Research Article
Systematic Analysis of the Association between Gut
Flora and Obesity through High-Throughput Sequencing and
Bioinformatics Approaches
Chih-Min Chiu,
1Wei-Chih Huang,
1Shun-Long Weng,
1,2,3,4Han-Chi Tseng,
5Chao Liang,
6Wei-Chi Wang,
6Ting Yang,
1Tzu-Ling Yang,
1Chen-Tsung Weng,
6Tzu-Hao Chang,
7and Hsien-Da Huang
1,8,9,101Institute of Bioinformatics and Systems Biology, National Chiao Tung University, Hsinchu 300, Taiwan 2Department of Obstetrics and Gynecology, Hsinchu Mackay Memorial Hospital, Hsinchu 300, Taiwan 3Mackay Medicine, Nursing and Management College, Taipei 104, Taiwan
4Department of Medicine, Mackay Medical College, New Taipei City 251, Taiwan 5Tseng Han-Chi General Hospital, Nantou 542, Taiwan
6Health GeneTech Corporation, Taoyuan 330, Taiwan
7Graduate Institute of Biomedical Informatics, Taipei Medical University, Taipei 110, Taiwan
8Department of Biological Science and Technology, National Chiao Tung University, Hsinchu 300, Taiwan 9Center for Bioinformatics Research, National Chiao Tung University, Hsinchu 300, Taiwan
10Department of Biomedical Science and Environmental Biology, Kaohsiung Medical University, Kaohsiung 807, Taiwan
Correspondence should be addressed to Tzu-Hao Chang; kevinchang@tmu.edu.tw and Hsien-Da Huang; bryan@mail.nctu.edu.tw Received 11 April 2014; Accepted 27 June 2014; Published 14 August 2014
Academic Editor: Li-Ching Wu
Copyright © 2014 Chih-Min Chiu et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Eighty-one stool samples from Taiwanese were collected for analysis of the association between the gut flora and obesity. The supervised analysis showed that the most, abundant genera of bacteria in normal samples (from people with a body mass index (BMI)≤ 24) were Bacteroides (27.7%), Prevotella (19.4%), Escherichia (12%), Phascolarctobacterium (3.9%), and Eubacterium (3.5%). The most abundant genera of bacteria in case samples (with a BMI≥ 27) were Bacteroides (29%), Prevotella (21%), Escherichia (7.4%), Megamonas (5.1%), and Phascolarctobacterium (3.8%). A principal coordinate analysis (PCoA) demonstrated that normal samples were clustered more compactly than case samples. An unsupervised analysis demonstrated that bacterial communities in the gut were clustered into two main groups: like and OB-like groups. Remarkably, most normal samples (78%) were clustered in the N-like group, and most case samples (81%) were clustered in the OB-N-like group (Fisher’s𝑃 value = 1.61𝐸−07). The results showed that bacterial communities in the gut were highly associated with obesity. This is the first study in Taiwan to investigate the association between human gut flora and obesity, and the results provide new insights into the correlation of bacteria with the rising trend in obesity.
1. Background
Enterobacteria, or gut microbiota, in the human gastroin-testinal (GI) tract play important roles in the body’s func-tions. For example, they can regulate immune responses and metabolic functions of the host [1–4]. The gut microbiota
is frequently used to study the association between human health and an individual’s lifestyle. In 2011, three major kinds of enterotypes consisting of Bacteroides, Prevotella, and Ruminococcus [5] provided new perspectives to classify individuals. Ruminococcus was reported to be an ambiguous enterotype, and the Bacteroides and Prevotella enterotypes
Volume 2014, Article ID 906168, 10 pages http://dx.doi.org/10.1155/2014/906168
were associated with dietary habits [6]. Animal protein and saturated fats were highly correlated with the Bacteroides enterotype. Low meat intake and plant-based nutrition with high carbohydrates were correlated with the Prevotella enterotype.
The Human Microbiome Project (HMP) [7] was launched by the US National Institutes of Health in 2008, and understanding the relationship between human health and microbiota that live in or on the human body was recognized as an important concept. To decipher this relationship, high-throughput sequencing, also called next-generation sequenc-ing (NGS), supportsequenc-ing a large number of sequences, can be used to sequence 16S ribosomal (r)RNA to construct complex microbial community profiles. 16S rRNA is considered the standard for studying microbial communities and assigning taxonomy to bacteria. Compared to conventional polymerase chain reaction- (PCR-) based or culture-based methods, 16S rRNA sequencing by NGS can detect hundreds to thousands of bacteria at one time and offer relative quantification of the bacteria. Interactions between different bacterial communi-ties and their environments can be comprehensively analyzed by metagenomics research. Associations between diseases and specific bacteria have been described in previous studies, for example, type 2 diabetes [8–11], irritable bowel syndrome (IBS) [12–14], and colorectal cancer (CRC) [15–17]. Moreover, some bacteria which are significantly associated with specific diseases were thought to be biomarkers for construction of a disease risk prediction model [8,18].
Obesity is a major public health problem worldwide, and its prevalence is rapidly increasing [19]. Obesity is related to several disorders, including type 2 diabetes [20–
23], cardiovascular disease [24–26], and cancer [26–28]. Recently, obesity was shown to be associated with an alter-ation of the gut microbiota, both in human [29–32] and animal models [30,33, 34]. It was observed that a reduced proportion of the Bacteroidetes and increased proportion of the Firmicutes were associated with human obesity [30,
35, 36]. Also, an increase of Actinobacteria in obese indi-viduals was reported [35]. In another study, amounts of Archaea and Methanobacteriales were positively correlated with obesity [37], and their amounts in obesity samples decreased or disappeared after gastric bypass surgery. In addition, another study also mentioned that the amounts of Bifidobacterium and Ruminococcus decreased in obe-sity samples [38]. However, some studies indicated that the ratio of the proportions of Bacteroidetes and Firmi-cutesis contradictory [39] or not associated [5, 40] with obesity.
With different ethnicities and regions, dietary habits and environmental factors can widely vary, and there is a lack of studies focusing on Taiwanese samples. Therefore, herein we collected 81 stool samples from Taiwanese for analysis of the association between gut flora and obesity. According to a study by Pan et al. [41], Taiwan adopted body mass index (BMI) values of 24 and 27 as the cutoff points for being overweight and obese, respectively. In this study, the stools of 36 obese (BMI≥ 27) and 45 normal persons (BMI ≤ 24) were
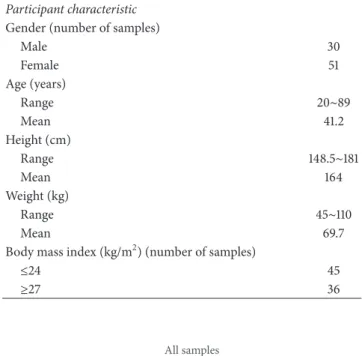
Table 1: Study participant characteristics and demographics. Participant characteristic
Gender (number of samples)
Male 30 Female 51 Age (years) Range 20∼89 Mean 41.2 Height (cm) Range 148.5∼181 Mean 164 Weight (kg) Range 45∼110 Mean 69.7
Body mass index (kg/m2) (number of samples)
≤24 45 ≥27 36 Bacteroides (28%) Prevotella (20%) Escherichia (9.7%) Phascolarctobacterium (3.9%) Eubacterium (3.2%) Megamonas (3%) Faecalibacterium (2.9%) Gemmiger (2.2%)Sutterella (2%) Fusobacterium (1.9%)Salmonella (1.9%) Megasphaera (1.3%)Dialister (1.2%) Bifidobacterium (1.1%)Akkermansia (1%) Others (17%) All samples
Figure 1: The distribution of genera among all samples.
collected, and 16S rRNA sequencing was used to assess the association between obesity and the taxonomic composition of the gut microbiota.
2. Results
Participant metadata are summarized inTable 1, and detailed sample profiles are given in Table S1 available online in Sup-plementary material athttp://dx.doi.org/10.1155/2014/906168, including the number of reads, gender, age, height, weight, and BMI. In total, 4,152,740 sequence reads were obtained from the 81 samples, and a mean of 51,268 reads with a median read length of 125 bp was obtained per study participant. Sequence reads were processed through our taxonomic map-ping process, and the distribution of genera in samples is depicted inFigure 1. The sequencing results showed that the most abundant genera in all samples were Bacteroides (28%),
Prevotella (20%), Escherichia (9.7%), Phascolarctobacterium
(3.9%), Eubacterium (3.2%), Megamonas (3%),
2.1. Unsupervised Clustering Analysis. Hierarchical clustering
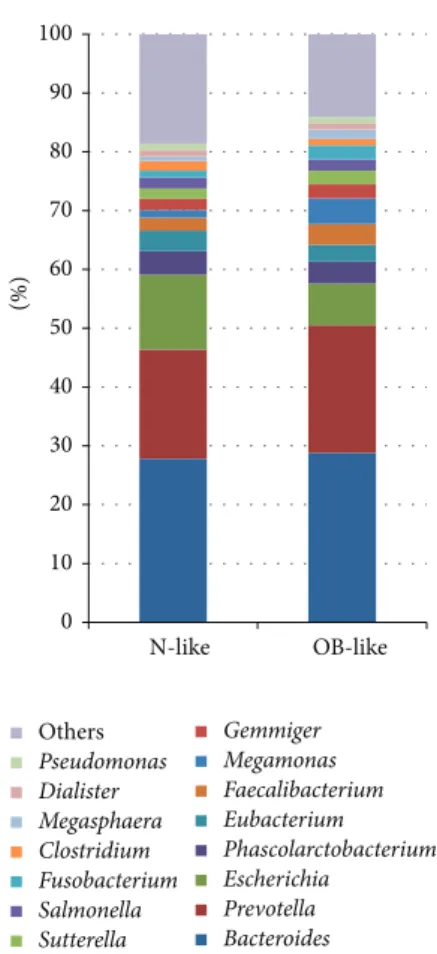
was performed using the UniFrac unweighted distance, and gut bacterial communities and clinical values of each sample are shown in Figure 2. The results demonstrate that the bacterial communities in the gut were clustered into two main groups: an N-like group (including the N1 and N2 subgroups) and an OB-like group (including the OB1, OB2, OB3, and OB4 subgroups). Figure 3 shows that the most abundant genera in N-like samples were Bacteroides (27.8%), Prevotella (18.6%), Escherichia (12.7%), Phascolarctobacterium (4%), and Eubacterium (3.5%). The most abundant genera in OB-like samples were Bacteroides (28.8%), Prevotella (21.7%),
Escherichia (7.1%), Megamonas (4.4%), and Phascolarctobac-terium (3.7%). Remarkably, most normal samples (78%) were
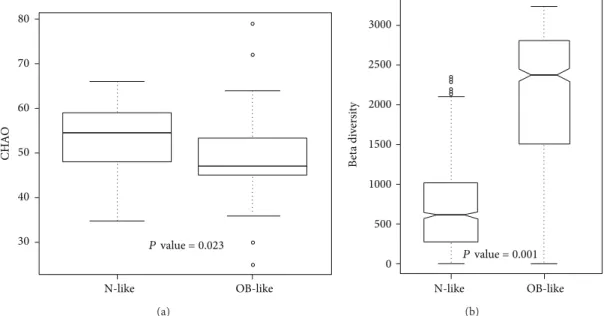
clustered in the N-like group, and most case samples (81%) were clustered in the OB-like group (Fisher’s𝑃 value = 1.61𝐸− 07). The results showed that gut bacterial community types were highly associated with obesity. The genera diversity analysis showed that the bacterial communities in the N-like group exhibited significantly higher alpha diversity and lower beta diversity than those in the OB-like group (Figure 4).
2.2. Supervised Clustering Analysis. To investigate the
asso-ciation between gut bacterial communities and obesity, 45 stool samples of participants with a BMI of≤ 24 were defined as normal samples, and 36 samples of participants with a BMI ≥ 27 were used as case samples. Figure 5shows that the most abundant bacteria in normal samples were
Bac-teroides (27.7%), Prevotella (19.4%), Escherichia (12%), Phas-colarctobacterium (3.9%), and Eubacterium (3.5%). The most
abundant bacteria in case samples were Bacteroides (29%),
Prevotella (21%), Escherichia (7.4%), Megamonas (5.1%), and Phascolarctobacterium (3.8%). Normal samples had a
signif-icantly higher proportion of Escherichia, while case samples had a higher proportion of Megamonas.
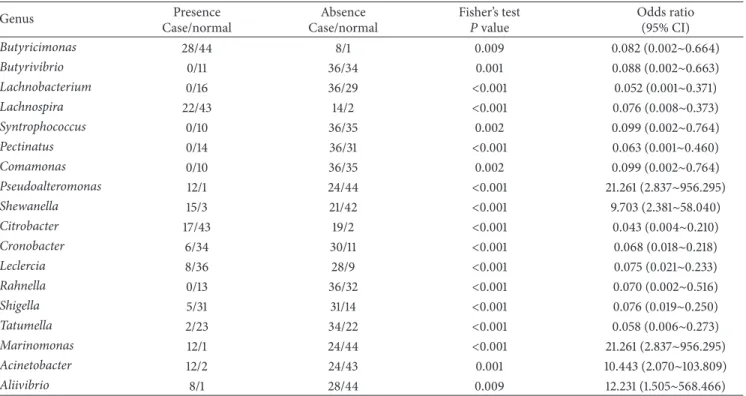
Genera with significantly different proportions between normal and case samples are listed inTable 2. Additionally, genera with a significantly different presence between normal and case samples are listed in Table 3, and significantly different species are also provided in Table S2. The genera of Shewanella, Citrobacter, Cronobacter, Leclercia, Tatumella,
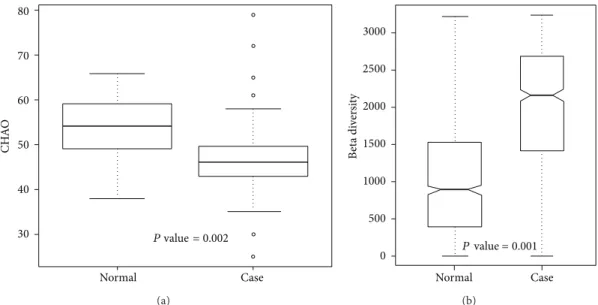
and Acinetobacter exhibited significant differences in both
proportions and presence. Unweighted alpha and beta diver-sities of genera in the normal and case samples are shown in Figures 6(a) and 6(b), respectively. The results showed that the bacterial communities in normal samples exhibited significantly higher alpha diversity and lower beta diversity than those in case
samples.
A PCoA of gut bacterial communities is shown in
Figure 7. The results showed that most normal samples (green nodes) were located in the bottom left area, and case samples (red nodes) were spread in other areas (Figure 7(a)). Samples in the N1, N2, OB1, OB2, OB3, and OB4 subgroups are depicted inFigure 7(b). The results show that bacterial communities of N1 and N2 were highly associated with normal-weight individuals, and others were associated with obese individuals. BMI Subgroups N1 N2 OB1 OB2 OB3 OB4 BMI ≥ 27 (case) BMI ≤ 24 (normal)
Figure 2: Bacterial communities in the samples.
0 10 20 30 40 50 60 70 80 90 100 N-like OB-like Others Pseudomonas Dialister Megasphaera Clostridium Fusobacterium Salmonella Sutterella Gemmiger Megamonas Faecalibacterium Eubacterium Phascolarctobacterium Escherichia Prevotella Bacteroides (%)
Figure 3: Relatively abundant genera in the N-like and OB-like groups.
Table 2: Genera with significantly different proportions between normal and case samples.
Genus Fold change
(case/control)
KS test
𝑃 value ANOVA𝑃 value Case mean Control mean
Collinsella 0.56 0.01 0.37 0.001 0.002 Barnesiella 0.61 0.01 0.26 0.002 0.003 Clostridium 0.52 0.01 0.01 0.009 0.017 Coprococcus 0.54 0.04 0.37 0.001 0.001 Lachnospira 1.68 0.02 0.27 0.003 0.002 Oscillibacter 0.55 0.01 0.02 0.001 0.001 Megamonas 5.70 0.08 0.01 0.051 0.009 Veillonella 0.47 0.01 0.27 0.002 0.004 Shewanella 4.69 0.00 0.04 0.001 <0.001 Citrobacter 0.26 0.00 0.11 0.002 0.006 Cronobacter 0.08 0.00 0.00 <0.001 0.002 Enterobacter 0.38 0.00 0.07 0.001 0.002 Erwinia 0.18 0.00 0.00 <0.001 0.002 Escherichia 0.62 0.00 0.04 0.074 0.120 Leclercia 0.09 0.00 0.06 0.001 0.005 Morganella 0.05 0.01 0.07 <0.001 0.001 Serratia 0.02 0.00 0.00 <0.001 0.018 Tatumella 0.06 0.00 0.00 <0.001 0.001 Halomonas 3.51 0.00 0.08 0.005 0.002 Acinetobacter 116.08 0.00 0.03 0.002 <0.001
KS: Kolmogorov-Smirnov; ANOVA: analysis of variance.
Table 3: Genera with a significantly different presence between normal and case samples.
Genus Presence
Case/normal
Absence Case/normal
Fisher’s test
𝑃 value Odds ratio(95% CI)
Butyricimonas 28/44 8/1 0.009 0.082 (0.002∼0.664) Butyrivibrio 0/11 36/34 0.001 0.088 (0.002∼0.663) Lachnobacterium 0/16 36/29 <0.001 0.052 (0.001∼0.371) Lachnospira 22/43 14/2 <0.001 0.076 (0.008∼0.373) Syntrophococcus 0/10 36/35 0.002 0.099 (0.002∼0.764) Pectinatus 0/14 36/31 <0.001 0.063 (0.001∼0.460) Comamonas 0/10 36/35 0.002 0.099 (0.002∼0.764) Pseudoalteromonas 12/1 24/44 <0.001 21.261 (2.837∼956.295) Shewanella 15/3 21/42 <0.001 9.703 (2.381∼58.040) Citrobacter 17/43 19/2 <0.001 0.043 (0.004∼0.210) Cronobacter 6/34 30/11 <0.001 0.068 (0.018∼0.218) Leclercia 8/36 28/9 <0.001 0.075 (0.021∼0.233) Rahnella 0/13 36/32 <0.001 0.070 (0.002∼0.516) Shigella 5/31 31/14 <0.001 0.076 (0.019∼0.250) Tatumella 2/23 34/22 <0.001 0.058 (0.006∼0.273) Marinomonas 12/1 24/44 <0.001 21.261 (2.837∼956.295) Acinetobacter 12/2 24/43 0.001 10.443 (2.070∼103.809) Aliivibrio 8/1 28/44 0.009 12.231 (1.505∼568.466)
OB-like N-like CH A O 80 70 60 50 40 30 P value = 0.023 (a) OB-like N-like B et a di ve rs it y 3000 2500 2000 1500 1000 500 0 P value = 0.001 (b)
Figure 4: Unweighted (a) alpha diversity and (b) beta diversity of bacterial communities in the N-like and OB-like groups.
Bacteroides (27.7%) Prevotella (19.4%) Escherichia (12%) Phascolarctobacterium (3.9%) Eubacterium (3.5%) Megamonas (0.9%) Faecalibacterium (2.5%)Gemmiger (1.9%) Sutterella (1.8%) Fusobacterium (1.2%)Salmonella (2%) Megasphaera (0.7%)Dialister (1.1%) Bifidobacterium (1%)Akkermansia (0.8%) Others (19.6%) Normal samples (a) Bacteroides (29%) Prevotella (21%) Escherichia (7.4%) Phascolarctobacterium (3.8%) Eubacterium (2.8%) Megamonas (5.1%) Faecalibacterium (3.2%) Gemmiger (2.5%) Sutterella (2.2%) Fusobacterium (2.5%) Salmonella (1.7%) Megasphaera (1.8%)Dialister (1.2%) Bifidobacterium (1.1%)Akkermansia (1%) Others (13.7%) Case samples (b)
Figure 5: Relatively abundant genera in the normal and case samples.
2.3. Potential Markers for Classification of Normal Weight and Obesity. The identified bacteria with statistical
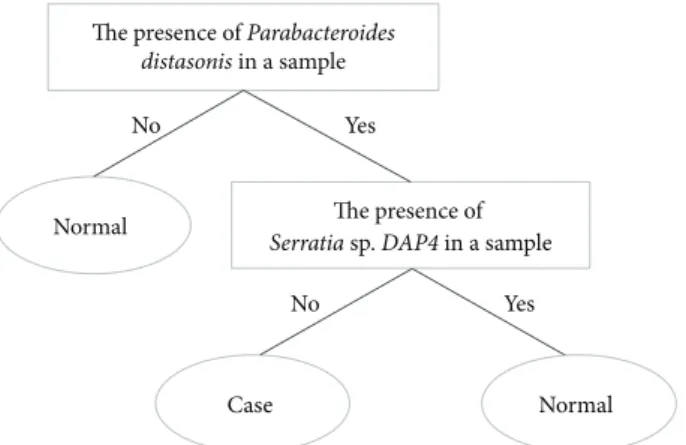
signifi-cance were used for rule-based clustering. Threefold cross-validation was used to evaluate the performance of the clas-sification model. Two out of the significant species in Table S2, Parabacteroides distasonis and Serratia sp. DAP4, were selected as discriminating factors in the J48 decision tree. As shown inFigure 8, the classification rules are described as follows. (1) a sample is classified as normal if Parabacteroides
distasonis was absent. (2) A sample with the presence of Parabacteroides distasonis and absence of Serratia sp. DAP4
was classified as a case; otherwise, it was classified as normal. As shown in Table S3, the classifier performed well, and the area under the receiver operating characteristic curve (AUC)
was 0.813. The results showed that Parabacteroides distasonis and Serratia sp. DAP4 might be potential markers for further clinical analysis and investigation of obesity.
3. Discussion
Several relatively abundant genera were identified in sam-ples (Figure 5), including Bacteroides, Prevotella, Escherichia,
Phascolarctobacterium, Eubacterium, Megamonas, Faecal-ibacterium, Gemmiger, Sutterella, Fusobacterium, Salmonella, Megasphaera, Dialister, Bifidobacterium, and Akkermansia.
In previous studies, the presence of Bacteroides, Prevotella, and Sutterella was negatively associated with obesity [36,42,
CH A O 80 70 60 50 40 30 P value = 0.002 Case Normal (a) B et a di ve rs it y 3000 2500 2000 1500 1000 500 0 P value = 0.001 Case Normal (b)
Figure 6: Unweighted (a) alpha diversity and (b) beta diversity of bacterial communities in case and control samples.
Case Normal 0.2 0.1 0.0 −0.1 −0.2 0.4 0.3 0.2 0.1 0.0 −0.1 −0.2 PC 2 (13 %) PC1 (17.3%) (a) d = 0.1 OB-1 OB-4 N-2 N OB-3 OB-2 (b)
Figure 7: Unweighted principal coordinate analysis plot of (a) case and normal samples, (b) samples in N1, N2, OB1, OB2, OB3, and OB4 subgroups.
43]. In other related gastrointestinal diseases, Prevotella was increased in children diagnosed with IBS [44]. Bacteroides,
Eubacterium, and Prevotella were increased, and Faecalibac-terium was reduced in CRC patients [45, 46]. Increased
Bacteroides and reduced Eubacterium and Prevotella were
also found in a rat model of CRC [47].
At the genus level, the presence of Acinetobacter,
Ali-ivibrio, Marinomonas, Pseudoalteromonas, and Shewanella
had positive associations with obesity (Table 3).
Acinetobac-ter is a genus of Gram-negative bacAcinetobac-teria, and the species Acinetobacter baumannii is a key pathogen of infections in
hospitals [48]. Aliivibrio is a reclassified genus from the “Vibrio fischeri species group” [49], and species of Aliivibrio are symbiotic with marine animals or are described as fish pathogens [50–52]. Shewanella is a genus of marine bacteria, and some species can cause infections [53,54]. Lachnospira
The presence of Parabacteroides
distasonis in a sample
The presence of
Serratia sp. DAP4 in a sample
Normal No Case Yes Yes No Normal
Figure 8: Classification rule and potential markers for discriminat-ing between obese and normal-weight individuals.
[37], Citrobacter [43], and Shigella [43] were reported to be positively associated with obesity. Lachnobacterium [18] showed a negative association with obesity.
At the species level, the presence of Parabacteroides
dis-tasonis, Lactobacillus kunkeei, Pseudoalteromonas piscicida, Shewanella algae, Marinomonas posidonica, and Aliivibrio fischeri was positively associated with obesity (Table S2).
Species of Bacteroides and Parabacteroides represent oppor-tunistic pathogens in infectious diseases, and they are able to develop antimicrobial drug resistance [55]. Parabacteroides
distasonis, previously known as Bacteroides distasonis [56], is prominently found in the gut of healthy individuals [57]. It is also related to improved human bowel health release [58] and negatively associated with celiac disease [59]. Our results revealed a positive association between
Parabacteroides distasonis and obesity. Blautia producta [60] and Enterobacter cloacae [61] were suggested to be related to a high-fat diet causing obesity in a mouse model. Serratia is a genus of Gram-negative, facultatively anaerobic, rod-shaped bacteria. In hospitals, Serratia species tend to colonize the respiratory and urinary tracts causing nosocomial infections [62,63]. In related studies of the GI tract, Serratia increased in formula-fed mice [64] and was positively correlated with infants with colic [65].
In the alpha diversity analysis (Figure 6(a)), the Chao richness index between normal and case groups exhibited a significant difference (𝑃 = 0.002). This shows that bacterial communities in normal samples had a greater genera richness than those in case samples. Results of the beta diversity analysis (Figure 6(b)) showed that bacterial communities in normal samples were more similar than those in case samples. The unweighted PCoA plot (Figure 7) showed that bacterial communities in the N-like group (including N1 and N2) were highly associated with normal individuals, and bacterial communities in the OB-like group (including OB1, OB2, OB3, and OB4) were more associated with obese individuals. The unsupervised clustering heatmap of all samples (Figure
S1(A)) was generated using Spearman correlations, and the results showed that most normal samples and most case samples were, respectively, clustered together, when all genera were used for clustering. However, when only relatively abundant genera were used for clustering, normal, and case samples were interwoven with each other (Figure S1(B)). This indicates that some genera found in small proportions might be important for distinguishing obese from normal individuals.
4. Conclusions
This is the first study in Taiwan to investigate the association between human gut microbiota and obesity using metage-nomic sequencing. The results showed that bacterial com-munities in the gut were clustered into N-like and OB-like groups which were highly associated with normal and obese subjects, respectively. Several relatively abundant bacteria with significantly different distributions between normal and case samples were identified and used to establish a rule-based classification model. Although detailed functional roles or mechanisms of these bacteria are needed for further validation, the results provide new insights about bacterial communities in the gut with a rising trend of obesity.
5. Methods
5.1. Sample Collection and DNA Extraction. Eighty-one stool
samples were collected by Sigma-transwab (Medical Wire) into a tube with Liquid Amies Transport Medium and stored at 4∘C until being processed. Fresh faeces were obtained from participants, and DNA was directly extracted from stool samples using a QIAamp DNA Stool Mini Kit (Qia-gen). A swab was vigorously vortexed and incubated at room temperature for 1 min. The sample was transferred to microcentrifuge tubes containing 560𝜇L of Buffer ASL, then vortexed, and incubated at 37∘C for 30 min. In addition, the suspension was incubated at 95∘C for 15 min, vortexed, and centrifuged at 14,000 rpm for 1 min to obtain pelletized stool particles. Extraction was performed following the protocol of the QIAamp DNA Stool Mini Kit. DNA was eluted with 50𝜇L Buffer AE, centrifuged at 14,000 rpm for 1 min, and then the DNA extract was stored at−20∘C until being further analyzed.
5.2. Library Construction and Sequencing of the V4 Region of 16S rDNA. The PCR primers, F515 (5 -GTGCCAGCMGCC-GCGGTAA-3) and R806 (5 -GGACTACHVGGGTWTCT-AAT-3), were designed to amplify the V4 region of bacterial 16S rDNA as described previously [66]. Polymerase chain reaction (PCR) amplification was performed in a 50𝜇L reac-tion volume containing 25𝜇L 2x Taq Master Mix (Thermo Scientific), 0.2𝜇M of each forward and reverse primer, and 20 ng of a DNA template. The reaction conditions included an initial temperature of 95∘C for 5 min, followed by 30 cycles of 95∘C for 30 s, 54∘C for 1 min, and 72∘C for 1 min, with a final extension of 72∘C for 5 min. Next, amplified products were checked by 2% agarose gel electrophoresis
and ethidium bromide staining. Amplicons were purified using the AMPure XP PCR Purification Kit (Agencourt) and quantified using the Qubit dsDNA HS Assay Kit (Qubit) on a Qubit 2.0 Fluorometer (Qubit), all according to the respective manufacturer’s instructions. For V4 library prepa-ration, Illumina adapters were attached to the amplicons using the Illumina TruSeq DNA Sample Preparation v2 Kit. Purified libraries were processed for cluster generation and sequencing using the MiSeq system.
5.3. Filtering 16S rRNA (rDNA) Sequencing Data for Quality.
The FASTX-Toolkit (http://hannonlab.cshl.edu/fastx toolkit) was used to process the raw fastq read data files from Illumina Miseq. The sequence quality criteria were as follows: (1) the minimum acceptable phred quality score of sequences was 30 with a score of>70% of sequence bases of ≥20; (2) after quality trimming from the sequence tail, sequences of>100 bp were retained, and they also had an acceptable phred quality score of 30; and (3) both forward and reverse sequencing reads which met the first and second requirements were retained for subsequent analysis. Sequencing reads from different samples were identified and separated according to specific barcodes in the 5end of the sequence (with two mismatches allowed).
5.4. Taxonomic Assignments of Bacterial 16S rRNA Sequences.
Paired-end sequences were obtained, and their qualities were assessed using the FASTX-Toolkit. To generate taxonomic assignments, Bowtie2 was used to align sequencing reads against the collection of a 16S rRNA sequences database. A standard of 97% similarity against the database was applied. 16S rRNA sequences of bacteria were retrieved from the SILVA ribosomal RNA sequence database [67]. Following sequence data collection, sequences were extracted using V4 forward and reverse primers. To prevent repetitive sequence assignments, V4 sequences from SILVA were then clustered into several clusters by 97% similarity using UCLUST [68]. Results of the taxonomic assignment were filtered to retain assignments with>10 sequences.
5.5. Bacterial Community Analysis. After taxonomic
assign-ment, an operational taxonomic unit (OTU) table was generated. To normalize the sample size of all samples, a rarefaction process was performed on the OTU table. Alpha and beta diversities were calculated based on a rarified OTU table. The Kolmogorov-Smirnov test and an analysis of variance (ANOVA) test with the Bonferroni correction were used to investigate significant differences between different sample groups. To observe relationships between samples and explore taxonomic associations, weighted and unweighted UniFrac [69] distance metrics were also generated based on the rarified OTU table. A principal coordinate analysis (PCoA) and unsupervised clustering were performed based on the UniFac distance matrix. To explore relationships between clinical features and different sample groups, Spear-man’s correlation coefficient and regression analysis were performed. The statistical analytical process was done in R language. The J48 machine learning method in Weka
3.6.7 [70] was used to construct a classification rule for discriminating between obese and normal individuals.
Conflict of Interests
The authors declare that there is no conflict of interests regarding the publication of this paper.
Authors’ Contribution
Chih-Min Chiu, Wei-Chih Huang, and Shun-Long Weng contributed equally to this work.
Acknowledgments
The authors would like to thank the National Science Council of the Republic of China for financially supporting this research under Grant nos. NSC 101-2311-B-009-005-MY3 and NSC 102-2627-B-009-001. This work was supported in part by the Ministry of Science and Technology under Grant no. MOST 103-2221-E-038-013-MY2. This work was supported in part by UST-UCSD International Center of Excellence in Advanced Bioengineering sponsored by the Ministry of Science and Technology I-RiCE Program under Grant no. NSC-102-2911-I-009-101-, and Veterans General Hospitals and University System of Taiwan (VGHUST) Joint Research Program under Grant no. VGHUST103-G5-1-2. This work was also partially supported by MOE ATU.
References
[1] J. R. Goldsmith and R. B. Sartor, “The role of diet on intestinal microbiota metabolism: downstream impacts on host immune function and health, and therapeutic implications,” Journal of Gastroenterology, vol. 49, no. 5, pp. 785–798, 2014.
[2] J. Chen, X. He, and J. Huang, “Diet effects in gut microbiome and obesity,” Journal of Food Science, vol. 79, no. 4, pp. R442– R451, 2014.
[3] O. O. Erejuwa, S. A. Sulaiman, and M. S. Wahab, “Modulation of gut microbiota in the management of metabolic disorders: the prospects and challenges,” International Journal of Molecular Sciences, vol. 15, no. 3, pp. 4158–4188, 2014.
[4] G. L. Hold, M. Smith, C. Grange, E. R. Watt, E. M. El-Omar, and I. Mukhopadhya, “Role of the gut microbiota in inflammatory bowel disease pathogenesis: what have we learnt in the past 10 years?” World Journal of Gastroenterology, vol. 20, no. 5, pp. 1192–1210, 2014.
[5] M. Arumugam, J. Raes, E. Pelletier et al., “Enterotypes of the human gut microbiome,” Nature, vol. 473, no. 7346, pp. 174–180, 2011.
[6] G. D. Wu, J. Chen, C. Hoffmann et al., “Linking long-term dietary patterns with gut microbial enterotypes,” Science, vol. 334, no. 6052, pp. 105–108, 2011.
[7] J. Peterson, S. Garges, M. Giovanni et al., “The NIH human microbiome project,” Genome Research, vol. 19, no. 12, pp. 2317– 2323, 2009.
[8] J. Qin, Y. Li, Z. Cai et al., “A metagenome-wide association study of gut microbiota in type 2 diabetes,” Nature, vol. 490, no. 7418, pp. 55–60, 2012.
[9] F. H. Karlsson, V. Tremaroli, I. Nookaew et al., “Gut metage-nome in European women with normal, impaired and diabetic glucose control,” Nature, vol. 498, no. 7452, pp. 99–103, 2013. [10] X. Zhang, D. Shen, Z. Fang et al., “Human gut microbiota
changes reveal the progression of glucose intolerance,” PLoS ONE, vol. 8, no. 8, Article ID e71108, 2013.
[11] F. Karlsson, V. Tremaroli, J. Nielsen, and F. Backhed, “Assessing the human gut microbiota in metabolic diseases,” Diabetes, vol. 62, no. 10, pp. 3341–3349, 2013.
[12] A. Lyra, T. Rinttil¨a, J. Nikkil¨a et al., “Diarrhoea-predominant irritable bowel syndrome distinguishable by 16S rRNA gene phylotype quantification,” World Journal of Gastroenterology, vol. 15, no. 47, pp. 5936–5945, 2009.
[13] S. O. Noor, K. Ridgway, L. Scovell et al., “Ulcerative colitis and irritable bowel patients exhibit distinct abnormalities of the gut microbiota,” BMC Gastroenterology, vol. 10, article 134, 2010. [14] M. Rajili´c-Stojanovi´c, E. Biagi, H. G. H. J. Heilig et al., “Global
and deep molecular analysis of microbiota signatures in fecal samples from patients with irritable bowel syndrome,” Gas-troenterology, vol. 141, no. 5, pp. 1792–1801, 2011.
[15] Y. Zhu, T. Michelle Luo, C. Jobin, and H. A. Young, “Gut microbiota and probiotics in colon tumorigenesis,” Cancer Letters, vol. 309, no. 2, pp. 119–127, 2011.
[16] M. A. Hullar, A. N. Burnett-Hartman, and J. W. Lampe, “Gut microbes, diet, and cancer,” Cancer Treatment and Research, vol. 159, pp. 377–399, 2014.
[17] H. R. Hagland and K. Soreide, “Cellular metabolism in colorec-tal carcinogenesis: influence of lifestyle, gut microbiome and metabolic pathways,” Cancer Letters, 2014.
[18] K. Korpela, H. J. Flint, A. M. Johnstone et al., “Gut microbiota signatures predict host and microbiota responses to dietary interventions in obese individuals,” PLoS One, vol. 9, no. 3, Article ID e90702, 2014.
[19] B. A. Swinburn, G. Sacks, K. D. Hall et al., “The global obesity pandemic: shaped by global drivers and local environments,” The Lancet, vol. 378, no. 9793, pp. 804–814, 2011.
[20] K. C. Portero McLellan, K. Wyne, E. T. Villagomez, and W. A. Hsueh, “Therapeutic interventions to reduce the risk of progression from prediabetes to type 2 diabetes mellitus,” Therapeutics and Clinical Risk Management, vol. 10, pp. 173–188, 2014.
[21] A. Everard and P. D. Cani, “Diabetes, obesity and gut micro-biota,” Best Practice and Research: Clinical Gastroenterology, vol. 27, no. 1, pp. 73–83, 2013.
[22] J. A. Bell, M. Kivimaki, and M. Hamer, “Metabolically healthy obesity and risk of incident type 2 diabetes: a meta-analysis of prospective cohort studies,” Obesity Reviews, vol. 15, no. 6, pp. 504–515, 2014.
[23] D. Nagakubo, M. Shirai, Y. Nakamura et al., “Prophylactic effects of the glucagon-like Peptide-1 analog liraglutide on hyperglycemia in a rat model of type 2 diabetes mellitus associated with chronic pancreatitis and obesity,” Comparative Medicine, vol. 64, no. 2, pp. 121–127, 2014.
[24] M. Kratz, T. Baars, and S. Guyenet, “The relationship between high-fat dairy consumption and obesity, cardiovascular, and metabolic disease,” European Journal of Nutrition, vol. 52, no. 1, pp. 1–24, 2013.
[25] G. M. Hinnouho, S. Czernichow, A. Dugravot et al., “Metaboli-cally healthy obesity and the risk of cardiovascular disease and type 2 diabetes: The Whitehall II Cohort Study,” European Heart Journal, 2014.
[26] K. A. Britton, J. M. Massaro, J. M. Murabito, B. E. Kreger, U. Hoffmann, and C. S. Fox, “Body fat distribution, incident cardiovascular disease, cancer, and all-cause mortality,” Journal of the American College of Cardiology, vol. 62, no. 10, pp. 921– 925, 2013.
[27] E. E. Frezza, M. S. Wachtel, and M. Chiriva-Internati, “Influence of obesity on the risk of developing colon cancer,” Gut, vol. 55, no. 2, pp. 285–291, 2006.
[28] K. Nakamura, A. Hongo, J. Kodama, and Y. Hiramatsu, “Fat accumulation in adipose tissues as a risk factor for the devel-opment of endometrial cancer,” Oncology Reports, vol. 26, no. 1, pp. 65–71, 2011.
[29] M. Remely, E. Aumueller, D. Jahn, B. Hippe, H. Brath, and A. G. Haslberger, “Microbiota and epigenetic regulation of inflam-matory mediators in type 2 diabetes and obesity,” Beneficial Microbes, vol. 5, no. 1, pp. 33–43, 2014.
[30] R. E. Ley, P. J. Turnbaugh, S. Klein, and J. I. Gordon, “Microbial ecology: human gut microbes associated with obesity,” Nature, vol. 444, no. 7122, pp. 1022–1023, 2006.
[31] J. Shen, M. S. Obin, and L. Zhao, “The gut microbiota, obesity and insulin resistance,” Molecular Aspects of Medicine, vol. 34, no. 1, pp. 39–58, 2013.
[32] M. Nieuwdorp, P. W. Gilijamse, N. Pai, and L. M. Kaplan, “Role of the microbiome in energy regulation and metabolism,” Gastroenterology, vol. 146, no. 6, pp. 1525–1533, 2014.
[33] P. J. Turnbaugh, F. B¨ackhed, L. Fulton, and J. I. Gordon, “Diet-induced obesity is linked to marked but reversible alterations in the mouse distal gut microbiome,” Cell Host and Microbe, vol. 3, no. 4, pp. 213–223, 2008.
[34] F. B¨ackhed, H. Ding, T. Wang et al., “The gut microbiota as an environmental factor that regulates fat storage,” Proceedings of the National Academy of Sciences of the United States of America, vol. 101, no. 44, pp. 15718–15723, 2004.
[35] P. J. Turnbaugh, M. Hamady, T. Yatsunenko et al., “A core gut microbiome in obese and lean twins,” Nature, vol. 457, no. 7228, pp. 480–484, 2009.
[36] J. Furet, L. Kong, J. Tap et al., “Differential adaptation of human gut microbiota to bariatric surgery-induced weight loss: links with metabolic and low-grade inflammation markers,” Diabetes, vol. 59, no. 12, pp. 3049–3057, 2010.
[37] H. Zhang, J. K. DiBaise, A. Zuccolo et al., “Human gut microbiota in obesity and after gastric bypass,” Proceedings of the National Academy of Sciences of the United States of America, vol. 106, no. 7, pp. 2365–2370, 2009.
[38] Y. N. Yin, Q. F. Yu, N. Fu, X. W. Liu, and F. G. Lu, “Effects of four Bifidobacteria on obesity in high-fat diet induced rats,” World Journal of Gastroenterology, vol. 16, no. 27, pp. 3394–3401, 2010. [39] R. E. Ley, “Obesity and the human microbiome,” Current
Opinion in Gastroenterology, vol. 26, no. 1, pp. 5–11, 2010. [40] The Human Microbiome Project Consortium, “Structure,
func-tion and diversity of the healthy human microbiome,” Nature, vol. 486, no. 7402, pp. 207–214, 2012.
[41] W. Pan, K. M. Flegal, H. Chang, W. Yeh, C. Yeh, and W. Lee, “Body mass index and obesity-related metabolic disorders in Taiwanese and US whites and blacks: Implications for definitions of overweight and obesity for Asians,” The American Journal of Clinical Nutrition, vol. 79, no. 1, pp. 31–39, 2004. [42] A. Santacruz, M. C. Collado, L. Garc´ıa-Vald´es et al., “Gut
microbiota composition is associated with body weight, weight gain and biochemical parameters in pregnant women,” British Journal of Nutrition, vol. 104, no. 1, pp. 83–92, 2010.
[43] S. Xiao, N. Fei, X. Pang et al., “A gut microbiota-targeted dietary intervention for amelioration of chronic inflammation underlying metabolic syndrome,” FEMS Microbiology Ecology, vol. 87, no. 2, pp. 357–367, 2014.
[44] L. Rigsbee, R. Agans, V. Shankar et al., “Quantitative profiling of gut microbiota of children with diarrhea-predominant irritable bowel syndrome,” The American Journal of Gastroenterology, vol. 107, no. 11, pp. 1740–1751, 2012.
[45] N. Wu, X. Yang, R. Zhang et al., “Dysbiosis signature of fecal microbiota in colorectal cancer patients,” Microbial Ecology, vol. 66, no. 2, pp. 462–470, 2013.
[46] W. Chen, F. Liu, Z. Ling, X. Tong, and C. Xiang, “Human intestinal lumen and mucosa-associated microbiota in patients with colorectal cancer,” PLoS ONE, vol. 7, no. 6, Article ID e39743, 2012.
[47] Q. Zhu, Z. Jin, W. Wu et al., “Analysis of the intestinal lumen microbiota in an animal model of colorectal cancer,” PLoS ONE, vol. 9, no. 3, Article ID e90849, 2014.
[48] D. Wang, D. Yan, W. Hou, X. Zeng, Y. Qi, and J. Chen, “Charac-terization of bla𝑂𝑥𝐴−23gene regions in isolates of Acinetobacter baumannii,” Journal of Microbiology, Immunology and Infection, 2014.
[49] H. Urbanczyk, J. C. Ast, M. J. Higgins, J. Carson, and P. V. Dunlap, “Reclassification of Vibrio fischeri, Vibrio logei, Vibrio salmonicida and Vibrio wodanis as Aliivibrio fischeri gen. nov., comb. nov., Aliivibrio logei comb. nov., Aliivibrio salmonicida comb. nov. and Aliivibrio wodanis comb. nov,” International Journal of Systematic and Evolutionary Microbiology, vol. 57, part 12, pp. 2823–2829, 2007.
[50] J. C. Ast, H. Urbanczyk, and P. V. Dunlap, “Multi-gene analysis reveals previously unrecognized phylogenetic diversity in Ali-ivibrio,” Systematic and Applied Microbiology, vol. 32, no. 6, pp. 379–386, 2009.
[51] R. Beaz-Hidalgo, A. Doce, S. Balboa, J. L. Barja, and J. L. Romalde, “Aliivibrio finisterrensis sp. nov., isolated from Manila clam, Ruditapes philippinarum and emended description of the genus Aliivibrio,” International Journal of Systematic and Evolutionary Microbiology, vol. 60, no. 1, pp. 223–228, 2010. [52] S. Yoshizawa, H. Karatani, M. Wada, A. Yokota, and K. Kogure,
“Aliivibrio sifiae sp. nov., luminous marine bacteria isolated from seawater,” Journal of General and Applied Microbiology, vol. 56, no. 6, pp. 509–518, 2010.
[53] Y. S. Chen, Y. C. Liu, M. Y. Yen, J. H. Wang, S. R. Wann, and D. L. Cheng, “Skin and soft-tissue manifestations of Shewanella putrefaciens infection,” Clinical Infectious Diseases, vol. 25, no. 2, pp. 225–229, 1997.
[54] N. Vignier, M. Barreau, C. Olive et al., “Human infection with Shewanella putrefaciens and S. algae: report of 16 cases in Martinique and review of the literature,” American Journal of Tropical Medicine and Hygiene, vol. 89, no. 1, pp. 151–156, 2013. [55] R. F. Boente, L. Q. Ferreira, L. S. Falc˜ao et al., “Detection of
resistance genes and susceptibility patterns in Bacteroides and Parabacteroides strains,” Anaerobe, vol. 16, no. 3, pp. 190–194, 2010.
[56] M. Sakamoto and Y. Benno, “Reclassification of Bacteroides distasonis, Bacteroides goldsteinii and Bacteroides merdae as Parabacteroides distasonis gen. nov., comb. nov., Parabac-teroides goldsteinii comb. nov and ParabacParabac-teroides merdae comb. nov,” International Journal of Systematic and Evolutionary Microbiology, vol. 56, part 7, pp. 1599–1605, 2006.
[57] J. Xu, M. A. Mahowald, R. E. Ley et al., “Evolution of symbiotic bacteria in the distal human intestine.,” PLoS Biology, vol. 5, no. 7, Article ID e156, 2007.
[58] J. M. Clarke, D. L. Topping, C. T. Christophersen et al., “Butyrate esterified to starch is released in the human gastrointestinal tract,” The American Journal of Clinical Nutrition, vol. 94, no. 5, pp. 1276–1283, 2011.
[59] E. S´anchez, E. Donat, C. Ribes-Koninckx, M. Calabuig, and Y. Sanz, “Intestinal Bacteroides species associated with coeliac disease,” Journal of Clinical Pathology, vol. 63, no. 12, pp. 1105– 1111, 2010.
[60] N. Becker, J. Kunath, G. Loh, and M. Blaut, “Human intestinal microbiota: characterization of a simplified and stable gnotobi-otic rat model,” Gut Microbes, vol. 2, no. 1, 2011.
[61] N. Fei and L. Zhao, “An opportunistic pathogen isolated from the gut of an obese human causes obesity in germfree mice,” ISME Journal, vol. 7, no. 4, pp. 880–884, 2013.
[62] A. Hejazi and F. R. Falkiner, “Serratia marcescens,” Journal of Medical Microbiology, vol. 46, no. 11, pp. 903–912, 1997. [63] M. Patankar, S. Sukumaran, A. Chhibba, U. Nayak, and
L. Sequeira, “Comparative in-vitro activity of cefoperazone-tazobactam and cefoperazone-sulbactam combinations against ESBL pathogens in respiratory and urinary infections,” Journal of Association of Physicians of India, vol. 60, no. 11, pp. 22–24, 2012.
[64] E. M. Carlisle, V. Poroyko, M. S. Caplan, J. Alverdy, M. J. Morowitz, and D. Liu, “Murine gut microbiota and transcrip-tome are diet dependent,” Annals of Surgery, vol. 257, no. 2, pp. 287–294, 2013.
[65] C. de Weerth, S. Fuentes, P. Puylaert, and W. M. de vos, “Intestinal microbiota of infants with colic: development and specific signatures,” Pediatrics, vol. 131, no. 2, pp. e550–e558, 2013.
[66] J. G. Caporaso, C. L. Lauber, W. A. Walters et al., “Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample,” Proceedings of the National Academy of Sciences of the United States of America, vol. 108, no. 1, pp. 4516– 4522, 2011.
[67] E. Pruesse, C. Quast, K. Knittel et al., “SILVA: a comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB,” Nucleic Acids Research, vol. 35, no. 21, pp. 7188–7196, 2007.
[68] R. C. Edgar, “Search and clustering orders of magnitude faster than BLAST,” Bioinformatics, vol. 26, no. 19, pp. 2460–2461, 2010.
[69] C. Lozupone, M. E. Lladser, D. Knights, J. Stombaugh, and R. Knight, “UniFrac: an effective distance metric for microbial community comparison,” ISME Journal, vol. 5, no. 2, pp. 169– 172, 2011.
[70] M. Hall, E. Frank, G. Holmes, B. Pfahringer, P. Reutemann, and I. H. Witten, “The WEKA data mining software: an update,” SIGKDD Explorations, vol. 11, no. 1, pp. 10–18, 2009.
Submit your manuscripts at
http://www.hindawi.com
Hindawi Publishing Corporation
http://www.hindawi.com Volume 2014 Anatomy
Research International
Peptides
Hindawi Publishing Corporation
http://www.hindawi.com Volume 2014
Hindawi Publishing Corporation http://www.hindawi.com
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporation
http://www.hindawi.com Volume 2014 Molecular Biology International
Genomics
International Journal of
Hindawi Publishing Corporation
http://www.hindawi.com Volume 2014
The Scientific
World Journal
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Hindawi Publishing Corporation
http://www.hindawi.com Volume 2014
Bioinformatics
Advances inMarine Biology
Journal ofHindawi Publishing Corporation
http://www.hindawi.com Volume 2014 Hindawi Publishing Corporation
http://www.hindawi.com Volume 2014
Signal Transduction
Journal of Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014 BioMed
Research International
Evolutionary Biology
International Journal of
Hindawi Publishing Corporation
http://www.hindawi.com Volume 2014
Hindawi Publishing Corporation
http://www.hindawi.com Volume 2014
Biochemistry Research International
Archaea
Hindawi Publishing Corporation
http://www.hindawi.com Volume 2014 Hindawi Publishing Corporation
http://www.hindawi.com Volume 2014 Genetics
Research International
Hindawi Publishing Corporation
http://www.hindawi.com Volume 2014 Advances in
Virology
Hindawi Publishing Corporation http://www.hindawi.com
Nucleic Acids
Journal ofVolume 2014
Stem Cells
International
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Hindawi Publishing Corporation
http://www.hindawi.com Volume 2014
Enzyme
Research
Hindawi Publishing Corporation
http://www.hindawi.com Volume 2014
International Journal of