行政院國家科學委員會專題研究計畫 成果報告
類固醇對牛角膜內皮細胞的生理影響(3/3)
研究成果報告(完整版)
計 畫 類 別 : 個別型 計 畫 編 號 : NSC 95-2314-B-002-010- 執 行 期 間 : 95 年 08 月 01 日至 96 年 07 月 31 日 執 行 單 位 : 國立臺灣大學醫學院眼科 計 畫 主 持 人 : 陳偉勵 共 同 主 持 人 : 胡芳蓉 計畫參與人員: 碩士級-專任助理:杜怡樺、林瑩美、黃玉華 研究生:周玉彬 處 理 方 式 : 本計畫可公開查詢中 華 民 國 96 年 10 月 31 日
行政院國家科學委員會補助專題研究計畫
█ 成 果 報 告
□期中進度報告
(計畫名稱)
類固醇對牛角膜內皮細胞的生理影響
計畫類別:▓ 個別型計畫
□ 整合型計畫
計畫編號:NSC95-2314-B -002 -010
執行期間:
95 年
8 月
1 日至 96 年
7 月
31 日
計畫主持人:陳偉勵 主治醫師
共同主持人:胡芳蓉 教授
計畫參與人員:
杜怡樺、林瑩美、黃玉華、周玉彬
成果報告類型(依經費核定清單規定繳交):□精簡報告 ▓完整
報告
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究
計畫、列管計畫及下列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,□一年□二年後可公
開查詢
執行單位:國立台灣大學醫學院眼科
中
華
民
國
96 年
10 月
31
日
中文摘要 角膜內皮細胞是影響角膜含水量的最重要組織。它可以經由細胞間的緊密相連, 和 Na-K-ATPase 作用而調節角膜含水量。然角膜內皮細胞雖有重要生理作用, 相關研究卻 相當缺乏。在角膜受到傷害,如發炎,手術傷害,角膜移植瓣排斥 時,類固醇 (Dexthamethasone)是廣泛利用的藥劑。若能了類固醇在角膜內皮細 胞上扮演的生理機轉,不但能擴展知識,甚至可引導至治療層面之上。近年來許 多研究指出類固醇在細胞的生理作用上扮演重要的腳色,例如它會影響傷口癒 合,細胞分裂,細胞移行,細胞凋零,Na-K-ATPase 活性和細胞骨骼的維持等等。 因此,我們利用初步培養的牛角膜內皮細胞做為細胞模式,並加入不同濃度的類 固醇 (Dexthamethasone) 。研究結果顯示,我們利用細胞免疫染色法,RT-PCR 和 西方墨點法先證實了牛角膜內皮細胞確實表現類固醇接受器。接著我們利用MTS 和flow cytometry 方式偵測類固醇是否影響牛角膜內皮細胞的細胞分裂及細胞 凋零,結果顯示在加入高濃度的類固醇確實會降低細胞分裂的能力而增進細胞凋 零。我們並利用非同位素的方法來測定類固醇是否影響牛類角膜內皮細胞的 Na-K-ATPase 酵素活性,結果顯示在加入某種濃度的類固醇會降低牛類角膜內皮 細胞的Na-K-ATPase 酵素活性。 關鍵字:角膜內皮細胞,細胞凋零,類固醇,細胞分裂,Na-K-ATPase 酵素活性
ABSTRACT
Purpose: To assess the in-vitro effects of dexamethasone (DEX) on the proliferation,
apoptosis, and Na+-K+-ATPase activity of bovine corneal endothelial cells.
Methods: Bovine corneal endothelial cells were cultured with DEX ranging from
10−10 to 10−3 M. The effect of DEX on the proliferation was analyzed by 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxy-methoxyphenyl)-2-(4-sulfophenyl)-2H-tetr azolium inner salt (MTS) assay. Apoptosis and necrosis were detected by staining with fluorescein-conjugated annexin V and propidium iodide, followed by flow cytometry. The effect of DEX on Na+-K+-ATPase activity was evaluated using non-isotopic methods.
Results: DEX did not affect cellular proliferation or induce apoptosis/necrosis from
10−10 to 10−5 M. At 10−4 and 10−3 M, DEX significantly decreased proliferation and increased apoptosis and/or necrosis. DEX significantly increased the Na+-K+-ATPase activity from 10−8to 10−6M, with the maximal effect at 10−6M (p < 0.01); this effect was inhibited by RU38486, an antiglucocorticoid molecule.
Conclusions: Bovine corneal endothelial cells express glucocorticoid receptor (GR)
mRNA and protein. DEX decreases cell proliferation and induces cellular apoptosis and/or necrosis at high concentrations. DEX also increases the Na+-K+-ATPase activity at certain concentrations.
KEYWORDSCorneal endothelial cells; apoptosis; dexamethasone; proliferation; Na+-K+-ATPase
Ocular Immunology and Inflammation, 14:215–223, 2006
INTRODUCTION
Dexamethasone (DEX), a glucocorticoid clinically used as an anti-inflammatory agent and immunosuppressant, exerts dramatic effects on various tissues. The action of DEX is believed to be mediated by glucocorticoid receptors (GR).1–3DEX inhibits
inflammation through multiple mechanisms.4–6Interestingly, the GR response gene
may positively or negatively regulate the actions according to different types of cells and different dosages of DEX treatment.7,8 Although not clearly defined, the dual
effects of glucocorticoids may be the result of cross-talk between nuclear comodulators, or of interactions of transcription factors.9,10Different types of ocular
tissues are known to express GR and are responsive to glucocorticoid treatment.7,8,11–13 Although DEX effectively reduces ocular surface inflammation,
delays excess corneal wound healing, and reduces pain, prolonged use of
DEX induces numerous ocular side effects such as glaucoma and cataract.14–16
Numerous studies have demonstrated the changes of corneal stromal and epithelial wound healing after DEX use, implying that steroid eyedrops impaired wound healing rather than having no effect.17,18 Under particular conditions, DEX also affected
proliferation and induced apoptosis and/or necrosis in corneal epithelial cells and keratocytes.7,8Corneal endothelium controls the hydration status of the cornea. The
cell density, pumping activity including Na+-K+-ATPase, and the cell-cell barrier are the major factors maintaining the functional homeostasis of the cornea.19–21 Unlike
corneal epithelium or keratocytes, human corneal endothelial cells have limited proliferative capacity in vivo.21,22 The aging process, trauma, corneal graft rejection,
stress caused by certain systemic diseases, endothelial dystrophies, and improper medications may all cause corneal endothelial cell loss and permanent damage.23–25
To avoid this, appropriate medications that diminish damage to corneal endothelial cells and promote corneal endothelial cellular function are extremely important. Clinically, DEX is frequently used for corneal endothelial diseases.26–28
Little is known, however, about the direct effects of DEX on these cells. In the present study, we used cultured bovine corneal endothelial cells as the experimental model. Whether or not GR mRNA and its corresponding protein are expressed in bovine corneal endothelial cells and the in-vitro effects of DEX on these endothelial cells, such as proliferation, apoptosis, necrosis, and Na+-K+-ATPase activity, were assessed.
MATERIALS AND METHODS
Cell Culture of Bovine Corneal Endothelial Cells
Adult bovine corneal buttons were obtained within 3 h after enucleation. Corneal endothelialcellswith Descemet’smembranewere dissected underthemicroscopeand separated from the stroma after treatment with trypsin-EDTA (Invitrogen-Gibco, Paisley, UK).Cellswere cultured in Dulbecco’smodified Eagle’smedium (DMEM) with 10% fetal calf serum (Gibco BRL/Life Technologies, Rockville, MD, USA).
Second passage cells were used in all the experiments. For the proliferation and apoptosis parts of the experiment, we added 2×10−5cells/well in 24-well tissue culture plates (Costar) and waited 1 d before DEX was added. For the Na+-K+-ATPase activity part of the study, we waited until the cells were confluent before DEX was added in order to mimic in-vivo conditions.
Drug Preparation
DEX, purchased from Sigma-Aldrich (Saint Quentin Fallavier, France), was diluted in 0.1% absolute ethanol before being added to the cultured medium. On the second day, tissue culture medium was replaced by 1 ml medium containing various concentrations of DEX (10−3, 10−4, 10−5, 10−6, 10−7, 10−8, 10−9, and 10−10 M). The culture media were changed daily for 6d. Mifepristone (RU38486; Roussel Uclaf, Romainville, France), an antiglucocorticoid molecule, was dissolved in absolute ethanol. RU38486 (10−5 M) was added daily to the culture medium, alone or in combination with 10−6M DEX, for six days. All experiments were repeated six times.
Glucocorticoid Receptor Analysis
GRs were studied using reverse transcriptasepolymerase chain reaction (RT-PCR). cDNA was prepared from 1 μg total RNA by RT in a 20 μl volume, using reagents from a commercially available kit (Promega, Pittsburgh, PA, USA). Primers specific for bovine GR were designed using MacVector 5.0 software (Oxford Molecular Group, Oxford, UK).The primer pairs used were: GR upstream sequence,
5_-GTTTCTGCGTCTTCACCCTCA-3_; downstream sequence,
5_-GTCTCTCCCATATACAGTCCC-3_. PCR was performed in a reaction mixture containing 1 μg cDNA and 2.5 μM each of the upstream and downstream primers, plus reagents from a commercially available kit (Gibco). PCR was performed in a thermal cycler. The PCR products were electrophoresed in 2% agarose gels containing ethidium bromide and photographed under UV illumination (665 film; Polaroid, Cambridge, MA, USA). Additionally, the PCR product was sequenced and checked for percent homology with the sequence from 1 to 158 bp of bovine GR, specified as BTU37383 in the Genbank.
Immunocytochemical
Localization
of
Glucocorticoid
Receptors
Bovine corneal endothelial cells cultured with 10−6 M DEX for 6 d were fixed, permeabilized, blocked, and incubated with primary antibody at room temperature for 2 h. The primary antibody, a rabbit polyclonal antibody(No. PAI-511, Dianova-Immunotech, Hamberg, Germany), was confirmed to have positive immunocytochemical staining for GR in bovine cells.29 The cells were then incubated with fluorescein isothiocyanate (FITC)-conjugated goat-anti-rabbit antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA) for 1 h at 37◦C. The cells were then washed and mounted in medium containing propidium iodide (PI; Vector Laboratories, Burlingame, CA, USA). Immunocytochemical study was also conducted on transverse cryosections of bovine cornea. The staining patterns were observed by conventional fluorescence microscopy using an Eclipse E800 Nikon
microscope with a VFM Epi-fluorescence attachment (Nikon, Melville, NY, USA) equipped with a Spot digital camera and Spot version 1.1 CE software (Diagnostic Instruments, Sterling Heights, MI, USA).
SDS-PAGE and Western Blotting of Glucocorticoid Receptors
Cultured cells were trypsinized, suspended, and homogenized. Equal amounts of protein were electrophoresed and transferred to a polyvinylidene difluoride membrane (Millipore Corp, Bedford, MA, USA). After blocking, the membranes were incubated with 1:750 polyclonal rabbit anti-human GR antibody (Rabbit, No. PAI-511, Dianova-Immunotech, Hamburg, Germany). The membranes were then washed and incubated with 1:6000 horseradish peroxidase-conjugated goat anti-rabbit antibody (Santa Cruz Biotechnology).The signal was detected using enhanced chemoluminescenceaccording to themanufacturer’sinstructions(Pierce,Rockford, IL, USA) and exposed to autoradiographic film.
MTS Assay
3-(4,5-Dimethylthiazol-2-yl)-5-(3-carboxy-methoxyphenyl)-2-(4-sulfophenyl)-2H-t etrazolium inner salt (MTS) and electron coupling reagent were purchased from Promega and Sigma, respectively. After 2, 4, and 6d of culture, 100 μl culture medium and 20 μl reagents were incubated at 37◦C for 1 h. In addition, corneal endothelial proliferation was analyzed by staining the nucleus with amidoblack (Sigma) and counted using a hemocytometer and a cell counter (Coulter, Hialeah, FL, USA). To clarify whether the change of living cellular amount was caused by cytotoxicity or change of proliferation, immunocytochemical staining with
Ki67 was performed to evaluate the proliferative status.
Apoptosis Assay
The cells, cultured in different concentrations of DEX for 6 d, were washed with cold PBS, and 1×106 cells were incubated in 195 μl annexin-binding buffer containing 5 μl annexin V and 5 μl 20 μg/ml PI for 15 min at room temperature. After incubation, 400 μl annexin-binding buffer was added. The cells were then analyzed by flow cytometry (FACSCalibur, Becton Dickinson, Rungis, France). At least 20,000 cells were analyzed for each experiment. AV-/PI- cells were considered viable, AV+/PI- cells were considered early apoptotic cells, and AV+/PI+ cells were considered necrotic cells. Bovine corneal endothelial cells cultured in DMEM medium with 0.1% absolute ethanol were used as the control group. All experiments were repeated six times.
Na
+-K+-ATPase Activity
Cells were harvested 2, 4, and 6 d afterDEX exposure for the determination of Na+-K+-ATPase activity. Aliquots of the suspension of bovine endothelium cell homogenates were used. Na+-K+-ATPase activity was quantified using the method of Hwang and colleagues with minor modifications.30–32 In brief, the sample was prepared in reaction buffer. After incubation, ice-cold trichloroacetic acid was added to stop the reaction. The vials were centrifuged, and the supernatant and the phosphate
standard were added with 2.5% acid molybdate solution, 10% SDS, and 0.025% 1-amino-2- naphthol-4-sulformic acid. After incubation, the inorganic phosphate production was measured at 700 nm according to the method described by Peterson.33 The enzyme activity of Na+-K+-ATPase was defined as the difference between the inorganic phosphate liberated in the presence and absence of 0.5 mM oubain in the reaction mixture, and calculated by comparison with the concentration of inorganic phosphate standard. In this study, the percentage of activity compared to the control group was used for analysis.
Statistical Analysis
Experimental data were analyzed using a one-factor analysis of variance (ANOVA), Dunnett’smultiplecomparison test,and Student’stwo-tailed t-test. The results are expressed as the mean ± standard error of the mean (SEM). The probability of error p
< 0.05 was considered to be statistically significant.
RESULTS
Detection of Glucocorticoid Receptor mRNA in Cultured
Bovine Corneal Endothelial Cells
RT-PCR demonstrated the existence of GR mRNA in cultured bovine corneal endothelial cells. A unique PCR product of 158 nucleotides was detected with ethidium bromide after gel electrophoresis (Fig. 1). The size of the band was consistent with the expected fragment size, determined from bovine GR cDNA. Additionally, the PCR product was sequenced and exhibited 97% homology with bovine GR BTU37383 in the Genbank.
Detection of Glucocorticoid Receptor Protein in Cultured
Bovine Corneal Endothelial Cells
Specific nuclear staining of GRs was observed in cultured cells (Fig. 2 A, B) and corneal endothelial cells on tissue sections (Fig. 2 C, D). The negative control showed no staining (data not shown). The Western blot results also demonstrated the existence of GR protein in the freshly obtained bovine corneal endothelial cells and cultured bovine corneal endothelial cells (Fig.2E–G).
Effect of DEX on Cellular Proliferation Evaluated by MTS
Assay
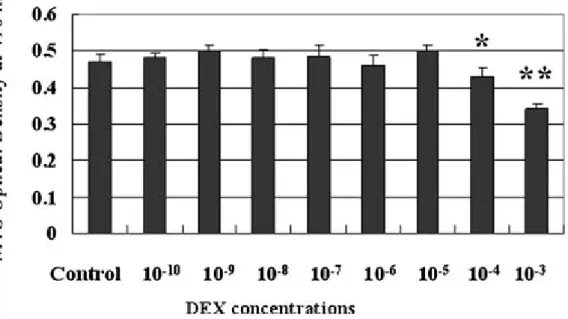
After six days of culture, DEX did not change the amount of living cells at concentrations ranging from 10−10 to 10−5 M. In contrast, DEX at 10−4 and 10−3 M significantly decreased the amount of living cells (p < 0.01; Fig. 3). There were no significant effects of DEX on the amount of living cells at Day 2 and Day 4 (data not shown). All of the results of the MTS assays were confirmed by flow cytometry and hemocytometer counting assays (data not shown).
Apoptosis Assay
After six days of culture, the percentage of viable cells, apoptotic cells, and necrotic cells did not change at DEX concentrations ranging from 10−5M to 10−10M. However, the percent viable cells decreased when 10−4 and 10−3 M DEX was added to the
cultured medium. Corneal endothelial apoptosis and necrosis were significantly enhanced when 10−4and 10−3M DEX was added (p < 0.01; Fig. 4). All of the results of the apoptosis/necrosis assay were compatible with the live/dead cell assay (data not shown). There were no significant effects of DEX on the percentage of viable cells, apoptotic cells, and necrotic cells on Day 2 and
Day 4 (data not shown).
Effect of DEX and RU38486 on Na
+-K+-ATPase Activity
After six days of culture, we observed a dose dependent effect of DEX on Na+-K+-ATPase activity of bovine corneal endothelial cells at DEX concentrations of 10−8M and 10−6M. However, at higher (10−4M) or lower (10−10M) concentrations of DEX, there was no significant difference compared to the control group (Fig. 5A). There were no significant effects of DEX on Na+-K+-ATPase activity on Day 2 and Day 4 (data not shown). The addition of 10−5 M RU38486 alone to the culture medium had no significant effect on the Na+- K+-ATPase activity compared to the effect of the ethanol control treatment. Nonetheless, it significantly decreased the promoting effect of Na+-K+-ATPase activity at 10−6M DEX (Fig. 5B).
DISCUSSION
Corneal endothelial cells participate in maintaining corneal transparency by barrier and ‘pump’ functions.21 Since human corneal endothelial cells have limited
proliferative capacity throughout the human lifespan and may lose the ability to maintain corneal transparency once cellular loss reaches a critical level, any insult that potentially causes corneal endothelial cell damage should be recognized and avoided.21,22 DEX is widely used for the treatment of ocular inflammation. The
originality of the present work is the possible effect of low-dose DEX on the various important cellular functions of cultured bovine corneal endothelial cells. We first used immunocytochemistry, RT-PCR, and Western blot analysis to show that GR mRNA and protein are expressed in cultured bovine corneal endothelial cells. We also used RU38486 (mifepristone), an 11-substituted 19-norsteroid steroid antagonist with antiprogesterone and antiglucocorticoid effects,34 to inhibit the function of DEX.
RU38486 inhibited the 10−6 M DEX-induced increase in Na+- K+-ATPase activity. This is pharmacologic evidence of the existence of functional GRs in corneal endothelial cells. Cenedella and Fleschner35 reported cholesterol biosynthesis by the
rabbit cornea and showed that possible endogenous metabolites synthesized from cholesterol can be found in the cornea. Since GRs were identified in all three layers of corneal cells, it is reasonable to suggest that endogenous steroids may have autocrine and/or paracrine roles in the cornea and that exogenous steroids may have direct functions on these cells.
It has been shown that AP1 components (c-Fos and c-Jun) are expressed in corneal endothelial cells.36,37 These proto-oncogenes may play an important role in
proto-oncogenes. GR and AP1 interactions have been described in the glucocorticoid response elements (GREs), where these two transcriptional factors are adjacent.38We
observed non-responsiveness to DEX for endothelial cell proliferation at DEX concentrations from 10−10to 10−5M, and significant inhibition of cell proliferation at higher concentrations (10−4 M and 10−3 M, p < 0.01). A previous study using immunostaining with Ki 67 demonstrated that confluent cultured rat corneal endothelial cells cease to proliferate.39 We also found no proliferation in all
experimental groups in this study on Days 2, 4, and 6 (data not shown). The change in the MTS assay result is thus supposed to be caused by cytotoxicity instead of altered proliferation.
The similar response curve for the hemocytometer cellcounts and the MTS assay results provides evidence that they measured similar effects. DEX induced an apparent paradoxical biphasic effect (i.e., proliferation after exposure to low-dose DEX and inhibition of growth after exposure to high-dose DEX) on human corneal epithelial cells, rabbit conjunctival cells, dermal fibroblasts, retinal pigment epithelial cells, and human corneal keratocytes.7,8,40,41 It is quite interesting to find that cultured
bovine corneal endothelial cells seem to be more refractive to survival/proliferation under treatment of DEX. In vivo, corneal endothelial cells are unique compared to other corneal cells for having a limited proliferative capacity due to various intrinsic and extrinsic factors.21 Although an in-vitro system differs from an in-vivo one, it is
not surprising that DEX does not increase cellular proliferation of cultured corneal endothelial cells. However, when the concentration of DEX reached 10−4 to 10−3M, cell numbers significantly decreased. Since the percentage of apoptotic and necrotic cells also increased at this concentration, we believe that the decreased cell number was mainly due to the significant increase in cellular death.
Apoptosis in the corneal endothelium has been suggested to play a major role in cell loss of various conditions.42,43 Glucocorticoid-induced cell death or apoptosis has
been described in various cell systems.44–46 However, glucocorticoid was shown to
protect other cell types, such as epithelial cells of the mammary gland, hepatocytes, and thymocytes.47,48 GR, as a signaling protein, can display protoapoptotic or
anti-apoptotic activity depending on cell context. Cross talks between ubiquitous signaling proteins can produce context-specific effects on cell survival.49–51We found
no significant change in the apoptosis or necrosis rate of corneal endothelial cells with DEX concentrations from 10−10 to 10−5 M (10−7 to 10−8 M, intracameral concentrations after topical usage).52,53 In contrast, the same concentrations of DEX
did induce apoptosis or necrosis of human keratocytes and corneal epithelial cells.7,8
The refractiveness of DEX to induce apoptosis on corneal endothelial cells can be roughly explained by two mechanisms: refractivenessto inducethe‘death gene’by activated GR or refractiveness to repress the survival function.8 At higher concentrations of DEX (10−4 to 10−3 M, close to eyedrop concentrations), however, toxicity which caused cellular apoptosis and necrosis was found. This finding differs from those of some other studies. Reinhard and Sundmacher54 showed that
intracameral steroid injection seemed to be a safe and helpful therapeutic measure in the treatment of moderate and severe endothelial immune reactions without complications to the human corneal endothelial cells. Wang et al.55 used an in-vitro
MTT viability assay and demonstrated relatively low complication rates for steroid therapy of porcine corneal endothelial cells compared to some antibiotics. Although their study results are different from ours, Solomon et al.56 demonstrated adverse
effects of corticosteroid on the morphology of cat corneal endothelial cells after trauma. In spite of the need for in-vivo studies to prove our DEX effects, our results point out the potential side effects of DEX on corneal endothelial cells and the necessity for using DEX with caution.
As for the effect of DEX on Na+-K+-ATPase activity, we found a small but statistically significant increase in enzyme activity at concentrations of 10−6 and 10−8 M (p < 0.01). Previously, very few reports demonstrated the possible pharmacologic increase in Na+-K+-ATPase activity in rabbit corneal endothelial cells,57 and our
study is the first to demonstrate that DEX increased Na+-K+-ATPase activity in these cells.There are several studies showing the effects of DEX to regulate Na+-K+-ATPase gene expression through multiple complex mechanisms, including transcription, posttranscriptional, translational, and enzyme activity in a variety of tissues.58–62 In some cells, one or more of these processes may coexist in the
regulatory process. Since we did not examine the sodium pump protein levels or mRNA levels, the mechanisms for DEX to increase enzyme activity of corneal endothelial cells await to be defined. The glucocorticoid-induced increase of Na+-K+-ATPase in this study, however, suggests a potential contribution of the effectiveness of DEX in treating corneal endothelial dysfunction. Although DEX seemed to increase Na+-K+-ATPase activity in this study, which may play a beneficial role in the enhancement of the recovery of corneal endothelial pumping dysfunction, DEX can also induce unwanted corneal endothelial cellular necrosis and apoptosis at higher concentrations. Long-term and high-dose DEX administration induces glaucoma, cataract, and other ocular complications.17,22–27 Therefore,
shortening the treatment course and adjusting the dosage of DEX are important. In addition, the in-vivo effect of DEX on Na+-K+-ATPase activity still needs to be confirmed.
In conclusion, our experimental results indicate the expression of GRs in cultured bovine corneal endothelial cells. Such receptors may influence bovine corneal endothelial proliferation, apoptosis/necrosis, and Na+-K+-ATPase activity in a dose-dependent way. Although the in-vitro effects do not necessarily correlate with those found in vivo, human corneal endothelial cells may differ from bovine cells, and young and non-confluent cells may not be similar to old and confluent cells, our results at least suggest that DEX may influence the physiological function of young bovine corneal endothelial cell in vitro instead of having no effect. The in-vivo function of DEX on corneal endothelial cells warrants further study.
REFERENCES
[1] Funder JW. Mineralocorticoids, glucocorticoids, receptors and response elements.
Science. 1993;259:1132–1133.
[2] Beato M. Gene regulation by steroid hormones. Cell. 1989;56:335–344.
[3] Webster JC, Cidlowski JA. Mechanism of glucocorticoid-receptormediated repression of gene expression. Trends Endocrinol Metab. 1999;10:396–402.
[4] Shimoyama M, Shimmura S, Tsubota K, Oguchi Y. Suppression of nuclear factor kappa B and CD18-mediated leukocyte adhesion to the corneal endothelium by dexamethasone. Invest Ophthalmol Vis Sci. 1997;38:2427–2431.
[5] Goulding NJ, Euzger HS, Butt SK, Perretti M. Novel pathways for glucocorticoid effects on neutrophils in chronic inflammation. Inflamm Res. 1998;47:158–165. [6] Adcock IM, Caramori G. Cross-talk between pro-inflammatory transcriptions factors and glucocorticoids. Immunol Cell Biol. 2001;79:376–384.
[7] Bourcier T, Borderie V, Forgez P, Lombet A, Rostene W, Laroche L. In vitro effects of dexamethasone on human corneal keratocytes. Invest Ophthalmol Vis Sci. 1999;40:1061–1070.
[8] Bourcier T, Forgez P, Borderie V, Scheer S, Rostene W, Laroche L. Regulation of human corneal epithelial cell proliferation and apoptosis by dexamethasone. Invest
Ophthalmol Vis Sci. 2000;41:4133–4141.
[9]MckennaNJ,Xu J,Nawaz Z,TsaiSY,TsaiMJ,O’Malley BW.Nuclearreceptor coactivators: multiple enzymes, multiple complexes, multiple functions. J Steroid
Biochem Mol Biol. 1999;69:3–12.
[10] Jenkins BD, Pullen CB, Darimont BD. Novel glucocorticoid receptor coactivator effector mechanisms. Trends Endocrinol Metab. 2001;12:122–126.
[11]Weinreb RN,Bloom E,BaxterJD,Alvarado J,Lan N,O’DonnellJ,Polansky JR. Detection of glucocorticoid receptors in cultured human trabecular cells. Invest
Ophthalmol Vis Sci. 1981;21:403–407.
[12] Southren AL, Dominguez MO, Gordon GG, Wenk EJ, Hernandez MR, Dunn MW, Weinstein BI. Nuclear translocation of the cytoplasmic glucocorticoid receptor in the iris-ciliary body and adjacent corneoscleral tissue of the rabbit following topical administration of various glucocorticoids. A rapid screeningmethod for glucocorticoid activity. Invest Ophthalmol Vis Sci. 1983;24:147–152.
[13] Lin MT, Eiferman RA, Wittliff JL. Demonstration of specific glucocorticoid binding sites in bovine cornea. Exp Eye Res. 1984;38:333–339.
[14] Becker B, Mills DW. Elevated intraocular pressure following corticosteroid eye drops. J Am Med Assoc. 1963;185:884–886.
[15] Cronin TP. Cataract with topical use of corticosteroid and idoxuridine. Arch
Ophthalmol. 1964;72:198–199.
[16] Burde RM, Becker B. Corticosteroid-induced glaucoma and cataract in contact lens wearers. J Am Med Assoc. 1970;213:2075–2077.
anti-inflammatory agents on corneal re-epithelization. Invest Ophthalmol Vis Sci. 1981;20:688–691.
[18] Petroutsos G, Guimaraes R, Giraud JP, Pouliquen Y. Corticosteroids and corneal epithelial wound healing. Br J Ophthalmol. 1982;66:705–708.
[19] Maurice DM. The location of the fluid pump in the cornea. J Physiol. 1972;221:43–54.
[20] Ewart HS, Klip A. Hormoneal regulation of the Na(+)-K(+)-ATPase: mechanisms underlying rapid and sustained changes in pump activity. Am J Physiol. 1995;269:295–311.
[21] Joyce NC. Proliferative capacity of the corneal endothelium. Prog Retinal Eye
Res. 2003;22:359–389.
[22] Joyce NC, Meklir B, Joyce SJ, Zieske JD. Cell cycle protein expression and proliferative status in human corneal cells. Invest Ophthalmol Vis Sci.
1996;37:645–655.
[23] Schultz RO, Matsuda M, Yee RW, Edelhauser HF, Schultz KJ. Corneal endothelial changes in type I and type II diabetes mellitus. Am J Ophthalmol. 1984;98:401–410.
[24] Bourne WM, Nelson LR, Hodge DO. Continued endothelial cell loss ten years after lens implantation. Ophthalmology. 1994;101:1014–1022.
[25] Wu KY, Hong SJ, Huang HT, Lin CP, Chen CW. Toxic effects of mitomycin-C on cultured corneal keratocytes and endothelial cells. J Ocul Pharmacol Ther. 1999;15:401–411.
[26] Sanchez J, Polack FM. Effect of topical steroids on the healing of corneal endothelium. Invest Ophthalmol. 1974;13:17–22.
[27] Chung JH, Paek SM, Choi JJ, Park YK, Lee JS, Kim WK. Effect of topically applied 0.1% dexamethasone on endothelial healing and aqueous composition during the repair process of rabbit corneal alkali wounds. Curr Eye Res. 1999;18:110–116. [28] Olsen EG, Dacanger M. The effect of steroids on the healing of the corneal endothelium. An in vivo and in vitro study in rabbits. Acta Ophthalmol (Copenh).. 1984;62:893–899.
[29] Boos A, Kohtes J, Stelljes A, Zerbe H, Thole HH. Immunohistochemical assessment of progesterone, oestrogen, and glucocorticoid receptors in bovine placentomes during pregnancy, induced parturition, and after birth with or without retension of fetal membranes. J Reprod Fertil. 2000;120:351–360.
[30] Hwang PP, Sun CM, Wu SM. Characterization of gill Na+- K+-activated adenosine triphosphatase from tilapia Oreochromis mossambicus. Zool Stud.. 1988;27:49–56.
[31] Weng CF, Chiang CC, Gong HY, Chen MH, Lin CJ, Huang WT, Cheng CY, Hwang PP,Wu JL. Acute changes in gill Na+-K+-ATPase and creatine kinase in response to salinity changes in the euryhaline teleost, tilapia (Oreochromis
mossambicus). Physiol Biochem Zool. 2002;75:29–36.
specific activity, protein determination. Methods Enzymol. 1988;156:105–115.
[33] Peterson GL. A simplified method for analysis of inorganic phosphate in the presence of interfering substances. Anal Biochem. 1978;84:164–172.
[34] Cadepond F, Ulmann A, Baulieu EE. RU486 (mifepristone): mechanisms of actions of action and clinical uses. Annu Rev Med. 1997;48:129–156.
[35] Cenedella RJ, Fleschner CR. Cholesterol biosynthesis by the cornea. Comparison of rates of sterol synthesis with accumulation during early development. J Lipid Res. 1989;30:1079–1084.
[36] Feldman ST, Gately D, Seely BL, Schonthal A, Feramisco JR. Stimulation of DNA synthesis and c-fos expression in corneal endothelium by insulin or insulin-like growth factor-1. Invest Ophthalmol Vis Sci. 1993;34:2105–2111.
[37] Nguyen V, Schonthal A, Feldman ST. Expression of c-Jun protooncogene in corneal endothelium. Exp Eye Res. 1994;59:335–341.
[38] Pearce D, Matsui W, Miner JN, Yamamoto KR. Glucocorticoid receptor transcriptional activity determined by spacing of receptor and nonreceptor DNA sites.
J Biol Chem. 1998;273:30081–30085.
[39] Joyce NC, Harris DL, Mello DM. Mechanisms of mitotic inhibitionin corneal endothelium: contact inhibition and TGF-ß2. Invest Ophthamol Vis Sci.
2002;43:2152–2159.
[40] Blumenkranz MS, Claflin A, Hajek AS. Selection of therapeutic agents for intraocular proliferative disease: cell culture evaluation. Arch Ophthalmol.
1984;102:598–604.
[41] He S, Wang HM, Ye J, Ogden TE, Ryan SJ, Hinton DR. Dexamethasone induce proliferation of cultured retinal pigment epithelial cells. Curr Eye Res.
1994;13:257–261.
[42] Wilson SE, Li Q, Weng J, Barry-Lane PA, Jester JV, Liang Q, Wordinger RJ. The Fas-Fas ligand system and other modulators of apoptosis in the cornea. Invest
Ophthalmol Vis Sci. 1996;37:1582–1592.
[43] Cho KS, Lee EH, Choi JS, Joo CK. Reactive oxygen species-induced apoptosis and necrosis in bovine corneal endothelial cells. Invest Ophthalmol Vis Sci. 1999;40:911–919.
[44] Dowd DR, Miesfeld RL. Evidence that glucocorticoid- and cyclic AMP-induced apoptosis in lymphocytes share distal events. Mol Cell Biol. 1992;12:3600–3608. [45] Evans-Storms RB, Cidlowski JA. Regulation of apoptosis by steroid hormones. J
Steroid Biochem Mol Biol. 1995;53:1–8.
[46] Cidlowski JA, King KL, Evans-Storms RB, Montague JW, Bortner CD, Hughes Jr, FM. The biochemistry and molecular biology of glucocorticoid-induced apoptosis in the immune system. Recent Prog Horm Res. 1996;51:457–490.
[47] Fenz Z, Marti A, Jehn B, Altermatt HJ, Chicaiza G, Jaggi R. Glucocorticoid and progesterone inhibit involution and programmed cell death in the mouse mammary gland. J Cell Biol. 1995;131:1095–1103.
dexamethasone of transforming growth factor beta-ainduced apoptosis in rat hepatoma cells: a possible association with Bcl-xl induction. Hepatology.
1998;27:959–966.
[49] Jamieson CAM, Yamamoto KR. Crosstalk pathway for inhibition of glucocorticoid-induced apoptosis by T cell receptor signaling. Proc Natl Acad Sci
USA. 2000;97:7319–7324.
[50] Cohen JJ, Duke RC. Glucocorticoid activation of a calciumdependent endonuclease in thymocyte nuclei leads to cell death. J Immunol. 1984;132:38–42. [51] Ohoka Y, Kuwata T, Asada A, Zhao Y, Mukai M, IwataM. Regulation of thymocyte lineage commitment by the level of classical protein kinase C activity. J
Immunol. 1997;158:5707–5716.
[52] Watson D, Noble MJ, Dutton GN, Midgley JM, Healey TM. Penetration of topically applied dexamethasone alcohol into human aqueous humor. Arch
Ophthalmol. 1988;106:686–687.
[53] Weijtens O, Schoemaker RC, Romijn FP, Cohen AF, Lentjes EG, Van Meurs JC. Intraocular penetration and systemic absorption after topical application of dexamethasone disodium phosphate. Ophthalmology. 2002;109:1887–1889.
[54] Reinhard T, Sundmacher R. Adjunctive intracameral application of corticosteroids in patients with endothelial immune reactions after penetrating keratoplasty: a pilot study. Transplant Int. 2002;15:81–88.
[55] Wang HZ, Chang CH, Lin CP, Tsai MC. Using MTT viability assay to test the cytotoxicity of antibiotics and steroid to cultured porcine corneal endothelial cells. J
Ocul Pharmacol Ther. 1996;12:35–43.
[56] Solomon A, Solberg Y, Belkin M, Landshman N. Effect of corticosteroids on healing of the corneal endothelium in cats. Graefes Arch Clin Exp Ophthalmol. 1997;235:325–329.
[57] Wigham CG, Turner HC, Swan J, Hodson SA.Modulation of corneal endothelial hydration control mechanisms by Rolipram.. Pflugers Arch. 2000;440:866–870. [58] Bhutada A, Wassynger WW, Ismail-Beigi F. Dexamethasone markedly induces Na,K-ATPase mRNA beta 1 in a rat liver cell line. J Biol Chem.
1991;266:10859–10866.
[59] Barquin N, Ciccolella DE, Ridge KM, Sznajder JI. Dexamethasone upregulates the Na-K-APTase in rat alveolar epithelial cells. Am J Physiol. 1997;273:825–830. [60] Hao H, Wendt CH, Sandhu G, Ingbar DH. Dexamethasone stimulates transcription of the Na+-K+-ATPase beta 1 gene in adult rat lung epithelial cells. Am
J Physiol Lung Cell Mol Physiol. 2003;285:593–601.
[61] Celsi G, Wang ZM, Akusjarvi G, Aperia A. Sensitive periods for glucocorticoids’regulation ofNa(+),K(+)-ATPase mRNA in the developing lung and kidney. Pediatr Res. 1993;33:5–9.
[62] Devarajan P, Benz Jr, EJ. Translational regulation of Na-K-ATPase subunit mRNAs by glucocorticoids. Am J Physiol Renal Physiol. 2000;279:1132–1138.
FIGURE 1 Electrophoretic profile of the bovine glucocorticoid receptor product obtained from reverse transcription-polymerase chain reaction (RT-PCR). One microgram of total RNA extracted from cultured corneal endothelial cells was reverse-transcribed using oligo(dN) and oligo(dT) primers. The obtained cDNA was electrophoresed, and the bands were visualized by ethidium bromide staining, Lane 1: cultured bovine corneal endothelial cells showing a length of 158 bp. Lane 2: GAPDH (BG223947. 1M0017C04 Bovine, 2001 in Genbank) as positive control. GAPDH is the product of a bovine skeletal muscle structural gene with 516 bp. Lane 3: PCR negative control. Lane M: 100 bp DNA ladder.
FIGURE 2 Immunocytochemical staining and Western blot analysis demonstrated the expression of GR protein in cultured bovine corneal endothelial cells and fresh obtained corneal endothelial cells. (A,B) Immunocytochemical staining of cultured corneal endothelia cells with antiglucocorticoid receptor antibody. (C,D) Immunocytochemical staining of corneal sections with antiglucocorticoid receptor antibody. Expression of GR was found in the nucleus in both cultured cells and tissue sections. Green: nuclear staining of GR. Red: stained for propidium iodide, which is a nuclear marker. Inverted fluorescent microscopy. Original magnification: ×400. (E–G) Western blot analysis confirmed the existence of GR in cultured bovine corneal in (E) medium with DEX, (F) medium without DEX, and (G) freshly obtained corneal endothelial cells. The band at 97 kD confirmed the expression of GR, while the band at 47 kD was β-actin (internal control).
FIGURE 3 Corneal endothelial survival studied by MTS assay. Bovine cornea1 endothelial cells were cultured with various concentrations of DEX (10−3, 10−4, 10−5, 10−6, 10−7, 10−8, 10−9, and 10−10M) diluted in 0.1% absolute ethanol for 6 d. Results of MTS assay are expressed in optical densities, measured
spectrophotometrically at 490 nm. Bars represent mean ± SEM (n = 6 in each group). The control consisted of corneal endothelial cells cultured in 0.1% absolute ethanol without DEX. DEX did not affect the number of living cells at DEX concentrations from 10−10to 10−5M. However, 10−4and 10−3M DEX had an inhibitory effect
FIGURE 4 Apoptosis, necrosis, and viability assay. Results are expressed for (A) apoptotic, (B) necrotic, and (C) vital cells as the percentage of cells (mean ± SEM). Apoptosis and necrosis were significantly enhanced by adding 10−4and 10−3M DEX to the culture medium. Viable cells were significantly decreased by
adding 10−4and 10−3M DEX to the culture medium. Blot shows mean ± SEM (n = 6). ∗p< 0.01,∗∗p< 0.001 by Dunnett’s test.
FIGURE 5(A) Na+-K+-ATPase activity assay. Na+-K+-ATPase activity was significantly enhanced by adding 10−8M and 10−6M DEX to the culture medium. Blot shows mean ± SEM (n = 6).∗p< 0.01,by Student’s t-test. (B) RU38486 antagonist effect. RU38486 (mifepristone), an 11-substituted 19-norsteroid steroid antagonist, which has antiprogesterone effects (at low concentrations) and antiglucocorticoid effects (at high concentrations). Bovine corneal endothelial cells were cultured for 6 d. Five groups were compared: control (0.1% final absolute ethanol in medium), RU486 (10−5M RU38486 diluted in 0.1% final absolute ethanol), DEX
and RU486 (10−6M DEX+ 10−5M RU38486 diluted in 0.1% final absolute ethanol), DEX (10−6M DEX diluted in 0.1% final absolute ethanol), andMed (mediumwithout adding ethanol, DEX, or RU38486). The addition of 10−5
M RU38486 inhibited increased Na+-K+-ATPase enzyme activity with 10−6M DEX treatment. However, the addition of 10−5M RU38486 did not significantly inhibit the Na+-K+-ATPase enzyme activity compared to the effect of ethanol control treatment. Blot shows mean ± SEM (n = 6). (A) ∗p< 0.01 by Dunnett’s test.(B)∗p< 0.01 by Student’s t-test.
Ocular Immunology and Inflammation, 14:215–223, 2006