Association between XRCC3 Thr241Met SNP and systemic lupus erythematosus in Han Chinese patients in Taiwan, and a meta-analysis of healthy populations
Yng-Tay Chen1, Shih-Yin Chen1, 2, Ying-Ju Lin1, 2, Chung-Ming Huang3, Yuan-Yen Chang,4 Fuu-Jen Tsai1, 2,5,*
1 Human Genetic Center, China Medical University Hospital, Taichung, Taiwan. 2 Graduate Institute of China Medical Science, China Medical University, Taichung, Taiwan.
3 Division of Immunology and Rheumatology, China Medical University Hospital, Taichung, Taiwan.
4 Department of Microbiology and Immunology, and Institute of Microbiology and Immunology, Chung Shan Medical University, Taichung, Taiwan.
5College of Health Science, Asia University, Taichung, Taiwan.
Running Title: XRCC3 SNP and SLE
Y-T Chen and S-Y Chen contributed equally to this paper.
*Correspondence to: Fuu-Jen Tsai, Human Genetic Center, China Medical University Hospital, 2 Yuh-Der Road, Taichung 40447, Taiwan.
Email: [email protected] 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21
Abstract
X-ray repair cross-complementing group 3 (XRCC3) plays a crucial role in mammalian DNA repair processes. The polymorphism of XRCC3, rs861539 (Thr > Met at codon 241), is common in populations worldwide. This study analyzed the relationship between this functional single nucleotide polymorphism (SNP) and systemic lupus erythematosus (SLE) in the Han Chinese population in Taiwan (HC-TW). Genotyping was performed using polymerase chain reaction restriction fragment length polymorphism (PCR-RFLP) on 163 SLE patients and 191 healthy participants in the control group. The data showed that the genotype frequency at codon 241 did not differ significantly between the SLE patients and the healthy participants in the control group; however, the allele frequency analysis indicated a significant difference between these groups. In addition, we used the genotype and allele frequencies of 191 healthy HC-TW participants for comparison with HapMap populations. The results indicated a significant difference of XRCC3 Thr241Met allele and genotype frequencies between the HC-TW population and HapMap populations, except for the other Han Chinese populations. A prior study showed that Thr241 > Met substitution in XRCC3 protein was positive as damaging and functional consequences as well. This is the first study to demonstrate the difference of XRCC3 Thr241 > Met variant between the HC-TW population and HapMap population.
Keywords: systemic lupus erythematosus (SLE); X-ray repair cross-complementing group 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40
3 (XRCC3); Single nucleotide polymorphisms (SNPs); Genotype; Haplotype.
Introduction
Systemic lupus erythematosus (SLE) is a complex multi-factorial auto-immune disease that is characterized by B-cell hyperactivity and auto-antigen production against nuclear constituents and other self-antigens (1, 2). SLE may be induced by genetic, hormonal, environmental factors, viral infection (3, 4). Exposure to ultraviolet (UV) light are considered crucial environmental factors (5). Defective immune regulatory mechanisms induced by genetic mutations can result in the dysregulation of inflammatory cytokine production and insufficient clearance of apoptotic cells or immune complexes, which activate anti-self-immune responses (6-9). Single nucleotide polymorphism (SNP) typing can be used to determine the potential molecular mechanisms in whole genomes (10, 11); the interactions between environmental factors and genetic variants in SLE remain unclear. Hypersensitivity to UV irradiation is frequently diagnosed in patients with SLE. Recently, deficiencies in the ability to repair DNA damage and the consequent abnormal levels of apoptotic bodies have been identified as causative factors for SLE. Clinical studies have further demonstrated that a number of SLE patients are highly deficient in DNA repair (12-14),suggesting that genetic variations in the genes encoding DNA repair proteins contribute to the susceptibility to SLE. Among the genes involved in the DNA 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59
repair pathways, the X-ray repair cross-complementing (XRCC) gene family can protect mammalian cells from DNA damage caused by ionizing radiation (15, 16). Consistent with this observation, previous studies have indicated that the XRCC 4, 5, 7, and 9 proteins function at DNA break sites to trigger the development of autoimmune diseases (17-20). A prior study indicated that XRCC1 is the genetic factor that interacts with environmental factors, such as UV irradiation, to promote the pathogenesis of SLE in the HC-TW population (21). DNA polymorphisms in the XRCC1 and XRCC3 have been an area of
intense current investigation (22).
XRCC3 is an X-ray repair cross-complementing gene family that can protect mammalian cells from DNA damage induced by ionizing radiation (15, 16).Cells lacking XRCC3 are unable to form Rad51 foci after radiation damage, and demonstrate genetic instability and increased sensitivity to DNA damaging agents (23, 24). The SNP within XRCC3 have been identified at codon 241, which is located in exons 7 (25). Molecular epidemiological studies have linked XRCC3 polymorphism to increased risk of breast cancer (26), lung cancer (27), skin cancer (28), and colorectal cancer (29). The results have been inconsistent (30). The SNP at codon 241 is also linked to reduced DNA repair activity (31). Typically, the allele frequencies of genes often differ substantially between populations, and thus, ethnic-specific association studies are required to confirm genetic association in different populations. In this present
60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78
study, we investigated whether the XRCC3 rs861539 SNP contributes to SLE susceptibility in HC-TW populations and confirm genetic association in different populations using a
meta-analysis approach.
Materials and Methods
Patients and sample collection
A total of 163 SLE patients and 191 healthy controls were recruited from the China Medical University Hospital in Taiwan. All SLE patients met the criteria for SLE as defined by the American Rheumatism Association. People who underwent health examinations at the same hospital were recruited for the healthy control group. Informed consent was obtained from all participants, and was approved by the Institutional Review Board (IRB) at the China Medical University.
Genomic DNA extraction and genotyping
All blood samples were collected using venipuncture for genomic DNA isolation. Genomic DNA was extracted from peripheral blood leukocytes according to standard protocols (Genomic DNA kit; Qiagen, Valencia, CA, USA). We used the following primers: Forward:
5'-GCTCGCCTGGTGGTCATC-3' and reverse: 5'-CTTCCGCATCCTGGCTAAAAA-5'-GCTCGCCTGGTGGTCATC-3'. The PCR products were amplified with an initial denaturation at 95°C for 5 min, followed by 40 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98
cycles of 95°C for 10 s, 56°C for 10 s, 72°C for 20 s, and one additional cycle of 72°C for 5 min. DNA fragments containing the polymorphism were detected using PCR-RFLP, and CviA II digestion was used to detect codon 722 C/T.
Identification of eligible studies and data extraction
A literature search was conducted for studies that examined associations between the
XRCC3 Thr241Met polymorphism and SLE. We utilized the MEDLINE citation index to
identify articles in which the XRCC3 Thr241Met polymorphism was determined in SLE patients and controls (until December 2012). In addition, all references mentioned in identified articles were reviewed to identify additional studies not indexed by MEDLINE. The following information was extracted for each study: author, year of publication, ethnicity of the study population, demographics, numbers of cases and controls.
Meta-analysis
Population data were selected using a population data source (www.hapmap.org) for meta-analysis. The HapMap database was used to identify the allele and genotype frequencies of XRCC3 Thr241Met polymorphisms in various ethnic populations. The populations were as follows: Tunisian (TUN), Utah residents with Northern and Western European ancestry from the CEPH collection (CEU), Tuscan in Italy (TSI), Luhya in Webuye , Kenya (LWK), Maasai in Kinyawa, Kenya (MKK), Mexican ancestry in Los Angeles, California (MEX), 99 100 101 102 103 104 105 106 107 108 109 110 111 112 113 114 115 116 117
Gujarati Indians in Houston (GIH), Yoruban in Ibadan, Nigeria (YRI), African ancestry in Southwest USA (ASW), Japanese in Tokyo, Japan (JPT), Han Chinese in Beijing China (HCB), Chinese in Metropolitan Denver, Colorado, Texas (CHD), and Han Chinese in Taiwan (HC-TW).
Statistical analysis
The allelic and genotype frequency distributions for the polymorphism of SLE were determined through χ2 analysis using SPSS software (version 10.0, SPSS Inc. Chicago, Illinois, US). A P value of less than 0.05 was considered statistically significant. Allelic and genotype frequencies were expressed as percentages of the total number of alleles and genotypes. Odds ratios (OR) were calculated from allelic and genotype frequencies with a 95% confidence interval (95% CI). Haplotypes were determined using the Bayesian statistical method, which was available in the program Phase 2.1.32. Adherence to the
Hardy-Weinberg equilibrium constant was tested using a χ2 test with one degree of freedom. Pair-wise Chi square (χ2) tests were also performed between HC-TW and
HapMap populations using the allele frequencies in a 22 contingency table to determine whether the HC-TW population significantly differed from other populations.
Result
The SNP rs861539: C > T at exon 7 causes Threonine to Methionine substitution at codon 241 of XRCC3 protein (Thr241>Met). PCR-RFLP product for rs861539 showed in Fig.1. 118 119 120 121 122 123 124 125 126 127 128 129 130 131 132 133 134 135 136
Table I shows the genotype distribution values of XRCC3 Thr241Met polymorphism in SLE patients and the healthy control. No statistically significant difference was observed in the genotype frequency of XRCC3 Thr241Met polymorphisms between the SLE patients and the healthy control. However, a statistically significant difference was observed in allele frequency of XRCC3 Thr241Met polymorphism between the SLE patients and the healthy control (P<0.05).
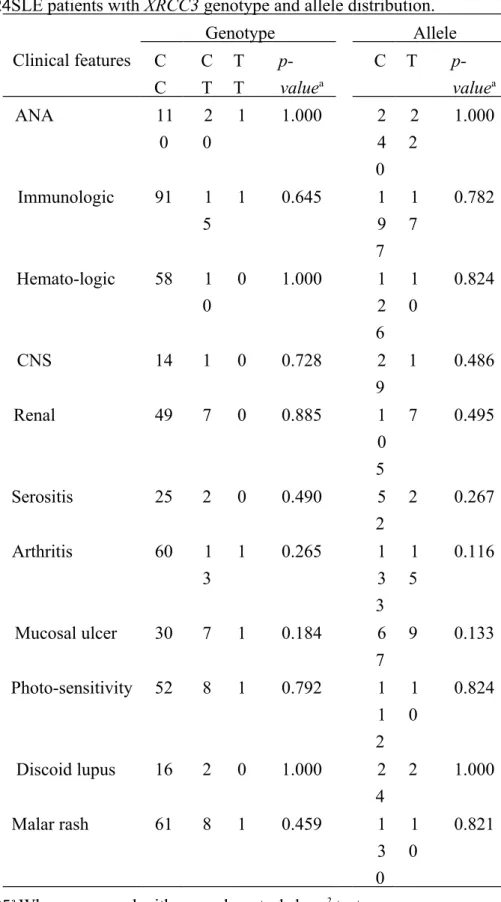
Furthermore, we determined the association among XRCC3 SNP and clinical and biochemical manifestations in SLE patients. The results showed that no significant difference occurred between the clinical and biochemical manifestations of SLE patients with XRCC3 genotype and allele distribution (Table 2).
We compared the XRCC3 Thr241Met polymorphism of healthy females and males in the HC-TW population, and the results indicated that no significant difference occurred between females and males in this population (Table 3). The XRCC3 Thr241Met T/T
genotype was not observed in the healthy HC-TW population.
We compared the XRCC3 Thr241Met between the healthy HC-TW population and the healthy HapMap population. The meta-analysis results, based on 13 studies, indicated that the genotype of XRCC3 Thr241Met polymorphism differed significantly between the HC-TW population and HapMap populations worldwide (Table 4). The Thr/Thr (CC), Thr/Met (CT), and Met/Met (TT) genotype frequencies in the HC-TW population were 137 138 139 140 141 142 143 144 145 146 147 148 149 150 151 152 153 154 155
0.922, 0.078, and 0, respectively (Table 4). The wild-type (C) and variant (T) alleles frequencies were 0.961 and 0.039, respectively. For HapMap populations, the variant allele frequency was 0.041 among the CHD population, and 0.474 among the TUN population (Table 4). The HC-TW population differed significantly from all HapMap populations
(P<0.01), except for Chinese populations, such as the HCB and CHD (P>0.05).
Discussion
UV irradiation is a powerful inducer of SLE, and genetic variation determines the susceptibility of a person to the development of SLE. We previously reported that SLE patients were associated with DNA repair gene XRCC1 polymorphism in the HC-TW population (21). Patients with SLE are commonly observed in RA patients. Previously report found that genetic polymorphisms of the DNA repair gene uracil-DNA glycosylase (UNG), 8-oxoguanine glycosylase 1 (OGG1), and N-methylpurine-DNA glycosylase (MPG) are associated with the susceptibility of RA in Taiwan population (32-34). We attempted to determine whether the functional SNP of XRCC3 Thr241Met is the genetic factor that reduces protection from DNA damage induced by UV exposure in SLE patients. The genetic polymorphism of DNA repair gene XRCC3 was significantly associated with SLE in the HC-TW population. The results suggest that the HC-TW population with genotype T/C at codon 241 of XRCC3 have a higher risk of developing SLE. No 156 157 158 159 160 161 162 163 164 165 166 167 168 169 170 171 172 173 174
association between XRCC1 Arg399Gln, XRCC3 Thr241Met polymorphism and SLE patients was observed in Brazilian populations (35). XRCC3 is one of the DNA repair genes, and encodes for a protein participating in the homologous recombination repair (HRR) of DNA double-strand breaks and cross-links (31, 36, 37). It is a member of an emerging family of Rad-51-related proteins that may participate in HRR to maintain chromosome stability and repair DNA damage (24). These phenotypes are a result of failure to initiate HRR and aberrant processing of HRR intermediates (31). Our results indicated that XRCC3 Thr241Met SNP associated with SLE disease in the HC-TW population and the distribution profile of the SNP is significant different compared with other non-Han-Chinese populations.
The variant T allele frequencies were 0.039 in HC-TW, 0.041 in CHD, and 0.062 in HCB populations. However, the variant T allele frequencies were 0.116 in JPT, 0.429 in CEU, 0.461 in TSI, and 0.474 in TUN populations. The results of meta-analysis, based on 13 studies, indicated that the genotype and allele frequencies of the XRCC3 Thr241Met polymorphism differed significantly between the HC-TW population and HapMap populations, except for the Han Chinese populations. The XRCC3 systematic function remains unclear. Walker Boxes A and B are two potential ATP-binding domains (38), and suggest that the transition from hydrophilic Threonine to a hydrophobic Methionine can affect ATP-binding and DNA repair efficiency (31, 39). A recent study indicated that the 175 176 177 178 179 180 181 182 183 184 185 186 187 188 189 190 191 192 193
variant allele of TUN differed significantly from all HapMap populations, except the CEU and TSI populations (38). We found that the variant allele of the HC-TW population differed significantly from all HapMap non-Han-Chinese populations. These results indicate that XRCC3 Thr241Met SNP differs among populations worldwide.
This study identified the polymorphism in the DNA repair gene XRCC3 Thr241Met and its associations with SLE risk in the HC-TW population. The meta-analysis of XRCC3 Thr241Met polymorphism showed a significant difference between the HC-TW population and the HapMap populations.
Acknowledgements
This work is supported in part by Taiwan Department of Health Clinical Trial and Research Center of Excellence (DOH101-TD-B-111-004), China Medical University (CMU100-S-06) and China Medical University Hospital (DMR-101-041) in Taiwan. The first author fellowship (YT Chen) is supported by China Medical University (CMU101-AWARD-01), Taiwan. 194 195 196 197 198 199 200 201 202 203 204 205 206 207 208
References
1. Yasutomo K. Pathological lymphocyte activation by defective clearance of self-ligands in systemic lupus erythematosus. Rheumatology (Oxford). 2003;42:214-222.
2. Mortensen ES, Fenton KA, Rekvig OP. Lupus nephritis: The central role of nucleosomes revealed. Am J Pathol 2008;172:275-283.
3. Parks CG, Cooper GS, Hudson LL, et al. Association of Epstein-Barr virus with systemic lupus erythematosus: Effect modification by race, age, and cytotoxic T
lymphocyte- associated antigen 4 genotype. Arthritis Rheum 2005;52:1148-1159. 4. Doria A, Canova M, Tonon M, et al. Infections as triggers and complications of systemic
lupus erythematosus. Autoimmun Rev 2008;8:24-28.
5. Rosenstein BS, Rosenstein RB, Zamansky GB. Repair of DNA damage induced in systemic lupus erythematosus skin fibroblasts by simulated sunlight. J. Invest. Dermatol 1992;98:469-474.
6. Mok CC, Lau CS. Pathogenesis of systemic lupus erythematosus. J Clin Pathol 2003;56: 481-490.
7. Lin YJ, Wan L, Lee CC, et al. Disease association of the interleukin-18 promoter polymorphisms in Taiwan Chinese systemic lupus erythematosus patients. Genes Immun 2007;8:302-307.
8. Lin YJ, Wan L, Sheu JJ, et al. A/C polymorphism in the interleukin-18 coding region among Taiwanese systemic lupus erythematosus patients. Lupus 2008;17:124-127. 209 210 211 212 213 214 215 216 217 218 219 220 221 222 223 224 225 226 227 228
9. Lin YJ, Wan L, Sheu JJ, et al. G/T polymorphism in the interleukin-2 exon 1 region among Han Chinese systemic lupus erythematosus patients in Taiwan. Clin Immunol 2008;129: 36-39.
10. Shirai T, Nishimura H, Jiang Y, Hirose S. Genome screening for susceptibility loci in systemic lupus erythematosus. Am J Pharmacogenomics 2002;2:1-12.
11. Cervino AC, Tsinoremas NF, Hoffman RW. A genome-wide study of lupus: Preliminary analysis and data release. Ann N Y Acad Sci 2007;1110:131-139.
12. Beighlie DJ, Teplitz RL. Repair of UV damaged DNA in systemic lupus erythematosus. J Rheumatol 1975;2:149-160.
13. Herrick AL, Rafferty JA, Margison GP. DNA repair deficiency in systemic lupus erythematosus; Cause or consequence of disease and implications for management.
Lupus 1995;4:423-424.
14. McCurdy D, Tai LQ, Frias S, Wang Z. Delayed repair of DNA damage by ionizing radiation in cells from patients with juvenile systemic lupus erythematosus and rheumatoid arthritis. Radiat Res 1997;147:48-54.
15. Thacker J, Zdzienicka MZ. The mammalian XRCC genes: Their roles in DNA repair and genetic stability. DNA Repair (Amst) 2003;2:655-672.
16. Thacker J, Zdzienicka MZ. The XRCC genes: expanding roles in DNA double-strand break repair. DNA Repair (Amst) 2004;3:1081-1090.
17. Takeda Y, Dynan WS. Autoantibodies against DNA double-strand break repair 229 230 231 232 233 234 235 236 237 238 239 240 241 242 243 244 245 246 247 248
proteins. Front Biosci 2001;6:D1412-D1422.
18. Lee KJ, Dong X, Wang J, et al. Identification of human autoantibodies to the DNA ligase IV/XRCC4 complex and mapping of an autoimmune epitope to a potential regulatory region. J Immunol 2002;169:3413-3421.
19. Kamei N, Yamane K, Yamashita Y, et al. Anti-Ku antibody-positive scleroderma-dermatomyositis overlap syndrome developing Graves’ disease and immune
thrombocytopenic purpura. Intern Med 2002;41:1199-1203.
20. Tajouri L, Mellick AS, Ashton KJ, et al. Quantitative and qualitative changes in gene expression patterns characterize the activity of plaques in multiple sclerosis. Brain Res Mol Brain Res 2003;119:170-183.
21. Lin YJ, Wan L, Huang CM, et al. Polymorphisms in the DNA repair gene XRCC1 and associations with systemic lupus erythematosus risk in the Taiwanese Han Chinese population. Lupus 2009;18:1246-1251.
22.
Goode EL, Ulrich CM, Potter JD. Polymorphisms in DNA repair genes and associations with cancer risk. Cancer Epidemiol Biomarkers Prev 2002;11(12):1513- 1530.
23. Griffin CS. Aneuploidy, centrosome activity and chromosome instability in cells deficient in homologous recombination repair. Mutat Res 2002;504:149-155.
24. Suwaki N, Klare K, Tarsounas M. RAD51 paralogs: roles in DNA damage signalling, recombinational repair and tumorigenesis. Semin. Cell Dev Biol 2011;22:898-905. 249 250 251 252 253 254 255 256 257 258 259 260 261 262 263 264 264 265 266 267
25. Matullo G, Palli D, Peluso M, et al. XRCC1, XRCC3, XPD gene polymorphisms, smoking and (32)P-DNA adducts in a sample of healthy subjects. Carcinogenesis
2001;22(9):1437-1445.
26. Kuschel B, Auranen A, McBride S, et al. Variants in DNA double-strand break repair genes and breast cancer susceptibility. Hum Mol Genet 2002;11(12):1399-1407. 27. Jacobsen NR, Raaschou-Nielsen O, Nexo B, et al. XRCC3 polymorphisms and risk of
lung cancer. Cancer Lett 2004;213(1):67-72.
28. Winsey SL, Haldar NA, Marsh HP, et al. A variant within the DNA repair gene XRCC3 is associated with the development of melanoma skin cancer. Cancer Res
2000;60(20): 5612-5616.
29. Mort R, Mo L, McEwan C, Melton DW. Lack of involvement of nucleotide excision repair gene polymorphisms in colorectal cancer. Br J Cancer 2003;89(2):333-337. 30. Jacobsen NR, Nexo BA, Olsen A, et al. No association between the DNA repair gene
XRCC3 T241M polymorphism and risk of skin cancer and breast cancer. Cancer Epidemiol Biomarkers Prev 2003;12(6):584-585.
31. Liu N, Lamerdin JE, Tebbs RS, et al. XRCC2 and XRCC3, new human Rad51-family members, promote chromosome stability and protect against DNA cross-links and
other damages. Mol Cell 1998;1:783-793.
32. Lo SF, Wan L, Huang CM, et al. Genetic polymorphisms of the DNA repair gene UNG are associated with the susceptibility of rheumatoid arthritis. Rheumatol Int 268 269 270 271 272 273 274 275 276 277 278 279 280 281 282 283 284 285 286 287
2012;32(12): 3723-3727.
33. Chen SY, Wan L, Huang CM, et al. Genetic polymorphisms of the DNA repair gene MPG may be associated with susceptibility to rheumatoid arthritis. J Appl Genet
2010;51(4):519-521.
34. Chen SY, Wan L, Huang CM, et al. Association of the C-285T and A5954G polymorphisms in the DNA repair gene OGG1 with the susceptibility of rheumatoid arthritis. Rheumatol Int 2012;32(5):1165-1169.
35. Bassi CL, Xavier DJ, Palomino GM, et al. Efficiency of the DNA repair and polymorphisms of the XRCC1, XRCC3 and XRCC4 DNA repair genes in systemic lupus erythematosus. Lupus 2008;17:988-995.
36. Pierce AJ, Johnson RD, Thompson LH, Jasin M. XRCC3 promotes homology- directed repair of DNA damage in mammalian cells. Genes Dev 1999;13:2633-2638.
37. Brenneman MA, Weiss AE, Nickoloff JA, Chen DJ. XRCC3 is required for efficient repair of chromosome breaks by homologous recombination. Mutat Res 2000;459:
89-97.
38. Walker JE, Saraste M, Runswick MJ, Gay N. Distantly related sequences in the a and b subunits of ATP synthase, myosin, kinases and other ATP-requiring enzymes and in common nucleotide binding fold. EMBO J 1982;1:945-951.
39. Salah GB, Fendri-Kriaa N, Kamoun H, et al. An interethnic variability and a functional prediction of DNA repair gene polymorphisms: the example of XRCC3 288 289 290 291 292 293 294 295 296 297 298 299 300 301 302 303 304 305 306 307
(p.Thr241>Met) and XPD (p.Lys751>Gln) in a healthy Tunisian population. Mol Biol Rep 2012;39: 9639-9647.
308 309
1 2 3
(CC) (CT) (TT)
Figure.1. Electrophoresis of PCR-RFLP product for rs861539. The sizes of PCR amplified products was 214 base pair (bp), after CviA II digestion, line 1 is CC genotype (214 bp). Line 2 was CT genotype, the fragments were 214 bp, 112 bp, and 102 bp. Line 3 was TT genotype, fragments were 112 bp and 102 bp.
310 311 312 313 314 315 316 317
Table 1. Allelic and genotype frequencies of XRCC3 polymorphisms among the Han Chinese SLE patients in Taiwan and the healthy control group.
Polymorphism s SLE N (%) Control N (%) p-valuea OR (95% CI) N=163 N=191 XRCC3 241 T T 1 (0.6) 0 (0) 0.06 -(rs861539) T C 24 (14.7) 15 (7.9) 2.04 (1.03-4.04) C C 138 (84.7) 176 (92.1) 1 T 26 (8.0) 15 (3.9) 0.02* 2.12 (1.10-4.08) C 300 (92.0) 367 (96.1) 1
aGenotype and allelic frequencies were compared between SLE patients and normal controls by x2 tests.
*Indicates statistical significance. 318
319
320 321 322
Table 2. Comparison of clinical and biochemical manifestations of SLE patients with XRCC3 genotype and allele distribution.
Clinical features Genotype Allele C C C T T T p-valuea C T p-valuea ANA 11 0 2 0 1 1.000 2 4 0 2 2 1.000 Immunologic 91 1 5 1 0.645 1 9 7 1 7 0.782 Hemato-logic 58 1 0 0 1.000 1 2 6 1 0 0.824 CNS 14 1 0 0.728 2 9 1 0.486 Renal 49 7 0 0.885 1 0 5 7 0.495 Serositis 25 2 0 0.490 5 2 2 0.267 Arthritis 60 1 3 1 0.265 1 3 3 1 5 0.116 Mucosal ulcer 30 7 1 0.184 6 7 9 0.133 Photo-sensitivity 52 8 1 0.792 1 1 2 1 0 0.824 Discoid lupus 16 2 0 1.000 2 4 2 1.000 Malar rash 61 8 1 0.459 1 3 0 1 0 0.821
a When compared with normal controls by x2 tests.
Abbreviations: ANA, antinuclear antibody; CNS, central nervous system; SLE, systemic lupus erythematosus.
323 324
325 326 327
Table 3. Genotype distributions and allele frequencies of XRCC3 SNP among the HC-TW population.
Genotype Total (N=191) Woman (N=157) Man (N=34) Allele frequency HWE
p-valuea
N Freq N Freq N Freq Wild-type Variant
rs861539: C > T (p.Thr241 > Met)
C/C 176 0.922 143 0.911 33 0.971
C/T 15 0.078 14 0.089 1 0.029 (C)0.961 (T)0.039 0.571
T/T 0 0 0 0 0 0
a When compared with normal controls by x2 tests. 328
Table 4. Genotype and allele frequencies of XRCC3 p.Thr241>Met in HapMap and HC-TW population.
Genotype frequency Allele frequency Pair-wise p-value between HC-TW and other
populations Population
Geno. Freq. Geno. Freq. Geno . Freq.
Wild-type Variant allele allele
XRCC3 C > T (p.Thr241 > Met) (C) (T) genotype allele
TUN (n = 154) C/C 0.312 C/T 0.428 T/T 0.260 0.526 0.474 9.19E-32 4.45E-41 CEU (n = 226) C/C 0.310 C/T 0.522 T/T 0.168 0.571 0.429 7.94E-36 2.45E-38 TSI (n = 204) C/C 0.324 C/T 0.431 T/T 0.245 0.539 0.461 1.28E-33 7.98E-42 LWK (n = 220) C/C 0.582 C/T 0.364 T/T 0.055 0.764 0.236 2.94E-14 1.15E-15 MKK (n = 310) C/C 0.645 C/T 0.310 T/T 0.045 0.800 0.200 2.13E-11 8.73E-13 MEX (n = 114) C/C 0.719 C/T 0.246 T/T 0.035 0.842 0.158 5.32E-06 3.04E-07 GIH (n = 202) C/C 0.574 C/T 0.356 T/T 0.069 0.752 0.248 1.70E-14 1.49E-16 YRI (n = 288) C/C 0.660 C/T 0.319 T/T 0.021 0.819 0.181 2.64E-10 8.47E-11 ASW (n = 114) C/C 0.684 C/T 0.281 T/T 0.035 0.825 0.175 2.43E-07 1.34E-08 JPT (n = 224) C/C 0.777 C/T 0.214 T/T 0.009 0.884 0.116 2.00E-04 5.15E-05
HCB (n = 274) C/C 0.876 C/T 0.124 T/T 0.000 0.938 0.062 0.116 0.126
CHD (n = 218) C/C 0.917 C/T 0.083 T/T 0.000 0.959 0.041 0.881 0.884
HC-TW (n =
191) C/C 0.922 C/T 0.078 T/T 0.000 0.961 0.039
Population data source: (www.hapmap.org) n number of individuals
TUN Tunisian, CEU Utah residents with Northern and Western European ancestry from the CEPH collection, TSI Tuscan in Italy, LWK Luhya in Webuye, Kenya, MKK Maasai in Kinyawa, Kenya, MEX Mexican ancestry in Los Angeles, California, GIH Gujarati Indians in Houston, YRI Yoruban in Ibadan, Nigeria, ASW African ancestry in Southwest USA, JPT Japanese in Tokyo, 330
331 332 333 334
Japan, HCB Han Chinese in Beijing China, CHD Chinese in Metropolitan Denver, Colorado, Texas, HC-TW Han Chinese in Taiwan.
p < 0.05, a significant difference between HC-TW and other populations p > 0.05, no significant difference between HC-TW and other populations
336 337 338 339