Abstract.
The effects of 50% ethanolic stem extracts of
Zanthoxylum ailanthoides Sieb and Zucc. (ZASZ) on the cell
viability, cell cycle and apoptosis were investigated in a
human colon adenocarcinoma cell line (colo 205). The results
demonstrated that ZASZ induced morphological changes and
decreased the cell viability. ZASZ promoted Wee1, checkpoint
kinase 2 (CHK2), p21 and p53 levels, decreased cyclin B and
cdc25c associated with that led to G
2/M phase arrest.
ZASZ-triggered apoptosis was confirmed by
4’-6-diamidino-2-phenylindole (DAPI) staining and DNA gel electrophoresis.
ZASZ increased the levels of glucose-regulated protein 78
(GRP78) and growth arrest and DNA damage inducible gene
153 (GADD153), and promoted an increase of reactive
oxygen species (ROS) and Ca
2+release, and loss of
mitochondrial membrane potential (ΔΨ
m) accompanied by
cytochrome c release that was due to the decrease of Bcl-2
and increase of Bax levels in the colo 205 cells. ZASZ also
induced the protein levels of apoptosis-inducing factor (AIF)
and endonuclease G (Endo G), increased the levels of
caspase-3, -7 and -9, and stimulated the levels of fatty acid
synthase (Fas) and Fas ligand in the colo 205 cells. ZASZ
contains phenolic compounds, including flavone, chlorogenic
acid and isofraxidin, among which, flavone was found to be
the most effective in reducing cell viability and proliferative
responses in the colo 205 cells. ZASZ induces cytotoxicity and
apoptosis in colo 205 cells which provides the rationale for
studies in animal models on the utilization of ZASZ as a
potential cancer therapeutic compound.
Colon cancer is a common malignancy, and one of the major
fatal diseases throughout the world (1). Furthermore, it is the
second most frequent cause of death from cancer in Taiwan,
where based on the reports from the Department of Health
of the Republic of China, 19.6 people per 100 hundred
thousand people die each year of colorectal cancer. The
therapy for human colorectal cancer in Taiwan includes
chemotherapy, radiotherapy and surgery. However, these
treatment strategies had limited success. New agents acting
on novel targets of carcinoma are needed.
The cyclin-dependent protein kinase (Cdk) family, cyclins
and Cdk inhibitors (CdKI) are essential for cellular
regulatory processes, particularly in the regulation of
transcription and cell cycle progression (G
1to S or G
2to M
phase) (2, 3). Cdk are regulated by multiple mechanisms
which include inhibitory phosphorylation of conserved
residues within the active site (4) and binding of additional
regulatory molecules (5). It is reasonable to propose that if
cyclin and/or Cdk are affected then cell cycle arrest and
apoptosis could be induced.
It is well known that apoptosis plays a variety of
important roles under normal physiological conditions (6).
Correspondence to: Jing-Gung Chung, Department of BiologicalScience and Technology, China Medical University, No 91, Hsueh-Shih Road, Taichung 404, Taiwan, R.O.C. Tel: +886 422053366, ext. 2161, Fax: +886 422053764, e-mail: [email protected]
Key Words: Zanthoxylum ailanthoides Sieb and Zucc., human colon
adenocarcinoma cell, colo 205 cells, caspases, apoptosis, flavone.
Zanthoxylum ailanthoides Sieb and Zucc. Extract
Inhibits Growth and Induces Cell Death through G
2
/M-phase
Arrest and Activation of Apoptotic Signals
in Colo 205 Human Colon Adenocarcinoma Cells
SU-TZE CHOU
1, HSIN-YI PENG
1, CHEN-TIEN CHANG
1, JAI-SING YANG
2,
HSIUNG-KWANG CHUNG
3,4, SU-TSO YANG
5, W. GIBSON WOOD
6and JING-GUNG CHUNG
7,8 1Department of Food and Nutrition, Providence University, Taichung 433, Taiwan, R.O.C.;
Departments of
2Pharmacology and
7Biological Science and Technology,
4
Graduate Institute of Clinical Medical Science, China Medical University, Taichung 404, Taiwan, R.O.C.;
Departments of
3Otorhinolaryngology-Head and Neck Surgery and
5
Radiology, China Medical University Hospital, Taichung 404, Taiwan, R.O.C.;
8Department of Biotechnology, Asia University, Taichung 413, Taiwan, R.O.C.;
6
Department of Pharmacology, University of Minnesota,
Impaired apoptosis is a crucial step in the process of cancer
development (7, 8). Cells undergo apoptosis through
distinct pathways based on stimulation of caspases, a
family of proteases that are important in regulating
apoptosis (5). It is well known that compounds that can
induce apoptosis can be divided into the fatty acid synthase
(Fas) and Fas ligand (FasL) pathways resulting in the
activation of the caspase-8, mitochondria-dependent
pathway and the caspase-3-dependent pathway. Both
pathways trigger the cytoplasmic release of pro-apoptotic
mitochondrial proteins and stimulate the endoplasmic
reticulum (ER) stress pathway which is attributed to the
activation of caspase-12 and eventual apoptosis (9).
Therefore, the resistance of cancer cells to treatment might
be associated with dysregulation or defects in different
steps of the apoptotic pathways.
Numerous naturally occurring compounds are thought to
have potential value for cancer prevention and cancer
therapy (10, 11) and they were recognized to be a potential
inhibitor of tumor cell proliferation, carcinogen-induced
carcinogenesis or inducers of apoptosis (12-14). The tender
leaves of Zanthoxylum ailanthoides Sieb and Zucc. have
been used as a dietary supplement in the Taiwanese
population for many years (15). In the Chinese population,
the plant Z. ailanthoides has been used as a herbal medicine
for the treatment of heart disease, bone-injury and cold
resistance. Our previous study demonstrated that a 50%
ethanolic stem extract of Z. ailanthoides (ZASZ) was safe
from a genotoxic perspective and acted as an antioxidant
(16). However, there is no information on effects of ZASZ.
Therefore, in the present study, the possible anticancer
activity of ZASZ and whether it acts on cell cycle arrest and
triggers tumor cell apoptosis were determined.
Materials and Methods
Plant material and preparation of extracts. The ZASZ (50%
ethanolic stem extract) was prepared as previously described (16). Briefly, the plant material was obtained from the mountain area of Sinshe Township, Taichung, Taiwan. Each 100 g stem was extracted with 500 ml 50% ethanol at 75˚C for 3 h and decocted three times. The decoctions were filtered and then dried by a vacuum freeze-dryer. The extraction yields were 9.0%. The extracts were sealed in plastic bottles and stored at –70˚C until used. For the present experiments, extracts were dissolved in PBS before adding to cell cultures.
Chemical and reagents. Propidium iodide (PI),
4’-6-diamidino-2-phenyindole (DAPI), ethidium bromide (EtBr), triton X-100, trypan blue and ribonuclease A (RNase A) were obtained from Sigma-Aldrich Corp. (St. Louis, MO, USA). RPMI 1640 medium, Dulbecco’s modified Eagle’s medium (DMEM), L-glutamine, fetal bovine serum (FBS), penicillin-streptomycin and trypsin-EDTA were obtained from Invitrogen Life Technologies (Carlsbad, CA, USA). All other chemicals were at least reagent grade.
Cell lines and culture conditions. Human colon adenocarcinoma cell
line (colo 205), human hepatoma cell line (Hep G2), human murine leukemia cell line (WEHI-3), mouse melanoma cell line (B16-F1) and promyelomonocytic cells line (HL60) were obtained from the Food Industry Research and Development Institution (Hsinchu, Taiwan). The colo 205 and WEHI-3 cells were cultured in RPMI 1640 medium, and the Hep G2 and B16-F1 cells were cultured in DMEM supplemented with 10% FBS, 2 mM L-glutamine, and 1% penicillin- streptomycin (100 U/ml penicillin and 100 μg/ml streptomycin) (17). All the cells were maintained in a humidified 5% CO2incubator at 37˚C and the cells were sub-cultured every 3-4 days to maintain logarithmic growth and were allowed to grow for 24 h before treatments were applied.
Morphological changes and cell viability analysis. The colo 205,
Hep G2, B16-F1 and WEHI-3 cells were plated in 12-well plates at a density of 1×105cells/ml. The cells were treated with different
concentrations (31.25, 62.5, 125.0, 250.0 or 500.0 μg/ml) of ZASZ or PBS as a solvent control and grown at 37˚C, 5% CO2and 95% air for 24, 48 or 72 h. To determine morphological changes, the cells were photographed by a phase-contrast microscope. Cell viability was determined by a PI exclusion method and by using a flow cytometer (Becton-Dickinson, FACSCalibur, San Jose, CA, USA) equipped with an argon ion laser at 488 nm wavelength as previously described (17).
Cell cycle and apoptosis analysis. The colo 205 cells were incubated
with different concentrations of ZASZ (0, 31.25, 62.5, 125.0, 250.0 or 500.0 μg/ml) for 48 h. The cells were then harvested by centrifugation and fixed gently in 70% ethanol (in PBS) at -20˚C overnight. The cells were then re-suspended in PBS containing 40 μg/ml PI and 0.1 mg/ml RNase A and 0.1% triton X-100 in a dark room for 30 min and analyzed by flow cytometry (17, 18). A FITC annexin V apoptosis detection kit from BD Pharmingen (San Diego, CA, USA) was used to determine and quantify the apoptotic cells by flow cytometry (19).
DAPI staining. The colo 205 cells were placed for 24 h and then
incubated with various concentrations of ZASZ (0, 31.25, 62.5, 125.0, 250.0 or 500.0 μg/ml) for 48 h. The cells were washed by PBS, fixed with 4% formaldehyde for 15 min, stained with DAPI (Sigma) at 37˚C for 30 min and photographed using a fluorescence microscope as previously described (17, 20).
DNA fragmentation assay. The colo 205 cells were incubated with
different concentrations of ZASZ (0, 31.25, 62.5, 125.0, 250.0 or 500.0 μg/ml) for 48 h. The cells were then harvested by centrifugation, the DNA was isolated and was then gel electrophoresized and stained with EtBr and photographed under UV light as previously described (17, 21).
Measurement of reactive oxygen species (ROS) production, mitochondrial membrane potential (ΔΨm) and Ca2+release. The
colo 205 cells were incubated with 250.0 μg/ml of ZASZ for 0, 2, 4, 6 or 24 h. The cells were harvested and washed twice with PBS then re-suspended with specific fluorochromes. For ROS analysis, the cells were re-suspended in 500 μl 10 μM of 2,7-dichlorodihydrofluorescein diacetate (DCFH-DA, Invitrogen) and incubated at 37˚C for 30 min and then analyzed by flow cytometry as previously described (22, 23). For ΔΨmanalysis, the cells were re-suspended in 500 μl of 3,3’-dihexyloxcarbocyanine (DiOC6, Invitrogen) (1 μmol/l) and incubated at 37˚C for 30 min and
analyzed by flow cytometry as previously described (22, 24). To detect ΔΨmrelease, the cells were re-suspended in Indo 1/AM (3 μg/ml, Invitrogen) and incubated at 37˚C for 40 min. After incubation, the cells were washed with PBS and analyzed by flow cytometry as previously described (22).
Determination of G2/M phase arrest and apoptosis-related protein levels. The colo 205 cells were incubated with 250.0 μg/ml of ZASZ
for 0, 6, 12, 24 or 48 h. After treatment, the cell lysates were used to determine the protein abundance of cyclin A and B, cell division control 2 (cdc2), cdc25c, checkpoint kinase 2 (CHK2), Wee1, ataxia-telangiectasia mutated (ATM), p21, p53, B-cell lymphoma 2 (Bcl-2), Bcl-2 associated X (Bax), cytochrome c, apoptosis-inducing factor (AIF) and endonuclease G (Endo G), caspase-9, -3, -7, poly (ADP-ribose) polymerase (PARP), fatty acid synthase (Fas), Fas ligand (FasL), caspase-8, truncated BH3 interacting domain death agonist (tBid), glucose-regulated protein 78 (GRP78), growth arrest and DNA damage inducible gene 153 (GADD153) and β-actin by sodium dodecylsulfate polyacrylamide gel electrophoresis (SDS-PAGE) and Western blotting as described previously (17, 22).
Characterization of phenolic compounds. Phenolic compounds in
the ZASZ extract were analyzed based on the method described by Li et al. (25) with a slight modification. Dried ZASZ (0.1 g) was dissolved in 10 ml of 50% ethanol, filtered through a 0.45 μm filter, and analyzed by high-performance liquid chromatography (HPLC). HPLC analysis was performed using a Hewlett- Packard HPLC System (HP 1100 series, Waldron, Germany), consisting of a quaternary pump and a variable wavelength detector (VWD) at 270 nm and equipped with a Li-Chrospher RP-18 cartridge column (250 mm ×4.6 mm, 5 μm, Merck KGaA, Darmstadt, Germany). The mobile phase was a stepwise gradient of water (0.1% v/v phosphoric acid)-acetonitrile (0.01 min, 94: 6; 50 min, 65: 35), and the injection volume was 30 μl. The identification of each compound was based on a combination of retention time and spectral matching by comparison with those of known standards. Peak areas and concentrations were determined using the Hewlett- Packard Chem Station Chromagraphic Management System.
Statistical analysis. Student’s t-test was used to analyze statistical
differences between the ZASZ treated and control groups. A p-value less than 0.05 was considered significant. All data are presented as means±S.D. of three independent experiments.
Results
Effects of ZASZ and its active phenolic compounds on cell
viability in human cancer cell lines. Each cell line responded
differently to the ZASZ treatment with the colo 205 cells
most susceptible to ZASZ-induced toxicity (Figure 1A). The
500 μg/ml ZASZ treatment over 48 h decreased cell viability
by 46.4%, 18.1%, 9.2% and 5.2% in the colo 205, Hep G2,
B16-F1 and WEHI-3 cells, respectively. Therefore, the colo
205 cells were used in the subsequent experiments because
of the greater effects of and sensitivity to ZASZ on those
cells compared with the other cell lines.
The ten standard polyphenols used for comparison showed
the retention times of gallic acid, chlorogenic acid, caffeic
acid, flavone, isofraxidin, rutin, genistin, daidzin, quercetin
and genistein to be 5.68, 14.78, 16.93, 21.28, 25.89, 26.87,
28.66, 39.43, 43.10 and 49.37 min, respectively, using HPLC
analysis (chromatograms not shown). The data in Table I
showed that ZASZ contained three major phenolic
substances, including chlorogenic acid, flavone and
isofraxidin with flavone the most abundant phenol. The
effects of chlorogenic acid, flavone and isofraxidin on cell
Table I. Phenolic contents of ZASZ.
Compound Content (mg/g extract) Chlorogenic acid 2.74±0.21 Flavone 22.68±0.90 Isoflaxidin 1.46±0.14 Reported values are the means±SD (n=3).
Figure 1. Effects of the ZASZ on the proliferation of colo 205, B16-F1,
Hep G2 and WEHI-3 cells (A) and the active phenolic compounds in ZASZ on the cell viability of colo 205 cells (B). Means±S.D. of three independent experiments. *p<0.05 compared with the control sample.
viability were examined, and results from Figure 1B
indicated that the induction of toxicity of the three phenolic
compounds was dose-dependent. The cell viability followed
the order flavone>chlorogenic acid>isofraxidin.
Effects of ZASZ on cell morphology, cell viability, cell cycle
distribution and apoptosis. The phase-contrast microscopic
examination indicated that ZASZ-induced apoptotic
morphological changes such as shrinking and cytoplasmic
Figure 2. Effects of ZASZ on cell morphology (A) and cell viability (B) up to 72 h and , DNA content for cell cycle distribution (C) and apoptosis
blebbing were found (Figure 2A) and a PI stain analysis by
flow cytometry showed that the percentage of viable cells was
significantly different between the ZASZ-treated and the
control groups (Figure 2B). These effects were dose- and
time-dependent. ZASZ induced G
2/M-phase arrest in the colo
205 cells (Figure 2C) and the effect was dose-dependent. It
can be seen in Figure 2D that ZASZ also induced apoptosis in
the colo 205 cells. To further confirm that ZASZ induced
apoptosis, the results determined by DAPI staining. Figure
3A shows that apoptotic cell death occurred after ZASZ
treatment and Figure 3B demonstrates evidence of DNA
fragmentation in the ZASZ -treated colo 205 cells.
Effects of ZASZ on ROS and Ca
2+levels and ΔΨ
m. The ROS
levels were stimulated when the colo 205 cells were incubated
with 250 μg/ml of ZASZ and peaked after a 2 h exposure,
then decreasing slightly up to 24 h. However, the ROS levels
were still higher in the ZASZ-treated cells than in the control
cells for at least 24 h (Figure 4A). Flow cytometric analysis
indicated that the ΔΨ
mdecreased in the ZASZ-treated cells as
Figure 3. The induction of apoptosis by ZASZ in colo 205 cells was determined by DAPI staining and fluorescence microscopy (A) and DNA gel
compared with the control cells after 24 h treatment (Figure
4B). Also, it can be seen in Figure 4C that the cytoplasmic
Ca
2+level peaked at 2 h and then began to decrease but were
higher than the control cells at 24 h (p<0.05).
Effects of ZASZ on G
2/M-phase arrest and
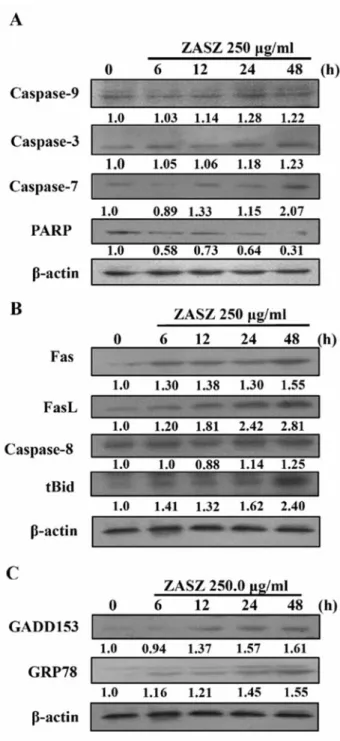
apoptosis-associated specific protein levels. The Western blotting
analysis is shown in Figures 5 and 6. Protein levels of
CHK2, Wee1, ATM, p21, p53 (Figure 5B), Bax (Figure 5C),
cytochrome c, AIF, Endo G (Figure 5D), caspases-9, -3, and
-7 (Figure 6A), Fas, FasL, caspase-8, tBid (Figure 6B),
GRP78 and GADD153 (Figure 6C) increased and cyclins A
and B , cdc2, cdc25c (Figure 5A), Bcl-2 (Figure 5C) and
PARP (Figure 6A) reduced following ZASZ treatment in the
colo 205 cells.
Discussion
Until now, a cure for colon cancer is not satisfying and
numerous studies focused on finding new agents for colon
cancer therapy. In this study, we showed that ZASZ induced
G
2/M arrest and Western blot analysis showed that it was
mediated through the decreased protein expressions
of cdc25c, Wee1 and CKI proteins (p21
CIP1/WAF1), a
simultaneous decrease in cyclins A and B and enhanced
binding of CKI-Cdk. The concentration of 250.0 μg/ml of
ZASZ was selected for all the Western blotting analysis based
on the viability studies (Figure 2) and was close to the IC
50for 72 h exposure. p21
CIP1/WAF1is a universal inhibitor of
Cdk(s) and p27
KIP1is commonly upregulated in response to
antiproliferative signals (26). The present data also showed
that ZASZ induced the expression of p53 in the colo 205 (p53
wild-type) cells (Figure 5B). The tumor suppressor gene p53
is activated in response to various genotoxic stresses,
resulting in cell cycle arrest or apoptosis. Cell cycle arrest that
is dependent on p53 requires transactivation of p21
WAF1/CIP1,
GADD45 and cyclin G through the binding of the cyclin-cdk
complex (27). The blockade of cell cycle progression by
ZASZ was through activation of p21
WAF1/CIP1involving a
p53 dependent pathway (Figure 5A and B). It has been
reported that p53 is responsible for the up-regulation of Fas,
Bax, Bad and Bcl-xl resulting in apoptotic cell death (28).
Therefore, ZASZ not only increased the cell cycle-related
protein (p21
WAF1/CIP1), but also enhanced the proapoptotic
expression of Bax, Fas and reduced levels of the
anti-apopototic protein Bcl-2 (Figure 5 and 6).
Normal cells maintain a balance of pro-apoptotic and
anti-apoptotic Bcl-2 family proteins which integrate the diverse
death- and survival-signals to control the fate of the cell (8,
29) including both Bcl-2 and Bax. The present data
demonstrated that the ZASZ-induced apoptosis was
associated with an increase of Bax and a reduction of Bcl-2
protein levels, and both Bcl-2 family proteins were in
agreement with the notion that an imbalance between
pro-and anti-apoptotic proteins of the Bcl-2 family results in
apoptosis (8, 29). Anti-apoptotic Bcl-2 family proteins are
also known to prevent the release of caspase-9 (30), and
regulate the release of cytochrome c from the mitochondria
Figure 4. Effects of ZASZ (250 μg/ml) on the levels of reactive oxygen
species (ROS) (A), mitochondria membrane potential (ΔΨm) (B) and Ca2+ release (C) in colo 205 cells. Mean±S.D. of triplicated experiments. *p<0.05 compared to control sample.
into the cytosol (31, 32). The over-expression of Bcl-xl
protein (anti-apoptotic) can block the release of cytochrome
c in response to a variety of apoptotis signals (32). However,
the Bax protein (pro-apoptotic) promotes cytochrome c
release from the mitochondria (31, 32). Western blotting
demonstrated that ZASZ decreased the ΔΨ
mand promoted
the levels of cytochrome c release from the mitochondria
(Figure 5D). The induction of the execution protease,
caspase-3 was also demonstrated. This was in agreement
with many reports demonstrating that released cytochrome c
from mitochondria activates initiator caspase-9 to activate a
sequential cascade of caspases, especially caspase-3 resulting
in the proteolysis of death substrates and subsequent DNA
degradation and apoptotic cell death (8, 29, 33, 34).
ZASZ also increased ROS and Ca
2+levels in the colo 205
cells (Figure 4A and C). ROS can stimulate ER stress
resulting in Ca
2+release and plays an important role in the
induction of apoptosis (35, 36). ZASZ increased the levels
of GADD153 and GRP78 (Figure 6C) also associated with
ER stress because GADD153 is a modulator and GRP78 is a
sensor of ER stress. Thus, ZASZ induced cytotoxicity in the
colo 205 cells which was mediated by several different
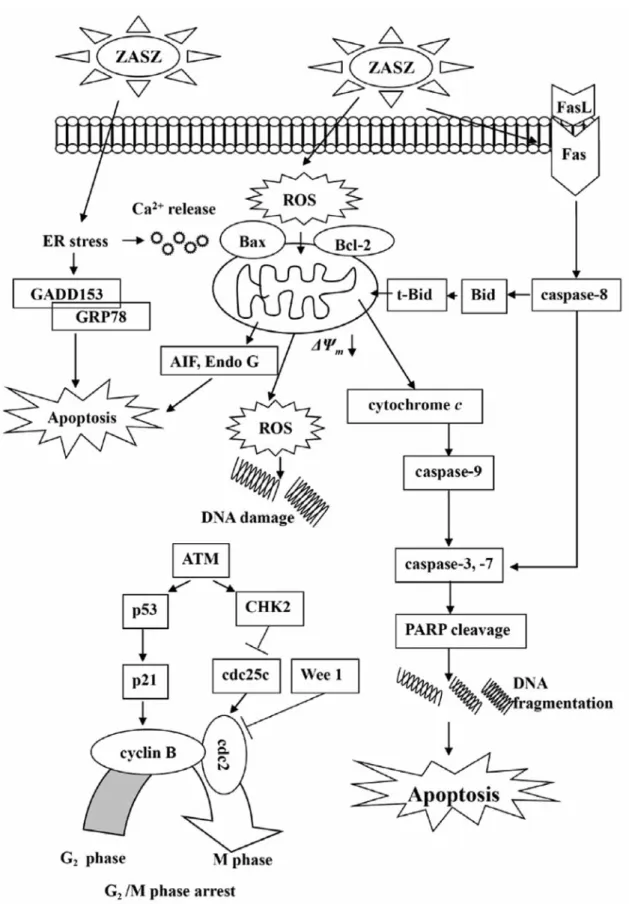
pathways associated with apoptosis as depicted in Figure 7.
ZASZ was found to contain approximately 3% phenolic
compounds mostly flavone, chlorogenic acid and isofraxidin.
Flavone was highest in abundance and was the least effective in
reducing cell viability in the colo 205 cells. Flavones are
polyphenolic compounds that occur naturally in many plants
used for food. In vitro studies on the potential anticancer activity
of flavones have been reported in different cell types (37-39).
Flavone may contribute to the antiproliferative and apoptotic
activities of the ZASZ extracts used in the present study.
In conclusion, ZASZ induces morphological changes,
decreases the percentage of viable cells, and induces G
2/M
phase arrest of cell cycle and apoptosis involving ROS
production, ER stress, caspase-3, AIF and Endo G signaling
pathways. Further studies are needed on the effectiveness of
ZASZ in cancer cells particularly studies in animal models.
Figure 5. Effects of ZASZ on the protein levels of A: cyclin A and B, cdc2 and cdc25c; B: CHK2, Wee1, ATM, p21 and p53; C: Bcl-2 and Bax; D:
Acknowledgements
This research was supported by the National Science Council, Taiwan (NSC 96-2313-B-126-002- MY2) and its financial support is greatly appreciated.
References
1 Yee J, Rosen MP, Blake MA, Baker ME, Cash BD, Fidler JL, Grant TH, Greene FL, Jones B, Katz DS, Lalani T, Miller FH, Small WC, Sudakoff GS and Warshauer DM: ACR Appropriateness Criteria on colorectal cancer screening. J Am Coll Radiol 7: 670-678, 2010. 2 Morgan DO: Cyclin-dependent kinases: engines, clocks, and
microprocessors. Annu Rev Cell Dev Biol 13: 261-291, 1997. 3 Collins I and Garrett MD: Targeting the cell division cycle in
cancer: CDK and cell cycle checkpoint kinase inhibitors. Curr Opin Pharmacol 5: 366-373, 2005.
4 Solomon MJ and Kaldis P: Regulation of CDKs by phosphorylation. Results Probl Cell Differ 22: 79-109, 1998. 5 Sherr CJ and Roberts JM: CDK inhibitors: positive and negative
regulators of G1-phase progression. Genes Dev 13: 1501-1512, 1999.
6 Alison MR and Sarraf CE: Apoptosis: regulation and relevance to toxicology. Hum Exp Toxicol 14: 234-247, 1995.
7 Johnstone RW, Ruefli AA and Lowe SW: Apoptosis: a link between cancer genetics and chemotherapy. Cell 108: 153-164, 2002.
8 Thompson CB: Apoptosis in the pathogenesis and treatment of disease. Science 267: 1456-1462, 1995.
9 Szegezdi E, Fitzgerald U and Samali A: Caspase-12 and ER-stress-mediated apoptosis: the story so far. Ann NY Acad Sci
1010: 186-194, 2003.
10 Cerella C, Sobolewski C, Dicato M and Diederich M: Targeting COX-2 expression by natural compounds: a promising alternative strategy to synthetic COX-2 inhibitors for cancer chemoprevention and therapy. Biochem Pharmacol 80: 1801-1815, 2010.
11 Trottier G, Bostrom PJ, Lawrentschuk N and Fleshner NE: Nutraceuticals and prostate cancer prevention: a current review. Nat Rev Urol 7: 21-30, 2010.
12 Steinmetz KA, Kushi LH, Bostick RM, Folsom AR and Potter JD: Vegetables, fruit, and colon cancer in the Iowa Women’s Health Study. Am J Epidemiol 139: 1-15, 1994.
13 Janakiram NB, Mohammed A, Zhang Y, Choi CI, Woodward C, Collin P, Steele VE and Rao CV: Chemopreventive effects of Frondanol A5, a Cucumaria frondosa extract, against rat colon carcinogenesis and inhibition of human colon cancer cell growth. Cancer Prev Res (Phila) 3: 82-91, 2010.
14 Nakamura Y: Chemoprevention by isothiocyanates: molecular basis of apoptosis induction. Forum Nutr 61: 170-181, 2009. 15 Chou ST, Chan HH, Peng HY, Liou MJ and Wu TS: Isolation of
substances with antiproliferative and apoptosis-inducing activities against leukemia cells from the leaves of Zanthoxylum
ailanthoides Sieb. and Zucc. Phytomedicine 18: 344-348, 2011.
16 Chung YC, Chang CT, Teng KY and Chou ST: Antioxidative and mutagenic properties of Zanthoxylum ailanthoides Sieb and
Zucc. Food Chem 97: 418-425, 2006.
17 Su CC, Chen GW, Lin JG, Wu LT and Chung JG: Curcumin inhibits cell migration of human colon cancer colo 205 cells through the inhibition of nuclear factor kappa B /p65 and down-regulates cyclooxygenase-2 and matrix metalloproteinase-2 expressions. Anticancer Res 26: 1281-1288, 2006.
18 Wu SH, Hang LW, Yang JS, Chen HY, Lin HY, Chiang JH, Lu CC, Yang JL, Lai TY, Ko YC and Chung JG: Curcumin induces apoptosis in human non-small cell lung cancer NCI-H460 cells through ER stress and caspase cascade- and mitochondria-dependent pathways. Anticancer Res 30: 2125-2133, 2010. Figure 6. Effects of ZASZ on the protein levels of A: caspase-9, -3, -7
and PARP; B: Fas, FasL, caspase-8 and tBid; C: GADD153 and GRP78 in colo 205 cells, shown by Western blotting. β-Actin, internal control.
Figure 7. Proposed possible signaling pathways included by ZASZ treatment. ZASZ induces ROS, ER stress, GRP78 and GADD153 expression
leading to Ca2+release, promotes Bax, but inhibits Bcl-2 before decreasing the ΔΨ
mleading to cytochrome c release, caspase-9 activation and caspase-3 activity causing apoptosis.
19 Chiang JH, Yang JS, Ma CY, Yang MD, Huang HY, Hsia TC, Kuo HM, Wu PP, Lee TH and Chung JG: Danthron, an anthraquinone derivative, induces DNA damage and caspase cascades-mediated apoptosis in SNU-1 human gastric cancer cells through mitochondrial permeability transition pores and Bax-triggered pathways. Chem Res Toxicol 24: 20-29, 2011. 20 Lu CC, Yang JS, Huang AC, Hsia TC, Chou ST, Kuo CL, Lu HF,
Lee TH, Wood WG and Chung JG: Chrysophanol induces necrosis through the production of ROS and alteration of ATP levels in J5 human liver cancer cells. Mol Nutr Food Res 54: 967-976, 2010.
21 Lu HF, Lai TY, Hsia TC, Tang YJ, Yang JS, Chiang JH, Lu CC, Liu CM, Wang HL and Chung JG: Danthron induces DNA damage and inhibits DNA repair gene expressions in GBM 8401 human brain glioblastoma multiforms cells. Neurochem Res 35: 1105-1110, 2010.
22 Wu CC, Lin JP, Yang JS, Chou ST, Chen SC, Lin YT, Lin HL and Chung JG: Capsaicin induced cell cycle arrest and apoptosis in human esophagus epidermoid carcinoma CE 81T/VGH cells through the elevation of intracellular reactive oxygen species and Ca2+productions and caspase-3 activation. Mutat Res 601:
71-82, 2006.
23 Lo C, Lai TY, Yang JH, Yang JS, Ma YS, Weng SW, Chen YY, Lin JG and Chung JG: Gallic acid induces apoptosis in A375.S2 human melanoma cells through caspase-dependent and -independent pathways. Int J Oncol 37: 377-385, 2010. 24 Lin SY, Lai WW, Ho CC, Yu FS, Chen GW, Yang JS, Liu KC,
Lin ML, Wu PP, Fan MJ and Chung JG: Emodin induces apoptosis of human tongue squamous cancer SCC-4 cells through reactive oxygen species and mitochondria-dependent pathways. Anticancer Res 29: 327-335, 2009.
25 Li Q, Jia Y, Xu L, Wang X, Shen Z, Liu Y and Bi K: Simultaneous determination of protocatechuic acid, syringin, chlorogenic acid, caffeic acid, liriodendrin and isofraxidin in
Acanthopanax senticosus Harms by HPLC-DAD. Biol Pharm
Bull 29: 532-534, 2006.
26 Olinari MM: Cell cycle checkpoints and their inactivation in human cancer. Cell Prolif 33: 261-274, 2000.
27 May P and May E: Twenty years of p53 research: structural and functional aspects of the p53 protein. Oncogene 18: 7621-7636, 1999.
28 Sheikh MS and Fornace AJ Jr.: Role of p53 family members in apoptosis. J Cell Physiol 182: 171-181, 2000.
29 Adams JM and Cory S: Life-or-death decisions by the Bcl-2 protein family. Trends Biochem Sci 26: 61-66, 2001.
30 Oltvai ZN, Milliman CL and Korsmeyer SJ: Bcl-2 heterodimerizes in vivo with a conserved homolog, Bax, that accelerates programmed cell death. Cell 74: 609-619, 1993. 31 Budihardjo I, Oliver H, Lutter M, Luo X and Wang X:
Biochemical pathways of caspase activation during apoptosis. Annu Rev Cell Dev Biol 15: 269-290, 1999.
32 Srivastava M, Ahmad N, Gupta S and Mukhtar H: Involvement of Bcl-2 and Bax in photodynamic therapy-mediated apoptosis. Antisense Bcl-2 oligonucleotide sensitizes RIF 1 cells to photodynamic therapy apoptosis. J Biol Chem 276: 15481-15488, 2001.
33 Kuida K: Caspase-9. Int J Biochem Cell Biol 32: 121-124, 2000. 34 Wang X: The expanding role of mitochondria in apoptosis.
Genes Dev 15: 2922-2933, 2001.
35 Oh SH and Lim SC: A rapid and transient ROS generation by cadmium triggers apoptosis via caspase-dependent pathway in HepG2 cells and this is inhibited through N-acetylcysteine-mediated catalase upregulation. Toxicol Appl Pharmacol 212: 212-223, 2006.
36 Wu XJ, Kassie F and Mersch-Sundermann V: The role of reactive oxygen species (ROS) production on diallyl disulfide (DADS) induced apoptosis and cell cycle arrest in human A549 lung carcinoma cells. Mutat Res 579: 115-124, 2005.
37 Kawaii S, Tomono Y, Katase E, Ogawa K and Yano M: Antiproliferative activity of flavonoids on several cancer cell lines. Biosci Biotechnol Biochem 63: 896-899, 1999.
38 Kuntz S, Wenzel U and Daniel H: Comparative analysis of the effects of flavonoids on proliferation, cytotoxicity, and apoptosis in human colon cancer cell lines. Eur J Nutr 38: 133-142, 1999. 39 Wenzel U, Kuntz S, Brendel MD and Daniel H: Dietary flavone is a potent apoptosis inducer in human colon carcinoma cells. Cancer Res 60: 3823-3831, 2000.