INTRODUCTION
White spot syndrome (WSS) has now become the most serious disease affecting the global shrimp aqua-culture industry. The causative agent, white spot syn-drome virus (WSSV), is an enveloped, ellipsoid, large, and double-stranded DNA virus (Wang et al. 1995, Wongteerasupaya et al. 1995, Lo et al. 1996a) that has a wide host range among crustaceans (Lo et al. 1996b, Flegel 1997, Lo & Kou 1998). The size of the WSSV genome has been reported to differ among isolates, and varies from 305 107 bp (GenBank Accession No. AF332093) to 292 967 bp (GenBank Accession No. AF369029) and 307 287 bp (GenBank Accession No. AF440570) for viruses isolated from China, Thailand and Taiwan, respectively. Recent research on WSSV structure using proteomic methods has revealed that its structural proteins have increased in number to 39 (Tsai et al. 2004). Now, research activities have begun to focus on the protein-protein interactions between the shrimp and virus, between shrimp proteins, and between virus proteins (Lu & Kwang 2004, Tonganunt
et al. 2005, Wang et al. 2005, Xie & Yang 2005). According to previous studies by Tsai et al. (2004, 2006), VP53A encoded by WSSV gene wssv067 is an envelope protein. Proteomic data indicates that VP53A (molecular weight 53 kDa) comprises the C terminus of WSSV067 that codes for a deduced protein of 150 kDa. We selected WSSV067C as the bait to perform a yeast 2-hybrid screen of a Penaeus monodon cDNA library. A positive clone containing a conserved chitin-binding Type 2 domain was obtained. Chitin-binding proteins (CBPs) such as chitinase and lectin that contain a chitin-binding Type 2 domain are known to participate in immune responses (Tjoelker et al. 2000, Gaines et al. 2003).
MATERIALS AND METHODS
Virus and genomic plasmid libraries. WSSV col-lected from infected Penaeus monodon (Wang et al. 1995) from Taiwan in 1994 was used to construct WSSV genomic libraries (Lo et al. 1996a, Tsai et al.
© Inter-Research 2007 · www.int-res.com *Email: [email protected]
White spot syndrome virus envelope protein
VP53A interacts with Penaeus monodon
chitin-binding protein (PmCBP)
Li-Li Chen
1,*, Li-Chen Lu
1, Wan-Jung Wu
1, Chu-Fang Lo
2, Wei-Pang Huang
2 1Department of Biomedical Sciences, Chung Shan Medical University, No. 110, Sec. 1, Chien-Kuo N. Road, Taichung 402,Taiwan, ROC
2Institute of Zoology, National Taiwan University, No. 1, Sec. 4, Roosevelt Road, Taipei, Taiwan 10617, ROC
ABSTRACT: White spot syndrome virus (WSSV) is the causative agent of a severe disease of culti-vated shrimp. Using purified WSSV virions, VP53A encoded by open reading frame wssv067 was identified as a structural protein by SDS-PAGE and proteomics. Immunoelectron microscopy with a gold-labeled secondary antibody revealed that VP53A was distributed on the viral envelope. In order to further explore the link between WSSV067 and host proteins, we performed a yeast 2-hybrid screening of a Penaeus monodon cDNA library, using WSSV067C as bait. One of the molecules that specifically interacted with WSSV067C was the P. monodon chitin-binding protein (PmCBP). An in vitro binding assay showed that c-myc-WSSV067C was capable of co-precipitating HA-PmCBP-C. Furthermore, PmCBP was expressed in almost all organs but appeared to be up-regulated at the late stage of WSSV infection.
KEY WORDS: Penaeus monodon · White spot syndrome virus · VP53A · Envelope protein · Chitin-binding protein
2000, Liu et al. 2001). This virus source has been main-tained in our laboratory since 1994 (GenBank Acces-sion No. AF440570). It was previously known simply as the WSSV Taiwan isolate (Lo et al. 1999), but to distin-guish it from other WSSV Taiwan isolates, it will henceforth be referred to as the WSSV Taiwan-1 strain (WSSV T-1 strain). WSSV T-1 strain was used as the basis for all of the WSSV genome sequence research conducted in the present study.
Antibody preparation.The DNA sequence encoding 200 deduced amino acids from the WSSV067 C-termi-nus was amplified from WSSV-infected shrimp by PCR with the 067CFBamHI/067CRSalI primer set (5’-GGGGATCCGATAACGGTTGAAGGAGTC-3’/5’-GG AAGCTTTTACATCAACACTGTAACTGC-3’; the un-derlined bases indicate respectively the BamHI and HindIII restriction sites that were used for subsequent cloning) and ligated to pGEM-T Easy (Promega). After confirming the sequence, the resulting plasmid pGEM-T-VP53AC was cleaved with BamH1 and Sal1, and the amplified fragment was then cloned to pET28b(+) (Novagen) at BamHI and HindIII sites. The resulting pET-VP53AC clone was transformed into BL21 Codon Plus Escherichia coli cells (Stratagene). For protein expression and purification, cells were grown overnight at 37°C in Luria-Bertani (LB) media supplemented with 50 µg ml–1 kanamycin and 34 µg ml–1 chloramphenicol. The cells were inoculated into new media at a ratio of 1:300, and grown at 37°C for 1.5 to 2 h. Expression was induced by addition of 1 mM isopropyl β-D-1-thiogalactopyranoside (IPTG) and incubation was continued for another 1.5 to 3 h. The induced bacteria were spun down at 4°C, suspended in ice-cold phosphate-buffered saline (PBS) containing 10% glycerol and a Protease Inhibitor Cocktail tablet (Roche Molecular BioChemicals), and sonicated for 30 s on ice. The insoluble debris was collected by cen-trifugation, suspended with PBS containing 1.5% sodium lauryl sarcosine, and solubilized by shaking at 4°C for 2 h. The supernatant was clarified by centrifu-gation and mixed with Ni-NTA agarose beads (Quia-gen) on a rotary wheel at 4°C for 16 h or overnight. The beads were then washed several times with ice-cold wash buffer (1M NaCl, 10 mM Tris-HCl, pH 7.5) to remove unbound material. The fusion proteins were directly eluted from the beads with SDS sample buffer and subjected to SDS-PAGE analysis. The protein bands containing the fusion proteins were sliced from the gel, minced, mixed with Freund’s adjuvant and used for antibody production.
Localization of VP53A by immunoelectron micro-scopy (IEM). A purified WSSV virion suspension was adsorbed to Formvar-supported and carbon-coated nickel grids (150 mesh) and incubated for 5 min at room temperature. The primary antibody and
pre-immune rabbit serum were diluted to a 1:50 ratio in incubation buffer (0.1% AURION BASIC-c, 15 mM NaN3, 10 mM phosphate buffer, 150 mM NaCl, pH 7.4). The grids were blocked with blocking buffer (5% BSA, 5% normal serum, 0.1% cold water skin gelatin, 10 mM phosphate buffer, 150 mM NaCl, pH 7.4) for 15 min and then incubated with the diluted primary antibody or pre-immune rabbit serum for 1 h at room temperature. After several washes with incu-bation buffer, the grids were incubated with goat anti-rabbit secondary antibody conjugated with 6 nm or 15 nm diameter gold particles (1:40 dilution ratio in incubation buffer) for 1 h at room temperature. The grids were then washed extensively with incubation buffer, washed twice more with distilled water to remove excess salt and stained with 2% phospho-tungstic acid (PTA, pH 7.2) for 30 s. Specimens were examined by transmission electron microscopy (TEM). Tissue tropism analysis of PmCBP, temporal analy-sis of WSSV067 and PmCBP transcription by RT-PCR. The stomach, gills, midgut, haemolymph, lymphoid organ, hepatopancreas, haematopoietic tissue, pleo-pods, nervous tissue and heart were collected from Penaeus monodon without artificial WSSV infection. To date, no WSSV-susceptible shrimp cell lines have become available; therefore, all RNA for the transcrip-tional analysis was taken from different tissues of WSSV-infected shrimp at various times after infection, using procedures described by Chen et al. (2002a). Total RNA was isolated as described previously (Tsai et al. 2000, Liu et al. 2001). The procedure for cDNA syn-thesis and primer sets for vp26, vp28 and actin were all as previously described by Chen et al. (2002a). A WSSV genomic DNA-specific primer set IC-F2/IC-R3 (5’-CAGACTATTAATGTACAAGTGCG-3’/5’-GAAT-GATTGTTGCTGGTTAGAACC-3’), derived from an inter-genic region of the WSSV genome, was used to confirm that the RNA was not contaminated by any viral DNA. The cDNA reaction products were sub-jected to PCR with the primer sets 067-RTF2/067-RTR8 (GATCCACATTCTCCTTGAGC/CGTCCAAGTTAGT GCCGATT) for WSSV067 and PmCBP-RT-F1/PmCBP-RT-R1 (5’-GGCAGTGGATTCACTGGATC-3’/5’-TGT TCGCATGAGTTGTTCATTG-3’) for PmCBP.
Yeast 2-hybrid assay. The plasmids, yeast strains and library construction kits used in this experiment were all adopted from BD Matchmaker library con-struction and screening kits (BD Biosciences Clon-tech).
Plasmid construction: The DNA sequence encoding
the 577 deduced amino acids from the WSSV067 C-ter-minus was amplified by PCR with the Y2H-WSSV067C F/R primer set (5’-ATTGAGCGCCCCGGGGACGA-3’/5’-ATGTGGATCCTTACCTATCCATTTTTATGA-3’; the underlined bases indicate respectively the SmaI
and BamHI restriction sites that were used for subse-quent cloning) from WSSV-infected shrimp, and lig-ated to pGEM-T Easy (Promega). After confirming the sequence, the resultant plasmid pGEM-T-WSSV067C was cleaved with SmaI and BamHI, and the amplified fragment was then subcloned in frame into the bait vector pGBKT7 that encodes a GAL4 DNA-binding domain. The plasmid was named BD-WSSV067C.
Library construction: Total RNA was isolated from
Penaeus monodon postlarvae 15 to 20 using Trizol re-agent (Invitrogen). About 5 µg of total RNA was used to generate cDNA that was ligated into the pGADT7-Rec prey vector and then transformed into Saccha-romyces cerevisiae strain AH109, according to the instructions provided by the manufacturer.
Screening: The bait vector BD-WSSV067C was used
to screen independent recombinant clones of the cDNA library in Saccaromyces cerevisiae strain Y187. After mating the library host strain with the bait strain, positive clones were selected for growth on a synthetic dropout (SD) plate lacking adenine, histidine, leucine and tryptophan (SD/-ade/-his/-leu/-trp) and contain-ing 5-bromo-4-chloro-3-indolyl-α-D-galactopyranoside (X-α-Gal). Plasmid DNA was isolated from those clones that activated all 4 yeast reporter genes (ADE2; HIS3; MEL1, encoding α-galactosidase; and lacZ, encoding β-galactosidase) and transformed into E. coli strain JM109 to recover the plasmid for sequencing. Gene database searches were performed through the National Center for Biotechnology Information (NCBI) using the basic local alignment search tool (BLAST). The longest cDNA clone encoding the Penaeus mon-odon chitin-binding protein (PmCBP) was designated AD-PmCBP-C (see Fig. 4 for sequence). To confirm the screening result, recovered plasmids (containing both BD-WSSV067C and AD-PmCBP-C) were re-trans-formed back into a host strain and plated on SD/-ade/-his/-leu/-trp. Positive interactions were re-tested by cotransforming bait and prey plasmids into Saccar-omyces cerevisiae strain AH109 according to the man-ufacturer’s instructions. Murine p53 (BD-p53) and SV40 large T-antigen (AD-T) that were supplied with the BD Matchmaker library construction and screen-ing kits were used as positive controls. Negative con-trols employed pGBKT7 (BD only) and pGADT7-Rec (AD only).
In vitro transcription and translation. In vitro
tran-scription and translation were performed under RNase-free conditions. BD-WSSV067C and AD-PmCBP-C DNA (1 µg) were respectively added to TNT Quick Master Mix (Promega) containing 35 S-methion-ine to yield c-myc-WSSV067C and HA-PmCBP-C fusion proteins, and the mixture was incubated at 30° for 60 to 90 min according to the manufacturer’s instructions.
Co-immunoprecipitation and SDS-PAGE analysis. In translated c-myc-WSSV067C and in vitro-translated HA-PmCBP-C were combined in a 1.5 ml mi-crocentrifuge tube and incubated at room temperature for 1 h. For the co-immunoprecipitation assay, protein A agarose (Amersham Pharmacia) was washed twice with PBS and restored to a 50% slurry with PBS. The optimal amount of c-myc monoclonal antibody or HA-Tag polyclonal antibody was added to the TNT reac-tants, followed by incubation at room temperature for 1 h. In order to capture the immunocomplex, an optimal amount of protein A agarose bead slurry was added to the TNT reactant/antibody mixture and gently rocked for at least 1 h. The agarose beads were collected by centrifugation at 13 000 × g at 4°C for 5 min and then washed 3 times with modified RIPA buffer (50 mM Tris-HCl, pH 7.4, 1% nonylphenyl-polyethylthylene 40 (NP-40), 0.25% Na-deoxycholate, 150 mM NaCl, 1 mM EDTA, 1 mM phenylmethylsulfonyl fluoride (PMSF), 1 µg ml–1aprotinin, 1 µg ml–1leupeptin, 1 µg ml–1 pep-statin, 1 mM Na3VO4, 1 mM NaF). The beads were re-suspended with PBS and then subjected to SDS-PAGE analysis to check the radiography.
Chitin-binding assay. Commercially available chitin beads (New England BioLabs, catalogue no. 6651S; pre-pared from shrimp Pandulus borealis chitin) were used to test binding. One volume of chitin beads was washed 3 times with 10 volumes of binding buffer (0.5 M NaCl, 10 mM Tris, pH 7.0, 0.05% Triton-X-100) before assay. After in vitro transcription and translation as described above, a 10 µl aliquot was removed (labeled as unbound protein) and the reactants were mixed with pre-washed chitin beads by gently shaking for 4 h at room tempera-ture. The supernatant was removed and kept after cen-trifugation. The pellet was washed twice with NaCl-Tris buffer (5 mM NaCl, 5 mM Tris-Cl, pH 7.0). The beads were resuspended with PBS and then subjected to SDS-PAGE analysis. In this experiment, murine p53 (de-scribed above) was used as the control.
RESULTS
Identification of WSSV structural protein VP53A RT-PCR analysis was used to detect VP53A-specific transcripts in DNase-treated total RNA from shrimp specimens before injection (0 h) and at 2, 4, 6, 8, 12, 18, 24, 36 and 60 h after intramuscular injection of WSSV. After 30 cycles of amplification, the transcripts were detected at low levels by 4 to 8 h post-injection (p.i.), increased at 12 h p.i. and remained fairly constant thereafter (Fig. 1). The overall transcriptional pattern of wssv067C was similar to those of vp26 and vp28 (which are known as late genes encoding WSSV
struc-tural proteins) and unlike the WSSV early genes dnapol, rr1 and rr2 (Chen et al. 2002b).
Immunoelectron microscopy confirmed that VP53A was localized in the envelope of the intact virus (Fig. 2A–D) and was not found when the envelope was removed (Fig. 2G). A similar result showed that an antibody against VP28 — one of the WSSV envelope proteins — also binds to the viral envelope (Fig. 2E,F).
Identification of PmCBP
To identify the shrimp proteins that bound WSSV067, we performed a yeast 2-hybrid screen with a 577 amino acid fragment of WSSV067 that
con-tained the C-terminal transmembrane domain (TMD) (WSSV067C, including the full length of VP53A) as bait. The screening of the cDNA library led to the iso-lation of 15 independent clones that were further sequenced and aligned with the NCBI or expression sequence tag (EST) database (unpubl. data) (Fig. 3). Among the clones, 9 consisted of the same incomplete cDNA that contained a conserved chitin-binding Type 2 domain, which is characterized by having a 6-cys-teine motif and several aromatic residues (Fig. 4). We named this protein Penaeus monodon chitin-binding protein (PmCBP).
The positive clone, PmCBP C terminus (PmCBP-C), was retransformed into the yeast strain AH109 to retest Fig. 1. Temporal transcription analysis of WSSV (A) wssv067,
(B) vp26 and (C) vp28 genes by RT-PCR. Internal controls:
total RNA treated with DNase and amplified with (D) actin-specific primers and (E) intergenic primers. Products were resolved in 1% agarose gel containing 0.5 µg ml–1ethidium
bromide. Lane M is a 100 bp DNA ladder (Promega); other lane headings show time (h) post-injection
Fig. 2. Immunoelectron microscopy analysis of purified virions probed with VP53A antibody. (A), (B), (C) and (D) show that the antibody specifically binds to the viral envelope, not the nucleo-capsid; (E) and (F) show that the antibody against VP28 also binds to the viral envelope; (G) shows that the antibody does not bind to the purified nucleocapsid; (H) shows that pre-immune rabbit
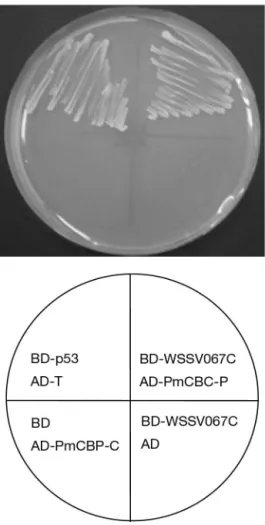
its putative interaction with WSSV067C. Murine p53 (BD-p53) and SV40 large T-antigen (AD-T) were used as positive controls. Negative controls employed pGBKT7 (BD only) and pGADT7-Rec (AD only). Yeast expressing BD-WSSV067C and AD-PmCBP-C formed
colonies on SD medium lacking adenine, histidine, leucine and tryptophan (Fig. 5). The results suggested strongly that, in yeast, WSSV067C (VP53A) and PmCBP-C interact, and that examination of this inter-action under different conditions was warranted.
In vitro binding assay
To confirm the interaction between WSSV067C (VP53A) and PmCBP-C, we carried out an in vitro co-immunoprecipitation. C-myc-WSSV067C, the myc tag-containing fusion protein expressed by BD-WSSV067C, and HA-PmCBP-C, the HA tag-contain-ing fusion protein expressed by AD-PmCBP-C, were expressed by in vitro transcription and translation described in ‘Materials and methods’. The C-myc monoclonal antibody and HA-tag polyclonal antibody coupled with protein A agarose were used to purify the specific proteins. As shown in Fig. 6, protein A agarose conjugated with c-myc antibody could capture
c-myc-1 1301 Bait 725 Clone Actin 1 1 4 9 Myosin EST clone P. monodon chitin-binding protein (PmCBP) Positive no.
Fig. 3. Schematic indicating the localization of bait (BD-WSSV 067C) and the positive clones obtained using the yeast
2-hybrid screen GGCAGTGGATTCACTGGATCCCAGTCTGGTTCATCAGGTGCCACTGGCTTCCAATCTGGA 60 G S G F T G S Q S G S S G A T G F Q S G 20 TCTGGCAGTGGATTCGGTCTTTCATCAGGATTTGGCGGATTCCTGTCTGGTTCAAGTGGT 120 S G S G F G L S S G F G G F L S G S S G 40 GCTGGATTTGGGGCAGCAGGTGGCAGTGGACTTCAGTTTGGTTCAGAATCAGATGATGTA 180 A G F G A A G G S G L Q F G S E S D D V 60 TTTGGTATCTTTGAGCCTCTGAATCTTCCTGCAGAAGCCACTCGCTTGCTTGGAAAAATC 240 F G I F E P L N L P A E A T R L L G K I 80 TCAACATCATTTGCCTGCCTTGATCGGCCTTATGGATATTATGCTGATGAAGAAAACTCC 300 S T S F A C L D R P Y G Y Y A D E E N S 100 TGTCACATCTTCCACATCTGTTACCCTGCTCTCTTTGCTAATGGTGTTATCGAGACCTCC 360 C H I F H I C Y P A L F A N G V I E T S 120 CAGTACAGCTTCCTGTGTGGTGAGGGTTCTCGATTCGACCAAAAGGAGCTTACTTGCGTG 420 Q Y S F L C G E G S R F D Q K E L T C V 140 GCAGAATCAGAAGCAATTCCTTGCCAGGAATCTTCCAACTTCTTTTTCAAGAACGAACAG 480 A E S E A I P C Q E S S N F F F K N E Q 160 TTTGGTCTTCCAAAGGAAAAGATTTAAGCCATTTCAGCCCACTTTGAGGACTTGATGTAT 540 F G L P K E K I * 168 CGCCCAATGAACAACTCATGCGAACATTTTGGATAAATAAAATGG 585 Fig. 4. Primary structure of the longest cDNA clone, designated AD-PmCBP-C, encoding the Penaeus monodon chitin-binding
protein (PmCBP). Chitin-binding Type 2 domain is in bold. In this domain, cysteine residues (shaded) and aromatic residues
(under-lined) are indicated
Fig. 5. Yeast 2-hybrid screen. Transformed cells were se-lected on synthetic dropout (SD/-ade/-his/-leu/-trp) medium.
WSSV067C (Lane 1) but not HA-PmCBP-C (Lane 6), and protein A agarose conjugated with HA antibody could capture HA-PmCBP-C (Lane 2) but not WSSV067C (Lane 5). After incubating c-myc-WSSV067C together with HA-PmCBP-C, protein A agarose conjugated with c-myc antibody could capture both c-myc-WSSV067C and HA-PmCBP-C (Lane 3), and protein A agarose conjugated with HA antibody could also capture both c-myc-WSSV067C and HA-PmCBP-C (Lane 4).
Chitin-binding assay
A chitin-binding assay was performed to determine whether PmCBP was capable of binding to chitin in the form of commercially available chitin beads prepared from shrimp Pandulus borealis chitin. HA-PmCBP-C, expressed by the in vitro transcription and translation system, bound to the chitin beads (Fig. 7) when com-pared with the control group p53 (Fig. 7B, Lane 5). PmCBP-C containing the conserved chitin-binding Type 2 domain was capable of binding to chitin (Fig. 7A, Lane 5). This indicates that the PmCBP is a chitin-binding protein.
Tissue tropism and expression pattern of PmCBP RT-PCR was used to reveal the tissue tropism of PmCBP. PmCBP was expressed in almost all organs including the stomach, gills, midgut, haemolymph, lymphoid organ, hepatopancreas, haematopoietic tis-sue, pleopods, nervous tissue and heart (Fig. 8A). Time-course RT-PCR analysis of total RNA samples from WSSV-infected shrimps revealed no significant changes in PmCBP during infection (Fig. 8C). This indicated that PmCBP was constitutively expressed.
DISCUSSION
As shown in Fig. 3, the screening of the cDNA library led to the isolation of 2 kinds of cytoskeletal Fig. 6. SDS-PAGE analysis of WSSV067C
co-immunoprecipi-tation with PmCBP-C. 35S-methionine was included in the
translation mixture to generate radioactively labeled products WSSV067C and HA-PmCBP-C. Lane 1: c-myc-WSSV067C + c-myc antibody (c-myc Ab, ø); Lane 2: HA-Pm-CBP-C + HA antibody (HA Ab, ø); Lane 3: c-myc-WSSV067C + HA-PmCBP-C + c-myc antibody; Lane 4: c-myc-WSSV067C + HA-PmCBP-C + HA antibody; Lane 5: c-myc-WSSV067C + HA antibody; Lane 6: HA-PmCBP-C + c-myc antibody
Fig. 7. SDS-PAGE analysis showing PmCBP-C binding with chitin beads. 35S-methionine was included in the translation
mixture to generate radioactively labeled products (A) Pm-CBP-C and (B) murine p53. Lane 1: unbound proteins; Lane 2: supernatants after centrifuging; Lanes 3 and 4: NaCl-Tris
proteins: myosin and actin. Similar results were revealed by Xie & Yang (2005) who used VP26 as bait. Some viruses, such as polyomavirus simian virus 40, vaccinia virus, herpesvirus, adenovirus etc. have been reported to exploit the host cytoskeleton in often very fascinating ways in order to facilitate important aspects of their life cycle such as entry, egress and intercellular spread (Sasaki et al. 1995, Favoreel et al. 2005, Gouin et al. 2005, Lehmann et al. 2005). Much of the protein-protein interaction with the host cytoskeleton occurs via virus structural proteins. Thus, it is possible that the WSSV envelope protein WSSV067C/VP53A could interact weakly with the cytoskeleton of host cells, but its role in viral infection, replication or spread remains to be tested.
Chitin is an important structural component of the cell walls of fungi and the exoskeletons of many
inver-tebrates, such as crustaceans, insects and nematodes. An arthropod chitin-binding domain has been charac-terized in some chitinases and peritrophic membrane proteins (Shen & Jacobs-Lorena 1999). However, the chitin-binding Type 2 domain with 6 conserved cys-teine residues and several conserved aromatic residues is not found in the cuticular proteins that are also CBPs. Other well characterized CBPs are agglu-tinins/lectins that exist in almost all living organisms. They are usually proteins or glycoproteins without cat-alytic activity that have the ability to bind to specific carbohydrates expressed on different cell surfaces (Marques & Barracco 2000). Agglutinin molecules may occur on the surfaces of viruses, bacteria, yeasts, proto-zoans and cells of both animals and plants (Sharon & Lis 1989). Most of the current research emphasizes the possible role of lectins as non-self-recognition mole-cules in vertebrate and invertebrate immunity (Vasta et al. 1999, Wilson et al. 1999). From studies of lectins conducted on different species of penaeids, the major biological activities were determined to be opsonic activity and agglutination of bacteria (reviewed by Marques & Barracco 2000). Although the term ‘chitin-binding protein’ denotes a family of proteins contain-ing one or more chitin-bindcontain-ing domains, it must be emphasized that the binding affinity of these proteins is not restricted to chitin, but may extend to various complex glycoconjugates that contain β-1,4-linked biopolymers of N-acetylglucosamine (GlcNAc), sialic acid (acetyl-D-neuraminic acid; NeuNAc) or N-acetyl-D-galactosamine (GalNAc) as building blocks (Raikhel et al. 1993, Marques & Barracco 2000). Func-tions of CBPs can thus range from simple binding (e.g. in the case of lectins) to chitinase activity, or even to antimicrobial and antifungal activities as in the case of tachycitin (Kawabata et al. 1996).
The chitin-binding Type 2 domain is mainly found in animal and baculovirus proteins, whereas Type 1 is found in plants and fungi. Type 2 is characterized by having a 6-cysteine motif, C-x(13, 20)-C-x(5, 6)-C-x(9,19)-C-x(10,14)-C-x(4,14)-C, and several aromatic residues. Proteins known to contain a chitin-binding Type 2 domain include animal class-II chitinase, tachycitin, peritrophin-A, Drosophila tequila protein and Drosophila hemolectin. All participate in immune responses, except peritrophin-A, which is a component of peritrophic matrices. These are acellular linings in the gut of most insects that comprise proteins and sugar polymers (Suetake et al. 2000, Tjoelker et al. 2000, Gaines et al. 2003).
Alignment of our new CBP to standard databases revealed no similar proteins. The full length PmCBP could not be obtained, even though many methods such as rapid amplification of the cDNA 5’ end (5’ RACE) were tried. Nevertheless, the characteristics of Fig. 8. Tissue tropism and temporal transcription analysis of
CBP. RNA samples were extracted from organs of Peneaus monodon that were not infected with WSSV and amplified
with (A) PmCBP-specific primer sets PmCBP-RT-F1/PmCBP-RT-R1 or (B) actin-specific primers. (C) Time-course transcrip-tion analysis after WSSV injectranscrip-tion using the same tested sam-ples as in Fig. 1. Products were resolved in 1% agarose gel containing 0.5 µg ml–1ethidium bromide. Lane M is a 100 bp
DNA ladder (Promega). Other lane headings show (A,B) Lane 1: stomach; Lane 2: gills; Lane 3: midgut; Lane 4: haemolymph; Lane 5: lymphoid organ; Lane 6: hepatopan-creas; Lane 7: haematopoietic tissue; Lane 8: pleopods; Lane 9: nervous tissue; Lane 10: heart; (C) time (h) post-injection
PmCBP can be inferred, owing to the partial C termi-nus. Because tissue tropism assays by RT-PCR revealed that PmCBP is a constitutively expressed gene with a chitin-binding Type 2 domain, we specu-late that it is not a cuticular protein. It might function like chitinase or lectin. If so, VP53A might play a role in the shrimp immune response. Further research is required to determine whether PmCBP is present on the cell surface or is a secreted protein.
Acknowledgements. This study was supported by the National Council grants NSC 93-2311-B-040-011 and NSC 94-2311-B-040-011. We are indebted to Mr. Vince J. Genis and Miss Angela Wang for their helpful criticism of the manuscript.
LITERATURE CITED
Chen LL, Leu JH, Huang CJ, Chou CM, Chen SM, Wang CH, Lo CF, Kou GH (2002a) Identification of a nucleocapsid protein (VP35) gene of shrimp white spot syndrome virus and char-acterization of the motif important for targeting vp35 to the nuclei of transfected insect cells. Virology 293 44–53 Chen LL, Wang HC, Huang CJ, Peng SE and 8 others (2002b)
Transcriptional analysis of the DNA polymerase gene of shrimp white spot syndrome virus (WSSV). Virology 301: 136–147
Favoreel HW, Van Minnebruggen G, Adriaensen D, Nauwynck HJ (2005) Cytoskeletal rearrangements and cell extensions induced by the US3 kinase of an alphaherpesvirus are asso-ciated with enhanced spread. Proc Natl Acad Sci USA 102: 8990–8995
Flegel TW (1997) Special topic review: major viral diseases of the black tiger prawn (Penaeus monodon) in Thailand. World J Microbiol Biotech 13:422–433
Gaines PJ, Walmsley SJ, Wisnewski N (2003) Cloning and char-acterization of five cDNAs encoding peritrophin-A domains from the cat flea, Ctenocephalides felis. Insect Biochem. Mol Biol 33:1061–1073
Gouin E, Welch MD, Cossart P (2005) Actin-based motility of intracellular pathogens. Curr Opin Microbiol 8:35–45 Kawabata S, Nagayama R, Hirata M, Shigenaga T and 6 others
(1996) Tachycitin, a small granular component in horseshoe crab hemocytes, is an antimicrobial protein with chitin-bind-ing activity. J Biochem 120:1253–1260
Lehmann MJ, Sherer NM, Marks CB, Pypaert M, Mothes W (2005) Actin- and myosin-driven movement of viruses along filopodia precedes their entry into cell. J Cell Biol 170: 317–325
Liu WJ, Yu HT, Peng SE, Chang YS and 9 others (2001) Cloning, characterization and phylogenetic analysis of a shrimp white spot syndrome virus (WSSV) gene that encodes a protein kinase. Virology 289:362–377
Lo CF, Kou GH (1998) Virus associated white spot syndrome of shrimp in Taiwan: a review. Fish Pathol 33:365–371 Lo CF, Leu JH, Ho CH, Chen CH and 8 others (1996a) Detection
of baculovirus associated with white spot syndrome (WSBV) in penaeid shrimps using polymerase chain reaction. Dis Aquat Org 25:133–141
Lo CF, Ho CH, Peng SE, Chen CH and 7 others (1996b) White spot syndrome baculovirus (WSBV) detected in culture and captured shrimp, crabs and other arthropods. Dis Aquat Org 27:215–225
Lo CF, Hsu HC, Tsai MF, Ho CH, Peng SE, Kou GH, Lightner DV
(1999) Specific genomic fragment analysis of different geo-graphical clinical samples of shrimp white spot syndrome virus. Dis Aquat Org 35:175–185
Lu L, Kwang J (2004) Identification of a novel shrimp protein phosphatase and its association with latency-related ORF427 of white spot syndrome virus. FEBS Lett 577:141–146 Marques MRF, Barracco MA (2000) Lectins, as
non-self-recognition factors, in crustaceans. Aquaculture 191: 23–44
Raikhel NV, Lee HI, Broekaert WF (1993) Structure and function of chitin-binding proteins. Annu. Rev. Plant Physiol Plant Mol Biol 44:591–615
Sasaki H, Nakamura M, Ohna T, Matsuda Y, Yuda Y, Nonomura Y (1995) Myosin-actin interaction plays an important role in human immunodeficiency virus type 1 release from host cells. Proc Natl Acad Sci USA 92:2026–2030
Sharon N, Lis H (1989) Lectins as cell recognition molecules. Sci-ence 246:227–234
Shen Z, Jacobs-Lorena M (1999) Evolution of chitin-binding pro-teins in invertebrates. J Mol Evol 48:341–347
Suetake T, Tsuda S, Kawabata S, Miura K, Iwanaga S, Hikichi K, Nitta K, Kawano K (2000) Chitin-binding proteins in inverte-brates and plants comprise a common chitin-binding struc-tural motif. J Biol Chem 275:17929–17932
Tjoelker LW, Gosting L, Frey S, Hunter CL, Trong HL, Steiner B, Brammer H, Gray PW (2000) Structural and functional defin-ition of the human chitinase chitin-binding domain. J Biol Chem 275:514–520
Tonganunt M, Phongdara A, Chotigeat W, Fujise K (2005) Iden-tification and characterization of syntenin binding protein in the black tiger shrimp Penaeus monodon. J Biotechnol 120: 135–145
Tsai MF, Lo CF, van Hulten MCW, Tzeng HF and 6 others (2000) Transcriptional analysis of the ribonucleotide reductase genes of shrimp white spot syndrome virus. Virology 277:92–99 Tsai JM, Wang HC, Leu JH, Hsiao HH, Wang AHJ, Kou GH, Lo
CF (2004) Genomic and proteomic analysis of 39 structural proteins of shrimp white spot syndrome virus. J Virol 78: 11360–11370
Tsai JM, Wang HC, Leu JH, Wang AHJ, Zhuang Y, Walker PJ, Kou GH, Lo CF (2006) Identification of the nucleocapsid, tegument, and envelope proteins of the shrimp white spot syndrome virus virion. J Virol 80:3021–3029
Vasta GR, Quesenberry M, Ahmed H, O’Leary N (1999) C-type lectins and galectins mediate innate and adaptive immune functions: their roles in the complement activation pathway. Dev Comp Immunol 23:401–420
Wang CH, Lo CF, Leu JH, Chou CM and 6 others (1995) Purifi-cation and genomic analysis of baculovirus associated with white spot syndrome (WSBV) of Panaeus monodon. Dis Aquat Org 23:239–242
Wang Z, Chua HK, Gusti AA, He F, Fenner B, Manopo I, Wang H, Kwang J (2005) RING-H2 protein WSSV249 from white spot syndrome virus sequesters a shrimp ubiquitin-conjugat-ing enzyme, PvUbc, for viral pathogenesis. J Virol 79: 8764–8772
Wilson R, Chen C, Ratcliffe NA (1999) Innate immunity in insects: the role of multiple, endogenous serum lectins in the recognition of foreign invaders in the cockroach, Blaberus discoidalis. J Immunol 162:1590–1596
Wongteerasupaya C, Vickers JE, Sriurairatana S, Nash GL and 6 others (1995) A non-occluded, systemic baculovirus that occurs in cells of ectodermal and mesodermal origin and causes high mortality in black tiger prawn Penaeus mon-odon. Dis Aquat Org 21:69–77
Xie X, Yang F (2005) Interaction of white spot syndrome virus VP26 protein with actin. Virology 336:93–99
Editorial responsibility: Timothy Flegel, Bangkok, Thailand
Submitted: August 16, 2006; Accepted: November 12, 2006 Proofs received from author(s): February 22, 2007