http://lup.sagepub.com
Lupus
DOI: 10.1177/0961203308095450 2009; 18; 206
Lupus
YH Hong, CJ Huang, SC Wang and BF Lin
MRL-lpr/lpr mice
The ethyl acetate extract of alfalfa sprout ameliorates disease severity of autoimmune-prone
http://lup.sagepub.com/cgi/content/abstract/18/3/206 The online version of this article can be found at:
Published by:
http://www.sagepublications.com
can be found at:
Lupus
Additional services and information for
http://lup.sagepub.com/cgi/alerts Email Alerts: http://lup.sagepub.com/subscriptions Subscriptions: http://www.sagepub.com/journalsReprints.nav Reprints: http://www.sagepub.co.uk/journalsPermissions.nav Permissions: http://lup.sagepub.com/cgi/content/refs/18/3/206 Citations
PAPER
The ethyl acetate extract of alfalfa sprout ameliorates
disease severity of autoimmune-prone MRL-
lpr/lpr mice
YH Hong, CJ Huang, SC Wang and BF Lin
Department of Biochemical Science and Technology, Institute of Microbiology and Biochemistry, College of Life Science, National Taiwan University, Taipei, Taiwan
Previous study showed that soy isoflavone supplement alleviates disease severity in autoimmune-prone mice. As the ethyl acetate extract of alfalfa sprout (AS) has selective oestrogenic and anti-inflammatory activity, this study evaluated the effects of alfalfa sprout ethyl acetate extract (ASEA) on disease severity of systemic lupus erythematosus, using autoimmune-prone female MRL-lpr/lpr mice. In Experiment 1, five groups of 12-week-old female mice were per oral treated with vehicle (control), lyophilized AS (550 mg wt/kg BW), ASEA (ASEA, 25 mg/kg BW), coumestrol (CUM, 0.075 mg/kg BW) and tamoxifen (TAM, 0.375 mg/kg BW) as the positive control. The onset of proteinuria was delayed, and the life span was significantly longer in the ASEA and TAM groups but neither in the AS nor in the CUM groups, compared to the control. To examine the changes in the immunological parameters related to disease process, three more groups of MRL-lpr/lpr female mice (control, ASEA and TAM) were fed in a similar manner for 6 weeks in the Experiment 2. Flow cytometric analysis of splenocytes showed a significantly lower percentage of activated T cells in the ASEA and TAM groups. The ex-vivo interferon-γ and interleukin (IL)-4 production from splenocytes and tumour necrosis factor-α and IL-1β production from peritoneal exudate cells were also significantly lower in the ASEA group compared with the control. The ASEA group also had less severe glomerulonephritis. Thus, ASEA attenuated cytokine and inflammatory responses of self-reactive lymphocytes, decreased the disease severity, increased survival and life span of the autoimmune-prone MRL-lpr/lpr mice, suggesting a potential of ASEA in the treatment of auto-immune diseases. Lupus(2009) 18, 206–215.
Key words: alfalfa sprout extract; autoimmune; IFN-γ; inflammation; systemic lupus erythematosus
Introduction
Systemic lupus erythematosus (SLE) is a heteroge-neous disorder with the feature of a multisystem inflammatory disease and the production of a variety of autoantibodies. Although the pathogenic mechan-isms are not entirely clear, environmental factors have been suggested to play a role in the aetiology and the exploration of which may help to advance the strate-gies to treat SLE.1The effect of dietary modifications
has been extensively studied in lupus animal models such as MRL-lpr/lpr and NZB×NZW F1 mice. Restriction of calorie, protein and fat, or addition
of n-3 fatty acid or vitamin E supplementation in the oxidised oil diet prolong the life span of lupus mice.2–4 In addition, Chinese herb such as Ganoderma tsugae, food component such as indole-3-carbinol, a synthetic oestrogen receptor (ER)-binding molecule such as bisphenol A were shown to delay onset of disease symptom or even prolong the life span of lupus mice. These results suggest that dietary change or sup-plementation may exert immunomodulatory effect on SLE.5–7
It is well known that SLE is nine times more preva-lent in women than in men,8,9and oestrogen enhanced
disease progress in animal model of SLE.10,11
Phyto-estrogens are plant-derived compounds that can inter-act with ER to exert oestrogenic/anti-oestrogen activities.12,13 In recent years, phytoestrogen
supple-ment, such as soy isoflavones, has become widely used as alternatives to alleviate menopausal syndrome or prevent chronic disease,14and thus, its oestrogenic
Lupus (2009) 18, 206–215
http://lup.sagepub.com
© 2009 SAGE Publications Los Angeles, London, New Delhi and Singapore 10.1177/0961203308095450
Correspondence to: Bi-Fong Lin, PhD, Department of Biochemical Science and Technology, Institute of Microbiology and Biochemistry, College of Life Science, National Taiwan University, No. 1, Roosevelt Road, Section 4, Taipei 10617, Taiwan, Republic of China.
Email: [email protected]
Received 1 April 2009; accepted 18 June 2008
at NATIONAL TAIWAN UNIV LIB on June 3, 2009 http://lup.sagepub.com
or anti-oestrogenic effect on SLE is worth examining. However, phytoestrogens have potential effects on immune modulation through the inhibition of inflam-matory responses which may be beneficial for SLE.15
Our previous study has shown that dietary supple-mentation of soy isoflavone did not aggravate but alle-viate disease severity in MRL-lpr/lpr mice. We also showed that the soy isoflavone supplement used in that study had higher affinity for ERβ than for ERα, the two isoforms of ERs in human.16 The ER
med-iates the function of oestrogen by binding to specific ligands and activating the transcription of specific tar-get gene. Some phytoestrogens with higher affinity for ERβ toward ERα are considered ‘natural’ selective oestrogen receptor modulators (SERMs) and may play protective roles in inflammatory diseases.17
Therefore, a transactivation assay that can measure the ligand-dependent transcription of reporter gene through ERα or ERβ has been instrumental in our screening of phytoestrogens from various food mate-rials extracted with methanol, ethyl acetate (EA) or n-hexane.18Among the plant foods we screened, AS
extracted in EA is one of the samples that have the potential for SERM activity.
AS contains high levels of dietary fibre and prevents hypercholesterolemia and atherosclerosis in rats and monkeys.19,20It has also been regarded as a functional
food to modulate lipid profile because it is enriched with phytoestrogens. In addition, AS extract has the potential to reduce pain from dermatitis in traditional remedy.21However, previous reports show that alfalfa
seed or sprout provokes SLE-like disease in monkeys and exacerbates disease severity in patients with SLE who ingested alfalfa tablet.22–24 Later, this causative
component has been identified as L-canavanine.25
However, both basic and clinical studies on alfalfa and the disease process of SLE are quite limited in that even the most recent review articles are based on findings reported more than 10 years ago.26 It
will be interesting to re-evaluate how alfalfa affects SLE in more detail based on more up-to-date knowl-edge the anti-inflammatory effect.
In this study, we first used MRL-lpr/lpr mice to evaluate the effects of AS (lyophilized powder), alfalfa sprout ethyl acetate extract (ASEA) and cou-mestrol (CUM), a known phytoestrogen in AS, on the disease course by following the proteinuria and life span of the lupus mice. Mice treated with tamoxi-fen (the TAM group), a potent oestrogen antagonist that was shown to alleviate disease severity in lupus mice, served as a positive control.27Part of the animal
experiment was repeated to examine the phenotypes of splenocytes, cytokine productions from peritoneal exudate cells (PEC) and splenocytes, autoantibody
productions and renal pathology of the ASEA-treated mice.
Materials and methods
Lyophilized AS and its extract
Fresh AS (Medicago sativa L.) were purchased from Goboul-Grange LTD (Taipei, Taiwan). AS were freeze-dried and finely grounded into the powder. The proximate compositions of this powder were 33.6% carbohydrate, 49.6% protein, 2.6% lipid, 10.5% water and 3.7% ash. AS powders were extracted with EA (1:40, w/v, g/mL) by stirring at room temperature for 2 days. The EA extract (ASEA) was obtained by removing the solvent in a rotary evaporator, with a yield of 43.1 mg/g. The ASEA was stored at−20 °C and was dissolved in sun-flower oil (SO) for tube feeding in the mice experi-ment. In ER transactivation assay, the extract was dissolved in absolute ethanol for cell treatment (final ethanol concentration never exceeded 0.1% in medium).
Experimental animals
Six-week-old-female MRL-lpr/lpr mice were purchased from the Animal Center of the College of Medicine at the National Taiwan University (Tai-pei, Taiwan). The mice were kept in stainless steel wire cages in a room with controlled temperature of 23 ± 2 °C and 12-h light–dark cycle. They were fed a non-purified diet (Lab Rodent Chow 5001, Ralston Purina Inc., St Louis, Missouri, USA) for adapta-tion, and then switched to soy-free and oestrogen-free AIN-76 diet, in which corn oil is substituted by SO. At age 12 weeks, the diet treatment was started.
Diet and supplementation of AS and its extract In the first experiment, 60 mice were assigned to the following five groups with similar average body weights, urinary protein levels and serum levels of anti–double stranded (ds) DNA immunoglobulin (Ig) G at age 12 weeks. The five groups received the following diet treatments: control group (tube fed with 50μL SO/day); the AS group (AIN-76 diet containing 0.55% AS powders and tube fed 50μL SO/day); the ASEA group (tube fed 25 mg/kg BW ASEA in 50μL SO/day); the CUM group (tube fed 0.075 mg/kg BW CUM in 50μL SO/day) and the TAM group (tube fed 0.375 mg/kg BW TAM in 50μL SO/day). Mice were tube fed six days per week. The mice of the AS group averagely received 550 mg/kg BW AS powder per day
from the diet. TAM was used as a positive control, and the dose used was according to the previous study that showed its alleviation of disease course in MRL-lpr/lpr mice.27 In the second experiment, the
control, ASEA and TAM groups were repeated to further investigate the immune modulation, and each group consisted of 14–18 mice. After 6 weeks’ supple-mentation, mice were sacrificed to determine the renal histology, phenotype of splenocytes and cellular cyto-kine production.
In both experiments, food intake, body weight and life span of the mice were explicitly recorded. In addi-tion, the urine and serum of the mice were regularly collected for further assay. Animal care and handling conformed to the National Institute of Health’s Guide for the Care and Use of Laboratory Animals.28
Assay of anti-dsDNA antibodies
Mice were retro-orbitally bled and serum samples were analysed for autoantibodies. Serum levels of anti-dsDNA IgG, IgG1 and IgG2a were measured by enzyme-linked immunosorbent assay (ELISA). The 96-well plates were coated with methylated bovine serum albumin (mBSA, Sigma Chemical Company, St Louis, Missouri, USA). After overnight incuba-tion, the plates were washed and calf thymus dsDNA was coated for overnight at 4 °C. The plates were washed and blocked with gelatin solution for 2 h. After washes, properly diluted serum samples were added in the plates. Two hours later, the plates were washed and peroxidase-conjugated Fcγ fragment-specific antibodies for IgG (Jackson Immu-noResearch, West Grove, Pennsylvania, USA), bioti-nylated A85-1 monoclonal antibodies for IgG1 (BD Pharmingen, San Diego, California, USA) or biotiny-lated R19-15 monoclonal antibodies for IgG2a (BD Pharmingen) were added for 1-h incubation. After washing, the plates for IgG1 and IgG2a assay were added with horseradish peroxidase-conjugated strep-tavidin (Pierce, Rockford, Illinois, USA) for addi-tionally 30-min incubation. After washing, the plates were incubated with ABTS [2, 20-azino-bis (3-ethylbenzthazoline-6-sulfonic acid); Sigma Chemi-cal Company, St Louis, Missouri, USA]. Absorbance was measured at 405 nm [optimal density (OD)] within 20 min.
Assay of cytokine production from immune cells In the second experiment, mice were sacrificed at age 18 weeks. Splenocytes and PEC were collected under sterile conditions. The culture medium used in the experiments was phenol-free RPMI-1640 supplemented with 10% charcoal-stripped fetal
bovine serum. Splenocytes, at a concentration of 5× 106 cells/mL/well, were treated without or with
5 mg/L concanavalin (Con) A (Sigma) for 24 h. PEC were seeded at 1.5× 106 cells/mL/well and treated
without or with 10 mg/L lipopolysaccharide (LPS, Sigma) for 48 h. After incubation, cell supernatants were collected for measurement of cytokine production.
The production of cytokines, including interleukin (IL)-2, interferon (IFN)-γ, IL-4, IL-10 (splenocytes), tumour necrosis factor (TNF)-α and IL-6 (PEC), was assayed by using commercial ELISA kit. Briefly, anti-IL-2, IFN-γ, IL-4, IL-6 (BD Pharmingen), TNF-α or IL-10 (R & D, Minneapolis, Minnesota, USA) antibodies were coated to 96-well plate and incubated at 4 °C overnight. After washes, the wells were blocked with 200μL blocking solution (phos-phate buffer saline (PBS) buffer containing 1% bovine serum albumin; Sigma) for 1 h. The plates were washed and added with properly diluted cell superna-tant for 2-h incubation. The plates were then washed before adding biotin-conjugated anti-IL-2, IFN-γ, IL-4, IL-6, TNF-α or IL-10 antibodies for 2 h. After washing, horseradish peroxidase-conjugated strepta-vidin (Pierce) was added and incubated for 30 min. After washing, the plates were incubated with ABTS or tetramethylbenzidine (Clinical Science Products, Mansfield, Massachusetts, USA) and measured for absorbance at 405 nm or 620 nm within 20 min. The data were calculated according to the cytokine stan-dard curve.
Flow cytometry
Phenotypic analysis of splenocytes was performed by flow cytometry. Cells at a concentration of 2× 106
were suspended in 0.1 mL of PBS (with 0.1 % sodium azide) and incubated at 4 °C for 30 min with predeter-mined optimal concentration of appropriate fluoros-cein isothiocyanate- or phycoerythrin-conjugated monoclonal antibodies (BD Pharmingen, San Diego, California, USA). The cells were washed and resus-pended in 0.5 mL of PBS (with 0.1% sodium azide) and subjected to fluorescence-activated cell sorting (FACScan analysis). A total of 10,000 cells were counted, the frequency of each cell surface marker was determined using appropriate software (FACScan, Becton Dickinson, Mountain View, California, USA).
Proteinuria and renal histology
Proteinuria was measured using Bio-Rad Protein Bradford assay to determine protein levels in urine. Briefly, 10μL of properly diluted urine sample or
Autoimmune and diet YH Honget al.
208
Lupus
at NATIONAL TAIWAN UNIV LIB on June 3, 2009 http://lup.sagepub.com
BSA at concentrations of 0.2–0.8 g/L as standards were added to 96-well plate. Then 200μL dye reagent (diluted five fold with deionised water) was added. After mixing for 5 min at room temperature, the absorbance was measured at 595 nm. Proteinuria was defined as a level of urinary protein higher than 1 g/L for two consecutive weeks. For renal histology, kidney samples were fixed in 3% formaldehyde, and 5-μm kidney sections were cut and stained with hae-matoxylin and eosin. The sections were observed for glomerular nephritis included glomerular enlarge-ment, hypercellularity, mesangial cell proliferation and fibrinoid necrosis.
Assay of oestrogenic activity
Oestrogenic activity was measured by the ER transac-tivation assay as described in the previous study.16
Briefly, 1.25× 105cells/mL CHO-K1 cells (CCRC
60006) were seeded at 96-well plate and transfected with 0.3μg of the chimeric receptor construct pBK-CMVGal4-hERα (or β) and pBK-CMV-(UAS)4
-tk-alkaline phosphatase (AP) (4:1 or 5:1) in 100μL serum-free medium OPTI-MEM (Gibco BRL, Gaithersburg, Maryland, USA) containing 1μL lipo-fectamine per well. After 5 h, the medium was chan-ged to Ham’s F-12 medium containing 10% TCM and 1 nM 17β-estradiol (E2, as the positive control) or the appropriate concentration of ASEA (0.4–50 μg/mL) or CUM (0.01–1 μM). After 48-h incubation, 20 μL (ERα) or 10 μL (ERβ) culture medium was trans-ferred to a new 96-well plate and mixed with 200μL of SEAP (secreted AP) assay solution. Finally, the absorbance was read at 405 nm for 15 min. Fold acti-vation was calculated by taking the AP activity of vehicle-treated cells as 1. Experiments were repeated four times.
Statistical analysis
Differences in cellular and serological parameters between the treatment and the control groups were analysed by Student’s t-test, whereas transactivation activity among groups was analysed by one-way ANOVA and Duncan’s multiple range test, using the SAS software program (SAS/STAT version 8; SAS Institute, Cary, North Carolina, USA). Data were presented as means ± standard deviation or standard error (SE) for each group. The data of the proteinuria cumulative occurrence and survival were analysed between two different curves using the Cox proportional hazards regression test (STATA version 6.0; Stata Corp., College Station, Texas, USA). Statis-tical significance was considered at a P value≤ 0.05.
Results
ASEA delayed the occurrence of proteinuria and increased mice survival
To investigate the effect of AS and its EA extract on SLE, MRL-lpr/lpr mice were fed AIN-76 diet and sup-plemented with either lyophilized AS powder, ASEA or CUM beginning at 12 weeks of age. There was no significant difference in body weight, food intake and food efficiency among five groups (data not shown). Figure 1A shows that proteinuria developed early in the control and the AS groups. At age 18 weeks, almost all mice of the control and the AS groups had protein-uria, but only one-third of the ASEA group had pro-teinuria, less than those of the TAM and CUM groups. The cumulative occurrence of proteinuria in the ASEA group (P = 0.047) and the TAM group (P = 0.049) were significantly lower than the control group.
Figure 1 Occurrence of proteinuria (A) and survival curve (B) of MRL-lpr/lpr mice supplemented without (control) or with alfalfa sprouts (AS), alfalfa sprout ethyl acetate extracts (ASEA), coumestrol (CUM) or tamoxifen (TAM). There were 12 mice, at the age 12 weeks, started on each supplement diet. The data were analysed by COX’s proportion hazards regression test. Lower occurrence of proteinuria (P = 0.047) and increased survival rate (P = 0.012) were in the ASEA group compared to those in the control group.
The survival rate of mice in the control group decreased with age and was approximately 50% at the age of 22 weeks (Figure 1B). At this age, 90% of mice in the ASEA group still survived, higher than the TAM and the CUM groups. The survival rate of the ASEA group was higher than that of the control group (P = 0.012, Cox proportion hazards regression test). The average life span in the ASEA group (199 ± 14 days) and the TAM group (189 ± 14 days) were significantly longer than the control group (152 ± 10 days) (P < 0.05). The life span and the sur-vival rate of the mice in the AS and the CUM groups were not significantly different from that of the control group.
Phenotypic analysis of surface marker expression of splenocytes
To examine the effects of ASEA on immune responses, three more groups of MRL-lpr/lpr mice
were treated in a similar manner to the control, ASEA and TAM groups of the Experiment 1 for 6 weeks. Phenotypic analysis of surface marker expression of splenocytes showed that the expressed level of CD69+T cells was significantly lower in the
ASEA and the TAM groups (P = 0.04 and 0.03, respectively, Figure 2). The CD69 molecule is an acti-vated marker for T cells. The percentage of B cells (B220+) tended to be lower in the ASEA group
(P = 0.09). No significant difference in the distribution of CD4+T cells, CD8+T cell or CD4−CD8−double
negative (DN) T cells was noted among these three groups.
Cytokines production by mitogen-stimulated PEC and splenocytes
Amount of cytokines secreted from LPS-stimulated PEC and Con A-stimulated splenocytes are shown in Table 1. The production of TNF-α was slightly lower
Figure 2 Phenotypic analysis of surface marker expression of splenocytes in MRL-lpr/lpr female mice supplemented without (control), or with alfalfa sprout ethyl acetate extract (ASEA) or tamoxifen (TAM). Splenocytes were collected from mice at age 18 weeks. Each group contains 14–18 mice. (A) Representative examples of two-colour fluorescence-activated cell sorting (FACScan analysis) of CD69+T populations of splenocytes from these three groups. (B) Cell populations by phenotypic analysis include CD3+(T), CD4+T,
CD8+T, CD4–CD8–double negative (DN) T, CD69+ T and B220+cells. The average values were represented means ± SE. The
significant difference between the control group and the experimental group was analysed by Student’s t-test, *P < 0.05. Autoimmune and diet
YH Honget al.
210
Lupus
at NATIONAL TAIWAN UNIV LIB on June 3, 2009 http://lup.sagepub.com
in the ASEA group (P = 0.09) and significantly lower in the TAM group. IL-1β was significantly lower in the ASEA group and slightly decreased in the TAM group (P = 0.07). IL-6 level was not significantly dif-ferent among the groups. The splenocytes of ASEA-supplemented group produced significantly lower IFN-γ and IL-4 while stimulated with 5 mg/L Con A. There was no significant difference in the level of IL-6, IL-2 and IL-10 among these three groups, except a slight decrease in IL-10 production of TAM group (P = 0.06).
Effects of ASEA on autoantibodies production and lupus nephritis
The serum levels of autoantibodies were monitored regularly to follow up the disease progress. Serum levels of anti-dsDNA IgG, anti-dsDNA IgM and anti-cardiolipin IgG increased with age but were not statistically different among the groups because of individual variation (data not shown). Further, the level of IgG2a anti-dsDNA antibody tended to be lower in ASEA- and TAM-treated groups but not sta-tistically significant (P = 0.26 and 0.22, respectively, Figure 3).
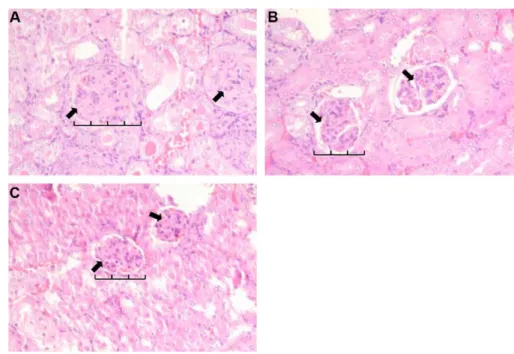
The renal histology shown in Figure 4 indicates that enlarged glomeruli and the proliferation of mesangial cells in the ASEA group were less severe than those in the control group. Less inflammatory cell infiltration in the kidneys of the ASEA and TAM groups was likewise noted.
Estrogenic activity of ASEA and CUM
To determine the oestrogenic activity of ASEA and CUM, ASEA or CUM was added to CHO-K1 cells co-transfected with constructs of either ERα or ERβ ligand binding domain, respectively, as well as a reporter. The results showed that ASEA exhibited a higher oestrogenic activity toward ERβ and a lower ERα oestrogenic activity (Figure 5A). The transacti-vation of ASEA on ERβ reached comparable levels to
1 nM E2 treatment at a concentration of 10μg/mL, whereas the activation of ERα was still below half of that of E2 treatment even at a concentration of 50 µg/mL ASEA. However, CUM exhibited a higher oestrogenic activity toward ERα than ERβ (Figure 5B) at the concentrations of 0.1–1 μM. The transactivation of CUM on both ERα and ERβ reached a level comparable to 1 nM E2 treatment at a concentration of 1μM.
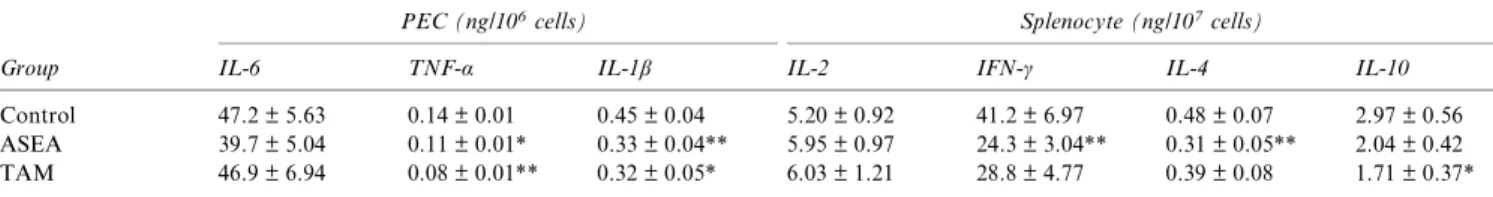
Table 1 The cytokine production from mitogen-stimulated peritoneal exudate cells and splenocytes of MRL-lpr/lpr micea
Group
PEC (ng/106cells) Splenocyte (ng/107cells)
IL-6 TNF-α IL-1β IL-2 IFN-γ IL-4 IL-10
Control 47.2 ± 5.63 0.14 ± 0.01 0.45 ± 0.04 5.20 ± 0.92 41.2 ± 6.97 0.48 ± 0.07 2.97 ± 0.56 ASEA 39.7 ± 5.04 0.11 ± 0.01* 0.33 ± 0.04** 5.95 ± 0.97 24.3 ± 3.04** 0.31 ± 0.05** 2.04 ± 0.42 TAM 46.9 ± 6.94 0.08 ± 0.01** 0.32 ± 0.05* 6.03 ± 1.21 28.8 ± 4.77 0.39 ± 0.08 1.71 ± 0.37*
aPeritoneal exudate cells (PEC) and splenocytes were collected from mice at age 18 weeks. Splenocytes and PEC were stimulated with 5 mg/L Con A
and 10 mg/L LPS, respectively. Values are means ± SE. Each group contains 14–18 mice. The significant difference between the control group and the experimental group was analysed by Student’s t-test.
*0.05 < P < 0.10. **P < 0.05.
Figure 3 The production of serum anti-dsDNA IgG2a (A) and anti-dsDNA IgG1 (B) of MRL-lpr/lpr mice supplemented without (control) or with alfalfa sprout ethyl acetate extract (ASEA) or tamoxifen (TAM). There were 14–18 mice, at the age 12 weeks, started on supplement diet. Bar value is mean ± SE and differs from the control group at the same age by Student’s t-test. † P = 0.26; ‡ P = 0.22.
Discussion
The attempt of dietary manipulation of disease sever-ity in lupus has been extended for decades. Suppres-sion of autoantibodies and inflammatory cytokine productions is suggested to be protective against dis-ease progression. In recent years, ERs are also sug-gested to play a role in oestrogen-related modulation of lupus.29In this study, ASEA, which may exert
anti-inflammatory and SERM activity by an in-vitro screening assay developed in our laboratory, was tested in murine lupus. ASEA significantly delayed the onset of proteinuria and prolonged the life span (Figure 1). However, the mice of the AS group had similar proteinuria progression and life span as the control group, suggesting that ingestion of whole AS had no beneficial effect.
Previous studies reported that alfalfa seed or sprout might be an SLE symptom eliciting food in both lab-oratory animal studies and a few human cases.22,23
Two inactive patients with SLE were also reported to reactivate serological and clinical symptoms after ingesting 8–15 alfalfa tablets daily.24 This causative
component was considered as a heat-liable amino acid L-canavanine25 and reported to increase antibody
production and higher renal histology scores in auto-immune mice.30 However, the AS group fed
550 mg/kg BW AS powder (0.55% in diet) in our study did not aggravate disease course. It may be
due to the counteraction of L-canavanine and ASEA, or the lower dose in our diet compared to the diet with 40% dried AS or with higher dose of L-canavanine treatment in the previous report.22,30
It was suggested that ERβ activation may have an immunosuppressive effect on murine lupus.29We also
showed that soy isoflavones with higher affinity for ERβ than ERα by transactivation activity alleviated disease severity in MRL-lpr/lpr in a previous study.16
In this study, ASEA also exhibited a higher oestro-genic activity toward ERβ than ERα. In contrast, CUM exhibited a higher oestrogenic activity toward ERα than ERβ at the concentrations of 0.1–1 μM (Figure 5). CUM is a well-known phytoestrogen in alfalfa.31 Our data suggested that phytoestrogenic
compounds other than CUM existed in ASEA as CUM did not show a selectivity toward ERβ as ASEA. The difference in ER subtype selectivity between ASEA and CUM seems to be in accordance with the difference in survival rate and life span in Experiment 1. Although in-vivo oestrogenic effect of ASEA needs to be further investigated, results of the present study confirmed our earlier observation that in-vitro screening of plant-derived fraction for both anti-inflammatory and SERM activity could be bene-ficial for murine lupus.
As previously reported by the other study,32CUM
neither ameliorated the disease severity nor prolonged life span in murine lupus model in this study.
Figure 4 Renal histological change of MRL-lpr/lpr mice supplemented without (A, control) or with alfalfa sprout ethyl acetate extract (B, ASEA) or tamoxifen (C) for 6 weeks. (A) shows enlarged glomeruli with hypercellularity and mesangial proliferation (arrows). (B) and (C) both show reductions in enlarged glomeruli, and had less mesangial proliferation (more space, arrows) while compared with the control. Sections were stained with haematoxylin and eosin (magnification, 200×).
Autoimmune and diet YH Honget al.
212
Lupus
at NATIONAL TAIWAN UNIV LIB on June 3, 2009 http://lup.sagepub.com
Although CUM supplementation lowered the occur-rence of proteinuria at early age, it increased spleno-cytic B-cell population (CUM, 72.5 ± 7.31% vs control, 45.6 ± 2.83%, P = 0.053), serum anti-dsDNA IgM level (CUM, 0.55 ± 0.06 OD vs control, 0.35 ± 0.02 OD, P < 0.01) (data not shown) and did not prolong life span while compared to the control. These immunological changes agree with the observa-tions that CUM did not ameliorate lupus onset and was not the main compound that contributed to the beneficial effect of the ASEA.
The isolation and chemical identification of the active compound(s) of ASEA with anti-inflammation and selective ER activation is currently under investi-gation. It has been reported that alfalfa contains flavo-noids such as conjugated apigenin,33isoflavones, such
as biochanin A, and formononectin,34 as well as
saponins.35 Although these compounds have been
reported to be oestrogenic,12 they are less selective
toward ERβ compared to ASEA observed in this
study. Saponins from soybean and Ginseng have also been reported to be oestrogenic.36,37 The
anti-inflammatory effects of these compounds have also been reported.38,39 Identification of the active
com-pound(s) in ASEA will provide more information for the future application in lupus attenuation.
TAM, a non-steroidal anti-oestrogen compound, has been reported to decrease the percentage of cell number of T cells, activated T cells (CD69+ T cells)
and DN T cells (CD4−CD8− T cells) of splenocytes and prolong the life span when administrated subcutaneously.27Although a decrease in DN T cells
was noted in the TAM group of this study but without statistical significance, the percentage of CD69+
T cells was significantly decreased (Figure 2). CD69 is an early T-cell activation marker. The percentages of activated T cells of peripheral blood mononuclear cells were significantly higher for patients with SLE than for healthy control. Activated T cells were reported to significantly correlate to SLE disease activity.40 Increased activated T cells were also
observed in splenocytes from lupus mice compared to normal mice.41In this study, ASEA
supplementa-tion, such as TAM, significantly decreased the num-ber of activated T cells, indicated that ASEA exerted immunosuppressive effect and thus might alleviate disease severity in murine lupus.
In addition, ASEA significantly suppressed the pro-duction of pro-inflammatory cytokines as shown in the ex-vivo IL-1β secretion from PEC and IFN-γ from splenocytes in the present study (Table 1). Serum IL-1β concentrations are increased in patients with SLE.42Mice deficient in IL-1β developed lower levels
of anti-dsDNA in experimental SLE suggested the role of IL-1β in the immunostimulatory and inflam-matory phenomena that mediate the development of SLE.43 Antagonist of IL-1β has been tested for the
alleviation of signs and symptoms of arthritis in SLE, but the results were not conclusive due to side effects.44 IFN-γ produced by lymph node cells from
MRL-lpr/lpr mice is higher than that from normal mice.45 The mRNA expression of IFN-γ and IL-10
in severe lupus-like mice increased and thus could be potential targets for immunotherapy.46 IL-10
secretion were slightly lower in TAM group (0.05 < P < 0.10) but not significantly different from the control group. Previous studies showed that die-tary manipulation, such as optimal dose of vitamin E, or soy isoflavone supplementation prolonged life span of murine lupus and significantly suppressed spleno-cytic IFN-γ secretion.4,16 Our results in the present
study with ASEA supplementation (Table 1) agree with our earlier observation that those lupus mice with prolonged life span (Figure 1) by diet treatment
Figure 5 Effects of alfalfa sprout ethyl acetate extract (A) and coumestrol (B) on the transcriptional activation of oestrogen receptors (Gal4-hERα or β). Values are expressed as means ± SE of four separate experiments, and 1 nM E2 (17β-estradiol) is positive control. Values not sharing the same letter are significantly different by Duncan’s multiple range test (P < 0.05, a–d represents the significant difference for ERα activities, whereas x–z represent for ERβ).
had significantly lower IFN-γ secretion from mitogen-stimulated splenocytes. The suppressive effect of ASEA on IL-1β and IFN-γ secretion implied a decreased risk of interplay of inflammatory and immune mediators.
Ex-vivo IL-4 secretion from Con A-stimulated sple-nocytes of MRL-lpr/lpr mice supplemented with ASEA also significantly decreased compared to the control. Higher serum levels of IL-4 were found in patients with SLE in several reports.47–49 IL-4
enhanced the production of anti-dsDNA IgG from splenocytes of lupus mice. Administration of mono-clonal antibodies against IL-4 before the onset of lupus inhibited the production of anti-dsDNA IgG and prevented the onset of lupus nephritis.50 Lupus
mice spontaneously secrete large quantities of patho-genic IgG1 and IgG2a autoantibodies, this isotype switching is known to be enhanced by IL-4 and IFN-γ, respectively.51In the present study, splenocytic
IL-4 and IFN-γ secretion in response to Con A were significantly reduced in the ASEA supplemented mice, although the lowering in serum levels of anti-dsDNA IgG1 and IgG2a did not reach statistical dif-ference. The results suggested that the beneficial effect of ASEA might be more attributable to the down-regulation of inflammatory mediators and cytokines instead of autoantibodies. This speculation is also sup-ported by the results of renal histology that also showed less enlarged glomeruli and mesangial prolif-eration in the ASEA and the TAM groups.
In conclusion, this study showed that autoimmune-prone MRL-lpr/lpr mice fed AS EA extract from age 12 weeks had significantly lower activated T-cell pop-ulation, IL-1β secretion by LPS-stimulated PEC, IFN-γ and IL-4 production by Con A-stimulated sple-nocytes and prolonged life span. The present study confirmed our earlier observation that plant-derived components with anti-inflammatory and SERM activity may have beneficial effects on disease course of murine lupus.
Acknowledgements
The authors have to disclose that there is no conflict of interest in this study. We are grateful for the help-ful advice and the analysis of oestrogenic activities by Ching-jang Huang and Ssu-Ching Wang. This work was kindly supported by a grant from the National Science Council of the Republic of China (NSC 94-2321-B-002-010).
References
1 Edwards CJ, James JA. Making lupus: a complex blend of genes and environmental factors is required to cross the disease threshold. Lupus 2005; 15: 713–714.
2 Leiba A, Amital H, Gershwin ME, Shoenfeld Y. Diet and lupus. Lupus 2001; 10: 246–248.
3 Lin BF, Jeng SJ, Chiang BL, Huang CC. Dietary fat affects lipids and anti-cardiolipin antibody levels in autoimmune-prone NZB/W F1 mice. Br J Nutr 1997; 77: 657–669.
4 Hsieh CC, Lin BF. The effects of vitamin E supplementation on autoimmune-prone New Zealand black× New Zealand white F1 mice fed an oxidised oil diet. Br J Nutr 2005; 93: 655–662.
5 Lai NS, Lin RH, Lai RS, Kun UC, Leu SC. Prevention of autoanti-body formation and prolonged survival in New Zealand Black/New Zealand White F1 mice with an ancient Chinese herb, Ganoderma tsugae. Lupus 2001; 10: 461–465.
6 Auborn KJ, Qi M, Yan XJ, et al. Life span is prolonged in autoimmune-prone (NZB/NZW) F1 mice fed a diet supplemented with indole-3-carbinol. J Nutr 2003; 133: 3610–3613.
7 Sawai C, Anderson K, Walser-Kuntz D. Effect of bisphenol A on murine immune function: modulation of interferon-gamma, IgG2a, and disease symptoms in NZB× NZW F1 mice. Environ Health Per-spect 2003; 111: 1883–1887.
8 Lahita RG. The importance of estrogen in systemic lupus erythema-tosus. Clin Immunol Immunopathol 1992; 63: 17–18.
9 Cooper GS, Dooley MA, Treadwell EL, St. Clair EW, Parks CG. Hormonal, environmental, and infectious, risk factors for developing systemic lupus erythematosus. Arthritis Rheum 1998; 41: 1714–1724. 10 Carlsten H, Tarkowski A, Holmdahl R, Nilsson LA. Oestrogen is a
potent disease accelerator in SLE-prone MRL-lpr/lpr mice. Clin Exp Immunol 1990; 80: 467–473.
11 Dhaher YY, Greenstein B, de Fougerolles Nunn F, Khamashta M, Hughes GR. Strain differences in binding properties of estrogen recep-tors in immature and adult BALB/c and MRL/MP-lpr/lpr mice, a model of systemic lupus erythematosus. Int J Immunopharmacol 2000; 22: 247–254.
12 Kuiper GG, Lemmen JG, Carlsson B, et al. Interaction of estrogenic chemicals and phytoestrogens with estrogen receptor beta. Endocrinol-ogy 1998; 139: 4252–4263.
13 Morito K, Aomori T, Hirose T, et al. Interaction of phytoestrogens with estrogen receptors alpha and beta (II). Biol Pharm Bull 2002; 25: 48–52.
14 Birt DF, Hendrich S, Wang W. Dietary agents in cancer prevention: 9 flavonoids and isoflavonoids. Pharmacol Ther 2001; 90: 157–177. 15 Verdrengh M, Jonsson IM, Holmdahl R, Tarkowski A. Genistein as
an anti-inflammatory agent. Inflamm Res 2003; 52: 341–346. 16 Hong YH, Wang TC, Huang CJ, Cheng WY, Lin BF. Soy
isofla-vones supplementation alleviates disease severity in autoimmune-prone MRL-lpr/lpr mice. Lupus (in press).
17 Middleton MR, Lorigan P, Owen J, et al. A randomized phase III study comparing dacarbazine, BCNU, cisplatin and tamoxifen with dacarbazine and interferon in advanced melanoma. Br J Cancer 2000; 82: 1158–1162.
18 Cheng WY, Kuo YH, Huang CJ. Isolation and identification of novel estrogenic compounds in yam tuber (Dioscorea alata Cv. Tainung No. 2). J Agric Food Chem 2007; 55: 7350–7358.
19 Malinow MR, McLaughlin P, Stafford C, Livingston AL, Kohler GO. Alfalfa saponins and alfalfa seeds. Dietary effects in cholesterol-fed rabbits. Atherosclerosis 1980; 37: 433–438.
20 Swain SM, Armentano LE. Quantitative evaluation of fiber from nonforage sources used to replace alfalfa silage. J Dairy Sci 1994; 77: 2318–2331.
21 Foster S, Duke JA. A field guide to medicinal plants and herbs: eastern and central North America. In: Peterson RT, (ed), Houghton Mifflin Co, Boston, Massachusetts; 1990. p. 211.
22 Malinow MR, Bardana EJ Jr, Pirofsky B, Craig S, McLaughlin P. Systemic lupus erythematosus-like syndrome in monkeys fed alfalfa sprouts: role of a nonprotein amino acid. Science 1982; 216: 415–417. 23 Montanaro A, Bardana EJ Jr. Dietary amino acid-induced systemic lupus erythematosus. Rheum Dis Clin North Am 1991; 17: 323–332.
Autoimmune and diet YH Honget al.
214
Lupus
at NATIONAL TAIWAN UNIV LIB on June 3, 2009 http://lup.sagepub.com
24 Roberts JL, Hayashi JA. Exacerbation of SLE associated with alfalfa ingestion. N Engl J Med 1983; 308: 1361.
25 Alcocer-Varela J, Iglesias A, Llorente L, Alarcon-Segovia D. Effects of L-canavanine on T cells may explain the induction of systemic lupus erythematosus by alfalfa. Arthritis Rheum 1985; 28: 52–57. 26 Akaogi J, Barker T, Kuroda Y, et al. Role of non-protein amino acid
L-canavanine in autoimmunity. Autoimmun Rev 2006; 5: 429–435. 27 Wu WM, Suen JL, Lin BF, Chiang BL. Tamoxifen alleviates disease
severity and decreases double negative T cells in autoimmune MRL-lpr/lpr mice. Immunology 2000; 100: 110–118.
28 National Research Council. Guide for the care and use of laboratory animals. Publication no. 85–23 (rev.), National Institutes of Health, Bethesda, MD, (1985).
29 Li J, McMurray RW. Effects of estrogen receptor subtype-selective agonists on autoimmune disease in lupus-prone NZB/NZW F1 mouse model. Clin Immunol 2007; 123: 219–226.
30 Prete PE. Effects of L-canavanine on immune functioning in normal and autoimmune mice: disordered B-cell function by a dietary amino acid in the immunoregulation of autoimmune disease. Can J Physiol Pharmacol 1985; 63: 843–854.
31 Bovee TF, Schoonen WG, Hamers AR, Bento MJ, Peijnenburg AA. Screening of synthetic and plant-derived compounds for (anti)estro-genic and (anti)andro(anti)estro-genic activities. Anal Bioanal Chem 2008; 390: 1111–1119.
32 Schoenroth LJ, Hart DA, Pollard KM, Fritzler MJ. The effect of the phytoestrogen coumestrol on the NZB/W F1 murine model of sys-temic lupus. J Autoimmun 2004; 23: 323–332.
33 Stochmal A, Piacente S, Pizza C, De Riccardis F, Leitz R, Oleszek W. Alfalfa (Medicago sativa L.) flavonoids. 1. Apigenin and luteolin gly-cosides from aerial parts. J Agric Food Chem 2001; 49: 753–758. 34 Saloniemi H, Wähälä K, Nykänen-Kurki P, Kallela K, Saastamoinen
I. Phytoestrogen content and estrogenic effect of legume fodder. Proc Soc Exp Biol Med 1995; 208: 13–17.
35 Story JA, LePage SL, Petro MS, et al. Interactions of alfalfa plant and sprout saponins with cholesterol in vitro and in cholesterol-fed rats. Am J Clin Nutr 1984; 39: 917–929.
36 Rowlands JC, Berhow MA, Badger TM. Estrogenic and antiproli-ferative properties of soy sapogenols in human breast cancer cells in vitro. Food and Chem Toxicol 2002; 40: 1767–1774.
37 Cho J, Park W, Lee S, Ahn W, Lee Y. Ginsenoside-Rb1 from Panax ginseng C.A. Meyer activates estrogen receptor-alpha and -beta, inde-pendent of ligand binding. J Clin Endocrinol Metab 2004; 89: 3510– 3515.
38 Nicholas C, Batra S, Vargo MA, et al. Apigenin blocks lipopolysaccharide-induced lethality in vivo and proinflammatory
cytokines expression by inactivating NF-kappaB through the suppres-sion of p65 phosphorylation. J Immunol 2007; 179: 7121–7127. 39 Kang JH, Sung MK, Kawada T, et al. Soybean saponins suppress the
release of pro-inflammatory mediators by LPS-stimulated peritoneal macrophages. Cancer Lett 2005; 230: 219–227.
40 Su CC, Shau WY, Wang CR, Chuang CY, Chen CY. CD69 to CD3 ratio of peripheral blood mononuclear cells as a marker to monitor systemic lupus erythematosus disease activity. Lupus 1997; 6: 449– 454.
41 Chae BS, Shin TY. Immunoregulatory abnormalities of T cells and hyperreactivity of B cells in the in vitro immune response in pristane-induced lupus mice. Arch Pharm Res 2007; 30: 191–198.
42 Nagahama M, Nomura S, Ozaki Y, Yoshimura C, Kagawa H, Fukuhara S. Platelet activation markers and soluble adhesion mole-cules in patients with systemic lupus erythematosus. Autoimmunity 2001; 33: 85–94.
43 Voronov E, Dayan M, Zinger H, et al. IL-1 beta-deficient mice are resistant to induction of experimental SLE. Eur Cytokine Netw 2006; 17: 109–116.
44 Moosig F, Zeuner R, Renk C, Schröder JO. IL-1RA in refractory sys-temic lupus erythematosus. Lupus 2004; 13: 605–606.
45 Manolios N, Schrieber L, Nelson M, Geczy CL. Enhanced interferon-gamma (IFN-γ) production by lymph node cells from autoimmune (MRL/1, MRL/n) mice. Clin Exp Immunol 1989; 76: 301–306. 46 Prud’homme GJ, Jono DH, Theofilopoulos AN. Quantitative
poly-merase chain reaction analysis reveals marked overexpression of IL-1β, IL-10 and IFN-γ mRNA in the lymph nodes of lupus-prone mice. Mol Immunol 1995; 32: 495–503.
47 Cuadrado MJ, Marubayashi M, Ortega C, et al. Relationship of IL-2, IL-2R (CD25+), soluble IL-2R and IL-4 with disease activity in SLE
patients. Lupus 1993; 2: 257–260.
48 Peng SL, Mosheli J, Craft J. Roles of IFN-γ and IL-4 in murine lupus. J Clin Invest 1997; 99: 1936–1946.
49 Wong CK, Ho CY, Li EK, Lam CW. Elevation of proinflammatory cytokine (IL-18, IL-17, IL-12) and Th2 cytokine (IL-4) concentrations in patients with systemic lupus erythematosus. Lupus 2000; 9: 589– 593.
50 Nakajima A, Hirose S, Yagita H, Okumura K. Roles of 4 and IL-12 in the development of lupus in NZB/W F1mice. J Immunol 1997;
158: 1466–1472.
51 Klinman DM. IgG1 and IgG2a production by autoimmune B cells treated in vitro with IL-4 and IFN-gamma. J Immunol 1990; 144: 2529–2534.