A strong endoplasmic reticulum retention signal in the stem
–anchor region of
envelope glycoprotein of dengue virus type 2 affects the
production of virus-like particles
Szu-Chia Hsieh
a, I-Jung Liu
a,d, Chwan-Chuen King
c, Gwong-Jen Chang
e, Wei-Kung Wang
a,b,⁎
a
Institute of Microbiology, College of Medicine, National Taiwan University, Taipei, Taiwan
b
Department of Internal Medicine, National Taiwan University Hospital, Taipei, Taiwan
c
Institute of Epidemiology, College of Public Health, National Taiwan University, Taipei, Taiwan
d
Cardinal Tien College of Healthcare and Management, Taipei, Taiwan
eDivision of Vector-Borne Infectious Diseases, Center for Disease Control and Prevention, Public Health Service, U. S. Department of Health and Human Service,
Fort Collins, Colorado, USA
Received 5 October 2007; returned to author for revision 25 October 2007; accepted 18 December 2007 Available online 5 February 2008
Abstract
Recombinant virus-like particles (VLPs) of flaviviruses have been shown to be produced efficiently by co-expressing the precursor membrane (PrM) and envelope (E) proteins with few exceptions, such as dengue virus type 2 (DENV2). It was reported previously that chimeric DENV2 PrM/E construct containing the stem–anchor region of E protein of Japanese encephalitis virus (JEV) produced VLPs efficiently (Chang, G. J., Hunt, A. R., Holmes, D. A., Springfield, T., Chiueh, T. S., Roehrig, J. T., and Gubler, D. J. 2003. Enhancing biosynthesis and secretion of premembrane and envelope proteins by the chimeric plasmid of dengue virus type 2 and Japanese encephalitis virus. Virology 306, 170–180.). We investigated the mechanisms involved and reported that compared with authentic DENV2 PrM/E-expressing cells, E protein in chimeric DENV2 PrM/E-expressing cells was also present in an endoglycosidase H (endo H)-resistant compartment and has shifted more to the pellets of the soluble fraction. Replacement of the transmembrane and cytoplasmic domains of CD4 with the stem–anchor of DENV2 (CD4D2) or JEV (CD4JEV) rendered the chimeric CD4 retained predominantly in the endoplasmic reticulum (ER). Flow cytometry revealed higher proportion of CD4JEV than CD4D2 expressed on the cell surface. Together, these findings suggested that the stem–anchor of DENV2 contained an ER retention signal stronger than that of JEV, which might contribute to the inefficient production of DENV2 VLPs. Moreover, co-expression of C protein can enhance the production of DENV2 VLPs, suggesting a mechanism of facilitating viral particle formation during DENV2 replication.
© 2007 Elsevier Inc. All rights reserved.
Keywords: Dengue virus; Stem–anchor region; Endoplasmic reticulum; Virus-like particles
Introduction
Dengue viruses (DENV) are members of the genus Flavi-virus in the family Flaviviridae. Among the 70 or so arthropod-borne flaviviruses, the four serotypes of DENV (DENV1, DENV2, DENV3, and DENV4) are the leading causes of arboviral diseases in the tropical and subtropical areas (Green and Rothman, 2006; Gubler, 2002; Guzman and Kouri, 2002;
Halstead, 1988). The clinical presentations after DENV in-fection range from asymptomatic inin-fection, to a debilitating but self-limited disease, dengue fever (DF), and severe and po-tentially life-threatening diseases, dengue hemorrhagic fever/ dengue shock syndrome (DHF/DSS). It has been estimated that about 50–100 million cases of DF and 250–500 thousands cases of DHF occur annually worldwide (Green and Rothman, 2006; Gubler, 2002; Guzman and Kouri, 2002; Halstead, 1988). The genome of DENV is a positive-sense, single-stranded RNA of approximately 10.6 kb in length. Flanked by the 5′ and 3′ non-translated regions, the single open reading frame encodes a polyprotein precursor, which is cleaved by cellular and viral
Available online at www.sciencedirect.com
Virology 374 (2008) 338–350
www.elsevier.com/locate/yviro
⁎ Corresponding author. Institute of Microbiology, College of Medicine, National Taiwan University, No1 Sec1 Jen-Ai Rd, Taipei, Taiwan. Fax: +886 2 2391 5293.
E-mail address:[email protected](W.-K. Wang).
0042-6822/$ - see front matter © 2007 Elsevier Inc. All rights reserved. doi:10.1016/j.virol.2007.12.041
proteases into three structural proteins, capsid (C), precursor membrane (PrM), and envelope (E), and seven nonstructural proteins, NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5 (Lindenbach and Rice, 2001). After the interaction of E protein with its cellular receptor, DENV is believed to enter the cell through receptor mediated endocytosis (Lindenbach and Rice, 2001; Mukhopadhyay et al., 2005). Following translation and genome replication, assembly occurs in the membrane structures derived from endoplasmic reticulum (ER), where immature virions are transported through the secretary pathway ( Linden-bach and Rice, 2001; Mukhopadhyay et al., 2005; Stadler et al., 1997). Prior to release from the cells, PrM protein is cleaved to Pr and M proteins by cellular furin or furin-like protease to generate mature infectious virions (Lindenbach and Rice, 2001; Mukho-padhyay et al., 2005; Stadler et al., 1997).
The E protein of DENV, which forms head-to-tail homo-dimers arranged in a dense lattice on the surface of virion, is the major determinant of host range, tissue tropism and virulence, as well as the major target of neutralizing and enhancing antibodies (Bray et al., 1998; Green and Rothman, 2006; Halstead, 1988; Lindenbach and Rice, 2001; Mukhopadhyay et al., 2005; Zhang et al., 2003). The N-terminal ecotodomain of DENV2 E protein (amino acid residues 1 to 395), of which the crystallographic structure has been solved, contains three well characterized domains (Modis et al., 2003, 2004; Rey et al., 1995). Domain I, an eight-strandedβ-barrel, is located in the center. Domain II, an elongated finger-like-domain containing the internal fusion loop, is involved in the dimerization and membrane fusion. Domain III, an immunoglobulin-like domain, is believed to participate in the receptor binding. The C-terminal of E protein contains the stem and anchor regions (Fig. 1A) (Allison et al., 1999; Zhang et al., 2003). The stem region (residues 396 to 450) consists of twoα-helices (H1 and E-H2) separated by a short sequence element, which is highly conserved among flaviviruses. The anchor region (residues 452 to 495) is composed of two α-helical transmembrane (TM) segments (E-TM1 and E-TM2), which cross the two leaflets of the lipid bilayer (Allison et al., 1999; Zhang et al., 2003). Based on the studies of the tick-borne encephalitis virus (TBEV), E-TM2 is the signal sequence of NS1, and E-TM1 anchors the membrane and is required for incorporation of E protein into particles. E-H2 is involved in the stabilization of PrM–E heterodimer, whereas E-H1 is involved in the irreversible trimerization of soluble E protein in low pH environment (Allison et al., 1999; Stiasny et al., 1996).
A common feature during flavivirus replication is the formation of subviral particles, which are smaller and sediment slower than the mature virions (Lindenbach and Rice, 2001). Co-expression of PrM and E proteins in the absence of C protein is sufficient to generate recombinant virus-like particles (VLPs). VLPs are similar to the infectious virions in the structural and physicochemical features (Ferlenghi et al., 2001; Konishi and Fujii, 2002; Schalich et al., 1996), and have been shown to be useful non-infectious serodiagnostic antigens (Davis et al., 2001; Hunt et al., 2001; Purdy et al., 2004) and potential vaccine candidates (Chang et al., 2000, 2003; Davis et al., 2001; Konishi and Fujii, 2002; Kroeger and McMinn, 2002). VLPs of several
members of flaviviruses can be generated efficiently with few exceptions, such as DENV2 and St. Louis encephalitis virus. It was reported previously that inefficient production of DENV2 VLPs was due to intracellular accumulation of PrM/E proteins and that replacement of the stem–anchor of DENV2 with that of Japanese encephalitis virus (JEV) resulted in efficient secretion of VLPs (Chang et al., 2003; Purdy and Chang, 2005).
In this study, we investigated the mechanisms involved in the production of VLPs of DENV2. Chimeric DENV2 PrM/E proteins containing the stem or anchor of JEV alone failed to produce VLPs efficiently, indicating that both stem and anchor regions of JEV are required. Replacement of the TM and cytoplasmic (CY) domains of CD4 with the stem–anchor of DENV2 (CD4D2) or JEV (CD4JEV) rendered the chimeric CD4 retained predominantly in the ER as revealed by double-label immunofluorescence analysis and supported by the sensitivity to endoglycosidase H digestion and flow cytometry. These findings suggested that the stem–anchor of DENV2 contained a strong ER retention signal, which might account for the inefficient production of VLPs of DENV2.
Results
Both stem and anchor of JEV E protein are required for efficient production of VLPs of DENV2
It was reported previously that pCB-JssD2J396, which encoded the signal sequence of JEV, PrM protein of DENV2 and chimeric E protein containing the stem–anchor of JEV, produced VLPs more efficiently than pCB-JssD2, which en-coded the signal sequence of JEV and PrM/E proteins of DENV2, suggesting that the stem–anchor of JEV contributes to efficient VLP production (Fig. 1A) (Chang et al., 2003). To investigate whether this is also true in the context of DENV2 signal sequence, we generated two constructs, pCB-DssD2 and pCB-DssD2J396, and examined the production of VLPs. As shown inFig. 1B, Western blot analysis of lysates derived from pCB-DssD2- and pCB-DssD2J396-transfected cells revealed similar amounts of PrM and E proteins, suggesting that protein expression was comparable in both constructs. In contrast, greater amounts of PrM and E proteins were found in pellets derived from culture supernatants of pCB-DssD2J396-trans-fected cells than those of pCB-DssD2-transpCB-DssD2J396-trans-fected cells, indi-cating that the stem–anchor of JEV contributes to efficient VLP production in the context of DENV2 signal sequence (Fig. 1C). Of note, the amounts of E protein in pellets derived from culture supernatants relative to those in lysates of the pCB-JssD2-transfected cells were not higher than those in the pCB-DssD2-transfected cells, suggesting that the signal sequence of JEV does not enhance VLP production (Fig. 1B and C). Taken together, these findings suggest that the stem–anchor of JEV is required for efficient production of VLPs of DENV2.
To further determine whether the stem or the anchor of JEV alone is sufficient to produce VLP efficiently, we generated an-other two constructs, pCB-JssD2J396D442 and pCB-JssD2J442, which contained the stem and the anchor of JEV, respectively
Fig. 1B and C, the amounts of E protein in pellets derived from culture supernatants relative to those in cell lysates of the pCB-JssD2J396D442- or pCB-JssD2J442-transfected cells were less than those of the pCB-JssD2J396-transfected cells, though slightly greater than those of the pCB-JssD2-transfected cells. These findings indicate that both stem and anchor regions of JEV are required for efficient production of VLPs of DENV2. Stem–anchor of JEV does not enhance heterodimeric interaction between DENV2 PrM and E proteins
A previous study of TBEV has shown that good heterodimeric interaction between PrM and E proteins was required for pro-duction of VLPs (Allison et al., 1999). To examine the possibility
that stronger interaction between PrM and E proteins in pCB-DssD2J396 and pCB-JssD2J396 may account for efficient pro-duction of VLPs, we carried out a radioimmunoprecipitation assay with an anti-E monocloncal antibodies (Mab). As shown in
Fig. 2, the amounts of PrM protein relative to those of E protein in the constructs producing VLPs efficiently (pCB-DssD2J396 and pCB-JssD2J396) were not higher than those producing VLPs inefficiently (pCB-DssD2, pCB-JssD2, pCB-JssD2J396D442 and pCB-JssD2J442). The ratios of the intensity of PrM band to that of E band for pCB-DssD2J396 and pCB-JssD2J396 were 0.18 and 0.22, respectively, which were within the range of the ratios for other 4 constructs (0.12 to 0.56). Of note, inclusion of the fast migrating E protein bands observed in the lysates of pCB-DssD2-, pCB-JssD2- and pCB-JssD2J396D442-transfected cells, Fig. 1. Schematic drawing of the stem–anchor region of E protein, the constructs and expression of PrM/E proteins in cells and pellets. (A) The stem consists of two α-helices (H1, H2) and the anchor twoα-helical TM segments (TM1, TM2). Open bars indicate DENV2 sequences and black bars JEV sequences. ss, signal sequence. (B, C) Forty eight hours after transfection of 10μg of each plasmid to 293T cells, cell lysates (B) and pellets (C) derived from culture supernatants by 20% sucrose cushion ultracentrifugation were subjected to Western blot analysis using serum from a confirmed DENV2 case (Wang et al., 2006). Arrow heads indicate PrM and E proteins recognized by the DENV2 serum, which did not react with lysates of mock-transfected cells (data not shown). The size of molecular weight markers is shown in kDa. The ratio of the intensity of E protein band in pellets to that in cell lysates (E pellet/E cell) was shown below each lane in panel C. One representative experiment of more than three was shown.
which may represent deglycosylated or misfolded E proteins, in the calculation revealed that the ratios of PrM band to E band for pCB-DssD2J396 and pCB-JssD2J396 were not higher than those of other 4 constructs. A similar trend was also found with another anti-E Mab (data not shown). These findings suggest that efficient production of VLPs by pCB-DssD2J396 and pCB-JssD2J396 could not be attributed to a stronger heterodimeric interaction between PrM and E proteins.
Stem–anchor of JEV affects the intracellular localization of DENV2 E protein
To investigate the intracellular localization of DENV2 E and chimeric DENV2 E proteins, pCB-JssD2- and pCB-JssD2J396-transfected BHK-21 cells were examined by double-label immunofluorescence analysis using Mab specific to PDI (an ER resident protein) and mannosidase II (Man II) (a marker of Golgi apparatus). As shown inFig. 3, the E protein in the pCB-JssD2-transfected cells was found in the cytoplasm with a staining pattern distinct from that of Man II and merging near completely with that of PDI, suggesting that the majority of DENV2 E protein retained in the ER (Fig. 3A and C). The E protein in the pCB-JssD2J396-transfected cells was also found in the cytoplasm with a slightly different staining pattern, which merged mainly with that of PDI and partially with that of MAN II (Fig. 3B and D). This finding suggests that the intracellular localization of the chimeric DENV2 E protein, though mainly in the ER, is slightly different from that of the DENV2 E protein. We next examined the glycosylation pattern of the DENV2 E and chimeric DENV2 E proteins in cell lysates by treatment with endo-β-N-acetylglucosaminidase H (endo H) and peptide N-glycosidase F (PNGase F). As shown inFig. 3E, E protein in the lysates of pCB-JssD2-transfected cells was sensitive to endo H digestion, suggesting that intracellular E protein remained primarily in a compartment prior to trans-Golgi. In
contrast, the chimeric E protein in the lysates of pCB-JssD2J396-transfected cells contained both a major band sensitive to endo H and a faint band resistant to endo H, suggesting that the chimeric E protein was present not only in an endo H-sensitive compartment, most likely ER, but also in a compartment beyond trans-Golgi, though only in a small amount. Moreover, treatment of the lysates of pCB-DssD2- and pCB-DssD2J396-transfected cells with endo H and PNGase F revealed a similar glycosylation pattern (Fig. 3F). Taken together, these findings, consistent with those of the immuno-fluorescence analysis, suggest that the stem–anchor of JEV affects the intracellular localization and glycosylation pattern of DENV2 E protein.
To further investigate whether intracellular PrM/E proteins retained in the membrane-bound fraction or formed VLPs in the soluble fraction of ER or other compartments, pCB-JssD2- and pCB-JssD2J396-transfected 293T cells were subjected to a previously described subcellular fractionation experiment to obtain the membrane-bound and soluble fractions (Xu et al., 1997). As shown inFig. 4, greater amounts of PrM/E proteins were found in the membrane-bound fraction of the pCB-JssD2-transfected cells than in that of the pCB-JssD2J396-pCB-JssD2-transfected cells. In contrast, greater amounts of PrM/E proteins were found in the pellets of soluble fraction of pCB-JssD2J396-transfected cells than in those of pCB-JssD2-transfected cells. As a control, similar amounts of calnexin, an integral ER membrane protein, were found in the membrane-bound fractions of both pCB-JssD2- and pCB-JssD2J396-transfected cells but not in the soluble fractions. Together, these findings suggested that the majority of PrM/E proteins in pCB-JssD2-transfected cells retained in the membrane of ER based on the immunoflures-cence analysis, though a relatively small amount of VLPs did form in the soluble fraction, probably in the lumen of ER. Moreover, replacement of the stem–anchor of DENV2 with that of JEV resulted in the release of PrM/E proteins from the membrane-bound fraction to form VLPs in the soluble fraction, probably in the lumen of ER and beyond trans-Golgi in a small amount.
Stem–anchor of E protein contains an ER retention signal To further investigate whether the stem–anchor of DENV2 and JEV contains a signal for retention in the ER, we generated two chimeric CD4 constructs (CD4D2 and pCB-CD4JEV), of which the TM plus CY domains were replaced by the stem–anchor of DENV2 and that of JEV, respectively (Fig. 5A). After transfection into 293T cells, cells and lysates were examined by immunofluorescence assay and endoglyco-sidase digestion, respectively. As shown in Fig. 5B, CD4 was readily detected by an anti-CD4 Mab, RPA-T4, in the pCB-CD4-transfected cells in the presence or absence of Triton X-100, indicating that CD4 expressed on the surface of cells. In contrast, CD4D2 and CD4JEV can be detected well only in the presence of Triton X-100. In the absence of Triton X-100, CD4JEV could be detected very weakly in a few cells and CD4D2 was much less visible, suggesting that they did not express well on the surface of cells (Fig. 5B). Consistent with Fig. 2. Interaction between PrM and E proteins. 293T cells transfected with PrM/
E-expressing constructs shown inFig. 1Aor mock were labeled with35S-Met at
20 h post-transfection, washed, lysed, immunoprecipitated with anti-E Mabs FL0232 and subjected to 12% PAGE as described in the Materials and methods. Arrow heads indicate E and PrM proteins.
this, lysates from the pCB-CD4D2- and pCB-CD4JEV-transfected cells were sensitive to endo H digestion, suggesting that CD4D2 and CD4JEV retained mainly in a compartment prior to trans-Golgi (Fig. 5C). In contrast, treatment of lysates of pCB-CD4-transfected cells with endo H revealed two protein
bands decreased in size by about 2 kDa and 7 kDa as reported previously (Cocquerel et al., 1998, 1999; Shin et al., 1991). Since only one of the two N-linked glycosylation sites on CD4 was converted to complex oligosaccharide, the protein band decreased by 2 kDa presumably represented CD4 transported Fig. 3. Intracellular localization and glycosylation pattern of DENV2 E and chimeric DENV2 E proteins containing the stem–anchor of JEV by double-label immunofluorescence assay (A to D) and by endoglycosidase digestion (E, F). (A to D) BHK-21 cells were transfected with mock, pCB-JssD2 or pCB-JssD2 J396, spotted on slides 48 h later and fixed with acetone. One set of cells was stained with mouse anti-E Mab 4G2 (goat FITC-conjugated anti-mouse IgG as secondary antibody) and rabbit anti-PDI Pab (goat rhodamine-conjugated anti-rabbit IgG as secondary antibody) (A, B), and another set was stained with serum from a confirmed DENV2 case (Wang et al., 2006) (goat FITC-conjugated anti-human IgG as secondary antibody) and mouse anti-ManII Mab (goat rhodamine-conjugated anti-mouse IgG as secondary antibody) (C, D), followed by observation under a SP2 confocal laser scanning microscope. The Mab 4G2 and DENV2 serum were specific to DENV proteins and did not react with mock-transfected cells (data not shown). (E, F) Cell lysates derived from 293T cells transfected with JssD2, pCB-JssD2J396, pCB-DssD2 and pCB-DssD2J396 were treated with endo H (H) or PNGase F (F) and subjected to Western blot analysis by using serum from a confirmed DENV2 case (Wang et al., 2006). Arrow heads indicate glycosylated (E) and deglycosylated E protein (Edg), recognized by the DENV2 serum, which did not react with lysates of mock-transfected cells (data not shown). The size of molecular weight markers is shown in kDa. One representative experiment of three experiments was shown.
beyond trans-Golgi and was resistant to endo H, whereas the band decreased by 7 kDa represented CD4 remaining in the ER and was sensitive to endo H (Shin et al., 1991). Together, these findings suggest that the stem–anchor of DENV2 and JEV can retain CD4 in an intracellular compartment prior to trans-Golgi, probably in the ER.
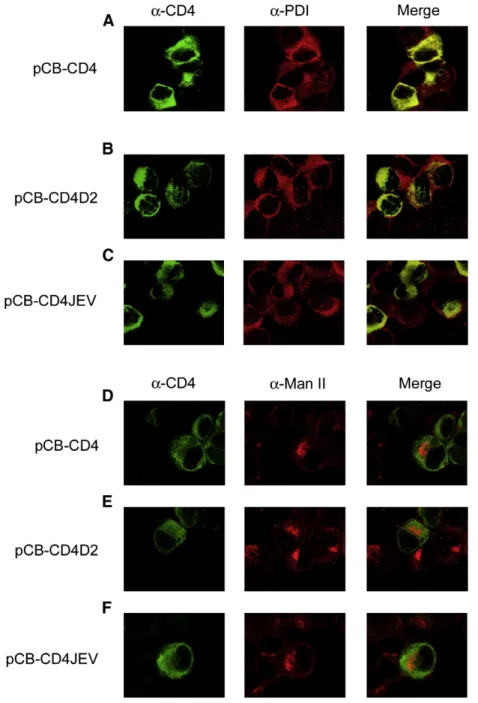
To further investigate this, double-label immunofluorescence analysis of pCB-CD4-, pCB-CD4D2- and pCB-CD4JEV-transfected BHK-21 cells was carried out by using Mab specific to protein markers of ER and Golgi. As shown inFig. 6, CD4 was stained by Mab RPA-T4 on the surface of cells as well as in the cytoplasm, which had a staining pattern distinct from that of Man II and merging partially with that of PDI, suggesting that after being synthesized in ER, CD4 transported beyond Golgi and expressed on the surface (Fig. 6A and D). In contrast, CD4D2 was stained mainly in the cytoplasm with a staining pattern different from that of Man II and merging almost completely with that of PDI, suggesting that CD4D2 retained primarily in the ER (Fig. 6B and E). Moreover, CD4JEV was stained mainly in the cytoplasm with a pattern similar to that of CD4D2 (Fig. 6C and F). Together, these findings suggested that the stem–anchor of DENV2 and JEV contained an ER retention signal. This was further supported by a similar staining pattern in 293T cells transfected with these three constructs (data not shown).
Of note, another conformation-sensitive Mab against CD4, Q4120 (Bijlmakers et al., 1997; Flint and McKeating, 1999), was found to bind to both CD4D2 and CD4JEV as good as
RPA-T4, suggesting that these two chimeric CD4 proteins do not affect the overall conformation of CD4 (data not shown). Moreover, immunofluorescence analysis by using Q4120 revealed a staining pattern similar to that by using RPA-T4 (data not shown). These findings suggest that the ER retention of CD4D2 and CD4JEV is unlikely due to misfolding of the chimeric proteins.
Fig. 5. CD4 and chimeric CD4 constructs, the surface expression and glyco-sylation pattern. (A) Schematic diagram of CD4 construct (pCB-CD4) and chimeric CD4 constructs containing the stem–anchor of DENV2 (pCB-CD4D2) or of JEV (pCB-CD4JEV). (B) 293T cells transfected with mock, pCB-CD4, pCB-CD4D2 or pCB-CD4JEV, were spotted on slide 48 h later, fixed with 4% paraformaldehyde, permeabilized with or without 0.5% Triton X-100, incubated with mouse anti-CD4 Mab RPA-T4 (goat FITC-conjugated anti-mouse IgG as secondary antibody), and observed under a fluorescence microscope. (C) Cell lysates derived from 293T cells transfected with the above constructs were treated with endo H (H) or PNGase F (F), and subjected to Western blot analysis by using Mab RPA-T4. Arrow heads indicate CD4 or chimeric CD4, and asters indicate the deglycosylated forms. The size of molecular weight markers is shown in kDa. The Mab RPA-T4 was specific to CD4 and did not react with mock-transfected cells and lysates (data not shown). One representative experiment of three experiments was shown.
Fig. 4. Subcellular fractionation experiment of DENV2 PrM/E and chimeric DENV2 PrM/E proteins containing the stem–anchor of JEV. 293T cells transfected with mock, pCB-JssD2, or pCB-JssD2J396 were washed with 1× PBS, resuspended in modified buffer B, and frozen–thawed 8 times (Xu et al., 1997). After clearing the nuclei and debris, the membrane-bound fraction and the pellets derived from the soluble fraction by 20% sucrose cushion ultracentrifugation were subjected to Western blot analysis by using serum from a confirmed DENV2 case (upper panel) (Wang et al., 2006) and then reprobing with anti-calnexin Mab (lower panel). Arrow heads indicate PrM, E and calnexin. The size of molecular weight markers is shown in kDa. One representative experiment of three experiments was shown.
A stronger ER retention signal in stem–anchor of DENV2 than in that of JEV
A closer examination of the immunofluorescence assay in
Fig. 5B revealed that in the absence of Triton X-100, faint staining signals detected by anti-CD4 Mab were observed in some of the CD4JEV-transfected cells, but not in the pCB-CD4D2-transfected cells. This suggests that the stem–anchor of JEV may contain a weaker ER retention signal than that of DENV2 (Fig. 5B). To further investigate this, CD4-,
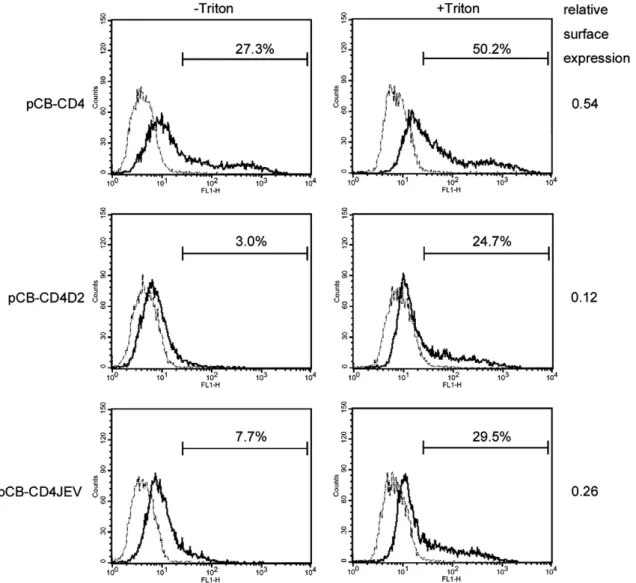
pCB-CD4D2- and pCB-CD4JEV-transfected cells were subjected to flow cytometric analysis in the presence or absence of Triton X-100. As shown inFig. 7, in the absence of Triton X-100, the percentage of positive cells stained with anti-CD4 Mab, compared with those with isotype control, was higher in cells transfected with pCB-CD4 (27.3%) than those transfected with pCB-CD4D2 (3.0%) or pCB-CD4JEV (7.7%), suggesting that more CD4 expressed on the surface than CD4D2 or CD4JEV. To adjust the expression level of each construct, we calculated the relative surface expression of CD4, which was the ratio of Fig. 6. Intracellular localization of CD4 and chimeric CD4 proteins containing the stem–anchor of DENV2 or JEV by double-label immunofluorescence assay. BHK-21 cells were transfected with mock, pCB-CD4, pCB-CD4D2 or pCB-CD4JEV, spotted on slides 48 h later and fixed with acetone. One set of cells was stained with mouse anti-CD4 Mab RPA-T4 (goat FITC-conjugated anti-mouse IgG as secondary antibody) and rabbit anti-PDI Pab (goat rhodamine-conjugated anti-rabbit IgG as secondary antibody) (A, B, C), and another set was stained with rabbit anti-CD4 Pab H-370 (goat FITC-conjugated anti-rabbit IgG as secondary antibody) and mouse anti-ManII Mab (goat rhodamine-conjugated anti-mouse IgG as secondary antibody) (D, E, F), followed by observation under a SP2 confocal laser scanning microscope. The Mab RPA-T4 and Pab H-370 were specific to CD4 and did not react with mock-transfected cells (data not shown). One representative experiment of three experiments was shown.
the percentage of CD4 positive cells in the absence of Triton X-100 to that in the presence of Triton X-X-100 and was 0.54, suggesting that approximately 54% of CD4 expressed on the surface in pCB-CD4-transfected cells. In contrast, the relative surface expression of CD4D2 and CD4JEV were 0.12 and 0.26, respectively, suggesting that approximately 12% and 26% of chimeric CD4 expressed on the surface in the pCB-CD4- and pCB-CD4-JEV-transfected cells, respectively. Of note was that the relative surface expression of CD4D2 was lower than that of CD4JEV, suggesting that the stem–anchor of DENV2 contains a stronger ER retention signal than that of JEV.
Capsid protein enhances VLP production of DENV2
The strong ER retention signal in the stem–anchor of DENV2 E protein and the inefficient VLP production raise a question how DENV2 overcomes this retention and produces
virus particles efficiently during its replication. One possibility is that RNA genome and viral proteins other than PrM and E, such as another structural protein, C protein, could facilitate particle formation. To examine the effect of C protein on VLP production, we constructed a plasmid expressing DENV2 C protein, pCDNA3-HAD2C (Fig. 8A), and examined VLP pro-duction by pCB-JssD2 in the presence or absence of C protein. As shown inFig. 8B, the amounts of PrM/E proteins in pellets derived from culture supernatants relative to those in lysates from the cells co-transfected with pCB-JssD2 and pCDNA3-HAD2C were greater than those from the cells transfected with pCB-JssD2 alone, indicating that C protein can enhance the VLP production by pCB-JssD2. Of note, C protein did not greatly enhance the VLP production by pCB-JssD2J396, which produced VLP efficiently by itself. This was further supported by quantification of the intensity of E protein band in pellets and cell lysates and calculation of the ratio (E pellet/E cell). As Fig. 7. Surface expression of CD4 and chimeric CD4 proteins containing the stem–anchor of DENV2 or of JEV by flow cytometry. 293T cells were transfected with mock, pCB-CD4, pCB-CD4D2 or pCB-CD4JEV, fixed with 4% paraformaldehyde, permeabilized with or without 0.5% Triton X-100, incubated with mouse anti-CD4 Mab RPA-T4 or isotype control (goat FITC-conjugated anti-mouse IgG as secondary antibody), resuspended in buffer containing 1% paraformaldehyde and subjected to FASCalibur and CellQuest analysis. Solid lines indicate anti-CD4 Mab. Dot lines indicate isotype control, which overlapped with those of mock-transfected cells (data not shown). The relative surface expression of CD4 or chimeric CD4 was the ratio of the percentage of RPA-T4-positive cells in the absence of Triton X-100 to that in the presence of Triton X-100. One representative experiment of three experiments was shown.
shown in Fig. 8C, the fold increase of E pellet/E cell in the presence of C protein was 2.2 for JssD2 and 1.3 for pCB-JssD2J396. Similarly, the VLP production by pCB-DssD2, but not by pCB-DssD2396, was also enhanced by co-transfection with pCDNA3-HAD2C (data not shown). Together, these findings suggest that C protein can enhance VLP production by DENV2 PrM/E proteins, which contain a strong ER retention signal, and may facilitate efficient production of DENV2 virus particles during replication.
Discussion
Because of the similarities in both structural and biochemical properties between VLPs and infectious virions, studying the mechanisms of VLP formation would provide important information not only to our understanding of the functions of PrM/E proteins but also to identification of potential anti-viral targets (Ferlenghi et al., 2001; Konishi and Fujii, 2002; Purdy and Chang, 2005; Schalich et al., 1996). Previous studies of
TBEV and West Nile virus (WNV) have shown the involvement of the stem region in the PrM–E interaction and the contribution of this heterodimeric interaction and N-linked glycosylation to the production of VLPs (Allison et al., 1999; Hanna et al., 2005). In this study, we generated a series of chimeric PrM/E and chimeric CD4 constructs to investigate the mechanisms involved in the VLP production by DENV2 PrM/E proteins and demonstrated that the stem–anchor of DENV2 E protein contained a strong ER retention signal, which might contribute to inefficient VLP production. To our knowledge, this is the first report that a strong ER retention signal in the flaviviral E protein may affect the production of VLPs.
Since the morphogenesis of flaviviral particles occurs in the membranous structures derived from ER, an ER localization signal that linked to the processes of genome packaging, virion assembly and budding during the replication cycle was thought to be present in the E protein of flaviviruses (Ishak et al., 1988; Lindenbach and Rice, 2001; Mackenzie and Westaway, 2001; Mukhopadhyay et al., 2005; Wang et al., 1997). This notion was supported by the recent reports that the TM domain of E protein of yellow fever virus (YFV) and those of E1 and E2 proteins of hepatitis C virus (HCV) contained an ER retention signal (Cocquerel et al., 1998, 1999; Op De Beeck et al., 2004). In agreement with this, chimeric CD4 proteins containing the stem–anchor of DENV2 (CD4D2) or of JEV (CD4JEV) retained in the ER, indicating that the stem–anchors of both DENV2 and JEV contained an ER retention signal (Figs. 5 and 6). Inter-estingly, the observations that the relative surface expression of CD4D2 was lower than that of CD4JEV (Fig. 7) and that faint surface staining was found in some of the CD4JEV-expressing cells but in none of the CD4D2-expressing cells (Fig. 5B) sug-gested that the stem–anchor of DENV2 contained a stronger ER retention signal than that of JEV. Consistent with this, E protein in the pCB-JssD2-transfected cells merged almost completely with PDI and retained mainly in an endo H-sensitive compart-ment, whereas the chimeric E protein in the pCB-JssD2J396-transfected cells merged partially with Man II (Fig. 3D), and were present not only in the endo H-sensitive but also in the endo H-resistant compartments (Fig. 3E). These findings, together with the observation that less amounts of VLPs were produced by pCB-JssD2 than those by pCB-JssD2J396 (Fig. 1), suggest that the stronger ER retention signal in the stem–anchor of DENV2 than that of JEV might account for the inefficient VLP production of DENV2.
Several specific signals for the retention and retrieval of proteins in ER have been identified. For soluble ER resident proteins, a stretch of amino acids consisting of“KDEL” was known to be recognized by the specific receptor for retrieval from Golgi to ER (Michelsen et al., 2005; Nilsson and Warren, 1994; Pelham, 1995). For TM proteins in the ER, ER retention and retrieval can be mediated through intracytoplasmic sequences. For examples, a di-lysine motif“K(X)KXX” at the C-terminal of the CY tail and an arginine-based motif “ΦΨ/ RRXR” at the N- or C-terminal of the CY domain have been identified (Michelsen et al., 2005; Nilsson and Warren, 1994; Pelham, 1995). In addition, several TM proteins retained in the ER based on the determinants present in the TM domains, Fig. 8. Production of VLPs by co-expressing PrM/E and C proteins. (A) The
construct, pCDNA3-HAD2C, expresses the mature C protein (residue 1 to 101) of DENV2 with HA tag at its N-terminal. (B) 293T cells were transfected with pCB-JssD2 or pCB-JssD2J396 alone or in the presence of pCDNA3-HAD2C. Forty eight hours later, cell lysates and pellets derived from culture supernatants by 20% sucrose cushion ultracentrifugation were subjected to Western blot analysis using mixture of anti-HA Mab and serum from a confirmed DENV2 case (Wang et al., 2006). Arrow heads indicate C, PrM and E proteins. The size of molecular weight markers is shown in kDa. One representative experiment of more than three experiments was shown. (C) The ratio of the intensity of E protein band in pellets to that in cell lysates (E pellet/E cell) was determined for each transfection. The fold increase was the ratio of E pellet/E cell in the presence of C protein to E pellet/E cell in the absence of C protein.
which may interact with membrane or other membrane proteins (Cocquerel et al., 1998; Op De Beeck et al., 2004; Szczesna-Skorupa and Kemper, 2000; Yang et al., 1997). Examination of the amino acid sequences of the stem–anchor of DENV2 and JEV revealed no above described ER retention or retrieval motif. Moreover, cryoelectron microscopic study of DENV2 virion at high resolution has shown that the linking residues between the TM1 and TM2 α-helices of E protein were associated with the phospholipid polar head groups of the inner leaflet of membrane, suggesting the lack of CY domain of E protein (Zhang et al., 2003). Therefore, the ER retention by the stem–anchor of DENV2 and JEV was most likely due to their intrinsic properties interacting with membrane or other membrane proteins as has been proposed for ER retention, but not retrieval, by the TM domains of E proteins of other two flaviviruses (Cocquerel et al., 1998, 1999; Duvet et al., 1998; Op De Beeck et al., 2004). Interestingly, substitutions of the charged residues between the TM1 and TM2 of HCV E2 protein were shown to alter the subcellular localization of E protein, whereas similar substitution of the arginine residue between TM1 and TM2 of YFV E protein did not, suggesting different mechanisms mediated by different TM domains of flaviviruses (Cocquerel et al., 2000; Op De Beeck et al., 2003).
By performing subcellular fractionation experiment, we showed that the majority of DENV2 PrM/E proteins retained in ER was in the membrane fraction and a relatively small amount of PrM/E proteins was in the pellets of the soluble fraction, suggesting that VLPs did form in the lumen of ER (Fig. 4). This finding resonated with a recent electron microscopic (EM) study of VLPs of TBEV, in which VLPs were found most frequently in the lumen of ER and less frequently in the smooth ER, ER-to-Golgi intermediate, trans-Golgi network and secre-tory vesicles (Lorenz et al., 2003). Interestingly, replacement of the stem–anchor of DENV2 with that of JEV resulted in a shift of PrM/E proteins from the membrane fraction to the pellets of the soluble fraction and to the supernatants, suggesting that the stem–anchor of JEV could enhance the formation of VLPs of DENV2 (Fig. 4). Although the budding of flaviviral particles or VLPs from the membrane to the lumen of ER was thought to be a very rapid process due to the lack of budding intermediates clearly observed thus far (Ishak et al., 1988; Lindenbach and Rice, 2001; Mackenzie and Westaway, 2001; Wang et al., 1997), studying the DENV2- and chimeric DENV2 PrM/E-expressing cells by EM may provide morphological details how the stem–anchor of JEV enhance the formation of VLPs. Moreover, further experiments to define the minimal domain required for ER retention and to elucidate the relationship between the property of ER retention and efficiency of VLP formation would be critical. In this regard, our findings that both stem and anchor regions of JEV were required for efficient VLP production (Fig. 1) were consistent with the previous report that amino acid substitutions of three critical residues (I398L, M401A and M412L) at the stem of DENV2 plus the anchor of JEV can enhance the VLP production by pCB-JssD2 to near 50% of that by pCB-JssD2J396, which contained both stem and anchor of JEV (Purdy and Chang, 2005). Analysis of the secondary structure in the stems of DENV2 and JEV suggested
that the increase in hydrophobicity and flexibility of E-H1 in the “hole” region near the E-dimers by these three substitutions may result in proper burial of these residues within the lipid mem-brane and optimal intra- or intermolecular interactions between PrM–E and E–E proteins, which might influence the curving and bending of lipid membrane and lead to the assembly and release of VLPs (Purdy and Chang, 2005).
The strong tendency of DENV2 PrM/E proteins to retain in the ER and produce VLPs inefficiently raises an important biological question, namely, how DENV2 produces virus particles efficiently during its replication. By co-expressing C protein and DENV2 PrM/E proteins, we demonstrated that C protein can enhance VLP production of DENV2. Although the increase in VLP production by providing C protein to pCB-DssD2 or pCB-JssD2 was not as good as that by providing the stem–anchor of JEV through chimeric constructs, pCB-DssD2J396 or pCB-JssD2J396, our findings that C protein has a role in the VLP production would suggest that the assembly and budding of virion during virus replication is a more complex and tightly regulated process than that of VLP production and that different viral or cellular factors, such as RNA genome and other viral proteins in the vicinity, could also play important roles in the whole process (Kummerer and Rice, 2002; Liu et al., 2003, 2002).
Materials and methods Plasmid constructs
Plasmid pCB8D2-2J-2-9-1, which encoded the signal se-quence of JEV, PrM protein of DENV2 and chimeric E protein with the N-terminal 395 residues from DENV2 and C-terminal 100 residues from JEV, was designated as pCB-JssD2J396 in this study (Fig. 1A) (Chang et al., 2003). RNA extracted from culture supernatants of DENV2 (16681 strain)-infected C6/36 cells was subjected to RT using random primers to generate cDNA template as described previously (Lin et al., 2004). PCR product containing the entire PrM/E gene and 3′ region of C gene (corresponding to amino acid residues 101 to 114, which encode the signal sequence) was amplified by using the primer pair (d2KpnSS, 5′-CTTGGTACCTCTAGAGCCGCCGC-CATGGC AGGCATGATCATTATG-3′ [genome positions 389 to 410] and d2NotI-2402, 5′-TTTCTCGAGC GGCCGCTC-AACTAATTAGGCCTGCACCATGACTC-3′ [position 2425 to 2397]) (Irie et al., 1989) and cDNA template, digested with KpnI and NotI, and cloned into respective sites of pCB-Jss D2J396 to generate pCB-DssD2. To construct pCB-DssD2J396, the Bsu36I–NotI fragment cut from pCB-JssD2J396 was cloned into respective sites of pCB-DssD2. To construct pCB-JssD2, PCR product containing PrM/E gene was amplified by using the primer pair (d2KasI-438, 5′-TGT GCAGGCGCCTTCCATT-TAACCACACGTAACG-3′ [positions 426 to 453], and d2NotI-2402) (Irie et al., 1989) and cDNA template, digested with KasI and NotI, and cloned into respective sites of pCB-JssD2J396. To construct pCB-JssD2J442, the BstXI–NotI fragment cut from pCB-JssD2J396 was cloned into respective sites of pCB-JssD2. To construct pCB-JssD2J396D442, the BstXI–NotI fragment
cut from JssD2 was cloned into respective sites of pCB-JssD2J396 (Fig. 1A). All the constructs were confirmed by sequencing the entire PrM/E region.
To construct pCB-CD4, PCR product containing CD4 was amplified by using the primer pair (KpnI CD41A, 5 ′-CTTGGTACCATGAATCGGGGAGTCCCT TTTAGG-3′ and
NotI CD4-435B, 5
′-ACGTCAGCGGCCGCTCAAA-TGGGGCTACATGTC-3′) and T4-pMV7 as template (Maddon et al., 1986), digested with KpnI and NotI, and cloned into respective sites of pCB-JssD2J396 (Fig. 5A). To construct pCB-CD4D2, PCR product containing region encoding stem–anchor of DENV2 E protein was amplified by using the primer pair (d2E395Bsu36I-A, 5′-GTT GCACCTGAGGGAAGTTC-TATCGGCCAAA-3′ [positions 2111 to 2130], and D2NotI-2402) (Irie et al., 1989) and pCB-JssD2 as template, digested with Bsu36I and NotI, and cloned into respective sites of pCB-CD4. To construct pCB-CD4JEV, PCR product containing region encoding stem–anchor of JEV E protein was amplified by using the primer pair (JEV400Bsu36IA, 5 ′-GTTGCACCT-GAGGGAAGCACGCTGGGCAAG-3′; JEVNotI495B, 5′-ACGTCAGCGGCC GCTCAAACTAATTAAGCATGCA-CATTGGTCGC-3′) and pCDJE2-7, a plasmid expressing JEV PrM/E proteins, as template (Chang et al., 2000), digested with Bsu36I and NotI, and cloned into respective sites of pCB-CD4. All constructs were confirmed by sequencing the insert.
To construct pCDNA3-HAD2C, PCR fragment containing region encoding mature C protein (residues 1 to 101) was am-plified by using the primer pair (d2C1ABamHI, 5′-CGGGATC-CATGA ATAACCAACGGAAAAAG-3′ [positions 89 to 110], and d2C100B XhoI, 5′-GTGGCTCGAG TCATCTGCG-TCTCCTATTCAA GATG-3′ [positions 389 to 367]) (Irie et al., 1989) and cDNA template derived from DENV2 16681 strain as described above, digested with BamHI and XhoI, and cloned into respective sites of pCDNA3-HAtag (Invitrogen, Carlsbad, CA). The construct was confirmed by sequencing the insert.
Cell lysates and recombinant VLPs
293T cells were prepared in a 10 cm-culture dish at 1 × 105 cells per dish one day earlier and transfected with 10 μg of plasmid DNA by calcium phosphate method (Chen and Okayama, 1987). At 48 h post-transfection, culture supernatants were collected for further process; and cells were washed with 1× PBS and treated with 1% NP40 lysis buffer (100 mM Tris [pH 7.5], 150 mM NaCl, 20 mM EDTA, 1% NP40, 0.5% Na deoxycholate) containing protease inhibitors (Roche Diagnos-tics, Indianapolis, IN), followed by centrifugation at 14,000 rpm (851 rotor, Thermo Electron Corporation, Waltham, MA) at 4 °C for 30 min to obtain cell lystates. Culture supernatants were clarified by centrifugation at 2,500 rpm (A-4-62 rotor, Eppendorf, Westbury, NY) for 20 min, filtered through a 0.22 μm pore-sized membrane (Sartorius, Goettingen, Ger-many), layered over a 20% sucrose buffer, and ultracentrifuged at 19,000 rpm (SW28 rotor, Beckman, Fullerton, CA) at 4 °C for 5 h. The pellets were resuspended in 30 °l TNE buffer (50 mM Tris [pH 8.0], 1 M NaCl, 100 mM EDTA) and subjected to Western blot analysis.
Antibodies
Human DENV sera that can recognize E and PrM proteins were obtained from confirmed DENV2 cases as described previously (Wang et al., 2006). Anti-CD4 mouse Mab RPA-T4 (IgG1,κ) and mouse isotype (IgG1, κ) control were purchased from eBioscience (San Diego, CA), anti-CD4 mouse Mab Q4120 from Sigma (St. Louis, MO), anti-CD4 rabbit polyclonal antibody (Pab) H-370 and anti-calnexin mouse Mab E-10 from Santa Cruz biotechnology Inc. (Santa Cruz, CA), anti-Man II mouse Mab 53FC3 from Covance (Berkely, CA), anti-PDI rabbit Pab SPA-890 from Stressgen Bioreagents (Ann Arbor, MI), and HA Mab from KPL (Guildford, UK). Mouse anti-E Mab FL0232 was purchased from Chance Biotechnology (Taipei, Taiwan) and 4G2 was from American Type Culture Collection (Rockville, MD) (Henchal et al., 1982).
Western blot analysis
Cell lysates or pellets from culture supernatants were subjected to 12% polyacrylamide gel electrophoresis (PAGE), followed by transferring to nitrocellulose membrane (Hybond-C; Amersham Biosciences, HK) with an electrotransfer apparatus (Bio-Rad, Hercules, CA). Membranes were blocked with 4% milk in wash buffer and incubated with primary (human sera or Mab) and secondary antibodies (horseradish peroxidase-conjugated anti-human or anti-mouse IgG) (Pierce, Rockford, IL) each at 37 °C for 1 h. After final washing, the signals were detected by enhanced chemiluminescence reagents (Perkin Elmer life sciences, Boston, MA) (Wang et al., 2006). The intensities of E and PrM bands were further analyzed by UVP Biochemi Image system and the software UVP lab. Work 4.5 (Upland, CA), and the ratio of the intensity of E protein band in pellets to that in cell lysates was determined. Subcellular fractionation
293T cells (1 × 105cells) were transfected with mock, pCB-JssD2, or pCBJssD2J396, washed 3 times with 1× PBS 48 h later, resuspended in modified buffer B (10% sucrose, 20 mM Tris, 150 mM NaCl, 10 mM Mg acetate, 1 mM EGTA [pH7.6]), and frozen–thawed 8 times as described previously (Xu et al., 1997). After clearing the nuclei and debris by centrifugation at 14,000 rpm (851 rotor, Thermo Electron Corporation, Waltham, MA) for 5 s, the membrane fraction was pelleted by cen-trifugation at 14,000 rpm for 30 min at 4 °C and subjected to Western blot analysis. The resulting supernatants were layered over a 20% sucrose buffer and ultracentrifuged at 45,000 rpm (SW55 rotor, Beckman, Fullerton, CA) at 4 °C for 1 h to obtain the pellets of soluble fraction, which were resuspended in 30μl TNE buffer and subjected to Western blot analysis.
Endoglycosidase digestion
Aliquots from cell lysates or pellets were treated with 500 U of endo H or PNGase F at 37 °C for 1 h according to the manufacturer's instructions (New England Biolabs., Beverly, MA), and subjected to Western blot analysis.
Radioimmunoprecipitation
293T cells prepared in 6-well plate were transfected with plasmid DNA by calcium phosphate method (Chen and Okayama, 1987). At 20 h post-transfection, cells were washed with 1× PBS, incubated with Met-free DMEM at 37 °C for 2 h, labeled with 30μCi35S-Met (Amersham Biosciences, HK) at 37 °C for 6 h. Cells were washed with 1× PBS twice, lysed and centrifuged as described above to obtain cell lysates. Following pre-clear with beads, cell lysates were incubated with anti-DENV E Mab at 4 °C overnight and then with protein A sepharose beads (Amersham Biosciences, HK) at 4 °C for 6 h. After washing with 1% NP40 buffer 4 times, the beads were mixed with 15μl 2× sample buffer (500 mM Tris [pH6.8], 4% SDS, 20% glycerol, 0.4% bromphenol blue), heated at 95 °C for 3 min, and centrifuged at 13,000 rpm for 5 min. The solubilized fraction was subjected to 12% PAGE.
Indirect immunofluorescence assay
Plasmid DNA was transfected to 293T cells prepared in 6-well plate by calcium phosphate method or to BHK-21 cells by lipofectamine 2000 (Invitrogen, Carsblad, CA). At 48 h post-transfection, cells were resuspended in 1× PBS, spotted onto 10-well Teflon-coated slide, air-dried and fixed with 4% paraf-ormaldehyde at room temperature for 30 min (Liu et al., 2005). Cells were permeabilized with or without 0.5% Triton X-100 at room temperature for 30 min, incubated with primary and secondary antibody each at 37 °C for 1 h. After final washing, drying and mounting, the slides were observed under a SP2 confocal laser scanning microscope (Leica) or fluorescence microscope.
Flow cytometry
293T cells prepared in 10 cm-culture dish were transfected with mock, pCB-CD4, pCB-CD4D2 or pCB-CD4JEV. At 48 h post-transfection, cells were washed with 1× PBS, resuspended in 1× PBS and treated with 2% paraformaldehyde at 4 °C for 1 h. Cells were then permeabilized with or without 0.5% Triton X-100 at room temperature for 30 min, incubated with primary antibody (anti-CD4 Mab RPA-T4 or mouse isotype control) and secondary antibody (FITC-conjugated anti-mouse IgG) each at 4 °C for 1 h. Following the final wash, cells were resuspended in buffer containing 1% paraformaldehyde and subjected to FASCalibur (BD Bioscience, San Jose, CA) and CellQuest analysis.
Acknowledgments
We thank Dr. L. K. Chen at the Tzu-Chi University Taiwan for kindly providing Mab FL0232, Dr. Richard Axel for providing T4-pMV7 through the NIH AIDS Research and Reference Reagent Program, Division of AIDS, NIAID, NIH, and Su-Ru Lin for technical assistance. This work was supported by the National Science Council Taiwan (NSC96-3112-B-002-038).
References
Allison, S.L., Stiasny, K., Stadler, K., Mandl, C.W., Heinz, F.X., 1999. Mapping of functional elements in the stem–anchor region of tick-borne encephalitis virus envelope protein. E.J. Virol 73, 5605–5612.
Bijlmakers, M.J., Isobe-Nakamura, M., Ruddock, L.J., Marsh, M., 1997. Intrinsic signals in the unique domain target p56lck to the plasma membrane independently of CD4. J. Cell Biol 137, 1029–1040.
Bray, M., Men, R., Tokimatsu, I., Lai, C.J., 1998. Genetic determinants responsible for acquisition of dengue type 2 virus mouse neurovirulence. J. Virol. 72, 1647–1651.
Chang, G.J., Hunt, A.R., Davis, B., 2000. A single intramuscular injection of recombinant plasmid DNA induces protective immunity and prevents Japanese encephalitis in mice. J. Virol 74, 4244–4252.
Chang, G.J., Hunt, A.R., Holmes, D.A., Springfield, T., Chiueh, T.S., Roehrig, J.T., Gubler, D.J., 2003. Enhancing biosynthesis and secretion of premembrane and envelope proteins by the chimeric plasmid of dengue virus type 2 and Japanese encephalitis virus. Virology 306, 170–180.
Chen, C., Okayama, H., 1987. High-efficiency transformation of mammalian cells by plasmid DNA. Mol. Cell Biol. 7, 2745–2752.
Cocquerel, L., Meunier, J.C., Pillez, A., Wychowski, C., Dubuisson, J., 1998. A retention signal necessary and sufficient for endoplasmic reticulum localization maps to the transmembrane domain of hepatitis C virus glycoprotein E2. J. Virol 72, 2183–2191.
Cocquerel, L., Duvet, S., Meunier, J.C., Pillez, A., Cacan, R., Wychowski, C., Dubuisson, J., 1999. The transmembrane domain of hepatitis C virus glycoprotein E1 is a signal for static retention in the endoplasmic reticulum. J. Virol 73, 2641–2649.
Cocquerel, L., Wychowski, C., Minner, F., Penin, F., Dubuisson, J., 2000. Charged residues in the transmembrane domains of hepatitis C virus glycoproteins play a major role in the processing, subcellular localization, and assembly of these envelope proteins. J. Virol 74, 3623–3633. Davis, B.S., Chang, G.J., Cropp, B., Roehrig, J.T., Martin, D.A., Mitchell, C.J.,
Bowen, R., Bunning, M.L., 2001. West Nile virus recombinant DNA vaccine protects mouse and horse from virus challenge and expresses in vitro a noninfectious recombinant antigen that can be used in enzyme-linked immunosorbent assays. J.Virol 75, 4040–4047.
Duvet, S., Cocquerel, L., Pillez, A., Cacan, R., Verbert, A., Moradpour, D., Wychowski, C., Dubuisson, J., 1998. Hepatitis C virus glycoprotein complex localization in the endoplasmic reticulum involves a determinant for retention and not retrieval. J. Biol. Chem 273, 32088–32095. Ferlenghi, I., Clarke, M., Ruttan, T., Allison, S.L., Schalich, J., Heinz, F.X.,
Harrison, S.C., Rey, F.A., Fuller, S.D., 2001. Molecular organization of a recombinant subviral particle from tick-borne encephalitis virus. Mol. Cell 7, 593–602.
Flint, M., McKeating, J.A., 1999. The C-terminal region of the hepatitis C virus E1 glycoprotein confers localization within the endoplasmic reticulum. J. Gen. Virol. 80 (Pt 8), 1943–1947.
Green, S., Rothman, A., 2006. Immunopathological mechanisms in dengue and dengue hemorrhagic fever. Curr. Opin. Infect. Dis 19, 429–436.
Gubler, D.J., 2002. Epidemic dengue/dengue hemorrhagic fever as a public health, social and economic problem in the 21st century. Trends Microbiol 10, 100–103.
Guzman, M.G., Kouri, G., 2002. Dengue: an update. Lancet Infect.Dis 2, 33–42. Halstead, S.B., 1988. Pathogenesis of dengue: challenges to molecular biology.
Science 239, 476–481.
Hanna, S.L., Pierson, T.C., Sanchez, M.D., Ahmed, A.A., Murtadha, M.M., Doms, R.W., 2005. N-linked glycosylation of West Nile virus envelope proteins influences particle assembly and infectivity. J. Virol 79, 13262–13274.
Henchal, E.A., Gentry, M.K., McCown, J.M., Brandt, W.E., 1982. Dengue virus-specific and flavivirus group determinants identified with monoclonal antibodies by indirect immunofluorescence. Am. J. Trop. Med. Hyg 31, 830–836.
Hunt, A.R., Cropp, C.B., Chang, G.J., 2001. A recombinant particulate antigen of Japanese encephalitis virus produced in stably-transformed cells is an effective noninfectious antigen and subunit immunogen. J. Virol. Methods 97, 133–149.
Irie, K., Mohan, P.M., Sasaguri, Y., Putnak, R., Padmanabhan, R., 1989. Sequence analysis of cloned dengue virus type 2 genome New Guinea-C strain. Gene 75, 197–211.
Ishak, R., Tovey, D.G., Howard, C.R., 1988. Morphogenesis of yellow fever virus 17D in infected cell cultures. J. Gen. Virol. 69 (Pt 2), 325–335. Konishi, E., Fujii, A., 2002. Dengue type 2 virus subviral extracellular particles
produced by a stably transfected mammalian cell line and their evaluation for a subunit vaccine. Vaccine 20, 1058–1067.
Kroeger, M.A., McMinn, P.C., 2002. Murray Valley encephalitis virus recombinant subviral particles protect mice from lethal challenge with virulent wild-type virus. Arch. Virol 147, 1155–1172.
Kummerer, B.M., Rice, C.M., 2002. Mutations in the yellow fever virus nonstructural protein NS2A selectively block production of infectious particles. J. Virol 76, 4773–4784.
Lin, S.R., Hsieh, S.C., Yueh, Y.Y., Lin, T.H., Chao, D.Y., Chen, W.J., King, C.C., Wang, W.K., 2004. Study of sequence variation of dengue type 3 virus in naturally infected mosquitoes and human hosts: implications for transmission and evolution. J. Virol 78, 12717–12721.
Lindenbach, B.D., Rice, C.M., 2001. Flaviviridae: the viruses and their replication. In: Knipe, D.M., Howley, P.M., Griffin, D.E. (Eds.), Fields Virology. Philadelphia, Lippincott William and Wilkins, pp. 991–1041. Liu, I.J., Chen, P.J., Yeh, S.H., Chiang, Y.P., Huang, L.M., Chang, M.F., Chen,
S.Y., Yang, P.C., Chang, S.C., Wang, W.K., 2005. Immunofluorescence assay for detection of the nucleocapsid antigen of the severe acute respiratory syndrome SARS-associated coronavirus in cells derived from throat wash samples of patients with SARS. J. Clin. Microbiol. 43, 2444–2448.
Liu, W.J., Chen, H.B., Khromykh, A.A., 2003. Molecular and functional analyses of Kunjin virus infectious cDNA clones demonstrate the essential roles for NS2A in virus assembly and for a nonconservative residue in NS3 in RNA replication. J. Virol 77, 7804–7813.
Liu, W.J., Sedlak, P.L., Kondratieva, N., Khromykh, A.A., 2002. Complemen-tation analysis of the flavivirus Kunjin NS3 and NS5 proteins defines the minimal regions essential for formation of a replication complex and shows a requirement of NS3 in cis for virus assembly. J. Virol 76, 10766–10775. Lorenz, I.C., Kartenbeck, J., Mezzacasa, A., Allison, S.L., Heinz, F.X., Helenius, A., 2003. Intracellular assembly and secretion of recombinant subviral particles from tick-borne encephalitis virus. J. Virol 77, 4370–4382. Mackenzie, J.M., Westaway, E.G., 2001. Assembly and maturation of the flavivirus Kunjin virus appear to occur in the rough endoplasmic reticulum and along the secretory pathway, respectively. J. Virol 75, 10787–10799. Maddon, P.J., Dalgleish, A.G., McDougal, J.S., Clapham, P.R., Weiss, R.A.,
Axel, R., 1986. The T4 gene encodes the AIDS virus receptor and is expressed in the immune system and the brain. Cell 47, 333–348. Michelsen, K., Yuan, H., Schwappach, B., 2005. Hide and run. Arginine-based
endoplasmic-reticulum-sorting motifs in the assembly of heteromultimeric membrane proteins. EMBO. Rep 6, 717–722.
Modis, Y., Ogata, S., Clements, D., Harrison, S.C., 2003. A ligand-binding pocket in the dengue virus envelope glycoprotein. Proc. Natl. Acad. Sci. U. S. A. 100, 6986–6991.
Modis, Y., Ogata, S., Clements, D., Harrison, S.C., 2004. Structure of the dengue virus envelope protein after membrane fusion. Nature 427, 313–319.
Mukhopadhyay, S., Kuhn, R.J., Rossmann, M.G., 2005. A structural perspective of the flavivirus life cycle. Nat.Rev.Microbiol 3, 13–22.
Nilsson, T., Warren, G., 1994. Retention and retrieval in the endoplasmic reticulum and the Golgi apparatus. Curr. Opin. Cell Biol. 6, 517–521. Op De Beeck, A., Molenkamp, R., Caron, M., Ben Younes, A., Bredenbeek, P.,
Dubuisson, J., 2003. Role of the transmembrane domains of prM and E proteins in the formation of yellow fever virus envelope. J. Virol 77, 813–820. Op De Beeck, A., Rouille, Y., Caron, M., Duvet, S., Dubuisson, J., 2004. The transmembrane domains of the prM and E proteins of yellow fever virus are endoplasmic reticulum localization signals. J. Virol 78, 12591–12602. Pelham, H.R., 1995. Sorting and retrieval between the endoplasmic reticulum
and Golgi apparatus. Curr.Opin.Cell Biol 7, 530–535.
Purdy, D.E., Chang, G.J., 2005. Secretion of noninfectious dengue virus-like particles and identification of amino acids in the stem region involved in intracellular retention of envelope protein. Virology 333, 239–250. Purdy, D.E., Noga, A.J., Chang, G.J., 2004. Noninfectious recombinant antigen
for detection of St. Louis encephalitis virus-specific antibodies in serum by enzyme-linked immunosorbent assay. J. Clin. Microbiol 42, 4709–4717. Rey, F.A., Heinz, F.X., Mandl, C., Kunz, C., Harrison, S.C., 1995. The envelope
glycoprotein from tick-borne encephalitis virus at 2 A resolution. Nature 375, 291–298.
Schalich, J., Allison, S.L., Stiasny, K., Mandl, C.W., Kunz, C., Heinz, F.X., 1996. Recombinant subviral particles from tick-borne encephalitis virus are fusogenic and provide a model system for studying flavivirus envelope glycoprotein functions. J.Virol 70, 4549–4557.
Shin, J., Dunbrack Jr., R.L., Lee, S., Strominger, J.L., 1991. Signals for retention of transmembrane proteins in the endoplasmic reticulum studied with CD4 truncation mutants. Proc. Natl. Acad. Sci.U. S. A. 88, 1918–1922. Stadler, K., Allison, S.L., Schalich, J., Heinz, F.X., 1997. Proteolytic activation
of tick-borne encephalitis virus by furin. J.Virol 71, 8475–8481. Stiasny, K., Allison, S.L., Marchler-Bauer, A., Kunz, C., Heinz, F.X., 1996.
Structural requirements for low-pH-induced rearrangements in the envelope glycoprotein of tick-borne encephalitis virus. J.Virol 70, 8142–8147. Szczesna-Skorupa, E., Kemper, B., 2000. Endoplasmic reticulum retention
determinants in the transmembrane and linker domains of cytochrome P450 2C1. J.Biol.Chem 275, 19409–19415.
Wang, J.J., Liao, C.L., Chiou, Y.W., Chiou, C.T., Huang, Y.L., Chen, L.K., 1997. Ultrastructure and localization of E proteins in cultured neuron cells infected with Japanese encephalitis virus. Virology 238, 30–39.
Wang, W.K., Chen, H.L., Yang, C.F., Hsieh, S.C., Juan, C.C., Chang, S.M., Yu, C.C., Lin, L.H., Huang, J.H., King, C.C., 2006. Slower rates of clearance of viral load and virus-containing immune complexes in patients with dengue hemorrhagic fever. Clin.Infect.Dis 43, 1023–1030.
Xu, Z., Bruss, V., Yen, T.S., 1997. Formation of intracellular particles by hepatitis B virus large surface protein. J.Virol. 71, 5487–5494.
Yang, M., Ellenberg, J., Bonifacino, J.S., Weissman, A.M., 1997. The transmembrane domain of a carboxyl-terminal anchored protein determines localization to the endoplasmic reticulum. J.Biol.Chem 272, 1970–1975. Zhang, W., Chipman, P.R., Corver, J., Johnson, P.R., Zhang, Y., Mukhopadhyay,
S., Baker, T.S., Strauss, J.H., Rossmann, M.G., Kuhn, R.J., 2003. Visualization of membrane protein domains by cryo-electron microscopy of dengue virus. Nat.Struct.Biol 10, 907–912.