行政院國家科學委員會專題研究計畫 成果報告
大豆過氧化同功
之生理生化研究
計畫類別: 個別型計畫 計畫編號: NSC91-2311-B-110-014- 執行期間: 91 年 08 月 01 日至 92 年 07 月 31 日 執行單位: 國立中山大學生物科學系(所) 計畫主持人: 劉景煌 計畫參與人員: 劉景煌 報告類型: 精簡報告 處理方式: 本計畫可公開查詢 中 華 民 國 93 年 2 月 11 日銅對大豆根部組織過氧化脢和木質素合成之快速影響
Rapid effect of copper on peroxidase activity and lignin synthesis in soybean roots
Chih-Cheng Lin, Li-Ming Chen, Zin-Huang Liu*
Department of Biological Sciences, National Sun Yat-Sen University, Kaohsiung, Taiwan 804, Republic of China
*Corresponding author. Fax: +88675253609
中文摘要 銅處理明顯抑制大豆根之生長,卻增強過氧化脢之活性。在銅處 理的組織,陽性( pI 8.9, pI 8.3 和 pI 7.7 )過氧化脢和陰性( pI 6.5, pI 5.6 and pI 4.4 ) 過氧化脢活性之增加伴隨木質素之上升。大豆 根尖對銅更敏感,在銅處理 1 小時內 pI4.4 的過氧化脢和 H2O2都明顯上 升,我們認為在銅處理的組織陽性和陰性過氧化脢的增加有部份用來 合成木質素同時並消除組織內有毒的 H2O2。 關鍵字: Glycine max,銅,過氧化氫,木質化作用,過氧化脢 Abstract
Copper-treated soybean ( Glycine max ) seedlings show significant inhibition in soybean root growth, and enhancement in peroxidase (POD) activity. The increase of cationic ( pI 8.9, pI 8.3 and pI 7.7 ) PODs and anionic ( pI 6.5, pI 5.6 and pI 4.4 ) PODs activities is
accompanied by a rise in lignin contents in Cu-treated tissues. Soybean root tips are more sensitive to Cu treatment, and response to Cu within an hour in terms of the increase of pI 4.4 POD activity and H2O2 levels.
We suggest that the increase in cationic ( pI 8.9, pI 8.3 and pI 7.7 ) and anionic ( pI 6.5, pI 5.6 and pI 4.4 ) PODs induced by Cu is in part responsible for lignin synthesis in soybean roots, and may remove excess hydrogen peroxide caused by Cu.
Keyword:Glycine max, copper, hydrogen peroxide, lignification,
peroxidase
Abbreviations: Cu, copper; H2O2, hydrogen peroxide; IAA,
Introduction
It has been known for a long time about the toxicity of excessive heavy metal. When plants are exposed to this environmental stress, it will cause growth inhibition, ion leakage of cell membrane, destruction of pigment, and even more the death of plants [1, 9, 25, 30]. The most common impact of the heavy metal is the accumulation of large
amounts of peroxide compounds rapidly attacking several kinds of biomolecules and disrupting the metabolism of plant cell [6]. Copper is an essential micronutrient for plants associated with enzymes involved in redox reactions. According to the reports, the excess copper increases the activity of peroxidase in tomato seeding [23]. Recently some
commercial fungicides are added with copper. Copper, in low concentrations, may not cause significant damage to the cell membranes, but may bring about the accumulation of peroxide compounds. This increase in peroxide compounds is known about oxidative burst [1, 9, 28, 33], which increases the levels of hydrogen peroxide (H2O2), and activates the host defense mechanisms to
pathogens. Moreover, the peroxide compound, H2O2, is reported to be
one part of the signal transduction cascade of the host defense mechanisms [5].
Peroxidases ( EC. 1.11.1.7 ) ( POD ) are widely distributed isozymes, and play an important role in physiological responses, including lignification, auxin metabolism, disease resistance, and growth regulation [18, 22]. According to the isoelectric point, the POD can be divided into cationic and anionic types. Cationic PODs are presumed to catalyze the degradation of indole-3-acetic acid ( IAA ) in
the presence of oxygen [10]. Anionic PODs are considered to catalyze the biosynthesis of lignin [15]. Transgenic tobacco with increasing anionic POD activity is associated with increasing resistance to
pathogens due to the lignification of the infected sites by anionic POD [17]. Meanwhile, some reports suggest that cationic POD may be involved in the lignin synthesis [27].
Lignin is the second abundant organic substance in vascular plant cell walls. Lignin is a highly branched polymer of phenylpropanoid groups, generally formed from three distinct phenylpropanoid alcohols including coniferyl, coumaryl, and sinapyl alcohols. The
polymerization of these precursors of lignin is catalyzed by POD in the present of H2O2 [22]. The accurate structure of lignin is still indistinct.
However, it is believed that lignin will covalently bound to cellulous, others polysaccharides, and proteins of the cell wall [12]. The complex structure of lignin provide the mechanical support, water transmission, as well as blocking the growth of pathogen and infection.
In this experiment, we investigated the effect of copper on the growth of soybean roots by examining the changes in H2O2 level, POD
activity, and lignin content during the period of copper treatment.
Materials and Methods
Plant material
Soybean ( Glycine max ) were germinated and grown in vermiculite at 27 ± 0.5℃ under 16 h / 8 h light/dark period and
m-2 s-1. After five days germination, soybean seedings were cultured in various concentrations of CuSO4 ( 1 – 10 µM ) for various times
( 0.5–72 h ). The fresh weight of root was measured at the end of culture. Roots were collected and stored in the freezer ( -80℃) for a
few days before H2O2, POD, and lignin assay.
Detection of H2O2
POD extraction and activity assay 2. Results
Isolelectric focusing gel electrophoresis and POD activity staining
RNA isolation and reverse transcription of RNA
Polymerase Chain Reaction ( PCR ) and quantification of PCR products
Lignin determination
In vivo detection of H2O2 in soybean roots
Results
Effect of Cu on root growth, H2O2 level, POD activity and lignin
content in soybean seedlings
Five-day-old soybean seedlings were cultured in various
concentrations of CuSO4 solutions ranging from 1 µM to 10 µM. As
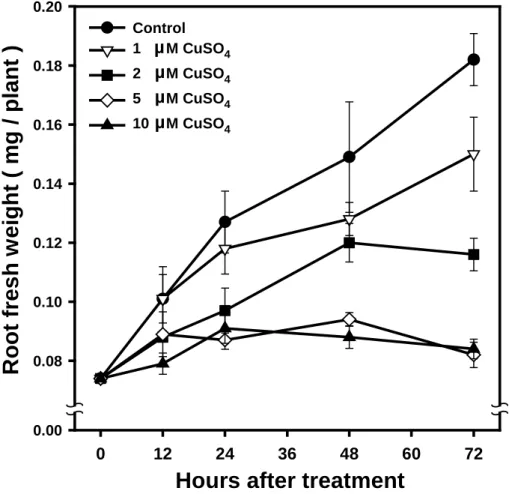
shown in Fig. 1, Cu remarkably inhibited the growth of soybean roots. As the incubation time was prolonged to 72 h, the reductions were
about 18 %, 36 % and 55 % respectively, in 1, 2 and 5 µM of CuSO4
compared with the control soybean roots. When the concentration of Cu increased to 10 µM, there is no further inhibition occurred in soybean roots.
Detection of the H2O2 level was following the methods by Jana
and Choudhuri [13]. The soybean seedlings were cultured in 2 µM, 5 µM and 10 µM CuSO4 solutions for 1 to 24 h. The root tissues were
assayed spectrophotometrically by measuring the increase in
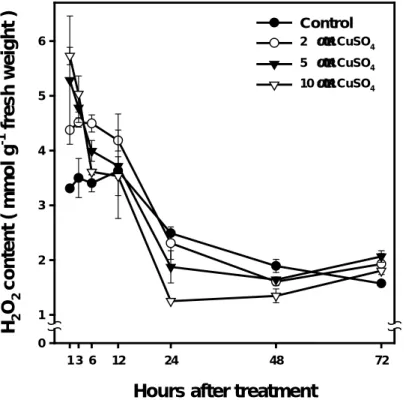
absorbance at 410 nm. As shown in Fig. 2, during the course of Cu treatment, the level of H2O2 rapidly increased 1 h after Cu treatment,
and decreased as the incubation time was prolonged to 12 h. The levels of H2O2 decreased in the 5 and 10 µM Cu-treated tissues, as the
incubation was prolonged to 24 h or 48 h. The amount of H2O2 in
Cu-treated tissue ( 10 µM ) was one – third compared with the control 24 h after incubation. The levels of H2O2 in control roots cultured in
water did not change much during 72 h of incubation. The localization of H2O2 in soybean roots was stained as a dark brown color described
by Thordal-Christensen et al. [31]. H2O2 appeared 1 h after Cu
treatment, and was mainly localized in the root tip (Fig. 3). As the incubation time was prolonged to 24 h, the brown colour became darker and spread evenly in the whole roots.
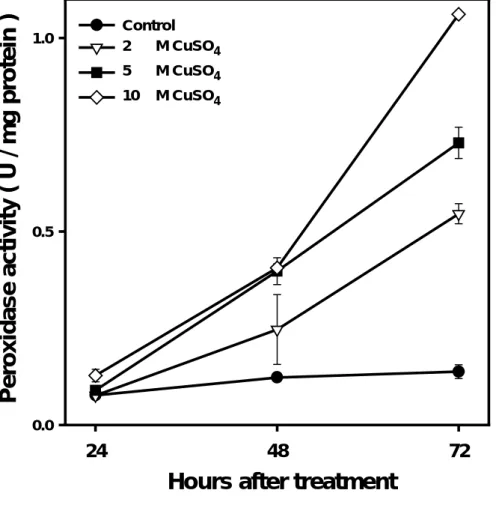
The POD activity was significantly enhanced by Cu treatment ( Fig. 4 ). In Cu treated tissues ( 10 µM ), the activity of POD was about 10-fold increase compared with the controls. As the incubation time was prolonged to 72 h, the activity of POD in the control did not
of POD activity by Cu was concentration dependent.
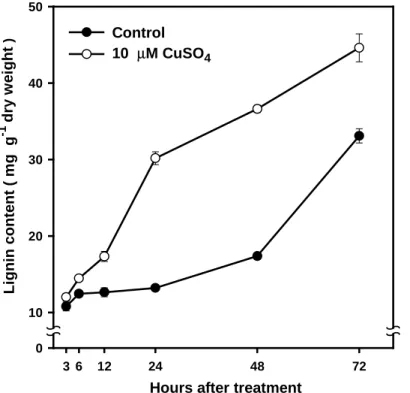
The lignin content in Cu-treated tissues ( 10 µM ) was about 3-fold higher than that of the controls after 24 h of incubation in CuSO4
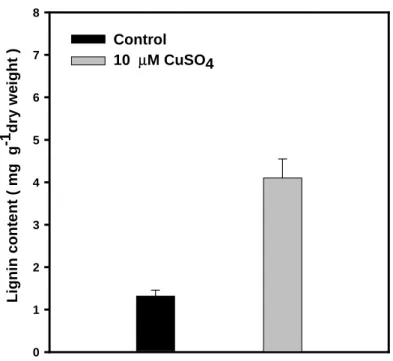
solution. As the incubation time was prolonged to 72 h, the difference in lignin content between Cu-treated tissues and the control become less ( Fig. 5 ). In addition, the lignin content in Cu-treated root tip (1 cm) was about 2.5-fold higher than that of the controls after 3 h of
incubation in CuSO4 solution (Fig. 6).
Change of cationic and anionic POD activity and POD gene transcripts in soybean roots during Cu treatment
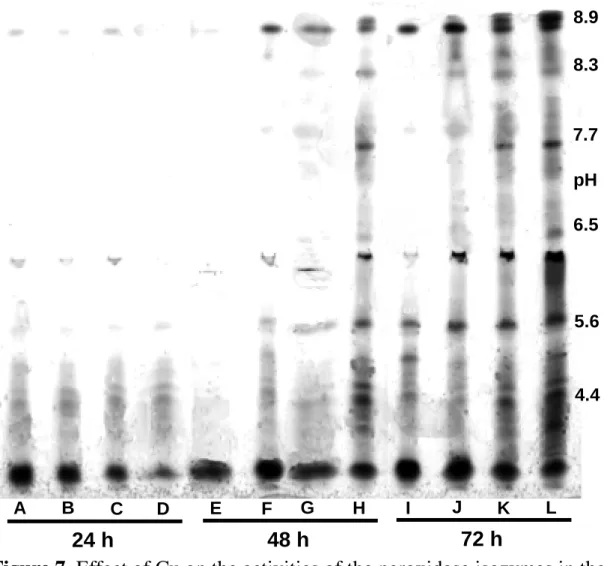
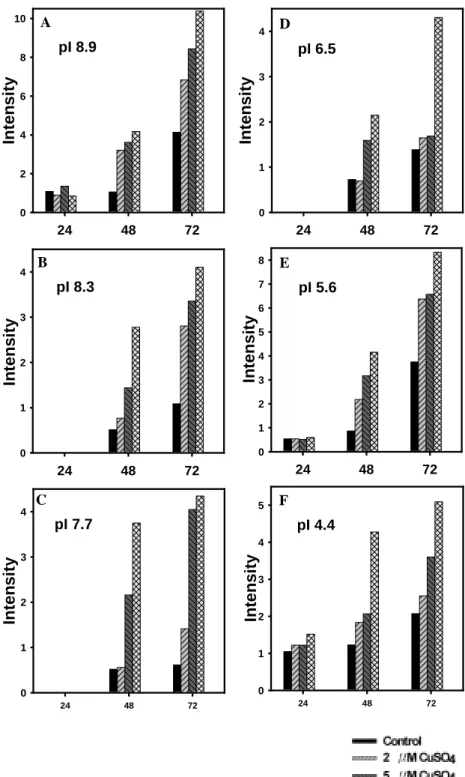
Time-course studies were designed to investigate the POD activity of Cu-treated and the control tissues. Soybean roots were homogenized, extracted and subjected to polyacrylamide gels for isoelectric focusing electrophoresis. As shown in Fig. 7, there were more than six isozymes of POD ( pI 8.9, pI 8.3, pI 7.7, pI 6.5, pI 5.6 and pI 4.4 ), identified by isoelectric point in the extracts of Cu-treated and the control roots. The activities of cationic ( pI 8.9, pI 8.3 and pI 7.7 ) and anionic ( pI 6.5, pI 5.6 and pI 4.4 ) POD were significantly enhanced by Cu treatments. The activity of pI 8.9 POD was enhanced in Cu-treated tissues 48 h after incubation ( Figs. 7 and 8A). As the incubation time was
prolonged to 72 h, POD activity was also enhanced to some extents. The activity of pI 8.9 POD in Cu-treated tissues was about 2 to 3-fold compared with the controls. The enhancement of pI 8.9 POD is
concentration dependent . The enhancement of the activity of pI 8.3 and pI 7.7 PODs in Cu-treated tissues showed similar pattern as that of pI 8.9 POD ( Figs. 7, 8B and 8C ) . The activity of pI 6.5 POD remarkably
increased in Cu-treated tissues 48 h after incubation ( Figs. 7 and 8D ). As the incubation time was prolonged to 72 h, POD activity was
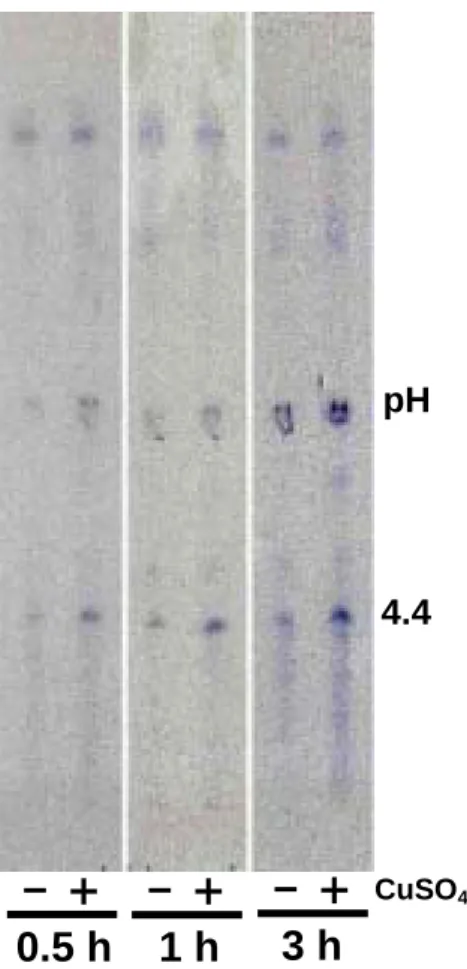
enhanced to great extents. The activity of pI 6.5 POD in Cu-treated tissues was about 3 to 4-fold compared with the control. Cu had similar effect on the other two anionic POD ( pI 5.6 and pI 4.4 ) (Figs. 7, 8E and 8F ). In Cu-treated root tip, the enhancement of the pI 4.4 POD activity occurred 0.5 h after Cu treatment (Fig. 9).
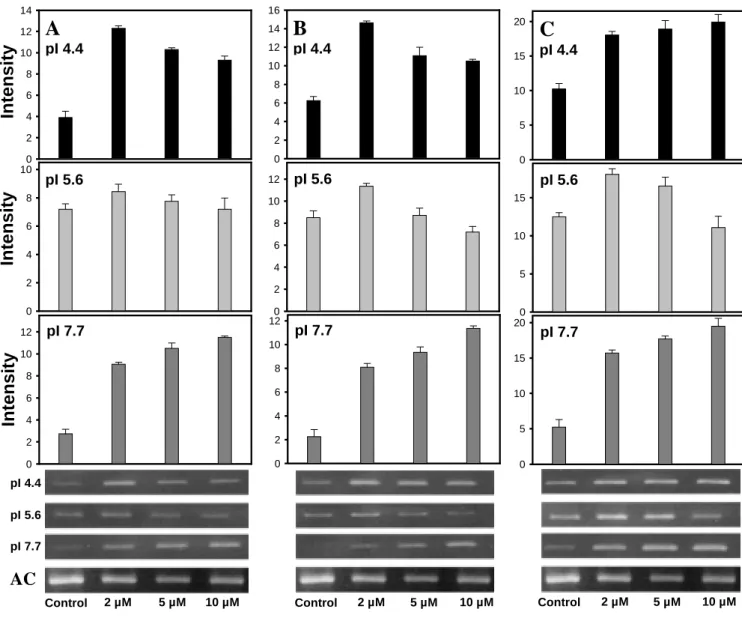
The transcript levels of gene encoding cationic pI 7.7 POD and anionic pI 5.6 and pI 4.4 PODs in soybean roots were examined by reverse transcriptase-polymerase chain reaction ( RT-PCR ). A linear calibration in the template and the PCR product of pI 7.7, pI 5.6, pI 4.4 and ACT were performed. In Cu-treated roots, the transcript level of cationic pI 7.7 POD was significantly enhanced 6 h after incubation ( Fig. 10A). As the incubation time was prolonged to 24 h, the
transcript levels of pI 7.7 POD did not change much. The enhancement of the transcript level of pI 7.7 POD is concentration dependent. The transcript level of anionic pI 4.4 POD in Cu-treated roots was also enhanced by Cu, the concentration of the transcript level was about 2 to 3-fold compared with the control. The increase of the transcript level of pI 4.4 POD in Cu-treated tissues is not concentration dependent. Low concentration of Cu(2 µM)is optimal. The effect of Cu on the
transcription level of pI 5.6 POD is less significant than that of pI 7.7 and pI 4.4 PODs. Higher concentration of Cu(5, 10 µM)seems to
Discussion
Cu-treated soybean seedling shows significant inhibition in root growth ( Fig. 1 ). As far as the relationship between root growth and POD activity is concerned. The reduction of soybean roots is correlated with the induction of PODs ( Fig. 4 ). Li et al. [19] reported that Cu suppresses the growth of peanut roots with the increase of the cationic and anionic PODs. In this experiment, we found that the POD activity was significantly enhanced by Cu ( Fig. 4 ). As reported, Cu is an efficient catalyst in the formation of several reactive oxygen species and free radical [33]. Cu at toxic concentration inhibits photosynthetic electron transport, promoting the formation of hydrogen peroxide [29]. The H2O2 level in Cu-treated tissues shows rapid increase 1 h after
treatment ( Figs. 2 and 3 ). A diminished level of H2O2 was observed in
the roots of Cu-treated ( 5 µM and 10 µM ) soybean seedling 12 h after incubation compared with the controls. The increase in POD activity induced by Cu might remove excess hydrogen peroxide caused by Cu, thus serving a detoxifying role during Cu-treatment [33].
Among the POD activity, anionic PODs are most likely involved in lignification [15, 22]. In our study, the isozyme pattern of PODs in isoelectric focusing gels showed an increase of the activities of cationic ( pI 8.9, pI 8.3 and pI 7.7 ) PODs and anionic ( pI 6.5, pI 5.6 and pI 4.4 ) PODs. The increase of cationic ( pI 8.9, pI 8.3 and pI 7.7 ) PODs and anionic ( pI 6.5, pI 5.6 and pI 4.4 ) PODs activities is accompanied by a rise of lignin contents in Cu-treated tissues ( Figs. 4 and 5 ). Quiroga et
al. [27] indicated that both of cationic and anionic PODs were involved
shown that POD present in tissues undergoing lignification display in
vitro activity toward syringaldazine [7]. The cationic ( pI 8.9, pI 8.3 and
pI 7.7 ) and anionic ( pI 6.5, pI 5.6 and pI 4.4 ) PODs also increase in the control tissues, causing more lignified tissues in the control. In this study, we suggest that the increase in cationic ( pI 8.9, pI 8.3 and pI 7.7 ) and anionic ( pI 6.5, pI 5.6 and pI 4.4 ) PODs induced by Cu is in part responsible for lignin synthesis in soybean roots [14, 26]. In soybean roots, the part of root tip is more srnsitive to Cu. The response of root tip to Cu in terms of the increase of pI 4.4 POD activity (Fig. 9) and H2O2 levels (Fig. 3) occurs within an hour. Soybean root tips rapidly
response to Cu stress, and remove excess H2O2 by the action of
peroxidase through lignification processes in plant cell wall. In addition to lignification, PODs probably also catalysed the linkages between phenolic acids and cell-wall polymers [8]. Furthermore, the transcript level of the genes encoding cationic pI 7.7 POD and anionic pI 4.4 POD were about 2 to 3-fold higher than that of control plant ( Fig. 10 ). But the effect of Cu on the transcript level of pI 5.6 POD is less significant than that of pI 7.7 and pI 4.4 PODs. The increased activity in cationic ( pI 7.7 ) and anionic ( pI 4.4 ) POD correlated with the increase in cationic and anionic POD transcripts ( Figs. 7 and 10), and therefore is mostly due to the de novo synthesis of the cationic ( pI 7.7 ) and anionic ( pI 4.4 ) PODs in Cu-treated soybean roots.
References
[1] S. Baccouch, A. Chaoui, E.E. Ferjani, Nickel-induced oxidative damage and antioxidant responses in Zea mays shoots. Plant Physiol. Biochem. 36 (1998 ) 689-694.
[2] R. Beffa, H.V. Martin, P.E. Pilet, In vitro oxidation of indoleacetic acid by soluble auxin-oxidase and peroxidases from maize root. Plant Physiol. 94 ( 1990 ) 485-491.
[3] D.J. Bradely, P. Kjellbom, C.J. Lamb, Elicitor- and wound- induced oxidative cross-linking of a proline-rich plant cell wall protein : a novel, radid defense response. Cell 70 ( 1992 ) 21-30.
[4] R. Bruce, C.A. West, Elicitation of lignin biosynthesis and isoperoxidase activity by pectic fragments in suspension cultures of castor bean. Plant Physiol. 91 ( 1989 ) 889-897
[5] A.C.Cazalé, M.A. Rouet-Mayer, H. Barbier-Brygoo, Y. Mathieu, and L. Christiane, Oxidative burst and hypoosmotic stress in tobacco cell suspensions. Plant Physiol. 116 ( 1998 ) 659-669.
[6] U.H. Cho, J.O. Park, Mercury-induced oxidative stress in tomato seedings. Plant Sci. 156 ( 2000 ) 1-9.
[7] J.H.Christensen, G. Bauw, K.G. Welinder, M.V. Montagu, W. Boerjan, Purification and characterization of peroxidases correlated with lignification in poplar xylem. Plant Physiol. 118 ( 1998 ) 125-135. [8] D.S. Everdeen, S. Kiefer, J.J. Willard, E.P. Muldoon, Enzymatic
3cross-linkage of monomeric extensin precursors in vitro, Plant Physiol. 87 (1988) 616-621.
[9] S.M. Gallego, M.P. Benavides, M.L. Tomaro, Effect of heavey metal ion excess on sunflower leaces : evidence for involvment of oxidative stress. Plant Sci. 121 (1996 ) 151-159.
[10] I.G.Gazaryan, L.M. Largimini, G.A. Ashby, R.N.F. Thorneley, Mechanism of indole-3-acetic acid oxidation by plant peroxidase: anaerobic stopped-flow spectrophotometric studies on horseradish and
tobacco peroxidases. Biochem. J. 313 ( 1996 ) 841-847.
[11] K.E. Hammond-Kosack, J.D.G. Jones, Resistance gene-dependent plant defense response. Plant Cell 8 ( 1996 ) 1773-1791.
[12] K.Iiyama, TB-T. Lam, B.A. Stone, Covalent cross-links in the cell wall. Plant Physiol. 104 ( 1994 )315-320.
[13] S. Jana, M.A. Choudhuri,. Glycolate metabolism of three submerged aquatic angiosperms during aging. Aquat. Bot. 12 ( 1982 ) 345-354. [14] K.L.Klotz, T.T-Y.Liu, L. Liu, and L. M. Largimini,. Expression of the
tobacco anionic peroxidase gene is tissue-specific and development regulared. Plant Mol. Biol. 36 ( 1998 ) 509-520.
[15] L.M. Lagrimini, Wound-induced deposition of polyphenols in
transgenic plants overexpressing peroxidase. Plant Physiol. 96 ( 1991 ) 577-583.
[16] L.M. Lagrimini, S. Rothstein, Tissue specificity of tobacco peroxidase isozymes and their induction by wounding and tobacco mosaic virus infection. Plant Physiol. 84 ( 1987 ) 438-422.
[17] L.M. Lagrimini, S. Bradford, S. Rothstein, Peroxidase-induced wilting in transgenic tobacco plants. Plant cell 2 ( 1990 ) 7-18.
[18] L.M.Lagrimini, V. Gingas, F. Finger, S. Rothstein, T.T. Liu, Characterization of antisense transformed plants deficient in the tobacco anionic peroxidase. Pant Physiol. 114 ( 1997 ) 1187-1196. [19] T.C. Li, T.Y. Feng, W.S. Chen, Z.H. Liu, The acute effect of copper on
the levels of indole-3-acetic acid and lignin in peanut roots. Aust. J. Plant. Physiol. 28 ( 2001 ) 1-6.
[20] C.C. Lin, C.H. Kao,. Abscisic acid induced changes in cell wall peroxidase activity and hydrogen peroxide level in roots of rice
seedling. Plant Sci. 160 ( 2001 ) 323-329.
[21] Z.H. Liu, M.J. Ger, Changes of enzyme activity during pollen
germination in maize and possible evidence of lignin synthesis. Aust. J. Plant Physiol. 21 ( 1997 ) 329-335.
[22] M. Mäder, R. Füssl, Role of peroxidase in lignification of tobacco cells: regulation by phenolic compounds. Plant Physiol. 70 ( 1982 )
1132-1134.
[23] S. Mazhoudi, A. Chaoui, M.H. Ghorbal, E.E. Ferjani, Response of antioxidant enzymed to excess copper in tomato(Lycopersicon
esculentum Mill). Plant Sci. 127 ( 1997 ) 129-137.
[24] G. Müsel, T. Schindler, R. Bergfeld, K. Ruel, G.Jacquet, G. Lapierre, V. Speth, P. Schoptfer, Structure and distribution of lignin in primary and secondary cell walls of maize coleoptiles analyzed by chemical and immunological probes. Planta 201 ( 1997 ) 146-159.
[25] O. Ouariti, N.Boussama, M. Zarrouk, A. Cherif, M.H. Ghorbal, Cadmium- and copper-induced changes in tomato membrane lipids. Phytochemistry 45 ( 1997 ) 1343-1350.
[26] A. Polle, T. Otter, T. Seifert, Apoplastic peroxidase and lignification in needles of norway spruce ( picea abies L. ). Plant physiol. 106 ( 1994 ) 53-60.
[27] M. Quiroga, C. Guerrero, M.A. Botella, A. Barcelo, I. Amaya, M.I.
Medina, F.J. Alonso, S.M. Forchetti, H. Tigier, V. Valpuesta, A tomato peroxidase involved in the synthesis of lignin and suberin. Plant Physiol. 122 ( 2000 ) 1119-1127.
Aluminum induces oxidative stress genes in Arabidopsis thaliana. Plant Physiol. 116 ( 1998 ) 409-418.
[29] G. Sandmann, P. Böger, Copper-mediated lipid peroxidation processes in photosynthetic membranes. Plant Physiol. 66 ( 1980 ) 797-800. [30] H. Teisseire, V. Guy, Copper-induced changes in antioxidant enzymes
actives in fronds of duckweed ( Lemna minor ). Plant Sci. 153, ( 2000 ) 65-72.
[31] H.Thordal-Christensen, Z. Zhang, Y. Wei, D. B.Collinge, Subcellular localization of H2O2 in plants. Accumulation in papillae and
hypersensitive response during the barley powdery mildew interaction. Plant J. 11 ( 1997 ) 1187-1194.
[32] W.C. Wang, Z.H. Liu, Harpinpss-induced peroxidase and lignin
accumulation in tobacco during the hypersensitive response. Aust. J. Plant Physiol. 26 ( 1999 ) 265-272.
[33] J.E.J. Wecekx, H.M.M. Clijsters, Oxidative damage and defense mechanism in primary leaves of Phaseolus vulgarus as a result of root assimilation of toxicity amounts of copper. Physiol. Plant. 96, (1996 ) 506-512.
Figure 1. Effect of Cu on the growth of soybean roots.
Values are the means ± SE of three replicates. Hours after treatment
0 12 24 36 48 60 72 R oot f res h w e igh t ( mg / plant ) 0.00 0.08 0.10 0.12 0.14 0.16 0.18 0.20 Control 1 M CuSO4 2 M CuSO4 5 M CuSO4 10 M CuSO4 µ µ µ µ
Figure 2. Effect of Cu on the levels of H2O2 in soybean roots. Values are
the means ±SE of three replicates.
Hours after treatment
1 3 6 12 24 48 72 H 2 O 2 co n ten t ( m m o l g -1 fresh w e ight ) 0 1 2 3 4 5 6 Control 2 µM CuSO4 5 µM CuSO4 10 µM CuSO4
Figure 3. Histochemical analysis of H2O2 in soybean roots. H2O2 is
observed as a dark brown staining.
CuSO4 − ª − ª − ª − ª − ª
Figure 4. Effect of Cu on the total peroxidase isozymes during the course of
treatment in soybean roots. One unit of peroxidase isozymes was defined as the amount enzyme that causes the formation of 1 nmol tetraguaiacol per min.(extinction coefficient is 26.6 mM-1cm-1 at OD. 470).Values are the means ± SE of three replicates.
Hours after treatment
24 48 72 Pero xid ase activ ity ( U / m g p rotei n ) 0.0 0.5 1.0 Control 2 M CuSO4 5 M CuSO4 10 M CuSO4
Figure 5. Effect of Cu on the lignin content during the course of treatment
in soybean roots. Values are the means ± SE of three replicates. Hours after treatment
3 6 12 24 48 72 Li gnin c onte n t ( m g g -1 dr y w e ight ) 0 10 20 30 40 50 Control 10 µM CuSO4
Figure 6. Effect of Vu on the lignin content in Cu-treated soybean root tips
(1 cm) after 3 h incubation. Values are the means ± SE of three replicates.
Lignin c ont e n t ( mg g -1 dr y w e ight ) 0 1 2 3 4 5 6 7 8 Control 10 µM CuSO4
Figure 7. Effect of Cu on the activities of the peroxidase isozymes in the
soybean roots. Lane A ( 24 h, control ), Lane B ( 24 h, 2 µM Cu ), Lane C ( 24 h, 5 µM Cu ), Lane D ( 24 h, 10 µM Cu ), Lane E ( 48 h, control ), Lane F ( 48 h, 2 µM Cu ), Lane G ( 48 h, 5 µM Cu ), Lane H ( 48 h, 10 µM Cu ), Lane I ( 72 h, control ), Lane J ( 72 h, 2 µM Cu ), Lane K ( 72 h, 5 µM Cu ), Lane L ( 72 h, 10 µM Cu ). Isolectric points of peroxidases are marked on the right. 8.9 8.3 7.7 6.5 5.6 4.4 pH A B C D E F G H I J K L 24 h 48 h 72 h
Figure 8. Quantification of peroxidase activity in soybean roots of
Cu-treated and control tissue. Data were converted from the Figure 5.
24 48 72 In te n s it y 0 1 2 3 4 24 48 72 In te n s it y 0 1 2 3 4 5 24 48 72 In te n s ity 0 1 2 3 4 24 48 72 In te n s it y 0 2 4 6 8 10 24 48 72 In te n s it y 0 1 2 3 4 5 6 7 8 24 48 72 In te n s it y 0 1 2 3 4 pI 8.9 pI 8.3 pI 7.7 pI 6.5 pI 5.6 pI 4.4 A B C D E F
Figure 9. Effect of Cu on the activities of peroxidase isozymes in soybean root tips (1 cm ). 4.4 pH CuSO4 « − − « − « 0.5 h 1 h 3 h
Figure 10. The effect of Cu on the transcript accumulation of peroxidase
isozymes. ( A ) after 6 h incubation, ( B ) after 12 h incubation, ( C ) after 24 h incubation. In te n s it y 0 2 4 6 8 10 12 14 Int e nsit y 0 2 4 6 8 10 Intensi ty 0 2 4 6 8 10 12 0 2 4 6 8 10 12 14 16 0 2 4 6 8 10 12 0 2 4 6 8 10 12 0 5 10 15 20 0 5 10 15 0 5 10 15 20 A pI 4.4 C pI 4.4 B pI 4.4 pI 5.6 pI 5.6 pI 5.6 pI 7.7 pI 7.7 pI 7.7 pI 4.4 pI 5.6 pI 7.7 AC