*Department of Zoology, ‡Department of Entomology, National Taiwan University, and †Institute of Biological Chemistry, Academia Sinica,
Taipei, Taiwan, Republic of China
Received August 13, 2001; returned to author for revision September 5, 2001; revised November 5, 2001
To identify the protein encoded by a 687-bp open reading frame (ORF) of a salI genomic DNA fragment of shrimp white spot syndrome virus (WSSV), we expressed the ORF in a baculovirus/insect cell expression system. The apparent molecular mass of the recombinant protein on sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) was 35 kDa in insect cells. Antibody raised against bacterially synthesized protein of the ORF identified a nucleocapsid protein (VP35) in the extracts of both the purified WSSV virions and the nucleocapsids which comigrated with the 35-kDa baculovirus-expressed recombinant protein on SDS-PAGE. We also show by transient expression in insect cells (Sf9) that VP35 targets the nucleus. Two potential nuclear localization signals (NLSs) were characterized, but only one of them was important for targeting VP35 to the nuclei of transfected insect cells. Replacement of a cluster of four positively charged residues (24KRKR27) at the N
terminus of the protein with AAAA resulted in mutant proteins that were distributed only in the cytoplasm, thus confirming that this sequence is a critical part of the functionally active NLS of VP35. © 2002 Elsevier Science (USA)
Key Words: Penaeus monodon; white spot syndrome virus (WSSV); nucleocapsid; nuclear localization signal (NLS); VP35.
INTRODUCTION
White spot syndrome virus (WSSV) is the causative agent of a disease that has led to severe mortalities of cultured shrimps in Taiwan and many other countries (Chou et al., 1995; Flegel, 1997; Lo et al., 1999). WSSV is an enveloped, ellipsoid, large, double-stranded DNA vi-rus (Wang et al., 1995; Wongteerasupaya et al., 1995) and it has a wide host range among crustaceans (Lo et al., 1996, 1998). Even while the molecular data was still limited, the morphological characteristics and general biological properties of the virus (Wang et al., 1995; Wongteerasupaya et al., 1995; Lo et al., 1996) already highlighted its uniqueness, and recent data, including the genome sequence and phylogenies based on DNA polymerase and protein kinase, suggest that WSSV is a member of a new virus family (van Hulten et al., 2001; Liu
et al., 2001). The size of the WSSV genome has been
differently reported for different isolates: 305107 bp (AX151396), 292967 bp (van Hulten et al., 2001) and 307287 bp (AF440570) for viruses isolated fromChina, Thailand, and Taiwan, respectively, with these size dif-ferences mostly being due to several small insertions and one large (⬃12 kb) deletion. The uniqueness of this
virus means that a thorough study of its molecular biol-ogy is still urgently needed for a better understanding of its nature and replication strategy and the molecular mechanisms of its pathogenesis.
All animal DNA viruses, except poxviruses (Wittek, 1982) and irridovirus (Tidona and Darai, 1997), replicate in the cell nucleus (Kasamatsu and Nakanishi, 1998). As a first step in infection, viruses import their genomic DNA into the nuclei of infected cells, and it is here that tran-scripts leading to viral protein synthesis are made. Most viruses utilize the nuclear import system of the cell, including the microtubules (Sodeik et al., 1997; Suoma-lainen et al., 1999), the nuclear pore complex (Greber et
al., 1996, 1997), receptors, and import factors (Marsh and
Helenius, 1989; Whittaker and Helenius, 1998; reviewed in Kasamatsu and Nakanishi, 1998) to access the nu-cleus. Often, viruses that are too large to easily enter the nucleus will first locate to the nuclear pore and then release their DNA for transport to the nucleus in asso-ciation with one or more mediating viral proteins. Thus for example, canine parvovirus (Vihinen-Ranta et al., 2000), hepatitis B virus (Kann et al., 1999), adenovirus (Greber et al., 1997), and simian virus 40 (Wychowski et
al., 1986, 1987; Nakanishi et al., 1996) all use one or
several viral proteins to mediate the import of their viral DNA into the nucleus. In a normal eukaryotic cell, for a cellular protein to pass through the central transporter channel into the nucleus, it must contain some kind of
1These two authors contributed equally.
2To whomcorrespondence and reprint requests should be
ad-dressed.
0042-6822/02 $35.00
© 2002 Elsevier Science (USA) All rights reserved.
nuclear localization signal (NLS); likewise if a viral pro-tein targets the nucleus, whether it acts to mediate the import of viral DNA or not, it too must include an NLS (Kalderon et al., 1984; reviewed in Jans and Hubner, 1996, and in Kasamatsu and Nakanishi, 1998). The classical versions of this signal consist of predominantly basic amino acids appearing in either one (simple type or monopartite NLS) or two (bipartite NLS) short clusters.
WSSV structural proteins are unique in that they do not show any obviously significant bands in SDS-PAGE, and fromthe WSSV genome sequence, no genes have yet been identified that show high homology to the structural protein genes of baculoviruses or other DNA viruses. It is therefore hard to directly apply other virus infection mod-els to interpret the infection strategy of WSSV; on the contrary, WSSV’s infection strategy must be investigated
ab initio. Thus, because WSSV structural protein(s) that
have an NLS might play a role in mediating the import of WSSV DNA into the nuclei of infected cells, we began by focusing on structural proteins that exhibit nuclear tar-geting behavior. WSSV VP35 is one such structural nu-cleocapsid protein. In the present paper, we identify and characterize its ORF in a WSSV salI genomic DNA frag-ment and express the protein in a baculovirus/insect system. We also use transient expression in insect cells (Sf9) to show that VP35 targets the nucleus, and define
and characterize a cluster of four positively charged residues at the N-terminus of VP35 that directs the pro-tein to the nucleus. This report is the first description of a functionally active NLS within a nucleocapsid protein of shrimp white spot syndrome virus.
RESULTS AND DISCUSSION
Location and sequence of the ORF encoding VP35 The virus used in this study was isolated froma batch of WSSV-infected Penaeus monodon collected in Taiwan in 1994 (Wang et al., 1995), which is now known as WSSV Taiwan isolate (Lo et al., 1999). Fromthis virus, a plasmid library (referred to as the pms library, where pm indicates
Penaeus monodon and s indicates SalI fragments) of
WSSV SalI genomic fragments was constructed (Wang et
al., 1995). The pms147 ORF5 (located at nt 1818 to 2504
of the 12002 bp pms147) was used in the present study (Fig. 1). This ORF encodes a protein of 228 amino acids with a theoretical size of 26 kDa and an isoelectric point of 4.51.
The nucleotide sequence surrounding the methionine start codon (AAAATGG; see Fig. 2) of the predicted pro-tein conformed to the Kozak rule of an efficient context for eukaryotic translation initiation (Kozak, 1989). A poly-adenylation signal (AATAAA) was located 7 nt down-FIG. 1. Restriction enzyme map of the pms147 SalI (S) fragment of WSSV genomic DNA showing the location (nt 1818 to nt 2504) of the WSSV vp35 gene (pms147 ORF5). Because a pms147 orf4-specific riboprobe is used later for Northern blotting analysis, the orf4 gene is also shown for reference. The pms147 segment contains six PstI (P) sites, six EcoRI (E) sites, and one BamHI (B) site. The relative positions on the entire WSSV genome for this fragment and for other published genes [rr genes (Tsai et al., 2000a); tk-tmk (Tsai et al., 2000b); vp22, vp25, and pk1 (Liu et al., 2001)] are also shown for reference. A protein kinase gene (pk2) that is distinctly related to the published protein protein kinase (pk1) is also indicated.
45 NUCLEAR TARGETING OF A SHRIMP WSSV NUCLEOCAPSID PROTEIN
streamof the translational stop codon. Based on the PROSITE database, the potential sites for posttransla-tional modifications of this protein include two sites (NTTK120 and NFSF152) for N-linked glycosylation, four
sites (SSR5, TKR25, SNK53, and TTK120) for PKC
phosphor-ylation, two sites (SHYE139 and SLID224) for CKII
phos-phorylation, and two sites (KRKR27 and KRPR56) for
po-tential endoprotease cleavage. Computer analysis of the protein showed that at the N terminus, there is quite a large continuous hydrophilic region that includes the acidic amino acid-rich domain shown in Fig. 2. The pms147 ORF5 protein was subsequently identified as a nucleocapsid protein by the following experiments. Expression of the pms147 ORF5 (VP35) DNA in eukaryotic cell-free system
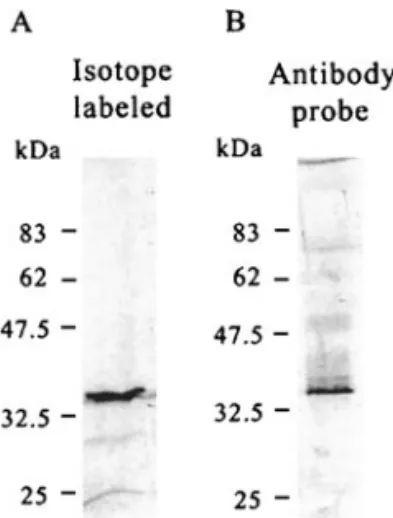
An expression vector, pcDNA3-HA-VP35, with the full length (nt 1818 to nt 2504) of pms147 ORF5 DNA was constructed for coupled in vitro transcription and trans-lation reactions in a cell-free systemwith rabbit reticu-locyte lysate. The translated protein was expected to be a hemagglutinin epitope (MCYPYDVPDYASLA)-tagged polypeptide (HA-tagged VP35). Following the TNT Quick Coupled Transcription/Translation System(Promega, Madison, WI) protocols, a single protein band visualized by autoradiography was synthesized. This had an appar-ent molecular mass higher than the theoretical size of the ORF5 protein and was about 35 kDa on SDS-PAGE (Fig. 3A). This polypeptide was specifically detectable with a commercial antibody against the tag (data not shown) and also was recognized by antibody raised against bacterially synthesized ORF protein (Fig. 3B). Thus, we conclude that the pms147 ORF5 protein ex-pressed in the eukaryotic cell-free translation system had an apparent molecular mass of 35 kDa on
SDS-PAGE. However, since the apparent size of this pms147 ORF5 in vitro translation product was significantly higher than the calculated molecular mass of 26,000, further verifications were made.
Expression of the pms147 ORF5 DNA in a baculovirus/insect cell systems
Using the BacVector Transfection system, an
Autogra-pha californica multicapsid nucleopolyhedrovirus
(AcMNPV)-based recombinant virus was constructed for the expression of the pms147 ORF5 in Sf9 cells. To this end, a recombinant AcMNPV (VL1392-VP35) with the full length of the pms147 ORF5 was constructed and propa-gated in Sf9 cells at 27°C in Sf-900 II SFM cell culture medium (Gibco BRL). The recombinant protein (rVP35; FIG. 2. The nucleotide sequence and deduced amino acid sequence of the WSSV vp35 gene. The two potential NLSs (boxes) and the acidic amino acid-rich domain (shaded) are indicated. The polyadenylation signal is marked with asterisks.
FIG. 3. VP35 yielded in eukaryotic in vitro translation systemvisual-ized by (A) autoradiography and (B) Western blot analysis. The size standards were determined using BenchMark prestained protein lad-der (Gibco BRL) and were corrected with BenchMark protein ladlad-der (Gibco BRL).
fusion protein of VP35 with additional 13 amino acids at N terminus) synthesized in insect cells infected with recombinant baculovirus VL1392-VP35 had an apparent molecular mass of 35 kDa on SDS-PAGE (Fig. 4A) (i.e., it was equal in size to the protein detected in the eukary-otic cell-free in vitro translation system) and it was rec-ognized by antibody raised against the bacterially syn-thesized pms147 ORF5 protein (Fig. 4B).
It is well documented that very acidic proteins such as the constitutive nucleolar protein (NO29; pI 3.75) of
Xe-nopus laevis protein (Zirwes et al., 1997) and brain
acid-soluble proteins (BASPs; pI 4.4–4.6) (Mosevisky et al., 1994) show an abnormally low mobility during SDS-PAGE electrophoresis. That is, they all show a SDS-PAGE mo-bility that corresponds to a molecular mass higher than the calculated molecular mass. This phenomenon pre-sumably accounts for the difference between the calcu-lated molecular mass and apparent molecular mass on SDS-PAGE of VP35 (pI 4.51).
Identification of the pms147 ORF5 protein as a nucleocapsid protein
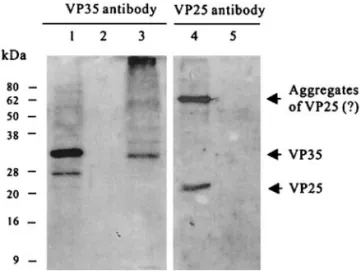
Antibody raised against bacterially synthesized tein of the pms147 ORF5 identified a nucleocapsid pro-tein (VP35) in the extracts of the purified WSSV virions which comigrated with the 35-kDa baculovirus-ex-pressed recombinant protein on SDS-PAGE (Fig. 5, lane 1). VP35 was further shown to associate with the nucleo-capsid fraction, but not with envelope fraction, in a triton-treated preparation of the purified virions (Fig. 5, lanes 2 and 3). Conversely, antibody against VP25 (a WSSV en-velope protein) reacted with protein in the enen-velope fraction, but did not react with the nucleocapsid fraction (Fig. 5, lanes 4 and 5). These results confirmthat VP35 is a WSSV nucleocapsid protein.
Transcriptional analysis of vp35
To discover when the vp35 mRNA is transcribed in the viral life cycle, and which mRNA is transcribed, we per-formed a transcriptional analysis of vp35 using RT-PCR and Northern blot analysis. Due to the lack of a shrimp cell line, it is impossible to do a temporal analysis of WSSV gene transcription in cells synchronously infected with the virus. However, WSSV is extremely virulent, and the onset of the disease is remarkably rapid in infected shrimp (Chou et al., 1995). Further, the disease progresses at a more or less similar pace in different individuals, especially in cuticular epithelial and connec-FIG. 4. Detection of the recombinant VP35 (rVP35) in insect cells by
Coomassie blue staining (A) and Western blotting (B). Lane 1: Cell lysate proteins fromthe Sf9 cells infected with recombinant AcMNPV (VL1392-VP35). Lane 2: Cell lysate proteins fromthe Sf9 cells infected with wild-type AcMNPV. Lane 3: Cell lysate proteins fromthe uninfected Sf9 cells. M1: BenchMark prestained protein ladder. M2: BenchMark protein ladder. The size standards indicated in (B) were based on prestained protein ladder corrected with BenchMark protein ladder.
FIG. 5. Analysis of VP35 localization by SDS-PAGE and Western blotting. Lane 1: Cell lysate proteins fromthe Sf9 cells infected with recombinant AcMNPV (VL1392-VP35). Lanes 2, 4: Envelop fraction of the purified WSSV virion. Lanes 3, 5: Nucleocapsid fraction of the purified WSSV virion. Proteins were separated on 12.5% SDS-PAGE and then probed with VP35 antiserum(lanes 1, 2, and 3) or VP25 (envelope protein) antiserum(lanes 4 and 5). The size standards were determined using BenchMark prestained protein ladder and were corrected with BenchMark protein ladder.
FIG. 6. Temporal transcription analysis of WSSV by RT-PCR. (A) RT-PCR with vp35-specific primers. (B) Internal positive control: total RNA treated with DNase and amplified with actin-specific primers. The products were resolved in 1% agarose gel containing 0.5 g/ml ethidiumbromide. The size standards are indicated using pGEM DNA marker (Promega). Lane headings show hours pi.
47 NUCLEAR TARGETING OF A SHRIMP WSSV NUCLEOCAPSID PROTEIN
tive tissues. We therefore performed the transcriptional analysis in experimentally infected shrimp using tissues fromthe pleopod as the source of total RNA, because this organ most frequently showed the highest preva-lence of the virus in early infection (Lo et al., 1997; Kou et
al., 1998).
RT-PCR analysis revealed that the vp35 transcript was first detected at 2 h p.i. and continued to be expressed at a very low level through to 12 h p.i. (Fig. 6A). The vp35 RT-PCR product then significantly increased at 18 and 24 h p.i. and declined again at 36 h p.i. (Fig. 6A). Northern blot analysis with vp35 specific riboprobe first detected two major transcripts of approximately 5.5 and 1.3 kb at 18 h p.i. (Fig. 7A). The temporal expression pattern of the 1.3 kb transcript (Fig. 7) is very similar to the pattern observed in the RT-PCR analysis (Fig. 6), and the length of the 1.3-kb transcript matched well with the length of the vp35 coding region (687 bp) (assuming 5⬘ and 3⬘ UTRs totally of several hundred nucleotides). The 5.5 kb band (Fig. 7) was very intense at 18 h p.i. and then dropped significantly. A riboprobe for orf4 located 3 kb upstreamof vp35 in the pms 147 fragment (see Fig. 1) also hybridized with the 5.5 kb transcript (Fig. 7B), indi-cating that this transcript must have been transcribed from a region in the pms 147 fragment that starts up-streamof the orf4 coding region and extends down-streamto vp35. The difference in intensities of the 1.3-kb transcript and the 5.5-kb transcript at 18 h p.i. also sug-gests that WSSV uses different promoters (with different strengths) to initiate transcription of the two vp35-con-taining (5.5 and 1.3 kb) transcripts. It is also worth noting that because the primer set used in RT-PCR was located in the vp35 coding region, Fig. 6A shows a combined
result amplified from both the 1.3 kb transcript and the 5.5 kb transcript.
Identification of VP35 protein nuclear localization signal (NLS)
Inspection of the 228 amino acids of VP35 revealed two clusters of four amino acid residues (24KRKR27, 53KRPR56) at the N terminus of the protein. These motifs
resemble many of the characterized classical NLSs, which are made essentially of a short stretch of posi-tively charged amino acids (Fig. 8). We therefore inves-tigated whether these two sequences function as the nuclear localization signal(s) of VP35.
FIG. 7. Northern blot temporal transcription analysis of total RNA isolated from WSSV-infected P. monodon using vp35- and pms 147 orf4-specific riboprobes. (A) Two transcripts of vp35 are approximately 5.5 and 1.3 kb. (B) Two transcripts of pms 147 orf4 are approximately 5.5 and 1.8 kb. The size standards were determined by RNA marker (Gibco BRL). Lane headings show hours pi.
FIG. 8. Alignment of the deduced NLS amino acid sequences of WSSV VP35 with the NLS motifs of other DNA virus proteins.
To engineer two mutant VP35s, in each of which one of the two potential NLS(s) (24KRKR27 and 53KRPR56) was
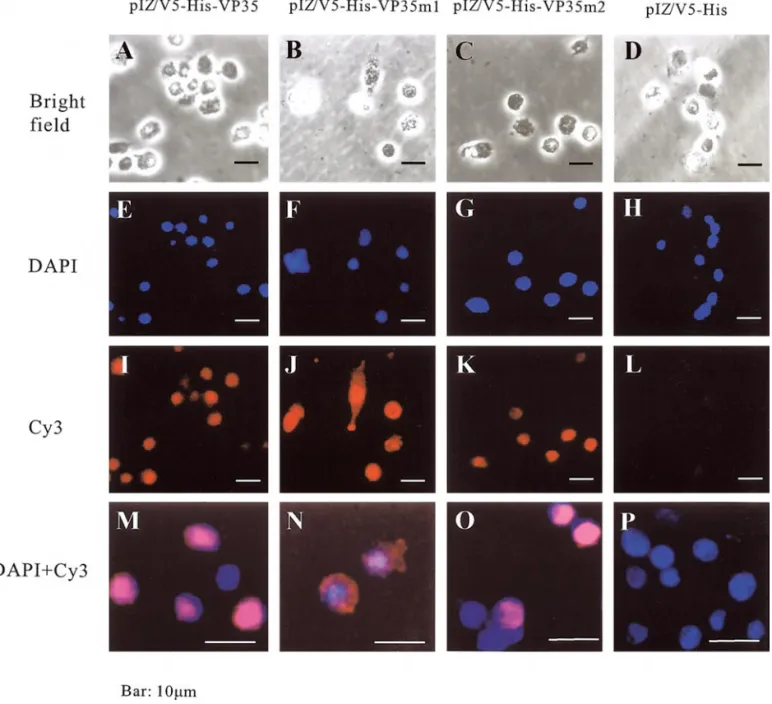
replaced with four alanines (AAAA), the genes of VP35 and the two mutant VP35 proteins were cloned into pIZ/V5-His vector to yield VP35, pIZ/V5-His-VP35m1, and pIZ/V5-His-VP35m2, respectively. The In-sectSelect Systemwas then used to transiently express VP35 and its mutants in Sf9 cells under the control of the baculovirus Orgyia pseudotsugata multicapsid nuclear polyhedrosis virus (OpMNPV) immediate-early gene pro-moter (OpIE2). The subcellular distribution of the result-ant VP35 proteins was visualized 48 h after transfection by indirect immunofluorescence microscopy with a
rab-bit antibody against bacterially synthesized VP35 in com-bination with Cy3-conjugated goat rabbit IgG anti-body as a secondary antianti-body. Nuclei were stained with 4⬘-6⬘-diamidino-2 phenylindole dihydrochloride (DAPI) (Fig. 9). The transfection efficiency was about 80%. The Cy3 signals for the wild-type VP35 (Figs. 9I and 9M) and the m2 53AAAA56 mutant VP35 (Figs. 9K and 9O) were
perfectly coincident with the nuclear DAPI signal, whereas expression of the m1 24AAAA27-mutant VP35
(Figs. 9J and 9N) was spread throughout the cytoplasmof the transfected cells. This result implies that the se-quence replaced in m1 mutant (i.e.,24KRKR27) is [a critical
part of] the signal that causes nuclear accumulation of FIG. 9. Intracellular localization of normal VP35 and two NLS-mutant VP35 proteins in Sf9 cells. Sf9 cells were transfected with expression vectors pIZ/V5-His-VP35, pIZ/V5-His-VP35m1, pIZ/V5-His-VP35m2, and pIZ/V5-His, respectively, where the m1 mutant has AAAA instead of24KRKR27, the m2
mutant has AAAA instead of53KRPR56, and pIZ/V5-His is a negative control. Twenty-four hours after transfection, cells were stained with indirectly
Cy3-conjugated antibody and counterstained with DAPI. The top row shows the transfected Sf9 cells under a bright field microscope, the second row shows the DAPI-counterstained nuclei of the same cells, the third row shows the Cy3 fluorescent signals from the transfected cells, and the last row shows the merged images of DAPI and Cy3 fluorescent signals from the transfected cells at higher magnification.
49 NUCLEAR TARGETING OF A SHRIMP WSSV NUCLEOCAPSID PROTEIN
(Fig. 9) show that mutation of the KRKR sequence to alanines completely abolished the nuclear accumulation of the protein, we cannot be certain whether the24KRKR27
sequence constitutes the entire NLS signal itself or if it is a crucial part of a bigger signal.
In summary, Western analysis of the VP35 structural protein studied here revealed that it was associated with the nucleocapsid, and transient expression in insect cells (Sf9) showed that it targeted the nucleus. We de-fined and characterized a cluster of four positively charged residues at the N terminus of VP35 that directs the protein to the nucleus. When the four basic amino acids of this motif were replaced with AAAA, the mutant protein remained totally cytoplasmic, thus indicating its function as the nuclear localization signal of VP35. It still remains to be shown that VP35 actually serves to medi-ate the import of viral DNA into the nucleus, but mean-while this report is the first description of an NLS found within a nucleocapsid protein of shrimp white spot syn-drome virus.
MATERIALS AND METHODS DNA sequencing and computer analysis
In 1994, WSSV was collected and purified fromin-fected Penaeus monodon (Wang et al., 1995) and then used to construct WSSV genomic libraries (Lo et al., 1996). This virus source (known as the Taiwan isolate; Lo
et al., 1999) was used as the basis for all of the sequence
work in the present study. Plasmid DNA for sequencing was purified using the QIAprep Miniprep System(Qia-gen, Germany). The DNA fragments were sequenced by primer walking on both strands. The nucleotide and the predicted protein sequences were analyzed using Gene-Works 2.5.1 (Oxford Molecular Group, Inc., Campbell, CA). The DNA and the deduced amino acids sequences were compared with GenBank/EMBL, SWISSPORT, and PIR databases using the programs FASTA (Pearson and Lipman, 1988) and BLAST (Altschul et al., 1997). The Prosite database was searched using Proscan (Bairoch
et al., 1997) and the hydrophobicity of VP35 was
pre-dicted according to the method published by Hopp and Woods (1981).
Virus purification
The virus used in this study was isolated fromnaturally infected P. monodon shrimp from Taiwan. It was first purified on a sucrose gradient as described previously (Wang et al., 1995) and then further purified on a CsCl gradient (initially 40% in TE buffer, 10 mM Tris-HCl, 1 mM EDTA, pH 7.6) at 74,000 g at 10°C for 16 h. The visible viral band was then removed and pelleted by centrifugation at 74,000 g at 4°C for 30 min. The virus envelope was removed from the virus particles by adding Triton X-100 (1% in TE buffer) to the virus solution and incubating for 1 h at 37°C (Nadala et al., 1998), and the nucleocapsids were then pelleted by centrifugation at 140,000 g at 4°C for 10 min. The pellet was dissolved in TE buffer for the SDS-PAGE and Western blot analysis.
Recombinant pms147 ORF5 protein (VP35) overexpression in Esherichia coli, antiserum production, and Western blot analysis
The DNA fragment encoding the full-length of WSSV VP35 (corresponding to ORF5 of pms147) was amplified from plasmid pms147 by PCR with the vp35FBamHI/R
Sal1 primer set
(GGATCCATGGTCTCTTCTAGAACA/GTC-GACTTACCAACAAGGATCATC; the underlined bases in-dicate, respectively, the BamH1 and Sal1 restriction sites that were used for subsequent cloning) and ligated to pGEM-T Easy (Promega). After confirming the sequence, the resultant plasmid, pGEM-T-VP35, was cleaved with
BamH1 and Sal1, and the amplified fragment was then
cloned to pQE30 (Qiagen) at BamH1 and Sal1 sites. The resultant recombinant plasmid, pQE30-VP35, was trans-formed into Escherichia coli JM109 strain. Liquid cultures were grown to an optical density of 0.6 at 600 nmand induced with 1 mM isopropyl--D-thiogalactopyranoside (IPTG) for 3 h at 37°C. The recombinant VP35 proteins tagged with six consecutive histidines (6⫻ His-tagged VP35) were purified by using QIAexpressionist nickel-nitrilotriacetic acid (Ni-NTA) metal-affinity chromatogra-phy (Qiagen) according to the manufacturer’s recommen-dations. New Zealand white rabbits were hyperimmu-nized by injection with 100 g proteins emulsified in complete Freund’s adjuvant. Subsequent booster injec-tions were carried out with 100g protein emulsified in
incomplete Freund’s adjuvant. The antiserum was col-lected after the antibody titer had peaked.
For Western blotting analysis, proteins that had been separated in SDS-PAGE were transferred onto a polyvi-nylidene difluoride (PVDF) membrane (Micron Separa-tions, Inc., Minnetonka, MN) by semidry blotting. Mem-branes were blocked in 5% skimmilk (Difco Laborato-ries, Detroit, MI) in TBS (0.2 M NaCl and 50 mM Tris-HCl, pH 7.4). Immunodetection was performed by incubation of the blot in a polyclonal rabbit anti-VP35 serumdiluted 1:2000 in TBS with 5% skimmilk for 1 h at roomtemper-ature. Subsequently, anti-rabbit IgG antibody conjugated with horseradish peroxidase (Sigma, St. Louis, MO) was used at a concentration of 1:2000 and detection was performed with a Western Blot Chemiluminescence Re-agent (NEN Life Sciences, Boston, MA).
Coupled in vitro transcription and translation
To facilitate detection of the HA-fusion protein in the in
vitro transfection experiment (described below), an
addi-tional hemagglutinin (HA) tag was inserted into pcDNA3 (Invitrogen, The Netherlands). The DNA fragment encod-ing the full-length of WSSV VP35 (correspondencod-ing to ORF5 of pms147) was amplified from plasmid pms147 by PCR with the vp35FBamHI/RNot1 primer set (GGATCCATG- GTCTCTTCTAGAACA/GCGGCCGCTTACCAACAAGGAT-CATC; the underlined bases indicate, respectively, the
BamH1 and Not1 restriction sites that were used for
subsequent cloning) and ligated to pGEM-T Easy (Pro-mega). After confirming the sequence, the resultant plas-mid was cleaved with BamH1 and NotI, and the amplified fragment was then cloned to pcDNA3-HA at BamH1 and
NotI sites to give pcDNA3-HA-VP35. Both
autoradiogra-phy and Western blotting were used for VP35 analysis. For autoradiography, pcDNA3-HA-VP35 DNA (1g) was added to TNT Quick Master Mix (Promega) containing [35S]methionine, and the mixture was incubated at 30°C
for 60 to 90 min. For Western blotting, the same proce-dure was followed except that there was no [35
S]methi-onine labeling. After incubation, all samples were sepa-rated by 10% SDS-PAGE (Laemmli, 1970). In the autora-diographic procedure, the expressed proteins were visualized by exposure to X-ray film, while for Western blotting the proteins were detected by antibodies against bacterially synthesized pms147 ORF5 protein.
Recombinant pms147 ORF5 protein (rVP35) overexpression in insect cells
The BacVector Transfection systemwas used to con-struct the Autographa californica multicapsid nucleopoly-hedrovirus (AcMNPV)-based recombinant virus for the expression of VP35 in Sf9 cells. To this end, the full-length coding region of vp35 was excised fromthe plas-mid pQE30-VP35 with HindIII (filling-in with Klenow en-zyme to generate a blunt end) and EcoRI and then cloned
into the baculovirus transfer plasmid pVL1392 EcoRI/
SmaI sites downstreamof the polyhedrin promoter. The
recombinant baculovirus was produced in Sf9 cells by cotransfection with the transfer plasmid (pVL1392-VP35) and linear viral DNA according to the BacVector Trans-fection Kit protocol (Novagen, Madison, WI). The purified recombinant baculovirus was then propagated in Sf9 cells at 27°C in Sf-900 II SFM cell culture medium (Gibco BRL, Frederick, MD). Forty-eight hours after infection, the infected cell proteins were separated by 10% SDS-PAGE and blotted onto PVDF membrane and then detected with antibodies against bacterially synthesized pms147 ORF5 protein.
Transcriptional analysis
Transcriptional analysis of vp35 was performed by Northern blot analysis using total RNA isolated from artificially infected black tiger shrimp as described pre-viously (Tsai et al., 2000a). Briefly, for the isolation of total RNA, frozen tissues (500 mg) from WSSV-infected shrimps were homogenized in 5 ml TRIzol-LS reagent (Gibco BRL) and then subjected to 2-propanol extraction and ethanol precipitation according to the manufactur-er’s recommendations. The total RNA was stored in 75% ethanol at⫺70°C.
Total RNA in 75% ethanol was centrifuged at 14,000 g for 30 min at room temperature. The pellet was resus-pended in DEPC-water and quantified by spectropho-tometer. An aliquot of 10g RNA was treated with 200 U of RNase-free DNase I (Roche, Germany) at 37°C for 30 min to remove any viral genomic DNA contamination and then reextracted with phenol-chloroform. The DNase-treated total RNA was denatured by heating at 85°C for 10 min in 10l DEPC-water containing 100 pmol oligo-dT primer (Roche). The first strand cDNA was synthesized by the addition of 4l Superscript II 5⫻ buffer, 1 l 100 mM DTT, 1l 10 mM dNTPs, 10 U rRNasin (Promega), and 100 U Superscript II reverse transcriptase (Gibco BRL). The reaction proceeded at 37°C for 1 h. The cDNA reaction products were subjected to PCR with the primer set vp35-F/vp35-R (ATGGTCTCTTCTAGAACATC/TCA-CACTTGTGGAGCAACTGG) for the vp35 gene. The -ac-tin gene served as an internal control for RNA quality and amplification efficiency as described previously (Tsai et
al., 2000a).
Detection of WSSV vp35 transcripts in WSSV-infected shrimp by Northern blot hybridization analysis with a
vp35 gene-specific riboprobe
A WSSV vp35-specific [␣-32P]rCTP-labeled riboprobe
was used for Northern blot analysis. To generate the riboprobe, the RNA polymerase promoter addition kit Lig’nScribe (Ambion, Austin, TX) was used in accordance with the manufacturer’s instructions to produce tem-plates fromWSSV vp35-specific PCR products for the in 51 NUCLEAR TARGETING OF A SHRIMP WSSV NUCLEOCAPSID PROTEIN
PCR product (50l) was then used to generate the WSSV
vp35-specific [␣-32P]rCTP-labeled riboprobe by in vitro
transcription (Sambrook et al., 1989) in a 20-l reaction mixture containing 40 U T7 RNA polymerase (Roche) and 0.02 mCi [␣-32P]rCTP for 2 h at 37°C. The reaction
mix-ture was then treated with 200 U RNase-free DNase I for 30 min at room temperature, terminated at 68°C for 15 min, and filtered through a Sephadex G50 column.
Total RNA (10g) was separated on 1% formaldehyde-agarose gel and transferred onto a Hybond-N⫹ mem-brane (Amersham Pharmacia Biotech, Inc., Piscataway, NJ) (Sambrook et al., 1989). The membrane was prehy-bridized for 1 h at 65°C in a prehybridization buffer (0.25 M phosphate buffer, 1 mM EDTA, 1% BSA, and 7% SDS) and then hybridized with a specific [␣-32
P]rCTP-labeled riboprobe that was added to the buffer. After hybridiza-tion for 16 h at 65°C the membrane was washed for 5 min with wash buffer I (2⫻ SSC, and 0.5% SDS) at room temperature, 30 min with wash buffer II (2⫻ SSC and 0.1% SDS) at roomtemperature, and 30 min with wash buffer III (0.1⫻ SSC and 0.1% SDS) at 65°C. The mem-brane was then exposed to Kodak BioMax MR filmvia an intensifying screen for several days at ⫺70°C and the filmwas then developed.
Nuclear localization of WSSV VP35 in Sf9 cell culture, transfection, and immunofluorescent staining
The DNA fragment encoding the full-length of WSSV VP35 (corresponding to ORF5 of pms147) was amplified from plasmid pms147 by PCR with the vp35-pIZ-F1/vp35-pIZ-R1 primer set (AAGCTTATGGTCTCTTCTAGAACA/ GGATCCCCAACAAGGATCATCAAT; the underlined bases indicate, respectively, the HindIII and BamH1 re-striction sites that were used for subsequent cloning) and ligated to pGEM-T Easy (Promega). After confirming the sequence, the resultant plasmid was cleaved with
HindIII and BamH1, and the amplified fragment was then
cloned to pIZ/V5-His at HindIII and BamH1 sites to give pIZ/V5-His-VP35. Four alanines (24AAAA27) were selected
to replace the first NLS motif (24KRKR27) using overlap
extension (Ho et al., 1989). Two small fragments were amplified with the vp35-pIZ-F1/vp35-NLS1-R1 (TTCTG-GCTCCGCCGCCGCCGCGGTGGGGAG) and vp35-pIZ-R1/vp35-NLS1-F1
(CTCCCCACCGCGGCGGCGGCGG-pIZ/V5-His-VP35m1, and pIZ/V5-His-VP35m2 at 70% con-fluency.
Sf9 cells were cultured at 27°C in Sf-900 II SFM (Gibco BRL). Transfection was performed in a 6-well format. About 106 cells were seeded in each well of the 6-well
culture plate and cultured overnight. Plasmid pIZ/V5-His-VP35 and pIZ/V5-His-mpIZ/V5-His-VP35 (⬃2.5 g) was mixed with Insectin-Plus liposomes (Invitrogen) in the medium and laid over the cultured cells according to the manufactur-er’s recommendations. Twenty-four hours after transfec-tion, cells were fixed in 10% paraformaldehyde for 10 min at 4°C, treated with 0.1% Triton X-100 for 10 min at 4°C, washed extensively with PBS, and reacted with anti-serumagainst VP35 at 4°C overnight. After washing, the cells were then reacted with carboxymethylindocyanine dye (Cy3) conjugated goat anti-rabbit IgG antibody (Sigma). Counterstaining of the nucleus was performed with 4⬘-6⬘-diamidino-2 phenylindole dihydrochloride (DAPI, Vector Laboratories, Burlingame, CA). Cy3 and DAPI were excited by light at 552 and 372 nm, respec-tively, and fluorescence was observed (at 565 nmfor Cy3 and 456 nm for DAPI) with an Olympus IX70 microscopy systemwith a fluorescent microscopy apparatus.
ACKNOWLEDGMENTS
This work was support by the National Science Council Grants NSC89-2311-B-002-042 and NSC90-2611-B-002-008 and Education Ad-ministration Grant 89-B-FA01-1-4. We are indebted to Mr. Paul Barlow and Miss Yu-Tsan Lin for their helpful criticismof the manuscript.
REFERENCES
Altschul, S. F., Madden, T. L., Schaffer, A. A., Zhang, J., Zhang, Z., Miller, W., and Lipman, D. J. (1997). Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 25, 3389–3402.
Bairoch, A., Bucher, P., and Hofmann, K. (1997). The PROSITE database, its status in 1997. Nucleic Acids Res. 25, 217–221.
Chou, H. Y., Huang, C. Y., Wang, C. H., Kou, G. H., and Lo, C. F. (1995). Pathogenicity of a baculovirus infection causing white spot syn-drome in cultured penaeid shrimp in Taiwan. Dis. Aquat. Org. 23, 165–173.
Flegel, T. W. (1997). Special topic review: Major viral diseases of the black tiger prawn (Penaeus monodon) in Thailand. World J. Microbiol.
Biotech. 13, 422–433.
Greber, U. F., Suomalainen, M., Stidwill, R. P., Boucke, K., Ebersold, M. W., and Helenius, A. (1997). The role of the nuclear pore complex in adenovirus DNA entry. EMBO J. 16, 5998–6007.
Greber, U. F., Webster, P., Weber, J., and Helenius, A. (1996). The role of the adenovirus protease in virus entry into cells. EMBO J. 15, 1766– 1777.
Ho, S. N., Hunt, H. D., Horton, R. M., Pullen, J. K., and Pease L. R. (1989). Site-directed mutagenesis by overlap extension using the polymer-ase chain reaction. Gene 77, 51–59.
Hopp, T. P., and Woods, K. R. (1981). Prediction of protein antigenic determinants from amino acid sequences. Proc. Natl. Acad. Sci. USA 78, 3824–3828.
Jans, D. A., and Hubner, S. (1996). Regulation of protein transport to the nucleus: Central role of phosphorylation. Physiol. Rev. 76, 651–685. Kalderon, D., Roberts, B. L., Richardson, W. D., and Smith, A. E. (1984). A short amino acid sequence able to specify nuclear localization.
Cell 39, 499–509.
Kann, M., Sodeik, B., Vlachou, A., Gerlich, W. H., and Helenius, A. (1999). Phosphorylation-dependent binding of hepatitis B virus core parti-cles to the nuclear pore complex. J. Cell Biol. 145, 45–55.
Kasamatsu, H., and Nakanishi, A. (1998). How do animal DNA viruses get to the nucleus? Annu. Rev. Microbiol. 52, 627–686.
Kozak, M. (1989). The scanning model for translation: An update. J. Cell
Biol. 108, 229–241.
Kou, G. H., Peng, S. E., Chiu, Y. L., and Lo, C. F. (1998). Tissue distribu-tion of white spot syndrome virus (WSSV) in shrimp and crabs. In “Advances in Shrimp Biotechnology” (T. W. Flegel, Ed.), pp. 267–271. National Center for Genetic Engineering and Biotechnology, Bangkok.
Laemmli, U. K. (1970). Cleavage of structural proteins during the as-sembly of the head of bacteriophage T4. Nature 227, 680–685. Liu, W. J., Yu, H. T., Peng, S. E., Chang, Y. S., Pien, H. W., Lin, C. J., Huang,
C. J., Tsai, M. F., Huang, C. J., Wang, C. H., Lin, J. Y., Lo, C. F., and Kou, G. H. (2001). Cloning, characterization and phylogenetic analysis of a shrimp white spot syndrome virus (WSSV) gene that encodes a protein kinase. Virology 289, 362–377, doi:10.1006/viro.2001.1091. Lo, C. F., Chang, Y. S., Cheng, C. T., and Kou, G. H. (1998). PCR
monitoring cultured shrimp for white spot syndrome virus (WSSV) in growout ponds. In “Advances in Shrimp Biotechnology” (T. W. Flegel, Ed.), pp. 281–286. National Center for Genetic Engineering and Biotechnology, Bangkok.
Lo, C. F., Ho, C. H., Chen, C. H., Liu, K. F., Chiu, Y. L., Yeh, P. Y., Peng, S. E., Hsu, H. C., Liu, H. C., Chang, C. F., Su, M. S., Wang, C. H., and Kou, G. H. (1997). Detection and tissue tropismof white spot syn-drome baculovirus (WSBV) in captured brooders of Penaeus
mon-odon with a special emphasis on reproductive organs. Dis. Aquat. Org. 30, 53–72.
Lo, C. F., Hsu, H. C., Tsai, M. F., Ho, C. H., Peng, S. E., Kou,. G. H., and Lightner, D. V. (1999). Specific genomic fragment analysis of different geographical clinical samples of shrimp white spot syndrome virus.
Dis. Aquat. Org. 35, 175–185.
Lo, C. F., Leu, J. H., Ho, C. H., Chen, C. H., Peng, S. E., Chen, Y. T., Chou, C. M., Yeh, P. Y., Huang, C. J., Chou, H. Y., Wang, C. H., and Kou, G. H. (1996). Detection of baculovirus associated with white spot syn-drome (WSBV) in penaeid shrimps using polymerase chain reaction.
Dis. Aquat. Org. 25, 133–141.
Makkerh, J. P. S., Dingwall, C., and Laskey, R. A. (1996). Comparative mutagenesis of nuclear localization signals reveals the importance of neutral and acidic amino acids. Curr. Biol. 6, 1025–1027. Marsh, M., and Helenius, A. (1989). Virus entry into animal cells. Adv.
Virus Res. 36, 107–151.
Mosevitsky, M. I., Novitskaya, V. A., Plekhanov A. Y., and Skladchikova, G. Y. (1994). Neuronal protein GAP-43 is a member of novel group of brain acidic-soluble proteins (BASPs). Neurosci. Res. 19, 223–228. Nadala, Jr., E. C. B., Tapay, L. M., and Loh, P. C. (1998). Characterization
of a nonoccluded baculovirus-like agent pathogenic to penaeid shrimp. Dis. Aquat. Org. 33, 221–229.
Nakanishi, A., Clever J., Yamada. M., Li, P. P., and Kasamatsu, H. (1996). Association with capsid proteins promotes nuclear targeting of sim-ian virus 40 DNA. Proc. Natl. Acad. Sci. USA 93, 96–100.
Pearson, W. R., and Lipman, D. J. (1988). Improved tools for biological sequence analysis. Proc. Natl. Acad. Sci. USA 85, 2444–2448. Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989). “Molecular Cloning:
A Laboratory Manual,” Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
Sodeik, B., Ebersold, M. W., and Helenius, A. (1997). Microtubule-mediated transport of incoming herpes simplex virus 1 capsids to the nucleus. J. Cell Biol. 136, 1007–1021.
Suomalainen, M., Nakano, M. Y., Keller, S., Boucke, K., Stidwill, R. P., and Greber, U. F. (1999). Microtubule-mediated plus- and minus end-directed motilities are competing processes for nuclear targeting of adenoviruses. J. Cell Biol. 144, 657–672.
Tidona, C. A., and Darai, G. (1997). Molecular anatomy of lymphocystis disease virus. Arch. Virol. Suppl. 13, 49–56.
Tsai, M. F., Lo, C. F., van Hulten, M. C. W., Tzeng, H. F., Chou, C. M., Huang, C. J., Wang, C. H., Lin, J. Y., Vlak, J. M., and Kou, G. H. (2000a). Transcriptional analysis of the ribonucleotide reductase genes of shrimp white spot syndrome virus. Virology 277, 92–99, doi: 10.1006/ viro.2000.0956.
Tsai, M. F., Yu, H. T., Tzeng, H. F., Leu, J. H., Chou, C. M., Huang, C. J., Wang, C. H., Lin, J. Y., Kou, G. H., and Lo, C. F. (2000b). Identification and characterization of a shrimp white spot syndrome virus (WSSV) gene the encodes a novel chimeric polypeptide of cellular-type thymidine kinase and thymidylate kinase. Virology 277, 100–110, doi: 10.1006/viro.2000.0957.
van Hulten, M. C. W., Witteveldt, J., Peters, S., Kloosterboer, N., Tarchini, R., Fiers, M., Sandbrink, H., Lankhorst, R. K., and Vlak, J. M. (2001). The white spot syndrome virus DNA genome sequence. Virology 286, 7–22, doi: 10.1006/viro.2001.1002.
Vihinen-Ranta, M., Yuan, W., and Parrish, C. R. (2000). Cytoplasmic trafficking of the canine parvovirus capsid and its role in infection and nuclear transport. J. Virol. 74, 4853–4859.
Wang, C. H., Lo, C. F., Leu, J. H., Chou, C. M., Yeh, P. Y., Chou, H. Y., Tung, M. C., Chang, C. F., Su, M. S., and Kou, G. H. (1995). Purification and genomic analysis of baculovirus associated with white spot syn-drome (WSBV) of Panaeus monodon. Dis. Aquat. Org. 23, 239–242. Whittaker, G. R., and Helenius, A. (1998). Nuclear import and export of
viruses and virus genomes. Virology 246, 1–23, doi: 10.1006/ viro.1998.9156.
Wittek., R. (1982). Organization and expression of the poxvirus genome.
Experientia 38, 285–297.
Wongteerasupaya, C., Vickers, J. E., Sriurairatana, S., Nash, G. L., Akarajamorn, A., Boonsaeng, V., Panyim, S., Tassankajon, A., Withya-nchumnarnkul, B., and Flegel, T. W. (1995). A non-occluded, systemic baculovirus that occurs in cells of ectodermal and mesodermal origin and causes high mortality in black tiger prawn Penaeus
mon-odon. Dis. Aquat. Org. 21, 69–77.
Wychowski, C., Benichou, D., and Girard, M. (1986). A domain of SV40 capsid polypeptide VP1 that specifies migration into the cell nucleus.
EMBO J. 5, 2569–2576.
Wychowski, C., Benichou, D., and Girard, M. (1987). The intranuclear localization of simian virus 40 polypeptides Vp2 and Vp3 depends on a specific amino acid sequence. J. Virol. 61, 3862–3869.
Zirwes, R. F., Schmidt-Zachmann, M. S., and Franke, W. W. (1997). Identification of a small, very acidic constitutive nucleolar protein (NO29) as a member of the nucleoplasmin family. Proc. Natl. Acad.
Sci. USA 94, 11387–11392.
53 NUCLEAR TARGETING OF A SHRIMP WSSV NUCLEOCAPSID PROTEIN