國立交通大學
資訊工程系
博士論文
複雜網路中具縮影性質之階層:
分析及應用
The Abstraction Hierarchy in Complex Networks:
Analyses and Applications
博士生: 鄭家胤
指導教授: 孫春在 教授
國立交通大學
資訊工程系
博士論文
複雜網路中具縮影性質之階層:
分析及應用
The Abstraction Hierarchy in Complex Networks:
Analyses and Applications
博士生: 鄭家胤
指導教授: 孫春在 教授
複雜網路中具縮影性質之階層:
分析及應用
The Abstraction Hierarchy in Complex Networks:
Analyses and Applications
研究生 :鄭家胤 Student:Chia-Ying Cheng
指導教授:孫春在 Advisor:Chuen-Tsai Sun
國立交通大學
資訊工程系
博士論文
A Dissertation Submitted toDepartment of Computer Science College of Computer Science National Chiao Tung University
for the Degree of Doctor of Philosophy
in
Computer Science
July 2009
Hsinchu, Taiwan, Republic of China
i
Abstract (in Chinese)
以複雜網路的型態來呈現複雜系統中的互動關係是一種方便且行之有年的研究 方法,包括在生物學、生態學、社會學等等的領域上,除了讓研究者有不同於以 往該領域傳統議題的新觀點外,許多新的方法也因此被提出來以解決在各種不同 複雜系統上的問題。其中,最重要也最具挑戰性的一個問題就是,如何將複雜網 路做分群(在社會學領域被稱之為共同體(community)或是群組(group),在生物學 上被稱為基塊(motif)或是模組(module))。如何(1)找出模組,(2)階層性的組織, 及(3)這兩者對應到真實世界的關係,一直是研究者的焦點所在。儘管已經有一 些成功的研究,但是至今仍沒有一個標準的衡量方法可以來找出模組或是階層性 組織。以階層式組織來說,大多數的研究專注於其模組在不同階層上垂直面向的 關係之探討-其可用來表示"包含(inclusion)","因果(causality)"和"調控(regulation)" 關係;但往往忽略了其在同一階層上水平面向的關係之研究-其可用來提供給研 究者在某一階層上的網路的縮影(abstraction)或是骨架(backbone)。在本論文研究 中,我提出了一雙向式尋找模組及建構階層組織的方法,其同時考慮了各個模組 間垂直和水平的關係來建構出該複雜系統的金字塔階層(pyramid hierarchies), 此方法除了被人工網路驗證外,也被應用在生物及社會網路上,其結果顯示該方 法在擷取複雜系統之資訊上卓越的效能。
ii
Abstract (in English)
The use of nodes and links to assemble networks is convenient for representing
interactions in complex systems. This benefits researchers in biology, ecology,
sociology and other biological and social sciences. In addition to supporting
alternative views of complex domains, network research is also supporting new
methods for solving problems in a range of domains. One particularly important and
challenging problem is partitioning networks into clusters (called communities or
groups in social science research and motifs or modules in biology). Research in these
areas has focused on identifying modules and hierarchical organizations that
correspond to real-world meanings (e.g., biological functions or economic and
political constraints). Despite a number of successful examples, no uniform measure
of modularity or standard hierarchical structure exists. Most current descriptions of
hierarchical organizations are limited to vertical relationships between modules at
different hierarchical levels, thus overlooking horizontal relationships that express
associations among modules at the same level. Vertical relationships can be used to
represent inclusion hierarchies and to describe causality/regulation. Horizontal
relationships complement these by providing abstractions of original networks of
iii
In this dissertation I describe a proposal for a two-way simultaneous module-finding
and hierarchy-building strategy. I take both vertical and horizontal relationships
between modules into consideration when building pyramid hierarchies in which each
layer represents an abstraction of lower-level networks. This dissertation also contains
descriptions of tests for this proposed approach, using networks consisting of
anywhere from tens to hundreds of nodes and links, and in domains that include
artificial random networks, social networks, and biological networks. The results
iv
Acknowledgements
I would like to express my deep gratitude to my advisor, Professor Chuen-Tsai Sun,
for teaching me how to define a problem, how to determine its essential components,
and how to identify relationships among those components. I am especially grateful
for the way he modeled the right attitude for students to take when addressing new
topics. That is a skill I will use both in my research and in everyday life.
I also thank my co-advisor, Professor Yuh-Jyh Hu, for his strong support during the
last two years of my Ph.D. research and writing. As a mentor he discussed in great
detail new ideas and ways to implement them. As a friend he shared his life and work
values with me, which helped me stay centered during the entire process.
For all others who were patient with me as I completed this project, I extend my
thanks for your support and patience.
Finally, I give thanks to my Lord Jesus Christ: "And we know that all things work
together for good to those who love God, to those who are called according to His
purpose" (Romans 8:28).
v
Contents
ABSTRACT (IN CHINESE) ... I
ABSTRACT (IN ENGLISH) ... II
ACKNOWLEDGEMENTS ... IV
CONTENTS ... V
LIST OF TABLES ... IX
LIST OF FIGURES ... XI
CHAPTER 1 INTRODUCTION ... 1
1.1 COMPLEX NETWORK TOPOLOGY ... 3
1.1.1 Randomness ... 4
1.1.2 Small-world property ... 5
1.1.3 Scale-free distributions ... 7
1.2 COMPLEX NETWORK STRUCTURE ... 8
1.2.1 Motifs ... 8
1.2.2 Communities ... 11
1.2.3 Hierarchical modularity ... 11
vi
1.3.1 Cellular automata ... 13
1.3.2 Preferential attachment ... 14
CHAPTER 2 STATIC NETWORKS AND DYNAMIC PROCESS CHARACTERIZATION AND ANALYSIS ... 16
2.1 NETWORK MOTIF DETECTION ... 18
2.1.1 General: Bridge and Brick Network Motif-Detecting Algorithm ... 21
2.1.2 Specific: Bridge and Brick Network Motif-Detecting Algorithm ... 26
2.2 SOCIAL NETWORK SIMULATION ... 27
2.3 EPIDEMIC DYNAMICS ANALYSIS ... 29
2.4 ABSTRACTION HIERARCHY ... 33
2.4.1 Proximity Measure ... 34
2.4.2 Network Abstraction ... 43
CHAPTER 3 NETWORK MOTIF EXPERIMENTS ... 45
3.1 GENERAL:BRIDGE AND BRICK NETWORK MOTIF-DETECTING ALGORITHMS ... 45
3.1.1 Validation ... 49
3.1.2 Experiments ... 52
vii
3.2 SPECIFIC:BRIDGE AND BRICK NETWORK MOTIF-DETECTING ALGORITHMS ... 63
3.2.1 Validation ... 63
3.2.2 Experiments ... 67
3.2.3 Conclusion ... 76
CHAPTER 4 SOCIAL NETWORK SIMULATION EXPERIMENTS ... 77
4.1 FRIENDSHIP EVOLUTION AND THE THREE-RULE MODEL ... 77
4.1.1 Friendship Selection Methods ... 79
4.1.2 Friendship Update Equation ... 80
4.1.3 Fitting a Normal Distribution ... 81
4.2 EXPERIMENT ... 83
4.2.1 Effects of Leaving and Arriving ... 85
4.2.2 Effects of Breakup Threshold ... 88
4.2.3 Effects of Resources ... 89
4.2.4 Effects of Initial Friendship ... 90
4.2.5 Distribution of Co-Directors... 92
4.2.6 Sampling ... 94
viii
CHAPTER 5 EPIDEMIC DYNAMICS EXPERIMENTS ... 97
5.1 EPIDEMIC DYNAMICS IN COMPLEX NETWORKS ... 98
5.2 EXPERIMENTS ... 102
5.3 CONCLUSION ... 111
CHAPTER 6 ABSTRACTION HIERARCHY EXPERIMENTS ... 112
6.1 BACKGROUND ... 113
6.2 VALIDATION ... 118
6.3 EXPERIMENTS ... 121
6.3.1 Club network analysis... 122
6.3.2 Football network analysis ... 125
6.3.3 PPI network analysis ... 127
6.3.4 Metabolic network analysis ... 134
CHAPTER 7 CONCLUSION ... 140
ix
List of Tables
TABLE.1.AN UPDATE RULE FOR A ONE-DIMENSIONAL, TWO-STATE CELLULAR AUTOMATON. ... 14
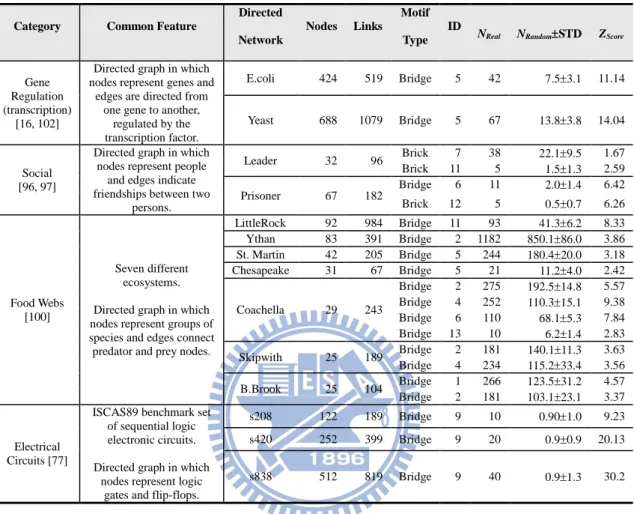
TABLE 2.BRIDGE AND BRICK SUBGRAPH FREQUENCIES IN FOUR COMPLEX NETWORK CATEGORIES (FOR VALIDATION PURPOSES). ... 51
TABLE 3.BRICK AND BRIDGE MOTIFS IN FOURTEEN REAL WORLD NETWORKS, INCLUDING EDGE AND NODE DEFINITIONS, NETWORK SIZES, AND REFERENCES. ... 60
TABLE 4.DESCRIPTIONS OF FIVE GENE REGULATION NETWORKS: EDGE AND NODE DEFINITIONS, NETWORK SIZES, AND REFERENCES. ... 70
TABLE 5.BRICK AND BRIDGE MOTIFS IN FIVE GENE REGULATION NETWORKS... 72
TABLE 6.TERMS AND ABBREVIATIONS FOR INITIALIZED PARAMETERS ... 84
TABLE 7.TERMS AND ABBREVIATIONS FOR STATISTICS ... 85
TABLE 8.EFFECTIVE DIRECTIONS OF THE PARAMETERS ON <K>,C,L ... 92
TABLE 9.CORRELATIONS BETWEEN <K>,C,L FROM EXPERIMENTS ... 92
TABLE 10.SUMMARY OF BIOLOGICAL SIGNIFICANCE OF MODULES BASED ON GO BIOLOGICAL PROCESS ANNOTATIONS ... 131
TABLE 11.SUMMARY OF WITHIN-MODULE CONSISTENCY OF METABOLIC PATHWAY CLASSIFICATION BASED ON KEGG. ... 137
xi
List of Figures
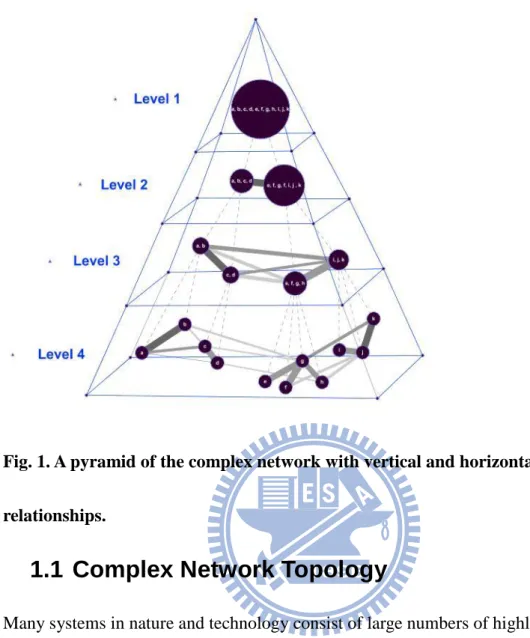
FIG.1.A PYRAMID OF THE COMPLEX NETWORK WITH VERTICAL AND HORIZONTAL RELATIONSHIPS. ... 3
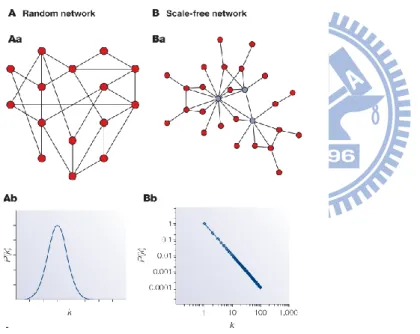
FIG.2.THE COMPARISON BETWEEN THE RANDOM NETWORK AND THE SCALE-FREE NETWORK. ... 8
FIG.3.13 POSSIBLE OF TRIAD MOTIFS DEFINED BY ALON. ... 9
FIG.4.COMMUNITIES CAN BE DEFINED AS GROUPS OF NODES SUCH THAT THERE IS A HIGHER DENSITY OF EDGES WITHIN GROUPS THAN BETWEEN THEM. ... 11
FIG.5.THE HIERARCHICAL NETWORK AND ITS DEGREE DISTRIBUTION. ... 13
FIG.6.NETWORK MOTIFS EXAMPLE. ... 20
FIG.7.LINK-WEIGHTED VALUE CALCULATING EXAMPLE.THE LINK-WEIGHTED VALUE WEIGHT (A, B) OF EDGE (A, B) IS 0 WHILE WEIGHT (B, C) ... 21
FIG.8. THE SMALL-WORLD MODEL.BLACK SIGNIFIES STRONG LINKS AND RED WEAK LINKS. ... 27
FIG.9.THREE-RULE MODEL FLOW DIAGRAM. ... 29
FIG.10FLOWCHART FOR A SIS EPIDEMIOLOGICAL SIMULATION MODEL. ... 33
FIG 11.FOUR SIMPLE NETWORKS TO ILLUSTRATE PROXIMITY MEASURES. ... 39
FIG 12. A SIMPLE UNDIRECTED WEIGHTED NETWORK. ... 41
FIG.13.PERCENTAGES OF BRIDGE AND BRICK MOTIFS IN SMALL-WORLD NETWORKS ACCORDING TO DIFFERENT REWIRING RATIOS. ... 52
xii
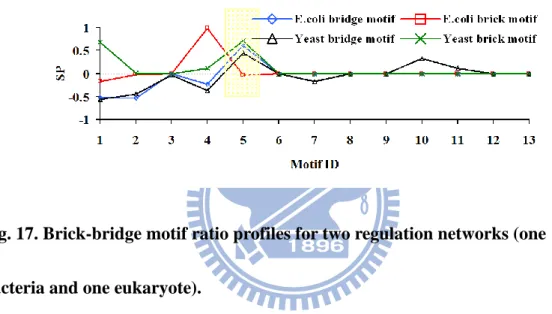
FIG.15.BRIDGE MOTIF RATIO PROFILES FOR TWO SOCIAL NETWORKS. ... 56 FIG.16.BRICK MOTIF RATIO PROFILES FOR TWO SOCIAL NETWORKS. ... 56 FIG.17.BRICK-BRIDGE MOTIF RATIO PROFILES FOR TWO REGULATION NETWORKS (ONE BACTERIA AND
ONE EUKARYOTE). ... 57 FIG.18.BRIDGE MOTIF RATIO PROFILES FOR SEVEN FOOD WEBS... 58 FIG.19. RELATIONSHIPS BETWEEN CLUSTERING COEFFICIENTS AND DIFFERENT REMOVAL RATIOS FOR
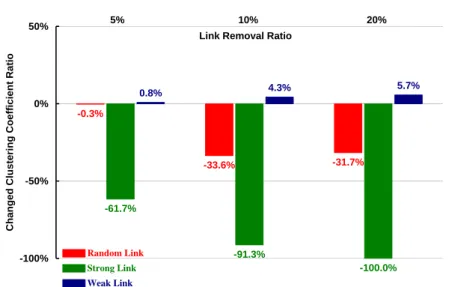
THREE E. COLI LINK TYPES.RED, RANDOM; GREEN, STRONG; BLUE, WEAK. ... 65 FIG.20. RELATIONSHIPS BETWEEN CLUSTERING COEFFICIENTS AND DIFFERENT REMOVAL RATIOS FOR
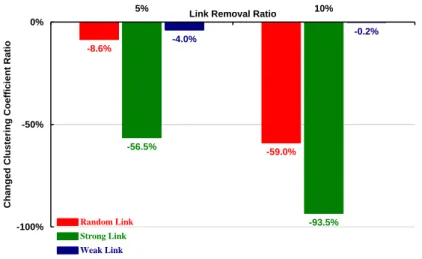
THREE S. CEREVISIAE (YEAST) LINK TYPES.RED, RANDOM; GREEN, STRONG; BLUE, WEAK. ... 66 FIG.21. COMPARISON OF ORIGINAL (BLUE CURVE) AND ALTERED (RED CURVE) BRICK MOTIF RATIO
PROFILES FOR E. COLI AFTER RANDOMLY REMOVING 40% OF ITS LINKS.ALTERED RESULTS
REPRESENT AVERAGE VALUES FOR 30 RUNS. ... 66 FIG.22.COMPARISON BETWEEN ORIGINAL BRICK MOTIF RATIO PROFILES AND ALTERED BRICK MOTIF
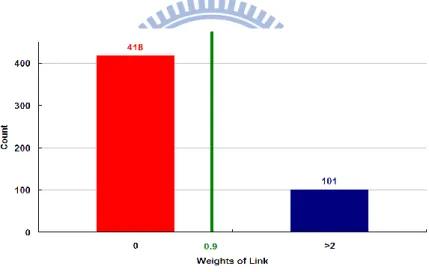
RATIO PROFILES FOR S. CEREVISIAE (YEAST) AFTER RANDOMLY REMOVING 40% OF ITS LINKS. ... 67 FIG.23.DISTRIBUTION OF LINK WEIGHTS IN E.COLI.AVERAGE MEAN AND STANDARD DEVIATION OF LINK
WEIGHTS FOR RANDOMIZED NETWORKS WERE CALCULATED AS 0.900.04. ... 67 FIG.24. COMPARISONS OF TRIAD SIGNIFICANCE PROFILES (TSPS) FOR OUR BRIDGE AND BRICK MOTIFS
xiii
FIG.25. COMPARISONS OF TRIAD SIGNIFICANCE PROFILES (TSPS) FOR OUR BRIDGE AND BRICK MOTIFS
AND MILO ET AL.‘S [7],[28]S. CEREVISIAE (YEAST) MOTIFS. ... 75
FIG.26. BRICK MOTIF RATIO PROFILES FOR TWO GENE REGULATION NETWORKS:E. COLI AND S. CEREVISIAE (YEAST). ... 75
FIG.27. BRIDGE MOTIF RATIO PROFILES FOR THREE GENE REGULATION NETWORKS:C. ELEGANS, SEA URCHIN, AND DROSOPHILA. ... 75
FIG.28.BETA14 PDF CURVES AT DIFFERENT AVERAGES OF 0.1,0.5, AND 0.9. ... 83
FIG.29.COMPARISON OF BETA AND NORMAL DISTRIBUTIONS. ... 83
FIG.30.EXAMPLE OF A STATISTICALLY STATIONARY STATE USING THE PROPOSED MODEL. ... 85
FIG.31.<K>,C AND L VARYING IN BREAKUP THRESHOLD Θ WITH DIFFERENT LEAVING AND ARRIVING PROBABILITY P. ... 87
FIG.32.<K>,C AND L VARYING IN FRIEND-REMEMBERING Q VALUE WITH DIFFERENT DISTRIBUTIONS OF FRIEND-MAKING RESOURCES. ... 88
FIG.33.<K>,C AND L VARYING IN FRIEND-REMEMBERING Q VALUE WITH DIFFERENT DISTRIBUTIONS OF INITIAL FRIENDSHIP F0. ... 89
FIG.34.TWO-RULE MODEL DEGREE DISTRIBUTION P(K). ... 90
FIG.35.<K>,C AND L VARYING IN LEAVING AND ARRIVING PROBABILITY P. ... 92
xiv
FIG.37PREVALENCE Ρ IN STEADY STATE AS A FUNCTION OF EFFECTIVE SPREADING RATE Λ. ... 101 FIG.38.RELATIONSHIP BETWEEN EFFECTIVE SPREADING RATE AND STEADY DENSITY OF THE SIS
EPIDEMIOLOGICAL MODEL ON THREE TYPES OF COMPLEX NETWORK PLATFORMS. ... 104 FIG.39.HOW THE AMOUNT OF AN INDIVIDUAL‘S ECONOMIC RESOURCES AFFECT STEADY DENSITY CURVES.
... 106
FIG.40.RELATIONSHIP BETWEEN RATIO OF TRANSMISSION COSTS TO AN INDIVIDUAL‘S ECONOMIC RESOURCES AND CRITICAL THRESHOLD... 106 FIG.41.HOW DIFFERENT DISTRIBUTION TYPES OF INDIVIDUAL ECONOMIC RESOURCES (DELTA, UNIFORM,
NORMAL, POWER-LAW) AFFECT STEADY DENSITY CURVES AND CRITICAL THRESHOLDS OF
INFECTIOUS DISEASE DIFFUSION IN A SCALE-FREE NETWORK. ... 108 FIG.42.A UNIFORM (N =5, R =2) AND NORMAL DISTRIBUTION (STANDARD DEVIATION =2) OF
INDIVIDUAL ECONOMIC RESOURCES WITH AVERAGE VALUE <R> OF 16. ... 109 FIG.43.INDIVIDUAL ECONOMIC RESOURCES IN A POWER-LAW DISTRIBUTION. ... 109 FIG.44.HOW DIFFERENT TYPES OF INDIVIDUAL ECONOMIC RESOURCE DISTRIBUTIONS (DELTA, UNIFORM,
NORMAL, AND POWER-LAW) AFFECT STEADY DENSITY CURVES AND CRITICAL THRESHOLDS OF INFECTIOUS DISEASE DIFFUSION IN A SCALE-FREE NETWORK. ... 110 FIG.45.A UNIFORM (N =5, R =3) AND NORMAL DISTRIBUTION (STANDARD DEVIATION =3) OF
xv
FIG.46.INDIVIDUAL ECONOMIC RESOURCES IN A POWER-LAW DISTRIBUTION. ... 111
FIG 47.EXAMPLE OF A DENDROGRAM FROM CONVENTIONAL HIERARCHICAL CLUSTERING. ... 118
FIG 48.VALIDATION OF TWO-WAY MODULE-FINDING-HIERARCHY-BUILDING STRATEGY. ... 120
FIG 49.ABSTRACT NETWORK CORRESPONDING TO HIERARCHICAL LEVEL THREE AND TWO. ... 121
FIG.50.CLUSTERING RESULTS OF ZACHARY‘S KARATE CLUB NETWORK. ... 124
FIG.51.THE ANALYSIS OF THE FOOTBALL NETWORK ... 127
FIG.52THE P-VALUE OF THE CORRESPONDING NODES AT DIFFERENT LEVELS. ... 132
FIG.53.THE MST OF PPI AT LEVEL FOUR AND FIVE, AND HAVE BLUE AND RED COLORS, RESPECTIVELY. ... 133
FIG.54.THE MAPPING BETWEEN THE MODULES WE FOUND AND THE REAL GO ID. ... 134
FIG.55.THE PYRAMID OF ABSTRACTION DISCLOSED FROM A METABOLIC NETWORK. ... 138
FIG.56.EXAMPLE OF THE VERTICAL RELATIONSHIPS IN AN ABSTRACTION PYRAMID DISCLOSED FROM A METABOLIC NETWORK. ... 138
FIG.57.EXAMPLE OF THE HORIZONTAL RELATIONSHIP AT THE THIRD LEVEL OF AN ABSTRACTION PYRAMID. ... 139
1
Chapter 1 Introduction
Complex networks consist of sets of items called vertices or nodes and connections
between them called edges. There are many examples of systems in the form of
networks (also called ―graphs‖ in mathematics): the World Wide Web, the Internet, social networks (acquaintances or other connections), distribution networks (e.g.,
blood vessels, postal delivery routes), organizations and business relations, neural
networks, metabolic networks, food webs, and research paper citations, among many
others.
The network concept is proving to be a very useful tool for studying complex systems
[1-3]. While no general theory of complexity exists [2, 4, 5], there is a growing
collection of related theories, paradigms, and tools, many of them associated with
physics and mathematics [5, 6]. They support explanations of complex phenomena
such as collective behavior observed in ferromagnetic phase transitions, herding
behavior, disease epidemics, and opinion formation—all examples of local
interactions that create global order [5, 7]. We also know that even very simple
systems such as discrete logistic growth models (logistic maps) can display rich and
2
systems manage to operate near criticality in the absence of fine-tuning [5, 8]. Fractal
geometry helps explain how and why certain forms and structures in nature arise—for
instance, vascular systems. As a new tool for studying complex phenomena, network
theory uses a mix of statistical mechanics, graph theory, and dynamical systems
theory [9-11].
The majority of network-related problems can be placed in one of two categories: for
static networks, relationships between network structure and function, and for
dynamic networks, global rules tied to network evolution [12-15]. In the following
3
Fig. 1. A pyramid of the complex network with vertical and horizontal relationships.
1.1 Complex Network Topology
Many systems in nature and technology consist of large numbers of highly
interconnected dynamical units [2, 16]. Examples include coupled biological and
chemical systems, neural networks, social interaction, and the Internet. An initial
approach to capturing the global properties of such systems is to model them as
graphs whose nodes represent dynamical units (e.g., neurons in the brain or
individuals in a social system) and whose links represent interactions between units.
4
between dynamical units (generally dependent on temporal, spatial, and many other
details) into simple binary numbers designating the existence or lack of links between
two corresponding nodes. Such approximations provide simple yet informative
representations of whole systems. The development of powerful and reliable data
analysis tools represent better mechanisms for exploring the topological properties of
multiple networked systems, thus supporting topological analyses of interactions in a
diverse range of systems (e.g., communication, social, and biological). These efforts
reveal that despite inherent differences, most real networks have the same topological
properties [1, 5]. The most significant are the small-world effect, degree scale-free
distributions, correlations, and clustering.
1.1.1 Randomness
The first non-regular network model [17, 18] was introduced by Paul Erdös and
Alfred Rényi in the late 1950s [19]. In this dissertation I will variously refer to this as
the random model, the Erdös-Rényi model, or the ER model. The ER model of a
random network starts with N nodes and connections between pairs of nodes at a p
probability, resulting in graphs with approximately N(N–1)/2 randomly placed links
5
indicating that most nodes have approximately the same number of links (close to the
average degree <k>). The tail (high k region) of the P(k) degree distribution decreases
exponentially, indicating the rarity of nodes that significantly deviate from the
average.
1.1.2 Small-world property
This property was first investigated in the 1960s in a social context, as part of a series
of experiments designed by Milgram [20, 21] to estimate the number of steps in
acquaintance chains. In his first experiment, Milgram asked randomly selected people in Nebraska to send letters that would eventually arrive at the home of an individual
living in Boston, identified only by his name, occupation, and city of residence. The step-by-step letters could only be sent to individuals that the current sender knew by
first name, and who were presumably closer to the final recipient. Milgram kept track
of the paths followed by the letters and of the demographic characteristics of their
handlers. At the time of these experiments, the commonly held belief was that it
would take hundreds of steps for letters to reach their final destination, but Milgram
found that the number of links needed to reach the targeted individual was six. Dodds
6
enough connecting chains so as to allow for a thorough statistical characterization.
The small-world property has been observed in a variety of real networks (including
biological and technological [2, 4, 23]), and is now an accepted mathematical
property in some network models (e.g., random graphs).
In 1998, Watts and Strogatz [21] proposed a new model for explaining small path
lengths and large clustering coefficients that are independent of network size—two
properties shared by many real networks. According to their model, the first step is to
construct a network with a one-dimensional ring lattice of N nodes (or d-dimensional
regular lattice) in which each node is wired to its neighbors up to kth nearest neighbor.
Such regular lattices have high average path lengths. Decreasing those lengths
requires the rewiring of each link with a p probability to another randomly picked
node—a process that establishes long-range connections. A small-world network
displays characteristics of a regular lattice for very small p values and an ER network
for very large p values, meaning that small-world networks lie somewhere between
order and randomness. Average path length in a small-world network is expressed as
) , (N p ∝ f( pKN) K N (1.1) u u u u u u f 4 tanh 4 4 ) ( 2 1 2 for u >> 1 or u << 1. (1.2)
7
the clustering coefficient for small-world networks is CSW ∝ (1 − p)3.
Small-world networks share some properties with a number of real networks.
However, their degree distribution has a pronounced peak at <k> = K and
exponentially decaying wings for large k, thereby distinguishing them from the power
law degree distributions of networks such as the WWW, the Internet, and many social
networks.
1.1.3 Scale-free distributions
Many scale-free networks are characterized by a power-law degree distribution [24] in
which the probability that a node has k links follows P(k) ~ k–γ, where γ is the degree
exponent. The probability that a node is highly connected is statistically more
significant than in a random graph(see Fig. 2, part Ba), with network properties often
determined by a relatively small number of highly connected nodes known as hubs. In
the Barabási–Albert scale-free model network model [24], a node with M links is
added to the network at each time point and connects to an already existing node I
with probability
I kI/
J kJ, where k is the degree of node I and J the index
denoting the sum over network nodes. The network generated by this growth process
8
distribution represented by a straight line on a log–log plot (see Fig. 2, part Bb). The
network created using the Barabási–Albert model [24, 25] does not have an inherent
modularity, meaning that C(k) is independent of k. Scale-free networks with degree
exponents 2<γ<3(a range that is observed in most biological and non-biological networks) are ultra-small, with average path lengths that follow l ~ log log N. This is
significantly shorter than log N, which is characteristic of random small-world
networks [21].
Fig. 2. The comparison between the random network and the scale-free network.
1.2 Complex Network Structure
1.2.1 Motifs
9
graphs (G) at a number that is significantly higher than in randomized versions (i.e. in
graphs with the same number of nodes, links and degree distribution as the original
one, but where the links are randomly distributed) [16, 26]. As a pattern of
interconnections, M is usually expressed as a connected (undirected or directed)
n-node graph that is a subgraph of G. All the possible three-node connected directed
graphs are illustrated in Fig. 3.
ID 1 2 3 4 5 6 7 8 9 10 11 12 13
Motif
Fig. 3. 13 possible of triad motifs defined by Alon.
The concept of motifs was originally introduced by Alon et al.[16] who studied small
n-node motifs in biological and other networks. Significant motif research in a G
graph consists of matching algorithms – that is, counting the total number of
occurrences of each n-node subgraph M in the original graph and in the randomized
graphs. The statistical significance of M is then described in terms of Z-score, defined
as rand n rand M M M M
n
n
Z
(1.3)Where nM is the number of times subgraph M appears in G, and
rand M
n and nrand
M
10
11
1.2.2 Communities
Community and the first network formalizations of the concept were proposed by
social scientists. Given a graph G(N, L), a community (or cluster, or cohesive
subgroup) can be expressed as subgraph G (N‘, L‘), whose nodes are tightly
connected, i.e. cohesive. Since the structural cohesion of the nodes of G quantified in
several different ways, there are different formal definitions of community
structures(Fig. 4)[27-29].
Fig. 4. Communities can be defined as groups of nodes such that there is a higher density of edges within groups than between them.
1.2.3 Hierarchical modularity
12
in many real systems, one must assume that clusters combine in an iterative manner to
generate hierarchical networks (Fig. 5, part A). The starting point for such
construction is a small cluster of four densely linked nodes (e.g., the four central
nodes in Fig. 5, part A). Next, three module replicas are generated and three external
nodes of the replicated clusters are connected to the central node of the old cluster,
thereby producing a large 16-node module. After generating three replicas of this
16-node module, the 16 peripheral nodes are also connected to the central node of the
old module, producing a new 64-node module. The hierarchical network model
seamlessly integrates a scale-free topology with an inherent modular structure by
generating a network that has a power-law degree distribution with degree exponent γ
= 1 + n4/n3 = 2.26(see Fig.5, part B) and a large, system-size independent average
clustering coefficient <C> ~ 0.6. A hierarchical architecture implies that sparsely
connected nodes are part of highly clustered areas, with communication between the
different highly clustered neighborhoods being maintained by a few hubs (see Fig. 5,
13
Fig. 5. The hierarchical network and its degree distribution.
1.3 Network Dynamics
Since actual complex networks are not necessarily static, simulating and/or studying
the dynamics of the complex networks is a difficult task. The following methods can
be used to address this problem in different domains.
1.3.1 Cellular automata
Cellular Automata (CA) [31]are simple examples of discrete dynamical systems. A
cellular automaton consists of a regular cell grid consisting a finite number of states.
Each ell state during time step t+1 is determined by states of cells in time step t. In the
14
bit-string CA) can be defined using the update rule defined in Table 1. In this example,
the state of a cell is determined by its own state and the states of its nearest neighbors.
This type of automaton can easily be schematized as a directed network in which each
cell takes inputs from two neighboring cells.
The time evolution of the cell states occur in discrete time steps with synchronous
update. Cellular automata are used to model several phenomena (e.g. pattern
formation) and to study various complexity theory concepts. Many types of cellular
automaton have been proposed, including random Boolean network and random
threshold networks. Three-cell block (t) 000 001 010 011 100 101 110 111 Center-cell (t+1) 0 1 0 0 1 1 1 0
Table. 1. An update rule for a one-dimensional, two-state cellular automaton.
1.3.2 Preferential attachment
15
connected nodes. Hubs are generated via ‗a rich-gets-richer‘ mechanism consisting of
growth and preferential attachment: the more connected a node is, the more likely
new nodes will link to it, meaning that highly connected nodes acquire new links
faster than their less connected peers. This mechanism ensures simultaneous the
16
Chapter 2 Static Networks and Dynamic
Process Characterization and analysis
No network in nature or technology is totally random—in other words, other
non-random mechanisms shape their evolution. The universality of various
topological characteristics, from degree distributions [25] to degree correlations [5,
22], motifs [16], and communities [2, 32], can be used as a springboard for studying
diverse phenomena and making predictions. Network theory has therefore
fundamentally reshaped our understanding of complexity. Even though researchers
still lack a universally accepted definition of complexity[32], the role of networks in
this area is obvious: all complex systems, from cells to the Internet and from social to
economic, consist of an extra-ordinarily large number of components that interact via
complex networks. We have long been aware of these networks, but only recently
have we acquired the data and tools to probe their topologies, thus giving us a clear
understanding of the strong impact of underlying connectivity on a system‘s behavior.
As a result, no single approach to complex systems can succeed unless it exploits
network topology.
17
behavior of systems that we perceive as being complex. We must be capable of
predicting how the Internet will respond to attacks and traffic jams, and how cells
react to environment changes. Progress in this direction demands an understanding the
dynamics of processes, a task made more difficult by the large number of dynamical
phenomena-almost as many as complex systems. Examples include the biological
study of reaction kinetics using metabolic networks, monitoring the flow of
information on computer networks; and exploring the spread of viruses and ideas via
social networks.
A major challenge is determining common characteristics among these diverse
dynamical processes. In this dissertation I will describe two approaches for
responding to this challenge:
For static networks, find relationships between network structure and function. I will
describe such relationships for two types of motifs in Chapter 3, and for network
hierarchy in Chapter 6
For dynamic networks, find global rules during network evolution. I will discuss
friendship evolution using three-rule model in Chapter 4 and epidemic dynamics with
18
2.1 Network Motif Detection
Commonalties have been found by complex network researchers in fields ranging
from biology to social and computer sciences. Three global features in complex
networks have been identified and investigated: highly clustered connections [1, 5],
small-world properties [1, 21], and the scale-free phenomenon [5, 7]. Approaches
based on quantitative and qualitative analyses of the topological properties of
complex networks are serving as the basis for studying how the global features of
network topological structures affect the dynamic behavior of networks [16, 33, 34].
This is currently considered one of the field‘s most important and challenging research topics [35, 36].
Some local structural motifs (also referred to as building blocks) reveal unique and
statistically significant patterns when compared with random [16], biological [1], and
food web [16, 26] motifs; all are perceived as containing important information.
However, the simple motifs of complex networks that are statistically significant but
functionally unimportant are inadequate for investigating network functions and
dynamic behaviors [16, 26]. In this dissertation, I will describe an algorithm that
19
and (b) identifies functionally and statistically significant network building blocks
from complex networks [37].
When considering the global features and local structural motifs of biological
networks, it is worth noting that link properties (weights) exert strong impacts on
network functions and dynamic behaviors [38-42]. Examples include the role of weak
links associated with the six degrees of separation (i.e., small-world) effect of
interpersonal networks [40, 41], and the strength of predator-prey interactions that
determine the stability of ecological communities [38]. Network researchers have
reported that weighted values representing interaction strength can be assigned to all
links (edges) in a real network [39, 43, 44]. I therefore considered network motif link
strength in terms of bridge motifs (consisting of weak links only or a minimum of one
weak link) and brick motifs (consisting of strong links only) (Fig. 6). Network motifs
can be separated into two categories: bridge and brick. Using the three-point
feed-forward motif as an example, it can be divided into two categories: a three-point
feed-forward brick motif (left box) composed of three strong (red) links, and a
three-point feed-forward bridge motif (right box) composed of at least one weak (blue)
link and a maximum of two strong (red) links as Fig. 6 shows. Bridge motifs connect
20
phenomenon of local clustering in biological networks.
21
2.1.1 General: Bridge and Brick Network Motif-Detecting
Algorithm
As shown in Figure 7, a link-weighted value that is dependent on the number of all
possible paths between two linked nodes equals the summation of the reciprocal
values of all possible path lengths except for the link itself. This is expressed as
i length pathi a b b a weight )) , ( ( 1 ) , ( (2.1)where pathI(a, b) indicates the i th path from node a to node b; pathi(a, b) ≠ edge(a, b);
and length(pathi (a, b)) ≤ average network diameter. The length of one path represents
its total number of nodes.
Average network diameter =
2 ) 1 | (| | | ) , ( ,
N N b a th ShortestPa b a N b a (2.2)ShortestPath(a, b) = Min(length(pathi (a, b)))
22
(a, b) of edge (a, b) is 0 while weight (b, c)
This definition implies clustering, with any increase in the number of possible paths
resulting in an increase in the clustering degree between two linked nodes.
Furthermore, the concepts and algorithms discussed in this dissertation are
generalizable to non-directed networks. To ensure that the proposed method can be
applied to any complex network, the link-weighted values calculated by the network
motif detection method are derived from the number of all possible paths between two
linked nodes within all network topological and local connection structures (no preset
link quantity). This definition is similar to that of betweenness [43, 45]—effects
resulting from the removal of network links. Accordingly, the proposed link-weighted
value calculation method is assumed to represent the importance of each link in a real
network [46, 47].
Also considered were the interactive strengths of individual links in a quantitative real
network. To validate the proposal for weighted links, they were compared with
quantitative links. However, interactive quantitative links are defined by
category-specific functions such as proteins, genes, species, and so on. It is difficult to
specify the overall impacts of these links on protein-protein interaction networks [48]
23
not reflect the significance of their connection within an overall food web.
Furthermore, each complex network type has its own measure for interactive strength.
A switching algorithm (i.e., AB, CD becomes AD, CB if AD and CB do not exist) was used to create random networks according to any given degree
sequence [16, 26]. Results from previous studies indicate that these random networks
have the same number of nodes and edges, as well as node in-degrees (incoming
edges) and out-degrees (outgoing edges) that are identical to those of real networks.
Furthermore, randomized networks preserve the same number of appearances of all
(n-1) node subgraphs as in real (original) networks [16]. The threshold that
determines the strength of an edge (link) is the mean weighted value of all edges in a
random network ensemble. Accordingly, 1,000 random networks were generated to
serve as a control. Edges were labeled ―weak‖ when their weighted values in these or real networks were smaller than the threshold minus a double standard deviation (p =
0.01); all other edges were labeled ―strong.‖ Researchers can define criteria for strong and weak links according to their own needs. Finally, all possible motifs were located,
and their distributions in real and random networks were compared.
Milo et al.‘s method [16] for identifying bridge and brick motifs in complex networks was expanded to include the following steps:
24
1. Calculate the weighted value of each link in a network of interest and an ensemble
of random networks to calculate the significance of n-node subgraphs. The goal is to
maintain the same number of appearances for all (n – 1) node subgraphs as in the
original network.
2. Label all weighted links in the network of interest and random network ensemble as
―strong‖ or ―weak‖ according to a benchmark of two standard deviations from the mean weighted value of all links in the ensemble. Links with weighted values below
the benchmark are considered weak.
3. Identify all n-node bridge/brick subgraph types in the network of interest and
random network ensemble.
4. Mark all n-node bridge/brick subgraph types by calculating their numbers in the
network of interest and random network ensemble. Each n-node bridge/brick
subgraph type is selected as a representative motif only if its frequency in the network
of interest far exceeds its frequency in the ensemble.
These steps can assist research efforts to understand the functions and roles of
identified motifs in a real network and to analyze the dynamic behaviors of complex
networks. Regarding method robustness, the proposed approach emphasizes the
25
functions of different network types.
Motif frequency can be used to measure levels of similarity between two networks of
interest. In addition, it is possible to calculate the Z-scores for all bridge/brick motifs
and significance profiles (SPs) in a network by expanding Milo et al.‘s [26, 49, 50] methods. As shown in the following formula, ZScore(Bridgei) represents the statistical
significance of the ith kind of bridge motif in a network:
( ) ( ) ( ) ( ( )) real i random i Score i random i N Bridge N Bridge Z Bridge STD N Bridge (2.3)
where Nreal(Bridgei) represents the time of appearance of the ith type of bridge motif in
a network, and <Nrandom(Bridgei)> and STD(Nrandom(Bridgei)) respectively represent
the mean and standard deviation of the time of appearance of the ith type of bridge motif in a randomized network ensemble. In the next equation, SP(Bridgei) is the
vector of ZScore(Bridgei) normalized to a length of 1. This normalization emphasizes
the relative significance of the ith type of bridge motif rather than the absolute significance. ZScore(Bricki) and SP(Bricki) can be derived in the same manner:
2
1 2 ( ) ( ) ( ) Score i i Score i Z Bridge SP Bridge Z Bridge
(2.4)(
)
(
)
(
)
(
(
))
real i random i Score i random iN
Brick
N
Brick
Z
Brick
STD N
Brick
(2.5)26
2
1 2(
)
(
)
(
)
Score i i Score iZ
Brick
SP Brick
Z
Brick
(2.6)2.1.2 Specific: Bridge and Brick Network Motif-Detecting
Algorithm
The previous method we proposed has solved some problems in different domains
successfully. However, since the concept of ―neighborhood‖ is very useful for
identifying the motifs or modules in biology. Another version of bridge and brick
network motifs is proposed for this specific domain. To ensure that the concepts and
methods described in this paper can be applied to any complex biological network, the
link-weighted value Link(v, w) for any edge between nodes v and w is expressed as its
hypergeometric coefficient Cv,w [51]. This value, which is frequently used to measure
cluster enrichment and co-occurrence significance, is expressed as:
min( ( ) , ( ) ) , ( ) ( )
( )
( )
( )
( , )
log
( )
N v N w V W i N v N wT
N v
N v
N w
i
i
Link v w
C
T
N w
(2.7)where |N(x)| is the neighborhood size of node x and T the total number of nodes in the
27
can be represented as the probability of obtaining a number of mutual neighbors
between nodes v and w at or above the observed number when the neighborhoods are
independent. The hypergeometric coefficient Cv,w is consequently defined as the
negative log of this summation. Given the neighborhood sizes of the v and w nodes
and the T total number of nodes in the biological network, the higher the value of Cv,w,
the higher the number of overlapping neighbors between v and w—an indication that
Link(v, w) has a higher clustering coefficient. Otherwise, it does not belong to any
specific cluster (Fig. 8). Different link definition differs between the general algorithm
and the specific algorithm for detecting bridge and brick motifs, other parts are all the
same.
Fig. 8. The small-world model. Black signifies strong links and red weak links.
2.2 Social Network Simulation
28
The first rule addresses how people make new friends (via introductions or
meetings-by-chance), and the second rule addresses how friendships are broken when
one party dies. The model entails a fixed number of N nodes and undirected links
between pairs of nodes representing individuals who know each other [53]. To reflect
friendship weakening and strengthening, I added a ―friend remembering‖ rule (Fig. 9). The model repeats the following three rules until the acquaintance network in
question reaches a statistically stationary state:
Rule 1 (friend making). Randomly chosen individuals introduce two friends to each
other. If this is their first meeting, a new link is formed between them. Randomly
chosen persons with less than two friends introduce themselves to one other person at
random. Note that the term ―introduce‖ is used to describe meetings by chance as well as meetings via a common friend.
Rule 2 (leaving and arriving). At a p probability, a randomly chosen individual and all
associated links are removed from a network and replaced by another person.
Accordingly, acquaintances can be viewed as circles of friends whose members can
leave for reasons other than death and enter the circle for reasons other than birth.
Rule 3 (friend remembering). A certain number of friendships are updated, with the
29
explained in detail in the following two sections.
Fig. 9. Three-rule model flow diagram.
2.3 Epidemic Dynamics Analysis
The state transfer concept of SIS models adopted by Pastor-Satorras [54, 55] was
applied as the core simulation model architecture. Parameters were incorporated to
simulate behavioral and transformative results arising from agent interactions [56-60].
Each agent (node) in a complex network owns a set of properties and behavioral rules
that are used to demonstrate the features and statuses of persons in social networks or
computers connected to the Internet. A link between two nodes means that the
30
interaction/communication channel. An infectious disease or computer virus can be
transmitted via this link. At each discrete time step, the epidemiological state of each
node is determined by its behavioral rules, original epidemiological state, neighbors‘
epidemiological states, infection rate, and recovery rate. As stated above, ρ(t) is
defined as the density of infected nodes present during time step t. When time step t
becomes infinitely large, ρ can be represented as a steady density of infected nodes. A
computational flowchart for the proposed simulation model is shown in Figure 10. A
complex network G(N, M) with N nodes and M links was constructed using the
algorithm described previously prior to setting relevant parameters and attributes for
the nodes involved in the simulation; discrete time t was set at 0. During simulations,
nodes take turns interacting with neighboring agents for specified time intervals. Note
that individual agent interactions do not result in immediate influences, and that
simultaneous state changes only occur when all agents in a complex network
complete their interactions. Accordingly, interaction sequences do not influence
interaction processes or results.
At the beginning of each discrete time step, the usable economic resources of each
agent vi are reset to R(vi), meaning that all agents renew and/or receive supplemental
31
of sleep. In our later experiments, the statistical distribution of individual economic
resources could be delta (rConstant), uniform, normal, or power-law, as long as the mean
value <r> satisfied: t Cons N i i r N v R r 1 tan ) (
(2.8)At each discrete time step, each vj agent interacts with one agent selected from all of
its Neighbors(vi). After the interaction process is completed, agents vi and vj must
have transmission costs c(vi) and c(vj) (0 ≤ c(vi) ≤ R(vi) and 0 ≤ c(vj) ≤ R(vj)) deducted
from their respective economic resources, regardless of the interaction result. If R(vi)
< c(vi) after the interaction, agent vi cannot interact with other neighbors because all
of its resources have been used up. Otherwise, it repeats the interaction process by
choosing another neighboring agent until its resources are exhausted.
Assume that infected and contagious agent vi is adjacent to susceptible and
infection-prone agent vj. When the two agents come into contact, a combination of
infection rate RateInfect, agent vj‘s resistance level, and a random number r
determines whether or not vj is infected by vi. If the random number r is lower than
the infection rate RateInfect, agent vj‘s epidemiological state becomes I (Infected).
32
recovery rate. Without a lack of generality, recovery rate RateReset can be assigned as
1, meaning if agent vj is infected by other agents at discrete time step t – 1, it will
recover and once again become susceptible at discrete time step t, since it only takes
affect according to the definition of the infection disease propagation time scale. At
the beginning of an infectious disease simulation, only ten individuals were given I
status; all others were given S(Susceptible). During each time step, agents randomly
interacted with several neighbors. All epidemic experiments discussed in this paper
33
Fig. 10 Flowchart for a SIS epidemiological simulation model.
2.4 Abstraction Hierarchy
The proposed two-way method is considered novel because it emerges from top-down
and bottom-up clustering algorithm synergy [61, 62]. Not only does it identify
modules in a top-down fashion and construct a hierarchy implied in a complex
34
at different levels in the hierarchy. The method consists of three steps: (i) computing
between-node proximity, (ii) extracting the backbone(represented by a spanning tree)
from the network and using it to partition the network [28, 63], and (iii) generating an
abstract network. Iteratively applying the same steps to a newly generated abstract
network supports the discovery of an abstraction hierarchy within a complex network
[2, 64].
2.4.1 Proximity Measure
Proximity between two nodes can be defined in many ways; since it affects resulting
module formation, selecting an appropriate proximity function is very important.
Commonly used measures include Euclidean distance, correlation coefficient and
cosine similarity [65, 66]. Module analysis is problem-dependent as stated earlier, in
this dissertation I investigate clustering based on network topology. Conventional
proximity measures are not applicable to clustering problems if network topology
represents the only available information-that is, Euclidean distance cannot be
calculated without node coordinates. Other proximity measures( e.g. edge
betweenness [45] and topological overlap[65, 67, 68]) have recently been proposed
35
While some successful applications have been reported, they have at least two
limitations: (a) edge betweenness of node pairs reflects the global characteristics in a
network, but they suffer from high computational costs [64] and are affected by the
network incompleteness and noise [32, 64]; and (b) sine topological overlap is a local
measure, it poses a challenge to identifying any module beyond a locally dense
connectivity pattern [69].
Most proximity measures in current use do not take link direction or weight into
account. Therefore, any directed weighted network is processed as an undirected
unweighted one. To expand its applicability, I propose using a new proximity function
for dealing with directions and weights. For the sake of simplicity, I will describe a
directed weighted network of n nodes by an nn adjacency matrix A, in which each element Aij is the weight of the link from node i to j. A zero-valued weight (Aij=0)
indicates the absence of a link between those nodes. The proximity function prox(i,j)
from node i to j, i j is defined as:
0, 0 , , , , , , , , { min( , )} ) , ( j k k i j k k i j k k i j i A A k out k j i out i A A W A A W A A j i prox , Wiout A i , m m n
(2.9)36
function considers not only the effects of common neighbors (i.e. node k), but also
link direction and weight. It treats the direct link from node i to j differently than
indirect paths between the same nodes through a common neighbor k. The weight of
the direct link contributes to prox(i, j), as indicated by the first term in the above
equation. To calculate i-to-j proximity based on an indirect path from i to j by way of
k, I divided the path into two sub-paths, from i to k and from k to j. Assuming on an
indirect path one node does not always affect all its neighbors, but instead acts
probabilistically. Thus, the probability that one node affects another(e.g. i affects k) is
defined as the ratio of the link weight between them to the sum of the weights of all
outgoing links from node i, not including the direct link from i to j. The probability of
an indirect path from i to j by way of k is therefore the product of the probability of
paths from i to k and the path from k to j. The proximity between i and j contributed
by the indirect path ikj is assigned to the probability times the minimum of Aik and
Akj. In cases where there is more than one common neighbor of i and j, the sum of the
proximity of each indirect path is used.
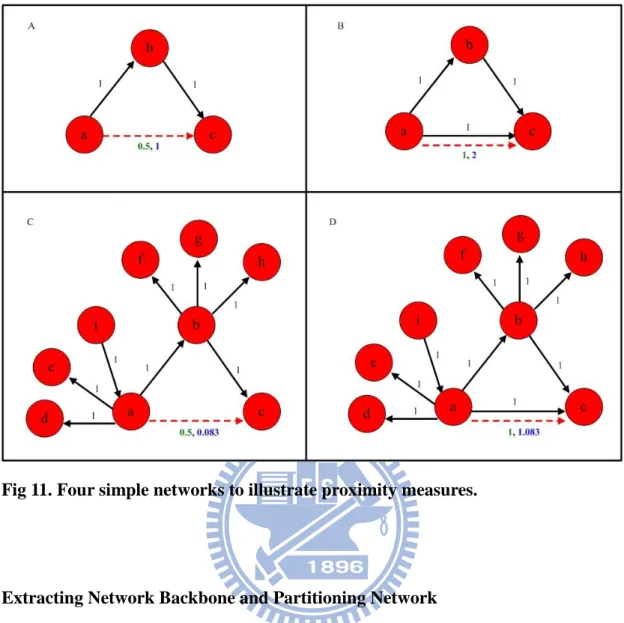
The examples shown in Fig. 11 illustrate our proximity function and compare it with a
related measure, topological overlap [67, 68, 70]; both take common neighbors into
37 defined as follows: Tij lijaij min( di,dj)1aij , (2.10)
where lij is the number of common neighbors shared by node i and j, di is the degree
of node i, and aij=1 if a direct link exists between i and j (otherwise, aij=0). The 1- aij
quantity in the denominator prevents the denominator from becoming zero in case
where min(di,dj)=0. The inclusion of aij in the numerator is to make Tij explicitly
dependent on the direct link between i and j.
Given the network in Fig. 11A, Tac=0.5 and prox(a,c)=1. To evaluate the effects of
direct links, one direct link was added between nodes a and c (Fig. 11B). If we
compare the network to a gene regulation model, a can be interpreted as a regulator, b
as an intermediate gene, and c as a target. Since gene a can regulate gene c either
directly, or through the intermediate gene b, the proximity between a and c in Fig.
11B should be higher than that in Fig. 11A. Increase in proximity were found in both
measures-that is, Tac=1 and prox(a,c)=2 vs. Tac=0.5 and prox(a,c)=1. The network
shown in Fig. 11C is different from the one in Fig. 11A in that node a and b both have
more neighbors. Considering the network as a model of gene regulation, it means that
38
of gene a on gene c may be diminished. For node a and node c in Fig. 11C, prox(a,c)
decreases to 1/12, which corresponds reasonably to the gene network model. In
contrast, according to the topological overlap measure, the proximity between nodes a
and c is 0.5, which is the same as that in Fig. 11A. Topological overlap measure fails
to distinguish between Fig. 11A and 11C. To complete the illustration, we added one
direct link between a and c, and created the network shown in Fig. 11D. An increase
in proximity results from either measure.
Even though the proposed proximity function is a local measure(similar to topological
overlap), it shows better discrimination in network topology and requires less in
computational costs than global measures such as edge betweenness. Incorporating
the proximity function into a two-way module-finding and hierarchy-building strategy,
it is possible to gather global characteristics and to detect the hierarchical structure of
a network. We validated our approach using hierarchically nested random networks as
39
Fig 11. Four simple networks to illustrate proximity measures.
Extracting Network Backbone and Partitioning Network
An optimal solution for network partitioning (based on a criterion function) emerges
after enumerating all possibilities, but it is computationally prohibitive for large
networks. In response to this problem, a graph-theoretic approach to partitioning was
adopted [71]. After computing the proximity between all node pairs, it is possible to
build a maximum spanning tree that includes all network nodes, which are all
40
proximities are discarded, the maximum spanning tree acts as the network backbone.
To reduce computational costs, partitioning is performed based on the maximum
spanning tree instead of the original network.
Two subtrees can be obtained by removing one link from a tree, with each subtree
representing one module. One tree can be partitioned into many subtrees (i.e.
modules/clusters) by repeating the same process on each subtree. Given the maximum
spanning tree, resulting modules can be examined by removing one link from a
(sub)tree. A link is selected for removal if the M={M1,M2,M3,…,Mn} set of modules
meets the following criteria after removal:
M
a,M
b
M, a
b S
int raMa
S
int erMa,Mb and
S
int raMb
S
int erMa,Mb (2.11) Where
S
int raMk = Ai, j i, jCk
is the sum of the proximity of each intralink within Mk, and
S
int erMa,Mb = Ai, j iCa, jCb
is the sum of the proximity of each interlink between Ma and Mb.
These criteria for modules are similar to those described in [64] and [72], except that
link weight (i.e., proximity) is considered instead of node degree. The simple example
network shown in Fig. 12 demonstrates the advantage of using the link weight criteria.
Without taking the weight into account, intuitively the most appropriate partition of
the network is to cut the link between node C and node F, and obtain two modules.
41
nodes only, the simple network will be partitioned in the way above. However, in
practice, if the weight represents the significance of connectedness, the network
should be considered as a whole. Our criteria for modules take weights into account;
therefore, the network cannot be divided based on our criteria. In the case of an
unweighted network, by treating each link as one with a constant weight, e.g. one, this
simple network will be partitioned into two modules as expected according to our
criteria. Without losing generality, this simple network demonstrates that our criteria
for modules are more realistic, and can subsume the previous definitions of modules
[72]. Note that the proximity sum is calculated for the network rather than the tree,
thus preventing information loss. In the proposed model, the tree is only used for
evaluating which nodes may form clusters, thus reducing the search space of the
original network.
42
Pseudocode for the partitioning procedure is shown below, starting with a single
module represented by a maximum spanning tree. The input includes the network Net
in question; M1 is its maximum spanning tree. The output consists of partitioning
result in clusters.
Procedure Network_Partition (Net, M1)
M={M1} //M keeps the modules for further analysis
Repeat
{
Select a cluster Mi M, and remove Mi from M.
Put Mi into D. //D stores the final clusters
Put all the links of Mi in Li.
While (Li is not empty)
{
Set the link in Li with min proximity as lmin.
Remove lmin from Li.
43
Add Ma and Mb to M.
If M does not satisfy criteria [4]
{
Remove Ma and Mb from M.
Restore lmin to Mi. //put the link lmin back to the tree Mi
}
Else
{
Remove Mi from D. // because Mi is legally split into Ma and Mb
Break; //break out of While loop
}
} until M is empty.
Output D.
2.4.2 Network Abstraction
After the partition of the network, each module is treated as a supernode [7, 73] and a
network of the supernodes is viewed as an abstraction of the original network. An
abstract network reveals the general framework of the original network without any
44
module Ma and Ma) is defined as
b a b a M n M m M M b a erM
M
prox
m
n
prox
, 1 sup(
,
)
(
,
)
(2.12)where |Ma| is the number of nodes in module Ma. Proximity between all possible
supernode pairs are computed and normalized to a z-score. Links that have z-scores
below a pre-set threshold are considered insignificant and therefore discarded. The
resulting supernode network (an abstraction of the original network) is placed one
level higher than the original network in the hierarchy. By repeating the same process
with other networks in the hierarchy, it is possible to generate additional abstract
networks and to consistently and systematically build a pyramid of abstraction from
45
Chapter 3 Network motif Experiments
Researchers are making progress toward defining organizing principles that govern
the formation and evolution of complex biological networks. Considered a major
challenge in computational system biology, predicting network behaviors [74] and
functions requires the identification of functionally and statistically important motifs.
To understand their structural organizing principles and evolutionary mechanisms,
bridge motifs can be defined as consisting of weak links only or at least one weak link
and multiple strong links; brick motifs can be defined as consisting of strong links
only. Next, an algorithm is proposed for performing two simultaneous tasks: detecting
global statistical features and local connection structures in biological networks, and
locating functionally and statistically significant network motifs.
3.1 General: Bridge and Brick Network
Motif-Detecting Algorithms
Commonalties have emerged from studies of complex networks in fields ranging from
biology to social and computer sciences. Three global features in complex networks
46
75], small-world properties [21, 75-78], and the scale-free phenomenon [1, 24, 39, 79].
Approaches based on quantitative and qualitative analyses of the topological
properties of complex networks are being utilized to study how the global features of
network topological structures affect the dynamic behavior of networks [1, 39, 80-84]
39-40]. This is currently considered one of the field‘s most important and challenging
research topics [1, 39].
Some local structural motifs (building blocks) reveal unique and statistically
significant patterns when compared with random [16, 80, 85-90], biological [16, 87],
and food web [16, 38] motifs; all are thought to contain important information.
However, simple motifs of complex networks that are statistically significant but
functionally unimportant are clearly inadequate for investigating network functions
and dynamic behaviors [16, 82, 88, 90-93]. An algorithm is therefore proposed to
perform two tasks: simultaneously detect global features and local structures in
complex networks, and identify functionally and statistically significant network
building blocks from complex networks.
When considering the global features and local structural motifs of biological
networks, it is worth noting that link properties (weights) exert strong impacts on