 protein in a
dose-dependent manner. In addition, a significant decrease in the expression could be
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
To assess whether the suppressive effect of DHT on Cx43 is mediated through the AR, cells
were incubated with DHT (10 ng/ml) and flutamide (100 ng/ml, a 10-fold excessabove that of
DHT). Flutamide, an AR antagonist, significantly blocked the inhibitoryeffects of DHT on
Cx43 expression as shown in Figure 1-A.
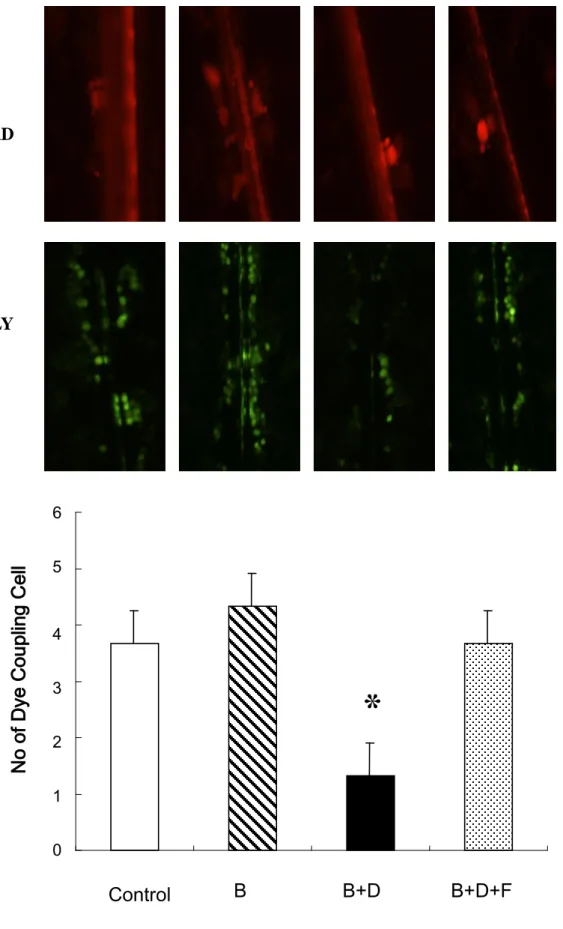
To understand the association between reduced Cx43 expression and GJIC activity, we
assessed the ability of HO-23 granulosa cells to transfer Lucifer yellow through gap junctions
using the scrape-loading dye transfer (SLDT) technique in corresponding cultures of cells
with reduced Cx43 expression. As shown in Figure 1-B, the number of dye-coupled cell
layers was significantly reduced on either side of the scrape in HO-23 granulosa cells grown
in 8-Br-cAMP with DHT medium compared with cells grown in 8-Br-cAMP medium. The
phenomenon wasprevented upon replenishing the medium with flutamide. This gap
junctional defect could be induced with DHT and the effect was recovered with addition of
flutamide to the culture medium.
Although anovulation is frequently observed in PCOS patients with androgen excess, the
underlying mechanism has not been fully elucidated. Several possibilities have been raised to
explain how androgens might mediate anovulation. In this study, we propose a new pathway
through which high androgen level down-regulates Cx43 expression, reduce GJIC in human
granulosa cells, and finally lead to impaired folliculogenesis.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
ovarian physiology in animals (17, 18). One of the targets of this disruptive action is the
granulosa cell within the ovarian follicle. While both androgen and gap junctions are reported
to have important roles in follicular development, our study provides a novel mechanism
which links androgen excess and Cx43 expression in the granulosa cells. We showed reduced
GJIC activity in the milieu of high androgen and reduced Cx43 expression which occurs at
the translation level mediated through androgen receptor.
A link between reduced GJIC activity and reduced connexin gene expression has been
previously reported in rat ovarian cells exposed to luteinizing hormone (LH). LH inhibited
GJIC activity by down-regulating Cx43 synthesis (19). Recently, Wang et al. (20) provided
the evidence that the strength of gap junctional conductance was positively correlated with
Cx43 level in human cumulus cells. Although it is possible that the effect of high androgen on
GJIC activity may be mediated via other junction protein components, our findings indicate
that reduced GJIC activity in human granulosa cells exposed to high androgen is at least in
part due to inhibition of Cx43 gene expression.
The expression and function of Cx43 protein in the ovary of mammalian species have
been discussed previously (21). In the mouse ovary, Cx43 is an important gap junction
protein required for granulosa cell to granulosa cell communication, which in turn is required
for follicle and oocyte maturation (7). Decreased Cx43 protein level is associated with
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
a coordinating role in the process of atresia (12). Disruption of granulosa cell couplingnot
only impairs follicle growth, but also impacts negativelyon the quality of the enclosed
oocytes. The knock-out mice of Cx43 showed that GJIC activity was reduced and oocyte
growth in mutant follicles was retarded (22). However, we know little about Cx43 in human
ovarian follicles and their roles in human folliculogenesis and fertility. A recent study shows
that Cx43 was primarily localized in the membrane where it forms gap junction-like plaques
between human cumulus cells, and its expression level may influence pregnancy outcome
from in vitro fertilization (20). Furthermore, high Cx43 levels in granulosa cells are linked to
good prognosis of human oocytes (9).
The roles of androgen and androgen receptor (AR) are well-established in the
development of male reproductive organs and spermatogenesis. However, the functions of
this steroid and its receptor in the ovary remain elusive. AR expression in the ovary has been
described at various stages of folliculogenesis in several species. In human ovaries, AR
protein expression is observed at different stages in granulosa and theca cells (23). Using a
Cre–loxP system to generate AR-knockout (ARKO) mice, investigators showed that activated
ARs are indispensable for normal folliculogenesis (24, 25). In this study, we demonstrate that
androgen regulates Cx43 expression in cultured human granulosa cells by an AR-dependent
process considering suppressive effectof DHT on Cx43 was blocked by an AR antagonist,
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
treatment ofpatients with PCOS and anovulation. In fact, flutamide is able to restore
ovulation in some women with PCOS (26, 27). However, there is still concern about the use
of flutamide in women with infertility considering its possible teratogenic effect (28).
The root cause of anovulation in PCOS is now thought to be associated with the
significant abnormalities in the very earliest stages offolliculogenesis (29). The normal
„dialogue‟ between oocyte and granulosa cells in these early growing follicles is altered. Recently, Li et al. (30) raised a hypothesis that if dysfunctional communications exist
between granulosa cells or granulosa cells and the oocyte, unnecessary androgen will enter
the oocyte and bind to the AR, provoking some follicles to undergo atresia. We therefore
propose a working mode for the effect of androgen excess on folliculogenesis and oogenesis.
High androgen level down-regulates Cx43 translation and impairs the communication
between granulosa cells and possibly communications between granulosa cells and oocyte.
Impaired communications negatively impacts folliculogenesis. The adverse effect of
androgen excess is further strengthened by “shunting” of excessive androgens to the oocytes secondary to impaired communications between granulosa cells.
In conclusion, the presentstudy provides the first evidence that increased levels of
androgen down-regulate Cx43 expression and impair communications between granulosa
cells. The effect may compromise folliculogenesis and oogenesis in PCOS patients. Caution
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
clinical setting considering few clinical data in this study. In addition, how DHT affects Cx43
translation still awaits further investigation.
Acknowledgment:
This study was supported by grants from the Changhua Christian Hospital
(94-CCH-IRP-07)1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 References
1. Jonard S, Dewailly D. The follicular excess in polycystic ovaries, due to intra-ovarian
hyperandrogenism, may be the main culprit for the follicular arrest. Hum Reprod Update
2004;10:107–17.
2. Homburg R. Androgen circle of polycystic ovary syndrome. Hum Reprod
2009;24:1548-55.
3. Jakimiuk AJ, Weitsman SR and Magoffin DA. 5alpha-reductase activity in women with
polycystic ovary syndrome. J Clin Endocrinol Metab 1999;84:2414–8.
4. Matzuk MM, Burns KH, Viveiros MM, Eppig JJ. Intercellular communication in the
mammalian ovary: oocytes carry the conversation. Science 2002;296:2178–80.
5. Russell DL and Robker RL. Molecular mechanisms of ovulation: co-ordination through
the cumulus complex. Hum Reprod Update 2007;13:289–312.
6. Kidder GM. Roles of gap junctions in ovarian folliculogenesis: Implications for female
infertility. In: Winterhager E ed. Gap Junctions in Development and Disease:
Springer-Verlag New York. 2005; p.223–37.
7. Tong D, Gittens JE, Kidder GM, Bai D. Patch-clamp study reveals that the importance
of connexin43-mediated gap junctional communication for ovarian folliculogenesis is
strain specific in the mouse. Am J Physiol Cell Physiol 2006;290:C290–7.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
membrane channels formed by connexins: their regulation and functions. Physiol. Rev
2003;83:1359–400.
9. Tsai MY, Lan KC, Huang KE, Huang FJ, Kung FT, Chang SY. Significance of mRNA
levels of connexin37, connexin43, and connexin45 in luteinized granulosa cells of
controlled hyperstimulated follicles. Fertil Steril 2003;80:1437–43.
10. Li TY, Colley D, Barr KJ, Yee SP, Kidder GM. Rescue of oogenesis in Cx37-null mutant
mice by oocyte-specific replacement with Cx43. Development 2007;120:4117-25.
11. Gittens JE, Kidder GM. Differential contributions of connexin37 and connexin43 to
oogenesis revealed in chimeric reaggregated mouse ovaries. J Cell Sci
2005;118:5071–8.
12. Cheng Y, Inoue N, Matsuda-Minehata F, Goto Y, Maeda A, Manabe N. Changes in
expression and localization of connexin 43 mRNA and protein in porcine ovary
granulosa cells during follicular atresia. J Reprod Dev 2005;51:627–37.
13. Hosokawa K, Dantes A, Schere-levy C, Barash A, Yoshida Y, Amsterdam A. Induction
of Ad4BP/SF-1, steroidogenic acute regulatory protein, and cytochrome P450scc
enzyme system expression in newly established human granulosa cell lines.
Endocrinology 1998;139:4679–87.
14. El-Fouly MH, Trosko JE, Chang CC. Scrape-loading and dye transfer. A rapid and
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 1987;168: 422–30.
15. Furger C, Cronier L, Poirot C, Pouchelet M. Human granulosa cells in culture exhibit
functional cyclic AMP-regulated gap junctions. Mol Hum Reprod 1996;2:541-8.
16.
diZerega GS, Marrs RP, Lobo R, Ujita EL, Brown J, Campeau JD. Correlation of
inhibin and follicle regulatory protein activities with follicular fluid steroid levels in
anovulatory patients. Fertil Steril 1984;41:849-55.
17. Pradeep PK, Li X, Peegel H, Menon KMJ. Dihydrotestosterone inhibits granulosa cell
proliferation by decreasing the cyclin D2 mRNA expression and cell cycle arrest at G1
phase. Endocrinology 2002;143:2930–5.
18. Zeleznik AJ, Little-Ihrig L, Ramasawamy S. Administration of dihydrotestosterone to
rhesus monkeys inhibits gonadotropin-stimulated ovarian steroidogenesis. J Clin
Endocrinol Metab 2004;89:860–6.
19. Granot I, Dekel N. Phosphorylation and expression of connexin-43 ovarian gap junction
protein are regulated by luteinizing hormone. J Biol Chem 1994;269:30502–9.
20. Wang HX, Tong D, El-Gehani F, Tekpetey FR, Kidder GM. Connexin expression and
gap junctional coupling in human cumulus cells: contribution to embryo quality. J Cell
Mol Med 2009;13:972-84.
21. Gershon E, Plaks V, Dekel N. Gap junctions in the ovary: expression, localization and
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
22. Ackert CL, Gittens JEI, O‟Brien MJ, Eppig JJ, and Kidder GM. Intercellular
communication via connexin43 gap junctions is required for ovarian folliculogenesis in
the mouse. Dev Biol 2001;233:258–70.
23. Walters KA, Allan CM, Handelsman DJ. Androgen actions and the ovary. Biol Reprod
2008;78:380-9.
24. Hu YC, Wang PH, Yeh S, Wang RS, Xie C, Xu Q, et al. Subfertility and defective
folliculogenesis in female mice lacking androgen receptor. Proc Natl Acad Sci USA
2004;101:11209–14.
25. Shiina H, Matsumoto T, Sato T, Igarashi K, Miyamoto J, Takemasa S, et al. Premature
ovarian failure in androgen receptor-deficient mice. Proc Natl Acad Sci USA
2006;103:224–9.
26. De Leo V, Lanzetta D, D‟Antona D, La Marca A, Morgante G. Hormonal effects of
flutamide in young women with polycystic ovary syndrome. J Clin Endocrinol Metab
1998;83:99–102.
27. Eagleson CA, Gringrich MB, Pastor CL, Azora TK, Bust CM, Evans WS, et al.
Polycystic ovarian syndrome: evidence that flutamide restores sensitivity of the
gonadotropin-releasing hormone pulse generator to inhibition by estradiol and
progesterone. J Clin Endocrinol Metab 2000;85:4047–52.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
hyperandrogenism in non-obese adolescents and women. Hum Reprod Update
2006;12:243-52.
29. Franks S, Stark J, Hardy K. Follicle dynamics and anovulation in polycystic ovary
syndrome. Hum Reprod Update 2008;14:367-78.
30. Li M, Schatten H, Sun QY. Androgen receptor's destiny in mammalian oocytes: a new
hypothesis. Mol Hum Reprod 2009;15:149-54.
Figure legends
Figure 1-A. Antiandrogen treatment blocked the inhibitoryeffects of DHT on Cx 43
expression. HO-23 cells were cultured for 24 h under serum-free conditions in the presence of
8-Br-cAMP (0.1 mM), DHT (10ng/ml), and the antiandrogen flutamide (100 ng/ml). Data are
expressed as the mean ± SD of three different experiments. (*: p < 0.05 vs. 8-Br-cAMP)
B: 8-Br-cAMP D: DHT F: flutamide
Figure 1-B: Analysis of GJIC activity in HO-23 cells using SLDT technique. RD remains in
the cells immediately bordering the scrape, while the LY diffuses to neighboring cells in gap
junction-enabled cells. GJIC was evaluated as the counts from dye-coupled cell layers and is
represented as a histogram. GJIC is blocked in HO-23 cells treated with 8-Br-cAMP (0.1 mM)
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
blockage of GJIC induced by DHT to a comparable level to 8-Br-cAMP (ns, not significant
vs 8-Br-cAMP). Data are expressed as the mean ± SD of three different experiments.
B: 8-Br-cAMP D: DHT F: flutamide
GJIC: gap junction intercellular communication
SLDT: scrape-loading and dye transfer
Figure 1-A.
Cx 43
β- Actin
Control B B+D B+F B+D+F
Effect of DHT on the connexin 43 level of human granulosa cell line (HO23)0 20 40 60 80 100 120
*
Opt
ic
D
en
si
ty
FigureFigure 1-B.
LY
RD
0 1 2 3 4 5 6
Control
B
B+D
B+D+F
*
N
o of
Dye
Cou
pling
C
ell
Layer
s
FigureF & S 8110 decline and resubmit highlighted
Running title: Androgen excess down-regulates Cx43 in granulosa cells
*Highlighted RevisionAndrogen Excess Down-regulates Connexin43 in a Human Granulosa Cells Line
Cheng-Hsuan Wu M.D.,
a,dJyuer-Ger Yang M.S.,
aJiann-Jou Yang Ph.D.,
cYueh-Min Lin M.D.,
bHong-Der Tsai M.D.,
a. Ching-Yuang Lin M.D. Ph.D.,
e,fPao-Lin Kuo M.D.
ga
Department of Obstetrics and Gynecology ,
bDepartment of Pathology ,
Changhua Christian Hospital, Changhua,
c
Department of Biomedical Science,
dInstitute of Medicine, Chung Shan
Medical University, Taichung,
eChildren‟s Medical Center,
fGraduate Institute of Clinical Medical Science,
China Medical University, Taichung,
gDepartment of Obstetrics and Gynecology , National Cheng Kung University
Hospital, Tainan, Taiwan, R.O.C
*To whom correspondence should be addressed at:
Ching-Yuang Lin, M.D., Ph.D.,
Children‟s Medical Center, China Medical University Hospital,
2 Yuh-Der Road, Taichung, Taiwan.
TEL: +886-4-22052121 ext 2077 FAX: +886-4-22032798
Pao-Lin Kuo,M.D.
Department of Obstetrics and Gynecology ,
National Cheng Kung University Hospital,
138 Sheng-Li Road, Tainan, Taiwan 704
Tel: +886-6-2353535 ext 5221
Fax: +886-6-2766185
Capsule:
Androgen excess reduces Cx43 expression and impairs GJIC between human
By using a cell model, we found high level androgen reduces connexin 43 expression
and impairs gap junction intercellular communication between human granulosa cells
through androgen receptors. This finding suggests that high level androgen may
impair folliculogenesis which in turn leads to ovulatory dysfunction in polycystic
Androgen excess is the central defect in polycystic ovarian syndrome (PCOS) patients and androgen excess is believed to be involved in the pathogenesisof follicular development arrest (1,2). Jakimiuk et al. (3) have shown that women with PCOS express elevated levels of 5α reductase mRNA in the granulosa cells. Thus, it is conceivable that the deleterious effects
of androgens in the ovary may be mediated through the conversion to 5α reduced metabolites.
Numerous literatures have shown that normal folliculogenesis relies on the bidirectional talk between granulosa cells and oocytes (4, 5). By facilitating the transfer of ions and small molecules from cell to cell, gap junction intercellular communication (GJIC) between granulosa cells and oocytes plays a critical role in folliculogenesis and oogenesis (6, 7). Connexins (Cx) are membrane-spanning proteins that assemble to form the intercellular channels of gap junctions. At least20 rodent and 21 human connexins have been identified and namedaccording to their molecular weights (8). Gap junctionsbetween ovarian granulosa cells contain predominantly Cx43 which is present at all stages of follicle development(6, 7); while Cx37 is expressed at the oocyte surface in oocyte-somatic cell gapjunctions, with little if any contribution from Cx43 (9; 10). Using chimeric ovaries, Gitten and Kidder (11) demonstrated that Cx37 is required for oocytes but not granulosa cells development; while Cx43 is required for granulosa cells but not oocytes development. The expression level of Cx43 protein is increased as the follicle grows and matures and decreased during follicular atresia (22). Taken together, these data suggested that Cx43 gap junction communication is criticalfor ovarian folliculogenesis.
hormone, regulate expression of connexins. However, to the best of our knowledge there have
been no reports regarding the effectof androgen on Cx43 expression in human granulosa cells.
We hypothesize that decreased Cx43 expression and GJIC activity are induced by high levels
of androgens in granulosa cells in PCOS ovaries, resulting in impaired folliculogenesis and
anovulation - a hallmark of PCOS. We treated the human granulosa cell line (HO-23) cultured
in vitro with different concentrations of dihydrotestosterone (DHT), a 5α reduced metabolite
of testosterone that has the highest affinity for the androgen receptor (AR) , to investigate the
Cx43 protein levels and to evaluate the effect of androgen on connexins expression in human
granulosa cells.
The HO23 human immortalized luteinized granulosa cell line was provided by Dr. Abraham
Amsterdam, Weizmann Institute of Science, Rehovot 76100, Israel (13). Granulosa cells were
maintained at 37°C in 5% CO2 /air in Dulbecco‟s minimal essential medium (DMEM)/Ham's
F12 (1:1), supplemented with 5% fetal calf serum and antibiotics (100 IU/ml penicillin and
100 μg/ml streptomycin). To test the effect of DHT on Cx43 expression, cells (2 x 106) were
seeded on 100 mm culture dishes and incubated for 24 hours. The media were removed, and
the cells were reincubated in fresh media with different concentrations of DHT. The cells
were harvested for protein assays at 24 hours.
Protein content in cell lysate was determined by the bicinchoninic acid (BCA) protein assay
polyvinylidene difluoride membrane (Bio-Rad Laboratories, Ca, USA). The membranes were
blocked with 5% nonfat dry milk in Tris-buffered saline (TBS) for 1 hours and incubated with
mouse anti-human Cx43 antibody (Sigma-Aldrich Co. , USA) (1:3000) in 0.1% nonfat dry
milk overnight at 4°C. After three washes in TBS containing 0.1% Tween-20, the membranes
were incubated with goat anti-mouse IgG (Chemicon, USA) (1:5000) conjugated with
alkaline phosphatase (Sigma-Aldrich) for 1 hour. The membranes were washed again as
above, applied to the enhanced chemilunimescence (ECL) detection kit (Amersham, UK), and
exposed to X-ray film (BioMAx films, Eastman Kodak Company, Rochester, NY, USA. To
assess the amount of Cx43 protein loaded, filterswere treated with 2% SDS and 100 mM β-mercaptoethanol in 62.5 mM Tris-HCl (pH 6.8) for 30 min at 60°C and reprobed for
β-actin. Films were scanned, and the optical density of the bandswas measured with Scion Image. Data shown are representativeof at least three independent experiments with similar
results.
Assessment of GJIC was determinedusing the scrape-loading and dye transfer (SLDT)
technique with a fluorescent dye, LuciferYellow (LY, Sigma, USA) (14). Briefly, human
granulosa cells, cultured as described above, were washed thoroughlywith PBS. SL was
performed applying three cuts on cell monolayerwith a surgical scalpel, and then a
mixture of 1% LY and 1% of rhodamine dextran in PBS was added to the cells.The cells
fluorescence. The cells were then fixed with 4% paraformaldehyde and photographed with a
fluorescent microscope equipped with a camera. Cells that receivedthe LY from the
scrape-loaded cells were considered as communicating.The dye-coupled cell layers on either
sides of the scrape were counted to evaluate the GJIC.
Data are expressed as mean ± SEM of at least three independent experiments performed at
different time points. All data were analyzed using one-way analysis of variance (ANOVA).
If the differences were significant, a Dunnett post hoc test was used for post-ANOVA
multiple comparisons. Statistical significance was determined asP < 0.05.
The effect of DHT on Cx43 protein expression in response to 8-Br-cAMP was examined by
Western blot analysis. 8-Br-cAMP, an analogue of cyclic AMP, is known to activate
adenylate cyclase and increase the adenosine cAMP pool in the granulosa cells and
up-regulates the Cx43 expression (15). A previous study showed the range of DHT in
follicularfluid of normal population is 0.77 ± 0.11 ng/ml (16). To demonstrate the effect of
androgen excess on Cx43 expression, cultured human granulosa cells were treated with
increasing doses of DHT at 0, 1, 10, or 100 ng/ml. Western blot analysis of DHT-treated
granulosa cell protein samples demonstrated decreasing amount of Cx43 protein in a
dose-dependent manner. In addition, a significant decrease in the expression could be
To assess whether the suppressive effect of DHT on Cx43 is mediated through the AR, cells
were incubated with DHT (10 ng/ml) and flutamide (100 ng/ml, a 10-fold excessabove that of
DHT). Flutamide, an AR antagonist, significantly blocked the inhibitoryeffects of DHT on
Cx43 expression as shown in Figure 1-A.
To understand the association between reduced Cx43 expression and GJIC activity, we
assessed the ability of HO-23 granulosa cells to transfer Lucifer yellow through gap junctions
using the scrape-loading dye transfer (SLDT) technique in corresponding cultures of cells
with reduced Cx43 expression. As shown in Figure 1-B, the number of dye-coupled cell
layers was significantly reduced on either side of the scrape in HO-23 granulosa cells grown
in 8-Br-cAMP with DHT medium compared with cells grown in 8-Br-cAMP medium. The
phenomenon wasprevented upon replenishing the medium with flutamide. This gap
junctional defect could be induced with DHT and the effect was recovered with addition of
flutamide to the culture medium.
Although anovulation is frequently observed in PCOS patients with androgen excess, the
underlying mechanism has not been fully elucidated. Several possibilities have been raised to
explain how androgens might mediate anovulation. In this study, we propose a new pathway
through which high androgen level down-regulates Cx43 expression, reduce GJIC in human
granulosa cells, and finally lead to impaired folliculogenesis.
ovarian physiology in animals (17, 18). One of the targets of this disruptive action is the
granulosa cell within the ovarian follicle. While both androgen and gap junctions are reported
to have important roles in follicular development, our study provides a novel mechanism
which links androgen excess and Cx43 expression in the granulosa cells. We showed reduced
GJIC activity in the milieu of high androgen and reduced Cx43 expression which occurs at
the translation level mediated through androgen receptor.
A link between reduced GJIC activity and reduced connexin gene expression has been
previously reported in rat ovarian cells exposed to luteinizing hormone (LH). LH inhibited
GJIC activity by down-regulating Cx43 synthesis (19). Recently, Wang et al. (20) provided
the evidence that the strength of gap junctional conductance was positively correlated with
Cx43 level in human cumulus cells. Although it is possible that the effect of high androgen on
GJIC activity may be mediated via other junction protein components, our findings indicate
that reduced GJIC activity in human granulosa cells exposed to high androgen is at least in
part due to inhibition of Cx43 gene expression.
The expression and function of Cx43 protein in the ovary of mammalian species have
been discussed previously (21). In the mouse ovary, Cx43 is an important gap junction
protein required for granulosa cell to granulosa cell communication, which in turn is required
for follicle and oocyte maturation (7). Decreased Cx43 protein level is associated with
a coordinating role in the process of atresia (12). Disruption of granulosa cell couplingnot
only impairs follicle growth, but also impacts negativelyon the quality of the enclosed
oocytes. The knock-out mice of Cx43 showed that GJIC activity was reduced and oocyte
growth in mutant follicles was retarded (22). However, we know little about Cx43 in human
ovarian follicles and their roles in human folliculogenesis and fertility. A recent study shows
that Cx43 was primarily localized in the membrane where it forms gap junction-like plaques
between human cumulus cells, and its expression level may influence pregnancy outcome
from in vitro fertilization (20). Furthermore, high Cx43 levels in granulosa cells are linked to
good prognosis of human oocytes (9).
The roles of androgen and androgen receptor (AR) are well-established in the
development of male reproductive organs and spermatogenesis. However, the functions of
this steroid and its receptor in the ovary remain elusive. AR expression in the ovary has been
described at various stages of folliculogenesis in several species. In human ovaries, AR
protein expression is observed at different stages in granulosa and theca cells (23). Using a
Cre–loxP system to generate AR-knockout (ARKO) mice, investigators showed that activated
ARs are indispensable for normal folliculogenesis (24, 25). In this study, we demonstrate that
androgen regulates Cx43 expression in cultured human granulosa cells by an AR-dependent
process considering suppressive effectof DHT on Cx43 was blocked by an AR antagonist,
treatment ofpatients with PCOS and anovulation. In fact, flutamide is able to restore
ovulation in some women with PCOS (26, 27). However, there is still concern about the use
of flutamide in women with infertility considering its possible teratogenic effect (28).
The root cause of anovulation in PCOS is now thought to be associated with the
significant abnormalities in the very earliest stages offolliculogenesis (29). The normal
„dialogue‟ between oocyte and granulosa cells in these early growing follicles is altered. Recently, Li et al. (30) raised a hypothesis that if dysfunctional communications exist
between granulosa cells or granulosa cells and the oocyte, unnecessary androgen will enter
the oocyte and bind to the AR, provoking some follicles to undergo atresia. We therefore
propose a working mode for the effect of androgen excess on folliculogenesis and oogenesis.
High androgen level down-regulates Cx43 translation and impairs the communication
between granulosa cells and possibly communications between granulosa cells and oocyte.
Impaired communications negatively impacts folliculogenesis. The adverse effect of
androgen excess is further strengthened by “shunting” of excessive androgens to the oocytes secondary to impaired communications between granulosa cells.
In conclusion, the presentstudy provides the first evidence that increased levels of
androgen down-regulate Cx43 expression and impair communications between granulosa
cells. The effect may compromise folliculogenesis and oogenesis in PCOS patients. Caution
clinical setting considering few clinical data in this study. In addition, how DHT affects Cx43
translation still awaits further investigation.
Acknowledgment:
This study was supported by grants from the Changhua Christian Hospital
(94-CCH-IRP-07)References
1. Jonard S, Dewailly D. The follicular excess in polycystic ovaries, due to intra-ovarian
hyperandrogenism, may be the main culprit for the follicular arrest. Hum Reprod Update
2004;10:107–17.
2. Homburg R. Androgen circle of polycystic ovary syndrome. Hum Reprod
2009;24:1548-55.
3. Jakimiuk AJ, Weitsman SR and Magoffin DA. 5alpha-reductase activity in women with
polycystic ovary syndrome. J Clin Endocrinol Metab 1999;84:2414–8.
4. Matzuk MM, Burns KH, Viveiros MM, Eppig JJ. Intercellular communication in the
mammalian ovary: oocytes carry the conversation. Science 2002;296:2178–80.
5. Russell DL and Robker RL. Molecular mechanisms of ovulation: co-ordination through
the cumulus complex. Hum Reprod Update 2007;13:289–312.
6. Kidder GM. Roles of gap junctions in ovarian folliculogenesis: Implications for female
infertility. In: Winterhager E ed. Gap Junctions in Development and Disease:
Springer-Verlag New York. 2005; p.223–37.
7. Tong D, Gittens JE, Kidder GM, Bai D. Patch-clamp study reveals that the importance
of connexin43-mediated gap junctional communication for ovarian folliculogenesis is
strain specific in the mouse. Am J Physiol Cell Physiol 2006;290:C290–7.
membrane channels formed by connexins: their regulation and functions. Physiol. Rev
2003;83:1359–400.
9. Tsai MY, Lan KC, Huang KE, Huang FJ, Kung FT, Chang SY. Significance of mRNA
levels of connexin37, connexin43, and connexin45 in luteinized granulosa cells of
controlled hyperstimulated follicles. Fertil Steril 2003;80:1437–43.
10. Li TY, Colley D, Barr KJ, Yee SP, Kidder GM. Rescue of oogenesis in Cx37-null mutant
mice by oocyte-specific replacement with Cx43. Development 2007;120:4117-25.
11. Gittens JE, Kidder GM. Differential contributions of connexin37 and connexin43 to
oogenesis revealed in chimeric reaggregated mouse ovaries. J Cell Sci
2005;118:5071–8.
12. Cheng Y, Inoue N, Matsuda-Minehata F, Goto Y, Maeda A, Manabe N. Changes in
expression and localization of connexin 43 mRNA and protein in porcine ovary
granulosa cells during follicular atresia. J Reprod Dev 2005;51:627–37.
13. Hosokawa K, Dantes A, Schere-levy C, Barash A, Yoshida Y, Amsterdam A. Induction
of Ad4BP/SF-1, steroidogenic acute regulatory protein, and cytochrome P450scc
enzyme system expression in newly established human granulosa cell lines.
Endocrinology 1998;139:4679–87.
14. El-Fouly MH, Trosko JE, Chang CC. Scrape-loading and dye transfer. A rapid and
1987;168: 422–30.
15. Furger C, Cronier L, Poirot C, Pouchelet M. Human granulosa cells in culture exhibit
functional cyclic AMP-regulated gap junctions. Mol Hum Reprod 1996;2:541-8.
16.
diZerega GS, Marrs RP, Lobo R, Ujita EL, Brown J, Campeau JD. Correlation of
inhibin and follicle regulatory protein activities with follicular fluid steroid levels in
anovulatory patients. Fertil Steril 1984;41:849-55.
17. Pradeep PK, Li X, Peegel H, Menon KMJ. Dihydrotestosterone inhibits granulosa cell
proliferation by decreasing the cyclin D2 mRNA expression and cell cycle arrest at G1
phase. Endocrinology 2002;143:2930–5.
18. Zeleznik AJ, Little-Ihrig L, Ramasawamy S. Administration of dihydrotestosterone to
rhesus monkeys inhibits gonadotropin-stimulated ovarian steroidogenesis. J Clin
Endocrinol Metab 2004;89:860–6.
19. Granot I, Dekel N. Phosphorylation and expression of connexin-43 ovarian gap junction
protein are regulated by luteinizing hormone. J Biol Chem 1994;269:30502–9.
20. Wang HX, Tong D, El-Gehani F, Tekpetey FR, Kidder GM. Connexin expression and
gap junctional coupling in human cumulus cells: contribution to embryo quality. J Cell
Mol Med 2009;13:972-84.
21. Gershon E, Plaks V, Dekel N. Gap junctions in the ovary: expression, localization and
22. Ackert CL, Gittens JEI, O‟Brien MJ, Eppig JJ, and Kidder GM. Intercellular
communication via connexin43 gap junctions is required for ovarian folliculogenesis in
the mouse. Dev Biol 2001;233:258–70.
23. Walters KA, Allan CM, Handelsman DJ. Androgen actions and the ovary. Biol Reprod
2008;78:380-9.
24. Hu YC, Wang PH, Yeh S, Wang RS, Xie C, Xu Q, et al. Subfertility and defective
folliculogenesis in female mice lacking androgen receptor. Proc Natl Acad Sci USA
2004;101:11209–14.
25. Shiina H, Matsumoto T, Sato T, Igarashi K, Miyamoto J, Takemasa S, et al. Premature
ovarian failure in androgen receptor-deficient mice. Proc Natl Acad Sci USA
2006;103:224–9.
26. De Leo V, Lanzetta D, D‟Antona D, La Marca A, Morgante G. Hormonal effects of
flutamide in young women with polycystic ovary syndrome. J Clin Endocrinol Metab
1998;83:99–102.
27. Eagleson CA, Gringrich MB, Pastor CL, Azora TK, Bust CM, Evans WS, et al.
Polycystic ovarian syndrome: evidence that flutamide restores sensitivity of the
gonadotropin-releasing hormone pulse generator to inhibition by estradiol and
progesterone. J Clin Endocrinol Metab 2000;85:4047–52.
hyperandrogenism in non-obese adolescents and women. Hum Reprod Update
2006;12:243-52.
29. Franks S, Stark J, Hardy K. Follicle dynamics and anovulation in polycystic ovary
syndrome. Hum Reprod Update 2008;14:367-78.
30. Li M, Schatten H, Sun QY. Androgen receptor's destiny in mammalian oocytes: a new
hypothesis. Mol Hum Reprod 2009;15:149-54.
Figure legends
Figure 1-A. Antiandrogen treatment blocked the inhibitoryeffects of DHT on Cx 43
expression. HO-23 cells were cultured for 24 h under serum-free conditions in the presence of
8-Br-cAMP (0.1 mM), DHT (10ng/ml), and the antiandrogen flutamide (100 ng/ml). Data are
expressed as the mean ± SD of three different experiments. (*: p < 0.05 vs. 8-Br-cAMP)
B: 8-Br-cAMP D: DHT F: flutamide
Figure 1-B: Analysis of GJIC activity in HO-23 cells using SLDT technique. RD remains in
the cells immediately bordering the scrape, while the LY diffuses to neighboring cells in gap
junction-enabled cells. GJIC was evaluated as the counts from dye-coupled cell layers and is
represented as a histogram. GJIC is blocked in HO-23 cells treated with 8-Br-cAMP (0.1 mM)
blockage of GJIC induced by DHT to a comparable level to 8-Br-cAMP (ns, not significant
vs 8-Br-cAMP). Data are expressed as the mean ± SD of three different experiments.
B: 8-Br-cAMP D: DHT F: flutamide
GJIC: gap junction intercellular communication
SLDT: scrape-loading and dye transfer
We have answered the editorial’s and reviewers’ comments in the revision as
below:
Editorial’s comment:
1) We had corrected
Reference 4.
2)
Our abstract had been shortened to two sentences.
Reviewer’s comment:
1) We had corrected into “induce” and “lead”. (
Abstract, lines 30, 31)
2)
We change mammalians into mammalian. (Page 10, line 39)
3)
Figure 1-B, bottom: change B+D+ into B+D+F
*Revision NotesManuscript number_______________________ Conflict of interest disclosure statement:
A copy of this form must be completed and signed by each author. All authors must disclose any commercial interest, financial interest, and/or other relationship with manufacturers of pharmaceuticals, laboratory supplies, and/or medical devices and with commercial providers of medically related services. All relationships must be disclosed. All non-FDA-approved uses of products must be clearly identified.
Commercial interest is defined as any proprietary entity producing, marketing, re-selling, distributing, or otherwise participating in or profiting from the distribution, promotion, or sale of health care goods or services consumed by, or used on, patients.
Financial interests/relationships are those in which the individual benefits by receiving a salary, royalty, intellectual property rights, consulting fee, honoraria, ownership interest (e.g., stocks, stock options, or other ownership interest, excluding diversified mutual funds), or other financial benefit. Financial benefits usually are associated with roles such as employment, management position, independent contractor (including contracted research), consulting, speaking and teaching, membership on advisory committees or review panels, board membership, and other activities from which remuneration is received or expected. This includes any financial relationships within the last twelve months, as well as known financial relationships of your spouse or partner.
Please check one of the following:
_V_ Neither I nor my spouse/partner has a commercial interest, financial interest, and/or other relationship with manufacturers of pharmaceuticals, laboratory supplies, and/or medical devices or with commercial providers of medically related services.
____ I have a commercial interest, financial interest or other relationship with manufacturers of pharmaceuticals, laboratory supplies, and/or medical devices or with commercial providers of medically relation services. Please list interest(s) or relationship(s). (If more room is needed, attach a second page.)
Source:____________________________Relationship:_________________________________ Source:____________________________Relationship:_________________________________ Source:____________________________Relationship:_________________________________
____ My spouse/partner has a commercial interest, financial interest, and/or other relationship with manufacturers of pharmaceuticals, laboratory supplies, and/or medical devices or with commercial providers of medically related services.
Please list source(s) or relationship(s). (If more room is needed, attach a second page.) Source:____________________________Relationship:_________________________________ Source:____________________________Relationship:_________________________________ Source:____________________________Relationship:_________________________________ ____ I refuse to disclose my commercial interest(s), financial interest(s), and/or other relationship(s) with manufacturers of pharmaceuticals, laboratory supplies, and/or medical devices and with commercial providers of medically related services. Authors who check this box will not be allowed to submit articles and reviewers will not be allowed to review
manuscripts for publication.
Print name:_____