An Observation on the RNA/DNA Ratio of Individual
Japanese Anchovy Larva from Four Hydrographic
Regions in the Northeastern Waters of Taiwan
TAI-SHENG CHIU AND JIUNN-BIN HUANG
Department of Zoology National Taiwan University
Taipei, Taiwan, R.O.C.
(Received August 20, 1998; Accepted December 29, 1998)
ABSTRACT
The RNA/DNA ratios of Japanese anchovy (Engraulis japonica) larvae caught from the northeastern waters of Taiwan were examined and compared in this study. Four types of water masses can be distinguished in the area; thus, four hydrographic regions are defined as the southern East China Sea (SECS), Kuroshio (KUR), surface mixing water (MIX) of SECS and KUR, and upwelling water (UWW) from subsurface KUR. The nucleic acid contents of indi-vidual larva were measured, and the RNA/DNA ratio was used as an index to compare the nutritional condition of larvae from among the waters. In general, the RNA and DNA contents were related to body size, but the RNA content had higher variability in correspondence to hydrographic differences. Larvae from the KUR showed a significantly lower RNA/DNA ratio than did those from the SECS, MIX and UWW. Finally, a relatively poor nutritional condition of Japanese anchovy larvae in the KUR is inferred, and the distribution pattern of Japanese anchovy in the waters around Taiwan is explained.
Key Words: nutritional condition, anchovy, East China Sea, Kuroshio
I. Introduction
The Japanese anchovy Engraulis japonica is an im-portant pelagic species in the East China Sea (Hayasi, 1967). During spring, its southern stock migrates to the northeast-ern waters of Taiwan for spawning (Young and Chiu, 1994). The area, including a coastal bay area, is an important nursery area for planktonic larvae of Japanese anchovy (Huang and Chiu, 1996).
The shelf area of northeastern Taiwan is part of the East China Sea, the water of which is affected seasonally by intrusion of the Kuroshio from the southeast (Chuang and Liang, 1994). This intrusion event induces mixing of Kuroshio surface water with that of the East China Sea and causes the local Kuroshio subsurface water to move up-ward (Chern et al., 1990; Liu et al., 1992; Gong et al., 1995). The upwelling in the northeastern waters of Taiwan has been found to be rich in nutrients (Gong et al., 1995) and chloro-phyll-a (Chen, 1995).
The quantity and quality of food items in a water en-vironment are major factors influencing fish stock in their early life stage. A lack of food or a mismatch in distribu-tion of larval fish and food organisms is, therefore, a major cause of high larval mortality (Fortier et al., 1995; Busch, 1996; Gotceitas et al., 1996). Since the RNA/DNA ratio
has been determined to be an effective index of the nutri-tional status in the larval stage (Buckley, 1984; Clemmesen, 1994), we adopt this method to examine the RNA/DNA ratio of Japanese anchovy larvae from four hydrographic regions in the northeastern waters of Taiwan, and to infer their rela-tive fitness in different marine environments.
II. Materials and Methods
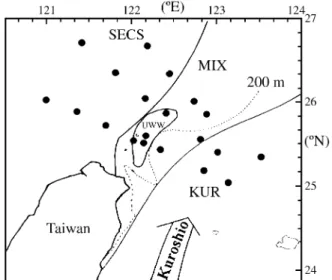
Japanese anchovy larvae were sampled during April 24-28, 1996 by R/V Ocean Research I in the northeastern waters of Taiwan, approximately bounded by 25º-27ºN and 121º-124ºE (Fig. 1). The larvae were collected using a Bongo net (Hydro-Bios), with a mouth diameter of 60 cm, length 250 cm and mesh size 500 µm, operated in a water column within a depth of 200 m. Obtained Japanese an-chovy larvae were sorted within five minutes and stored in liquid nitrogen immediately.
At each station, profiles of the temperature and salin-ity of the water were measured using a gauge with conductivity, temperature and depth (CTD) sensors (SBE 9/11, Sea-Bird Electronics, Inc.). The patterns of tempera-ture-salinity diagrams, the bottom depth, and satellite im-ages of the sea surface temperature were used to classify four hydrographic regions. These include two basic
end-members of the southern East China Sea (SECS, low tem-perature and low saline situated on the continental shelf) and the Kuroshio (KUR, high saline and high temperature situated far offshore, with a bottom much deeper than 500 M), and two derived members of the mixing water (MIX, relatively low surface temperature situated on the continental slope between the SECS and KUR) and the upwelling wa-ter (UWW, surface temperature less than 20 ºC, forming a significant cold dome in satellite images). The identifica-tion of water categories was carried out by generally fol-lowing Chiu and Lee (1991), Chiu and Hsyu (1994), and Huang and Chiu (1998). For each larva, the body length was measured using the notochord length (pre-flexion and flexion larvae) or total length (post-flexion larvae). Mea-surements of RNA and DNA contents basically followed Clemmesen (1993), with minor modification. The RNA and DNA were first purified from individual larva homogenate, and the contents were subsequently measured using the fluorescence photometric method with the nucleic acid fluorescence dye, ethidium bromide. The content of total nucleic acid (RNA+DNA) was determined first, and the content of DNA was determined after RNA having been digested by RNase. The fluorescence index was read by using a spectrophotometer (F-2000, Hitachi), where a wave-length of 365 nm was selected for excitation, and the emit-ted light was observed at 590 nm.
Thirty individuals from each classified region were randomly selected for this study. Since the RNA/DNA ra-tio may be confounded by the body size (Westerman and Holt, 1988), analysis of variance was used to separate the size effect, and comparison of the ratios among different waters was carried out within a relatively small body size
interval of 2 mm total length (TL). Duncan’s multiple range test (Snedecor and Cochran, 1980) was used to test the differences in the RNA/DNA ratios among the 4 regions.
III. Results
1. General Status
During the spring of 1996, four water types, viz. SECS, MIX, UWW and KUR, were identified in the northeastern waters of Taiwan (Fig. 1). The environmental factors per-taining to the stations from these waters, including bottom depth, water temperature and salinity, were measured at a depth of 50 m, and the results are shown in Table 1. The SECS, located on the continental shelf, had a shallower bottom depth of 67-128 m, a temperature of 15.86-20.11 ºC and a salinity of 33.49-34.25 ppt. The MIX, lying on the borders of the continental shelf, had an intermediate bottom depth of 113-709 m, a temperature of 18.46-20.41 ºC and a salinity of 34.42-34.56 ppt. The UWW was discovered in a relatively smaller range with a depth of 97-194 m, a temperature of 17.71-18.84 ºC and a salinity of 34.40-34.51 ppt. The temperature of the UWW was lower than that of the water in its vicinity although the extreme cold water found in the SECS were located very close to the coast of Mainland China with no ambiguous overlap in geographic range. Therefore, the UWW can be identified reasonably precisely. The KUR, which stretched out into the open sea, had a very deep bottom of 839-1343 m, hot water in the range 21.61-24.49 ºC and a high level of salin-ity of 34.55-34.81 ppt.
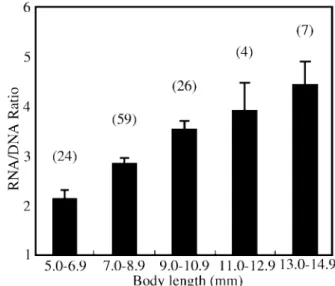
2. Variation in RNA/DNA Ratio with Body Size
The scatter plots of DNA and RNA contents for each larva are shown in Fig. 2. The magnitude of the RNA con-tent per individual larva apparently increased with the in-crease in fish size, but that of DNA remained fairly constant. A linear relationship between body length and RNA was found as follows: RNA (µg/ind.) = –10.291 ± 1.992 TL (mm) (r = 0.828, p < 0.001, n = 120). The RNA/DNA ratio was also found to be body size dependent (Fig. 3). The lowest Fig. 1. Schematic diagram showing four hydrographic regions in the
waters off northeastern Taiwan: the southern East China Sea (SECS), the mixing water (MIX), the upwelling water (UWW) and the Kuroshio (KUR). Solid circle (•): Survey stations occupied.
Table 1. Basic Information of Four Hydrographic Regions where
Japa-nese Anchovy Larvae Were Sampled
Hydrographic Environmental factor (ranges) Regions Depth (m) Temperature (ºC)a Salinity (ppt)a
SECS 67 – 128 15.86 – 20.11 33.49 – 34.25
MIX 113 – 709 18.46 – 20.41 34.42 – 34.56
UWW 97 – 194 17.17 – 18.84 34.40 – 34.51
KUR 839 – 1343 21.61 – 24.49 34.55 – 34.81
value of the ratio (2.166 ± 0.176) was found for fish with 5.0-6.9 mm TL. The ratio increased as the size of the fish increased: 2.872 ± 0.115 (7.0 – 8.9 mm TL), 3.550 ± 0.165 (9.0 – 10.9 mm TL), 3.911 ± 0.554 (11.0 – 12.9 mm TL), and 4.409 ± 0.469 (13.0 – 14.9 mm TL), for each size group, respectively.
3. Variation in RNA/DNA Ratio with Hydro-graphic Region
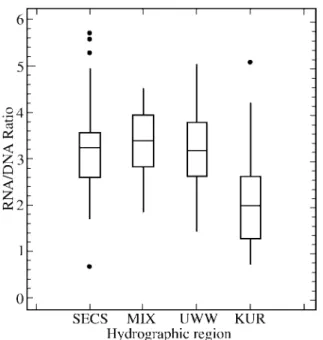
The ratios for fish obtained from four hydrographic regions were found to be different (Fig. 4; p < 0.001). Multiple range test revealed a group of SECS + MIX + UWW to be in contrast to KUR. The average ratios and standard deviation were 3.259 ± 0.213, 3.388 ± 0.130, and 3.180 ± 0.144, for SECS, MIX and UWW, respectively. However, they were found to be significantly lower for KUR (2.181 ± 0.204).
IV. Discussion
Upwelling and intrusion are the two mechanisms which transport water from the Kuroshio onto the southern East China Sea shelf of northeastern Taiwan. The upwelling Kuroshio water (UWW) carries surplus nutrients to the pho-tosynthetic layer of the shelf water and causes high primary productivity in the northeastern waters of Taiwan (Gong et al., 1995). The intrusion of the Kuroshio into the southern East China Sea causes water mixing and forms an ecotonal region (MIX) between the southern East China Sea and the Kuroshio. Therefore, four hydrographic regions and, thus, ecological regions can readily be found during the spring blooming season of Japanese anchovy larvae.
It has been suggested that fish production is high in upwelling areas (Ryther, 1969). However, this phenom-enon did not occur in the core upwelling region in the wa-ters of northern Taiwan (Chu et al., 1993). This study is the first time to compare the nutritional condition of anchovy larvae across four hydrographic regions in the waters of NE Taiwan. In our study, no high values of the RNA/DNA ratio, an index of nutritional condition (Canino, 1994), were found for larval Japanese anchovy from the UWW (Fig. 4). This result further shows that the persistent upwelling wa-ter of northeaswa-tern Taiwan might not provide additional food to Japanese anchovy larvae. Probably, this further shows that the upwelling water has little benefit for fish species of non-phytoplankton eaters other than Japanese anchovy. However, this inference should not be extended prematurely to other fish larvae, such as scombers (Auxis spp.) and scads (Decapterus spp.), which have been found to be abundant in the UWW (Huang and Chiu, 1998).
The migration of adult Japanese anchovy to the coast Fig. 2. Scatter plots showing RNA (••) and DNA (•) contents (µg) and total
length (mm) of individual Japanese anchovy larva.
Fig. 3. Plot of RNA/DNA ratio by body length (mm) for Japanese anchovy
larvae. Vertical bar, within group standard deviation; number in (), number of larva examined.
of Taiwan is related to reproduction (Young and Chiu, 1994). The earliest larvae were found in I-lan Bay and Pan-Cha-Yu, adjacent to the UWW location (Chiu et al., 1997). Relatively few larvae had been transferred across the hy-drographic front into the KUR (Young et al., 1995). The RNA/DNA ratios of the anchovy larvae collected in the KUR were the lowest in the studied hydrographic regions (Fig. 4). Based on assessment using the height of midgut cells, Hung (1995) found that 75% of the anchovy larvae in the Kuroshio were starving. Shimizu et al. (1989) also showed that the nutritional condition of the sardine Sardinops melanostictus collected from the offshore wa-ters of the Kuroshio is poorer than that of the same sardine collected from the coastward side. It is, hence, hypothesized that the oligotrophic Kuroshio water along the western boundary of the Pacific is incapable of supporting anchovy larvae with sufficient quantity and suitable quality of food. On the other hand, the better food quality found in non-KUR regions, as indicated by an increase of 20-40% in the protein level (Chen and Chen, 1992), greatly enhances the nutritional conditions for Japanese anchovy. As a result, the Kuroshio water is not a favorable environment for the development of Japanese anchovy larvae. Accordingly, no adult Japanese anchovy were found in the Kuroshio and the offshore area of eastern Taiwan (Young et al., 1994).
Acknowledgment
We thank the crew members of the R/V Ocean Research I for their
assistance in collecting the samples. Special thanks are also extended to Dr. C.T. Shih for kindly reviewing this manuscript and to Dr. C. Clemmesen for technical consultation on nucleic acid analysis. This study was par-tially funded by the National Science Council, Republic of China, under grant No. NSC 85-2611-M003-001-K2.
References
Buckley, L.J. (1984) RNA-DNA ratio: an index of larval fish growth in the sea. Mar. Biol., 80:291-298.
Busch, A. (1996) Transition from endogenous to exogenous nutrition: lar-val size parameters determining the start of external feeding and size of prey ingested by Ruegen spring herring Clupea harengus. Mar. Ecol. Prog. Ser., 130:39-46.
Canino, M.F. (1994) Effects of temperature and food availability on growth and RNA/DNA ratios of walleye pollock Theragra chalocgramma (Pallas) eggs and larvae. J. Exp. Mar. Biol. Ecol., 175:1-16. Chen, H.Y. and Chen, Y.L.L. (1992) Quantity and quality of summer
sur-face net zooplankton in the Kuroshio current induced upwelling north-east of Taiwan. Terrestrial, Atmosphere and Oceanography, 3:321-334. Chen, Y.L.L. (1995) Phytoplankton composition and productivity in re-sponse to the upwelling off northeastern Taiwan. Proc. Natl. Sci. Counc. ROC(B), 19(1):66-72.
Chern, C.S., Wang, J. and Wang, P.D. (1990) The exchange of Kuroshio and East China Sea shelf water. J. Geophys. Res., 95:16017-16023. Chiu, T.S. and Hsyu, Y.H. (1994) Interannual variation of ichthyoplankton
density and species composition in the waters off northeastern Taiwan. Mar. Biol., 119:441-448.
Chiu, T.S. and Lee, P.Y. (1991) Initial ichthyoplankton studies in the Kuroshio edge exchange area. Bull. Inst. Academia Sinica., 30:261-292.
Chiu, T.S., Young, S.S. and Chen, C.S. (1997) Monthly variation of larval anchovy fishery in I-lan Bay, NE Taiwan, with an evaluation for opti-mal fishing season. J. Fish. Soc. Taiwan, 24(4):273-282.
Chu, T.J., Yeh, S.Y. and Liu, H.C. (1993) Hydroacoustic biomass distri-bution in the waters off northern Taiwan in 1990 winter season. Acta Oceanogr. Taiwanica., 30:22-24.
Chuang, W.S. and Liang, W.D. (1994) Seasonal variation of the Kuroshio water across the continental shelf northeast of Taiwan. J. Oceanogr.,
50:531-542.
Clemmesen, C.M. (1993) Improvements in the fluorometric determina-tion of the RNA and DNA content of individual marine fish larva. Mar. Ecol. Prog. Ser., 100:177-183.
Clemmesen, C.M. (1994) The effect of food availability, age or size on RNA/DNA ratio of individually measured herring larvae: laboratory calibration. Mar. Biol., 118:377-382.
Fortier, L., Ponton, D. and Gilbert, M. (1995) The match/mismatch hy-pothesis and the feeding success of fish larvae in ice-covered south-eastern Hudson Bay. Mar. Ecol. Prog. Ser., 120:11-27.
Gong, G.C., Liu, K.K. and Pai, S.C. (1995) Prediction of nitrate concen-tration from two end member mixing in the southern East China Sea. Continental Shelf Res., 15:827-842.
Gotceitas, V., Puvanendran, V., Leader, L.L. and Brown, J.A. (1996) An experimental investigation of the match/mismatch hypothesis using larval Atlantic cod. Mar. Ecol. Prog. Ser., 130:29-37.
Hayasi, S. (1967) A note on the biology and fishery of the Japanese an-chovy Engraulis japonica (Houttuyn). Rep. California Coop. Oceanic Fish Invest., 11:44-57.
Huang, J.B. and Chiu, T.S. (1996) Daily growth and length distribution of larval Japanese anchovy (Engraulis japonica) in the neritic waters off northeastern Taiwan. Acta Zool. Taiwanica., 7:29-42.
Huang, J.B. and Chiu, T.S. (1998) Seasonal and hydrographic variations of ichthyoplankton density and composition in the Kuroshio edge ex-change area off northeastern Taiwan. Zoological Studies, 37(1):63-73.
Fig. 4. Box-and-whisker plots showing the variation of the RNA/DNA
ratio of Japanese anchovy larvae among four hydrographic environments. Bar within box, group median; box, range of 50% individuals; whiskers, the extremes of data; four outliers (•) ob-served in SECS and one in KUR.
Hung, S.J. (1995) Histological Study on Nutritional Status of Larval Japa-nese Anchovy (Engraulis japonica T & S), p. 63. M.S. Thesis. Na-tional Taiwan University, Taipei, Taiwan, R.O.C.
Liu, K.K., Gong, G.C., Lin, S., Yang, C.Y., Wei, C.L., Pai, S.C. and Wu, C.K. (1992) The year-round upwelling at the shelf break near the north-ern tip of Taiwan as evidenced by chemical hydrography. Terrestrial, Atmosphere and Oceanography, 3:243-276.
Ryther, J.H. (1969) Photosynthesis and fish production in the sea. The production of organic matter and its conversion to higher forms of life vary throughout the world ocean. Science, 166:72-76.
Shimizu, H., Nakata, K. and Nakano, H. (1989) Comparison of nutritional condition of sardine larvae, Sardinops melanostictus (T & S), taken from the coastal and offshore region of the Kuroshio current. Nippon Suisan Gakkaishi, 55:179.
Snedecor, G.W. and Cochran, W.G. (1980) Statistical Methods, p.507. The Iowa State University Press., Ames, IA, U.S.A.
Westerman, M.E. and Holt, G.J. (1988) The RNA/DNA ratio: measure-ment of nucleic acids in larval Sciaenops ocellatus. Contrib. Mar. Sci.,
30(Suppl.):117-124.
Young, S.S. and Chiu, T.S. (1994) Ovarian maturation of Japanese anchovy, Engraulis japonica T. & S., from I-lan bay, northeastern Taiwan. Zoo-logical Studies, 33(4): 302-309.
Young, S.S., Chiu, T.S. and Shen, S.C. (1994) A revision of the family Engraulidae (Pisces) from Taiwan. Zoological Studies, 33(3): 217-227. Young, S.S., Chiu, T.S. and Shen, S.C. (1995) Taxonomic description and distribution of larval anchovy (Engraulidae) occurred in the waters around Taiwan. Acta Zool. Taiwanica, 6(1):33-60.