Mycological Society of America
A New Species of Endophragmiella from Taiwan
Author(s): S. S. Tzean and J. L. Chen
Source: Mycologia, Vol. 81, No. 5 (Sep. - Oct., 1989), pp. 800-805
Published by: Mycological Society of America
Stable URL:
http://www.jstor.org/stable/3759887
Accessed: 28/01/2010 21:05
Your use of the JSTOR archive indicates your acceptance of JSTOR's Terms and Conditions of Use, available at
http://www.jstor.org/page/info/about/policies/terms.jsp
. JSTOR's Terms and Conditions of Use provides, in part, that unless
you have obtained prior permission, you may not download an entire issue of a journal or multiple copies of articles, and you
may use content in the JSTOR archive only for your personal, non-commercial use.
Please contact the publisher regarding any further use of this work. Publisher contact information may be obtained at
http://www.jstor.org/action/showPublisher?publisherCode=mysa
.
Each copy of any part of a JSTOR transmission must contain the same copyright notice that appears on the screen or printed
page of such transmission.
JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of
content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms
of scholarship. For more information about JSTOR, please contact [email protected].
Mycological Society of America is collaborating with JSTOR to digitize, preserve and extend access to
Mycologia.
Mycologia, 81(5), 1989, pp. 800-805.
? 1989, by The New York Botanical Garden, Bronx, NY 10458-5126
A NEW SPECIES OF ENDOPHRAGMIELLA FROM TAIWAN
S. S. TZEAN AND J. L. CHENDepartment of Plant Pathology and Entomology, National Taiwan University, Taipei, Taiwan, 10476 Endophragmiella was proposed by Sutton
(1973) to accommodate two species, E. palles- cens Sutton, and E. canadensis (Ell. & Ev.) Sut- ton. Separation of Endophragmiella from the morphologically similar genus Endophragmia was based on several features. For example, En- dophragmiella was characterized by thin-walled conidiophores and conidiogenous cells, conid- iophores simple or irregularly branched, without producing flared cup-shaped collarettes, and conidia bearing a distinct frill (Sutton, 1973). Hughes (1979) indicated that many species in- cluded in Endophragmia were untenable, be- cause they seceded conidia rhexolytically and showed percurrent proliferation arising from the penultimate cells of the conidiophores. The mechanism of conidiogenesis and conidiophore proliferation actually did not comply with the definition of Endophragmia proposed by Du- vernoy and Maire (1920). Many previously de- scribed species of Endophragmia (Ellis, 1971, 1976), therefore, were transferred to Endophrag- miella after its generic concept was redefined (Hughes, 1979). Accepting the concept of Hughes (1979), Kirk published a series of papers describ- ing many new species of Endophragmiella (Kirk, 1981a, b, 1982a, b, c), and provided a key to the 46 described species (Kirk, 1985). During inves- tigations of the hyphomycetes of Taiwan asso- ciated with decaying leaves and stems, a conidial fungus was isolated, conidia of which secede rhexolytically and had a frill at their base. The terminal cell of the conidiophore proliferated percurrently to form a new conidiogenous cell. These characteristics suggested a relationship to Endophragmiella. Nevertheless, the shape, size, septation and pigmentation of conidia and co- nidiophores distinguish this fungus from other described Endophragmiella species (Ellis, 1971, 1976; Hughes, 1979; Kirk, 1981a, b, 1982a, b, c, 1985), and warrant accommodation in a new species. For the diagnosis, Kornerup and Wanscher (1978) was used as a color standard.
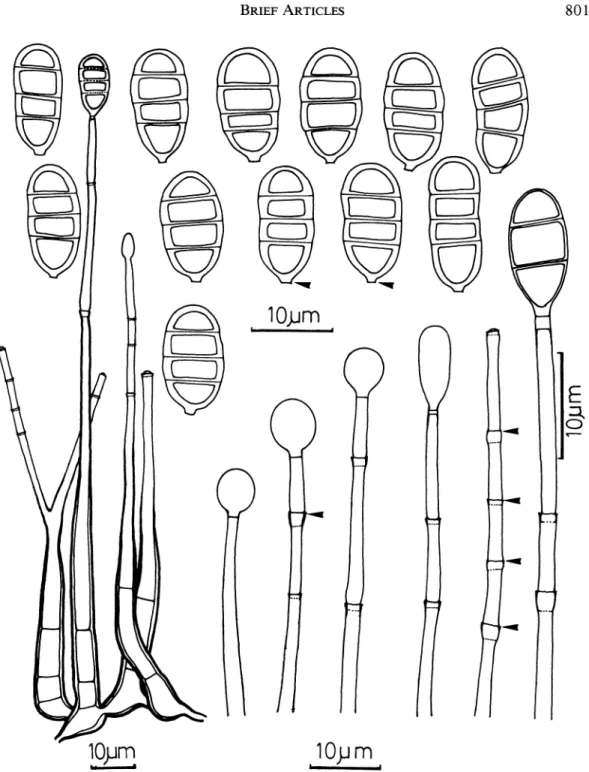
Endophragmiella achromatica sp. nov. FIGS. 1-3 Coloniae on CMA (Zea mays L. farinosis agaribus) effusae, aurantiacae albae ad pallidam aurantiacam; reversae obscure aurantiacae albae ad obscure auran- tiacas umbras. Mycelium fere immersum in substrato, constituta laeve, ramosum, septatum; hyphae hyalinae. Conidiophorum macronematoideum, mononemato- ideum, oriens singulatim vel 3-5 in caterva ab mycelio vel intercalaris moniliformis cellulae, gracile, strictum vel leviter flexuosum, laeve, septatum, hyalinum ad laete pallidum brunneum, 70-160 ,m altum, cylin- dricum ad subulatum, contractum ad finem, cum cras- so-pariete inflata base, 4.3-11 im latum, simplex vel raro ramosum, cum usque ad 10 percurrens prolife- rum. Cellulae conidiogenae, monoblastic, integrae, ter- minales, percurrentes, contractae ad apicem. Conidia solitaria, exsiccata, acroportata, ovoidea, vel late ob- longata, ferens angustus, cylindraceus protuberans abs- cissus hilum ad basim, 0.4-1.6 im longam et 2 um latam, 3-septata, paries conidicus usque ad 1.6 Am crassum, et septa usque ad 1.3 um crass, laeves, hy- alinae, 12.2-21.3 x 5.8-9.2 ,m.
Colonies on corn meal agar effused, orange white to pale orange; reverse dull orange-white to dull orange shades. Mycelium mostly im- mersed in the substratum, composed of smooth, branched, septate, hyaline hyphae. Conidio- phores macronematous, mononematous, arising singly or 3-5 in a group from mycelium or in- tercalary moniliform cells, slender, straight or slightly flexuous, smooth, septate, hyaline to light pale brown, 70-160 Lm high, cylindrical to subu- late, tapered toward the end, with thick-walled swollen base, 4.3-11 Am wide, simple or rarely branched. Conidiogenous cells monoblastic, in- tegrated, terminal, tapered toward apex, with up to 10 percurrent proliferations. Conidia solitary, dry, acrogenous, ovoid or broadly oblong, 12.2- 21.3 x 5.8-9.2 ium bearing a basal narrow, cy- lindrical protuberant abscission scar, 0.4-1.6 um long and 2 Am wide; conidial wall up to 1.6 Am thick, smooth, hyaline, 3-septate; septa up to 1.3 tzm thick.
HABITAT. On rotten stems collected from Wulai, Tai-
pei, Taiwan, on 4.III. 1987. HOLOTYPUS: PPH4 (dried culture) and ex-typus PPH4E, deposited in the De-
800
BRIEF ARTICLES
10pJm
IL I
10im
1 0j m
FIG. 1. Conidiophores, conidiogenous cells, conidia, and conidiogenesis of Endophragmiella achromatica. Arrows indicate the conidium basal frill, collarette, and percurrent proliferation of conidiogenous cells.
partment of Plant Pathology and Entomology, Nation- ISOTYPUS: PPH4-2 (dried culture), and ex-typus in al Taiwan University, Taipei, Taiwan; ex-typus de- Commonwealth Mycological Institute.
posited in the Culture Collection and Research Center
(CCRC), Hsinchu, Taiwan. ISOTYPUS: PPH4-1 (dried Hughes (1979) redefined Endophragmiella to culture) also deposited in New York Botanical Garden; include only species in which conidia secede
801
B
*A
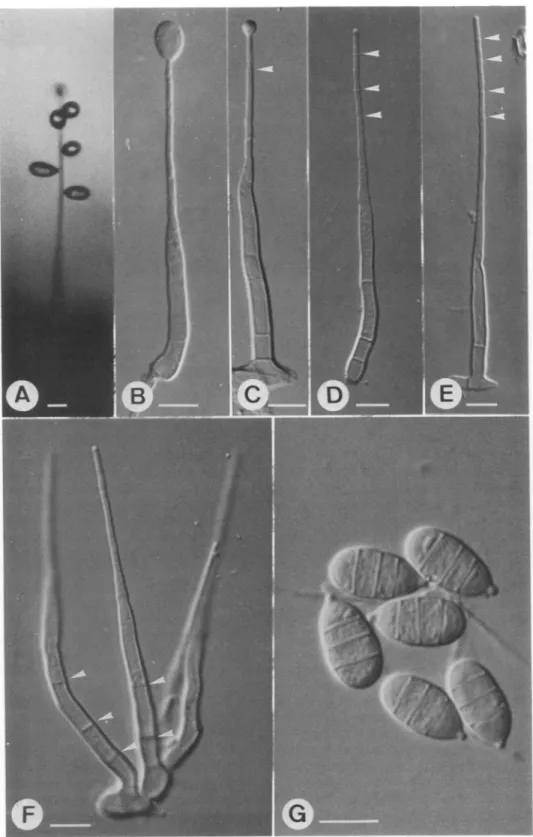
FIG. 2. Morphological characteristics ofEndophragmiella achromatica. A. Habit, without disturbance, under humid condition. The seceded conidia push aside, but still remain adherent on the conidiophore. B. First conidial initial on the conidiophore, judging by the lack of any collarette. C. Percurrent proliferation of a second conidium. The first conidium has seceded, and the collarette remains (arrow). D, E. Third and fourth collarettes (arrows),
implying that 2-3 conidia have been formed and seceded. F. Conidiophores in a group of four arising from the foot-like cells showing permanent septa (arrows). G. Conidia broadly ovoid, some with prominent abscission
hilum. Bar = 10 l,m.
'? ?-.
?b;:'-?-?:;
.?.
I
'
,? ?
: :r
.r....' .:. +: 1.? ';"?? -.p*? ?t . r - -? -- ...: .. .i ? ?. - r ;?; - ?1 ? a'?""'' .; ? :..:i i. ..* L .r.? ? ? ccrr- ?,'A
^^-^^
i.
-
,1fc
^^?
^!
w-
sL~c
. .. -. --ij '-4i(
w-
AWO
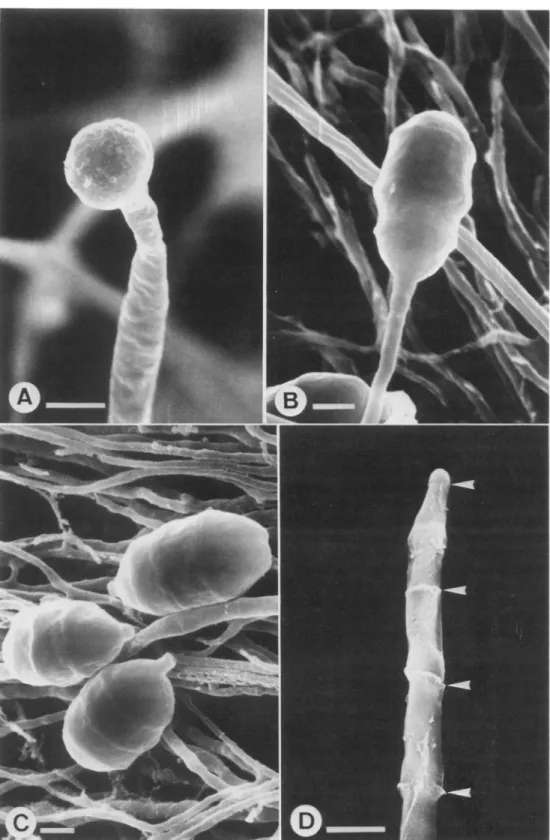
4Jm.-FIG. 3. Scanning electron micrographs of Endophragmiella achromatica. A. Conidiogenous cell bearing a monoblastic, terminal, immature conidium. In this incipient stage, the conidium is more or less subglobose. B. An almost mature conidium on a conidiogenous cell. C. Mature and detached conidia with hilum. D. Collarettes left over on the conidiophore (arrows) after conidia have seceded. Bar = 2 Mm.
MYCOLOGIA
TABLE I
MORPHOLOGICAL CHARACTERISTICS OF ENDOPHRAGMIELLA SPECIES
Conidia
Species Shape Septation Pigmentation Size (tm)
E. bisbyi obovoid to pyri- predominantly brown above, pale be- 12.5-16 x 6.3-7.6 form, distal 3-septate low, distal septum
cell largest partly obscured by
dark band
E. eboracensis narrowly ellipsoi- (1-)3-septate evenly brown to dark 17-21.5 x 5.4-6.6
dal, cylindri- with basal or apical
cal, narrowly cell paler
obvoid
E. theobromae obovoid, central predominantly very pale brown, bas- 18-27 x 9-11.7 cell shortest 2-septate al cell subhyaline
E. tripartita ellipsoid, broadly predominantly brown to dark brown 18-21.5 x 8-9.5
obovoid 2-septate
E. achromatica ovoid to broadly predominantly hyaline 12.2-21.3 x 5.8-9.2
oblong 3-septate
rhexolytically and in which conidiogenous cells proliferate percurrently from the penultimate cell of conidiophores. Conidium secession and per- current proliferations could be divided into three major models. First, like E. pallescens, the break in the wall of the conidiogenous cell is well below the apex, resulting in a conspicuous frill around the conidium scar. Second, as in E. boewei (Crane) Hughes, the mature conidium secedes by a clean, rhexolytic break at a visible line of dehiscence just below the base of the conidium. The conid- ium, therefore, is not ornamented with a prom- inent basal frill. Third, in some Endophragmiella species a septum may fail to develop during pro- liferation, so that no new conidiogenous cell is delimited. Thus, the conidiogenous cells contin- ue to elongate after completion ofconidiogenesis and bear longer linear fragments (Hughes, 1979). In E. achromatica, more or less similar to E. albiziae (Ellis) Hughes, E. boewei, or E. ebora- censis Sutton, at first the apex of the conidio- phore expands into a conidial initial (FIGS. 1, 3A). The initial developed and became ovoid or broadly oblong in shape, and laid down three transverse septa (FIG. 1, 3B). Subsequently, a thin septum is then formed at a very short distance (0.5-2 ,um) below the thickened base of the co- nidium. The conidium matures and a newly dif- ferentiated terminal cell of the conidiophore is vacuolated or presumably emptied of its con- tents, judging by lack of staining by 0.1% cotton blue in lactophenol. The mature conidium then secedes by a rhexolytic break along a dehisced line just below the thickened but narrow hilar
basal abscission scar, and bears a barely visible frill. When the conidium matures and secedes, a collarette is left over; the septum (adventitious or temporary) which becomes elastic, then pro- liferates percurrently. After ca 5-10 percurrent proliferations, occasional additional secondary (permanent) septa are then usually laid down at irregular intervals in acropetal succession.
The length and width of the conidium scar in E. achromatica is comparable to E. albiziae (Hughes, 1979). The holoblastic, integrated, ter- minal and percurrent conidiogenesis exhibited by E. achromatica is further verified by scanning electron microscopy (FIG. 3A-D). The conidi- ogenous cell bulges out to form a conidial initial which eventually matures and becomes broadly oblong, smooth but with slight constriction.
In light of the key to Endophragmiella species provided by Kirk (1985), E. achromatica shows morphological similarities to E. bisbyi (Sutton) Hughes, E. eboracensis, E. theobromae Ellis and E. tripartita Hughes. On the other hand, based on distinctions of shape, size, septation, pigmen- tation, basal frill of the conidia, conidiophore and conidiogenous cell (Hughes, 1979; Kirk, 1985), separation of these four species is main- tained (TABLE I).
ACKNOWLEDGMENTS
This work was supported by the National Science Council Taiwan, grant NSC-0409-B002-64. The au- thors are indebted to T. Matsushima for invaluable taxonomic comments, and to Dr. J. C. Liao for prep- aration of the Latin diagnosis.
BRIEF ARTICLES BRIEF ARTICLES Key Words: Endophragmiella, hyphomycetes, taxon-
omy
LITERATURE CITED
Duvernoy, A., and R. Maire. 1920. Une nouvelle
Dematiee a conidies pseudo-endogenes. Bull. Soc.
Mycol. France 36: 86-89.
Ellis, M. B. 1971. Dematiaceous Hyphomycetes.
Commonwealth Mycological Institute.
. 1976. More Dematiaceous Hyphomycetes. Commonwealth Mycological Institute.
Hughes, S. J. 1979. Relocation of species of Endo- phragmiella auct. with notes on relevant generic names. New Zealand J. Bot. 17: 139-188. Kirk, P. M. 198 la. New or interesting microfungi. I.
Dematiaceous hyphomycetes from Devon. Trans. Brit. Mycol. Soc. 76: 71-87.
. 198 b. New or interesting microfungi. II. Dematiaceous hyphomycetes from Esher Com- Key Words: Endophragmiella, hyphomycetes, taxon-
omy
LITERATURE CITED
Duvernoy, A., and R. Maire. 1920. Une nouvelle
Dematiee a conidies pseudo-endogenes. Bull. Soc.
Mycol. France 36: 86-89.
Ellis, M. B. 1971. Dematiaceous Hyphomycetes.
Commonwealth Mycological Institute.
. 1976. More Dematiaceous Hyphomycetes. Commonwealth Mycological Institute.
Hughes, S. J. 1979. Relocation of species of Endo- phragmiella auct. with notes on relevant generic names. New Zealand J. Bot. 17: 139-188. Kirk, P. M. 198 la. New or interesting microfungi. I.
Dematiaceous hyphomycetes from Devon. Trans. Brit. Mycol. Soc. 76: 71-87.
. 198 b. New or interesting microfungi. II. Dematiaceous hyphomycetes from Esher Com-
mon, Surrey. Trans. Brit. Mycol. Soc. 77: 279- 298.
. 1982a. New or interesting microfungi. IV. Dematiaceous hyphomycetes from Devon. Trans. Brit. Mycol. Soc. 77: 55-74.
. 1982b. New or interesting microfungi. V. Microfungi colonizing Laurus nobilis leaf litter. Trans. Brit. Mycol. Soc. 78: 293-303.
1982c. New or interesting microfungi. VII. Two new hyphomycetes from Pericopsis angolen- sis leaf. Mycologia 74: 872-876.
? 1985. New or interesting microfungi. XIV. Dematiaceous hyphomycetes from Mt. Kenya. Mycotaxon 23: 305-352.
Kornerup, A., and J. H. Wanscher. 1978. Methuen
Handbook of Colour. 3rd Ed. Eyre Methuen, Lon- don.
Sutton, B. C. 1973. Hyphomycetes from Manitoba and Saskatchewan, Canada.Mycol. Pap. 132: 1- 143.
mon, Surrey. Trans. Brit. Mycol. Soc. 77: 279- 298.
. 1982a. New or interesting microfungi. IV. Dematiaceous hyphomycetes from Devon. Trans. Brit. Mycol. Soc. 77: 55-74.
. 1982b. New or interesting microfungi. V. Microfungi colonizing Laurus nobilis leaf litter. Trans. Brit. Mycol. Soc. 78: 293-303.
1982c. New or interesting microfungi. VII. Two new hyphomycetes from Pericopsis angolen- sis leaf. Mycologia 74: 872-876.
? 1985. New or interesting microfungi. XIV. Dematiaceous hyphomycetes from Mt. Kenya. Mycotaxon 23: 305-352.
Kornerup, A., and J. H. Wanscher. 1978. Methuen
Handbook of Colour. 3rd Ed. Eyre Methuen, Lon- don.
Sutton, B. C. 1973. Hyphomycetes from Manitoba and Saskatchewan, Canada.Mycol. Pap. 132: 1- 143.
Mycologia, 81(5), 1989, pp. 805-811.
? 1989, by The New York Botanical Garden, Bronx, NY 10458-5126
INFECTION OF WOODLAND GRASSES BY FUNGAL ENDOPHYTES
KEITH CLAY AND ADRIAN LEUCHTMANN
Department of Biology, Indiana University, Bloomington, Indiana 47405 Mycologia, 81(5), 1989, pp. 805-811.
? 1989, by The New York Botanical Garden, Bronx, NY 10458-5126
INFECTION OF WOODLAND GRASSES BY FUNGAL ENDOPHYTES
KEITH CLAY AND ADRIAN LEUCHTMANN
Department of Biology, Indiana University, Bloomington, Indiana 47405 Many grasses are known to harbor systemic
clavicipitaceous endophytes within leaves, stems, and seeds. The fungi are classified in the tribe Balansieae (Clavicipitaceae) including the form genus Acremonium sect. Albo-lanosa Morgan- Jones & Gams which accommodates the ana- morphs ofEpichlo (Fr.) Tul. (Diehl, 1950; Mor- gan-Jones and Gams, 1982). Previous work has shown that this association is often mutualistic with host plants exhibiting increased growth and resistance to insect herbivores compared to un- infected plants (Clay, 1988). Most research has focused on two economically important forage and turf grasses, Festuca arundinacea Schreb. and Lolium perenne L., infected by A. coenophialum Many grasses are known to harbor systemic clavicipitaceous endophytes within leaves, stems, and seeds. The fungi are classified in the tribe Balansieae (Clavicipitaceae) including the form genus Acremonium sect. Albo-lanosa Morgan- Jones & Gams which accommodates the ana- morphs ofEpichlo (Fr.) Tul. (Diehl, 1950; Mor- gan-Jones and Gams, 1982). Previous work has shown that this association is often mutualistic with host plants exhibiting increased growth and resistance to insect herbivores compared to un- infected plants (Clay, 1988). Most research has focused on two economically important forage and turf grasses, Festuca arundinacea Schreb. and Lolium perenne L., infected by A. coenophialum
Morgan-Jones & Gams and A. lolii Latch et al., respectively (Funk et al., 1983; Latch et al., 1985; Clay, 1986; Bacon and Siegel, 1988).
Little is known about community level pat- terns of infection frequency; for example, are there differences between infection frequency of grass- es from early vs late successional habitats, wood- land vs grassland habitats, or tropical vs temper- ate communities? While there have been several surveys of endophyte-infected grasses based on herbarium sheets and seed collections (White, 1987; Latch et al., 1987), they have been at a large scale and not focused on all the grass species within a particular community. There have also been several field surveys of endophyte-infected Morgan-Jones & Gams and A. lolii Latch et al., respectively (Funk et al., 1983; Latch et al., 1985; Clay, 1986; Bacon and Siegel, 1988).
Little is known about community level pat- terns of infection frequency; for example, are there differences between infection frequency of grass- es from early vs late successional habitats, wood- land vs grassland habitats, or tropical vs temper- ate communities? While there have been several surveys of endophyte-infected grasses based on herbarium sheets and seed collections (White, 1987; Latch et al., 1987), they have been at a large scale and not focused on all the grass species within a particular community. There have also been several field surveys of endophyte-infected
805 805